
Method Article
Анализ трубчатой мембраны сетей в кардиомиоциты от предсердий и желудочков
* Эти авторы внесли равный вклад
В этой статье
Резюме
В кардиомиоцитов, трубчатые мембранные структуры формирования внутриклеточных сетей. Опишем оптимизированы протоколы I) изоляции миоцитов от мыши сердце включая контроль качества, II) живой окрашивания клеток для государством в самых современных флуоресцентной микроскопии, и в) анализ прямой образ для количественной оценки сложности компонентов и пластичность внутриклеточного мембраны сети.
Аннотация
In cardiac myocytes a complex network of membrane tubules - the transverse-axial tubule system (TATS) - controls deep intracellular signaling functions. While the outer surface membrane and associated TATS membrane components appear to be continuous, there are substantial differences in lipid and protein content. In ventricular myocytes (VMs), certain TATS components are highly abundant contributing to rectilinear tubule networks and regular branching 3D architectures. It is thought that peripheral TATS components propagate action potentials from the cell surface to thousands of remote intracellular sarcoendoplasmic reticulum (SER) membrane contact domains, thereby activating intracellular Ca2+ release units (CRUs). In contrast to VMs, the organization and functional role of TATS membranes in atrial myocytes (AMs) is significantly different and much less understood. Taken together, quantitative structural characterization of TATS membrane networks in healthy and diseased myocytes is an essential prerequisite towards better understanding of functional plasticity and pathophysiological reorganization. Here, we present a strategic combination of protocols for direct quantitative analysis of TATS membrane networks in living VMs and AMs. For this, we accompany primary cell isolations of mouse VMs and/or AMs with critical quality control steps and direct membrane staining protocols for fluorescence imaging of TATS membranes. Using an optimized workflow for confocal or superresolution TATS image processing, binarized and skeletonized data are generated for quantitative analysis of the TATS network and its components. Unlike previously published indirect regional aggregate image analysis strategies, our protocols enable direct characterization of specific components and derive complex physiological properties of TATS membrane networks in living myocytes with high throughput and open access software tools. In summary, the combined protocol strategy can be readily applied for quantitative TATS network studies during physiological myocyte adaptation or disease changes, comparison of different cardiac or skeletal muscle cell types, phenotyping of transgenic models, and pharmacological or therapeutic interventions.
Введение
У здоровых поперечно-полосатых мышечных клеток, трубчатые мембранные структуры с «поперечных» ориентации (Т-трубочки) перпендикулярно главной оси клеток в изобилии. Следовательно, Т-трубочки были охарактеризованы как непрерывных расширений мышечной клетки главным "боковое" поверхность мембраны (сарколеммы), которые глубоко проникают в цитозоль к центру клетки. Физиологическая роль Т-канальцев непрерывных с внешней поверхностью мембраны является быстрое электрическое соединение удаленных внутриклеточные компартменты, образованных SER органелл контактных областей в течение относительно большого объема сердечной мышцы клеток на нанометровом связи близости от напряжения активируется L-типа Са 2 + каналы (Cav1.2) внутрь тока (I Ca) активации рядом рецепторов рианодиновый (RyR2) SER Ca 2 +-релизы. В желудочковой миоцитов (VM), не-непрерывные мембранные контакты ("переходах") между узловой ГЭЭ доменов и Т-tubules, как полагают, контролировать тысячи отдельных внутриклеточных нанодоменов Ca 2 + высвобождения в каждой ячейке 1.
Для любого данного контактной области, противолежащие друг другу мембранные части каждого из Т-канальцев и периферической (SER) соединительного примерно 15 нм, близкие друг к другу, следовательно, определенные как нанодоменного. Таким образом, очень маленькие индивидуальные цитоплазматические подпространства отделены которые позволяют квази клеточно-автономным поведения отсека. При входящем потенциал действия активирует Cav1.2 каналов в Т-канальцев виртуальных машин, сравнительно небольшой Са 2 + внутрь ток будет быстро увеличиваться подпространство Са 2 + концентрации [Ca2 +] S в attoliter размера нанодоменного 1. Далее, [Ca 2 +] S рост активирует Ca 2 + -gated рецепторы Рианодин (RyR2) в нанометровом близости в этом сопоставлении SER мембраны перехода, и этот процесс, связь возникает на протяжении всего электрического соединенияд миоцитов нанодомены. RyR2s произойти в плотных скоплений многоканальные, с общим стехиометрии 1 Cav1.2 канала в течение 5-10 RyR2 каналов 2. Поскольку ГЭЭ-к-цитозоле [Ca 2 +] градиент очень крутой (соотношение 10 4: 1) и RyR2s функционировать как высоко-проводимости, Ca каналов 2 + выпуску в функционально связанных кластеров, RyR2 активация приводит количественно большим Са 2 + выпустить ток от T-канальцев в сочетании соединительных ГЭЭ доменов увеличения местного подпространство [Ca 2 +] S до 100 мкм или выше в течение 1-2 мсек 3,4. Такое поведение усиление сигнала сердца также называют Са 2 + индуцированного Са 2+ выпуска (CICR). Взятые вместе, Т-канальцы являются важнейшими мембранные структуры, которые быстро активировать сигналы релиз Ca 2 + через соединительных нанодоменного ГЭЭ контактов и клеток-широкий CICR во возбуждения-сжатия (ЕК) связи.
В дополнение к Т-трубочек, осевая трубочкаS (A-трубочки) с существенно разной ориентации параллельно главных (продольных) оси клеток были зарегистрированы с помощью электронной микроскопии (ЭМ), конфокальной и 2-фотонных микроскопических исследований. Например, непрерывный решетки клеток в масштабе А-канальцев между миофибрилл, соединенных с Т-канальцев вблизи саркомера Z-линий было показано внеклеточных индикаторов и EM визуализации фиксированной морской свинки виртуальных машин 5. Использование внеклеточный декстран-связаны окрашивание флуоресцеина и жить 2-Фотон изображений крыс виртуальных машин, комплекс ретикулярная 3D трубочка сети визуализировали, состоящий из ~ 60% Т-трубочек и ~ 40% A-трубочки 6. Это исследование не только привело к 3D визуализации обильных А-трубочек, но и к пониманию, что секционирования для ЭМ визуализации в своей основе ограничена для анализа сложных и динамичных мембранных сети, такие как поперечно-осевой системы канальцев (Таты). Следовательно, конфокальной живая клетка томография Таты мембран непосредственно окрашивали ди-8-ANNEPS была разработана. Если живая клеткаТаты сети анализируются преобразования Фурье, регулярное появление Т-трубочка компонентов в пространстве вблизи саркомера Z-линий отражается ансамбля спектра мощности от области поперечно-полосатых сигналов 7. Эта стратегия косвенным анализ был использован для обнаружения клеток в масштабе региональных изменений в TATS компонента регулярностью в модели болезни 7. Например, ShRNA опосредованной junctophilin-2 нокдауна вызвало сердечная недостаточность и изоформы конкретных дефицит белка привело к Т-канальцев реорганизации с нанодоменного Са 2 + релиза дисфункции 8. Недавно мы расширили анализ Таты мембранных сетей посредством прямых количественных подходов и далее по живой клетки сверхразрешения микроскопии отдельных компонентов Таты в мыши виртуальных машин с использованием эффекта истощения излучения (STED) наноскопия 9. Нанометровом разрешении допускается прямой анализа небольших отдельных компонентов татов, приближенным распределение поперечных 50:50по сравнению с осевыми ориентаций канальцев, количественно подтвердив два обильные еще дифференциально ориентированные отдельные компоненты татов здоровым сердцем мыши 9. Эти стратегии будут более подробно изложены в разделе протокола ниже.
В то время как физиологическая роль обильным-канальцев компонентов во взрослом сердце осталась загадочной, исследования EM документально SER мембранные структуры, связанные с А-канальцев предлагая эндогенный Ca 2 + выпуску нанодоменов в морской свинки и крысы виртуальных машин 5,10. Конфокальной анализ Cav1.2 и RyR2 нашли высокую степень колокализации в-канальцев переходов 10. Поскольку ~ 20% спонтанных Ca 2 + искр в крысу ВМ возникших относительно далеко от Z-линии страт, где обычно происходят Т-трубочки, один аргумент был, что-трубочка, связанные нанодомены действительно может существовать и функционировать как Ca 2 + сайтов выпуска 11,12. Интересно, что образование Т-канальцев и созревание оспсы только после рождения и параллельно росту сердечных клеток, например, через прорастания сарколемных предшественником инвагинаций в Р5 и незрелые разветвленной TATS сетевых узлов на P10 у мышей 13. Похоже, что Junctophilin-2 является особенно важным для послеродового TATS сети созревания с ShRNA нокдауна предотвратить закрепление T-канальцев мембран ГЭЭ переходов приводит к замедленной Са 2 +-релизе и патологической организации Таты в соответствии с незрелыми-трубочка преобладают архитектур в ВМ 13. Эти наблюдения могут в конечном счете привести к доказательством правильности концепции, что Т-трубочки образуют через мембрану инвагинации процессов, тогда как А-канальцев может трансформироваться за счет дополнительных или даже альтернативных внутриклеточных механизмов 14.
Характеристика Таты мембранных изменений в сердечно-сосудистых заболеваний стала важным направлением исследований для патофизиологических вопросы. Первоначальные доклады в собачьей модели стимуляции, вызванной сердечной failuRe показал убыток Т-трубочек и Cav1.2 тока (I Ca) 15. Свинья модель ишемической кардиомиопатией, показали снижение плотности Т-канальцев и уменьшается синхронность внутриклеточного Са 2 +-релиз 16. Использование крыс со спонтанной гипертонией (SHR) модель сердечной недостаточности, потери Т-трубочек было связано со снижением нанодоменного сцепления Cav1.2 и RyR2 предлагаемым механизмом "RyR2 сиротами" 7. Потери Т-канальцев Также было показано, в человеческой виртуальных машин от ишемических, расширенными и гипертрофической кардиомиопатии образцов 17. Кроме того, увеличение в А-канальцев сообщили в срезах ткани человеческого дилатационной кардиомиопатией 18. После инфаркта миокарда, мы показали дифференциальный механизм реорганизации Таты в мыши виртуальных машин с значительным снижением Т-канальцев в отличие от повышений-канальцев компонентов 9. Важно отметить, что улучшилось местного мембрана контраст достигается за счет живой superre клетокРешение STED микроскопии позволило детальный анализ количественного компонента путем прямых измерений, которые показали значительное распространение А-трубочек с общим увеличением длины TATS сети и ветвления сложности 9. Кроме того, было показано, что учения могут обратить вспять Т-канальцев ремоделирования у крыс после инфаркта миокарда 19 и что СРТ может привести к обратной реконструкцию Т-трубочек у собак с мерцательной tachypacing индуцированной сердечной недостаточности 20. Взятые вместе, исследования как в больного человека и животного виртуальных машин, а также потенциальных терапевтических вмешательств будет возможно извлечь выгоду из процедур выделения клеток высококачественных и детальных стратегий количественного анализа, как указано в протоколе и приводит разделы ниже.
Кроме того, как показано недавно боковой поверхности по сравнению с TATS оборот мембрана KATP канала изоформы 21, важно рассмотреть предсердия мyocytes (AMS) в качестве биологически различны, а также сравнительной модели сердечной клетки против виртуальных машин. Т-трубочки были недавно зарегистрированы в овец и человеческого AMs 22. Текущие данные показывают, что существует несколько T-канальцев в АМ клеток и, как правило, в более крупных млекопитающих, таких как овцы и человека, но не в мелких грызунов 23. В отличие от виртуальных машин, в AMs появляется внутриклеточного Ca2 +, происходит высвобождение из клеточной поверхности, распространяющейся за счет диффузии в направлении центра клетки, что приводит к отмеченной пространственной и временной Са 2 + градиентов 23. В рамках этой структуры представляется важным выяснить механизмы внутриклеточного Са 2 + нестабильности для распространенных форм заболевания сигнализации, например, фибрилляции предсердий 24. В целом, как изолятор AM и В.М. и каждый для здоровых и больных сердец обычно используются протоколы. Только если изолятор сделано правильно судить по микроскопическим документации достаточного качества клеток, образцы AM и В.М. должно быть окrried вперед для количественного анализа татов. Соответственно, в следующих разделах протокола критически зависят от высокого качества клетки изолирует от мыши или других видов с последующим живых клеток микроскопии для анализа нетронутыми Таты мембраны. Как отметил ранее, характеристика Таты мембран является сложной областью исследований с склонностью к фиксации и подготовки артефактов 6, изменений мембранных вследствие осмотических изменений и ограничений разрешающей способности обычной световой микроскопии 9. Отметим, что последние государство-оф-искусство протоколы для изоляции человека AMs для Са 2 + изображений и патч-зажим и крыс виртуальных машин для культуры клеток были ранее опубликованы в этом журнале 25,26.
протокол
ПРИМЕЧАНИЕ: Все процедуры животных были рассмотрены и утверждены уходу и использованию комитета Институциональная животных Университетского медицинского центра Геттингена по в соответствии с гуманной помощи и использования лабораторных животных.
1 Выделение предсердий и желудочков миоцитов от сердца мыши
- Ручка животных как можно осторожнее и в соответствии с утвержденными протоколами, чтобы минимизировать стресс в целом и конкретно, чтобы избежать возможных мощных непреднамеренных эффектов от нейрогормональных эксцессов на изолированных клетках сердца. Кроме того, вводят каждый мышь с гепарином (500 МЕ / кг массы тела подкожно), по крайней мере 20 минут до извлечения сердца, чтобы предотвратить свертывание крови и микро эмболии, которые могут значительно компромиссу выход и целостность сердечных клеток в процессе выделения.
- Обезболить мышей в возрасте 12 недель и старше по изофлурана ингаляции, подтвердить отсутствие отмены боль рефлексов, и усыпить животных путем смещения шейных позвонков.
- Извлеките сердце стремительно следующий ранее утвержденных протоколов экспертных (например, видеть Kaestner соавт. 26 и Louch др. 27). Избегайте случайного повреждения предсердий ненужными сжатия или растяжения.
- Тщательно сохранить ткани проксимального восходящей аорты с помощью пары культи щипцов и прямых ножниц установление постоянного поперечная режущую кромку через аорты стенки сосуда, что очень важно для успешного катетеризации и перфузии сердца.
- Немедленно передать вырезали сердце в ледяную номинально Ca 2 + бесплатный буфер перфузии (для решения таблица 1). Держите большие сосуды зажаты во время передачи через воздух в буфер, чтобы избежать случайного воздушной эмболии, пока сердце не будет полностью погружен в воду. Используйте BDM в растворах буферных и охлаждаемых льдом ингибировать сокращение сердечной.
- Используйте Бинокулярный микроскоп с достаточным 3D Illumминации.
- Иглу аорту под панорамным стереоскопического сердца с гладкой, полированной поверхности 21 G канюли (наружный диаметр 0,81 мм; для нормального веса мыши сердца), который должен быть полностью заполнен буфер. Убедитесь, что нет никаких пузырьков воздуха в канюли, подключив ближний резервуар решение (например, шприц) через 2-полосная Luer клапана, позволяющего для быстрого контроля потока раствора.
- Подтвердите под бинокулярного увеличения, что канюля правильно расположенными внутри аорты, что примерно на 1 мм над аортального клапана и филиалов коронарных артерий. Абсолютно избежать прохода через или случайного перфорацию аортального клапана с канюли (это будет постоянно нарушать аорты закрытие клапана и, следовательно, нарушить сердечную перфузии).
- Свяжите аорты осторожно, чтобы заказных, ориентированные в кольцевом направлении канавок противоскользящих в конце канюли с помощью двух шелковых швов. Не смывать коронарного arteriэс силой в любой момент. Подключите решение заполнено канюли, привязанную к аорты и сердца, чтобы плотно пригнанной оттока коннектору настроены и предварительно калиброванного системы перфузии, он же модифицированного установки Лангендорфа (либо с помощью постоянного давления или постоянного потока; см также дискуссионный раздел ниже).
- Заливать сердце как можно скорее в течение 4 мин, используя кислородом буфер перфузии при 37 ° С (скорость перфузии цель: 4 мл / мин). Начать пищеварение путем переключения перфузии, чтобы содержащих коллагеназу пищеварения буфера (600 Ед / мл коллагеназы типа II) в течение 8-10 мин при температуре 37 ° С. Контроль за ходом ткани пищеварения, подтвердив аналогичные изменения тканей в том числе увеличением непрозрачности, мягкость, и вялость во всем видимой поверхности сердца.
- Проанализируйте сердечные камеры по мере необходимости, после расщепления. Поместите канюлированной сердце под бинокулярным микроскопом и визуализировать заднюю стенку сердца. Проанализируйте остаточной ткани например несердечная, легких Aй части судна, чтобы избежать загрязнения клеток в буфере пищеварения с использованием микро ножницы (например, пружинные ножницы с 8 мм лезвия), как показано на рисунке 1.
- Следуйте контрольный список, какие именно камеры, регионы и / или клетки коллагеназы переваривается сердце должно быть собраны (рисунок 1): левый и / или правое предсердие, свободный левый и / или стенки правого желудочка, и / или желудочковая перегородка.
- Для вскрытия конкретных сердечных тканей, использовать относительно широкий и плоский рассечение ванну, покрытую толстым слоем несколько мм силиконового пластика эластомера. Закрепите верхушке сердца с тонкой насекомых стальной шпильки до нижнего слоя эластомера.
- Отклонить правого предсердия придатком и анализировать в правое предсердие чуть выше атриовентрикулярных клапанов. Продолжить рассечение с левого предсердия. Проанализируйте и отбросить аппарат волокнистую клапана. Наконец, рассекают слева и справа бесплатно желудочковых стены и перегородки и / или небольшойэ ткани части по мере необходимости.
- Примечание только для стажеров: чтобы получить практику, начать с не переваренной сердцах мыши. Для простоты анатомической ориентации, практиковать обработку тканей, включая все последовательные шаги рассечение под бинокулярного зрения, как показано на рисунке 1. После того, как 3D анатомии, ручной обработки под бинокулярного зрения, и шаги рассечение в достаточной ознакомился, продолжить с коллагеназы переваривается мыши сердцах как описано выше.
- Для желудочка миоцитов (VM) выделения клеток: передать желудочка ткани в 2,5 мл свежего буфера пищеварения. Если одновременные сотовые выделения из нескольких частей органов, например, предсердий и желудочков попытка, второй человек может взять на одной ноге процедуры клеточной диссоциации, как свести к минимуму и оптимизировать использование мышей путем скоординированных обработки нескольких сердечных тканей. Продолжить с шагами 1.9.1-1.9.4, после 1,10.
- Проанализируйте либо весь желудочка тканиили определенные части их (например, ПЖ, свободные стены, и / или перегородки) в приблизительно 1 мм 3 штук в 2,5 мл пищеварения буфера, используя острые ножницы (например, пружинные ножницы с 8 мм прямыми лезвиями) в 60 мм чашки Петри .
- Аккуратно отделить виртуальных машин в клеточной суспензии медленным растирания тканевых штук с пипетки. Избегайте пузырьков воздуха в клеточной суспензии.
- Добавить 8 мл стоп-буфера к клеточной суспензии В.М. и передачи клеточной суспензии в 15 мл коническую пробирку. Разрешить оставшиеся кусочки ткани, чтобы поселиться в нижней примерно 15 сек, но достаточно коротким для изолированных клеток остаются во взвешенном состоянии. Далее, собирают суспензию VM через передачу объема надосадочной в новую пробирку на 15 мл. Если избыточные части ткани присутствуют, в качестве альтернативы использовать минимально 200 мкм нейлоновую сетку разнесенных отделить куски ткани из клеточной суспензии.
- Пусть суспензию клеток В.М. оседают на дно 15 мл COское трубки под действием силы тяжести в течение 8 мин.
- Промыть шаг: Удалить супернатант и осторожно ресуспендируют оставшееся VM осадок в 10 мл буфера перфузии. Повторите шаг мыть 1.9.5 (опционально: добавить дополнительные шаги мыть постепенно увеличивать Ca 2 + концентрации по мере необходимости).
- Ресуспендируют оседлое VM осадок в 10 мл перфузии буфера и распространять оставшийся объем клеточной суспензии в 1,5 мл микропробирок (примерно 50 000 В.М. клеток на пробирку).
- Для миоцитов (AM) изоляции предсердий клеток: передать переваренной / расчлененный предсердия ткани в 1 мл свежего буфера пищеварения.
- Разрежьте частично переваренной предсердия ткани в приблизительно 1 мм 3 штук в 1 мл пищеварения буфера, используя микро ножницы в небольшой чашке Петри (например, диаметром 60 мм). Аккуратно отделить AM клетки из переваренных кусков ткани в клеточной суспензии с помощью растирания с 1 мл пластиковую пипетку с вырезом наконечником, чтобы не повредить струи жидкости. Во времярастирание строго избежать пузырьков воздуха в клеточной суспензии. После механического перемешивания, добавить 4 мл стоп-буфера (50 мкМ CaCl 2; 10% BCS) арестовать оставшуюся активность коллагеназы в клеточной суспензии.
- Передача клеточной суспензии АМ в 15 мл коническую пробирку. Разрешить оставшиеся кусочки ткани, чтобы поселиться в нижней примерно 15 сек, но достаточно коротким для изолированных клеток остаются во взвешенном состоянии. Урожай объем надосадочной жидкости, содержащей свободные AM клеток с помощью передачи раствора в новую пробирку на 15 мл.
- Центрифуга клеточной суспензии AM, например, 2 мин при 20 мкг при комнатной температуре или - предпочтительнее для мембранных исследований - пусть клетки располагаются медленно под действием силы тяжести в течение 20 мин в 15 мл коническую трубку.
- Вымойте шаг: отбросить супернатант и осторожно ресуспендируют осадок АМ в 5 мл перфузии буфера. Повторите 1.10.4.
- Ресуспендируйте AM клетки аккуратно в 5 мл перфузии буфера. Распределите объем суспензии клеток в 1,5 мл микроцентрифужных ваннойES (примерно 1000 AM клеток на трубе).
- Анализировать и документировать изолированное качество клеточной популяции для каждого сердца, включая выход клеток с использованием трипанового синего окрашивания.
- Для этого, разбавленные 500 мкл клеточной суспензии в 1: 1 об / об с трипанового синего раствора (конечная концентрация 0,02%) с использованием 1 мл советы разрез пипетки. Смешайте клетки и трипановым синим нежно очень медленный вверх / вниз пипетирование. Немедленно применить трипановым синим, содержащий суспензию клеток в Neubauer-типа улучшенной цитометра и считать интактные миоцитов использованием инвертированного микроскопа.
- Исключите все клетки с ясным повреждением, мембранных пузырьков, разрушенных страт, контрактуры, и клеток, накапливающихся внутриклеточный трипановым синим (Рисунок 2). Также исключите спонтанно заражения клетки, которые чувствительны к последующей гибели клеток. Для оценки подсчет целых клеток в суспензии, использовать только кардиомиоцитов с регулярными страт, которые исключают трипанового синего всейобъем ячейки.
- Судите целостность отдельных предсердных и желудочковых миоцитов в проходящем свете микроскопии. Сохраните светлого поля изображения в качестве Tif файл документации и дальнейшего анализа.
- Используйте следующие критерии для анализа целостности клеток сердца:
- подтвердить наличие обычных бороздок по всему объему видимого клеток;
- подтверждения целостности непрерывной боковой поверхности мембраны на обеих клеточных сторонами, параллельными миофиламентов;
- визуализировать острые зубцы на интеркалирован- дисков с обеих сторон элемента, которые отражают целостность отдельных поверхность мембранных структур; и
- визуализировать флуоресцентные сигналы татов мембран (или immunolabeled кавеолин-3 белок или другие маркеры мембраны), как описано в разделе 3 для локализации корреляции рядом с основной клеточно-специфической яркий поля изображения морфологии (например, сочетать оба изображения с ImageJ как наложением соединения ИМАГе).
- Определите длину саркомера от изображений в светлом поле. Для средней длины саркомера, измерить расстояние последовательно выровнены саркомера страт, и разделить расстояние по количеству саркомерах. Мера по крайней мере, два места на клетку. Проведение анализа с коммерческим программным обеспечением или в автономном режиме с ImageJ.
ПРИМЕЧАНИЕ: Если обрабатывали разобщающих агентов, нетронутыми расслабленным виртуальные машины от мыши сердце показывать среднее саркомера длину ~ 1,9 мкм 28.
- Используйте следующие критерии для анализа целостности клеток сердца:
- Количественная морфологию и размеры отдельных предсердных и желудочковых миоцитов от проходящего света микроскопии изображения. Рассмотрим, что AM и В.М. клетки значительно отличаются по размеру. Измерьте длину ячейки, ширину и площадь, и вычислить длину: к ширине.
- Анализ размеров 2D клеток из передаваемого светового изображения в ImageJ помощью команды полигон инструмент выделения и добавить в менеджер ROI, аналогично Рисунок 3. Если морфологической сhanges, как ожидается, в конкретных контекстах исследования, дополнительно документом для всех клеток конкретного штамма мыши, возраст, пол, размер сердца, и любые вмешательства для последующей классификации данных.
2 Окрашивание Таты мембран в живых предсердий и желудочков миоцитов
- Загрузите изображения камеры (например, ВОУ-R2) с 42 мм покровного стекла. Для стабильного крепления миоцитов к покровным, готовят 20 мкл раствора ламинина 1:10 разбавлением ламинина складе в физиологическом буфере перфузии (конечная концентрация 0,2 мг / мл). Спрэд 20 мкл ламинином решения равномерно на покровного стекла.
- Подготовьте 800 мкл 50 мкМ раствора ди-8-ANEPPS в перфузии буфера. Для этого, разбавленные 20 мкл 2 мМ ди-8-ANEPPS маточного раствора в 780 мкл физиологического буфера.
- Чтобы окрасить виртуальных машин, пусть клетки урегулировать под действием силы тяжести в течение 8 мин в 1,5 мл реакционной трубы. Чтобы окрасить AMS, либо использовать гравитационный седиментации или зрВ клеточной суспензии в течение 2 мин (см 1.10.3). Для обоих AMs и виртуальных машин, тщательно удалите супернатант избегая ненужного перемешивание осадка клеток и аккуратно ресуспендируют осадок клеток в 800 мкл ди-8-ANEPPS содержащий решение (50 мкм). Немедленно передать ди-8-ANEPPS / миоцитов суспензии на ламинин покрытием покровное в камере формирования изображения.
- Пятно подвеска VM в течение 15 мин при комнатной температуре в темноте.
- Медленно удалить избыточный объем вверх через мениска жидкости по бокам изображения камеры с механической пипетки. Подтвердите, что большинство ди-8-ANEPPS окрашенные миоциты остается надежно прикрепленной к ламинином покрытием покровное и не стали видны в воздухе. Далее, мыть прилагаемый миоцитов суспензии один раз, медленно добавляя 1 мл буфера перфузии с последующим удалением избытка жидкости, включая не-адгезивных клеток.
- Осторожно накладывать окрашенных и поверхность прикрепленные миоцитов с 1 мл перфузии буфера медленно со стороны тон визуализации камеру. Поместите камеру изображений на столике микроскопа.
3 Визуализация TATS мембранных структур в Living предсердий и желудочков миоцитов
- В общем, тщательно выбирать наилучшее вариант (ы) флуоресцентного микроскопа, доступной для мембранного изображений татов. Для конфокальной микроскопии, рассмотреть недавнее поколение, современные флуоресценции микроскопы с оптимизированными PMT массива детекторов и переработки фотон путей, которые максимизируют интенсивность флуоресцентного сигнала. Для конфокальной микроскопии мелкие детали татов мембранные структуры использовать 63X 1,4 NA масло цель или - в зависимости от наличия - использовать сверхразрешение микроскоп STED для мельчайших деталей таты как отзывы для миоцитов конкретных заявлений Коль др 34 Для общих принципах высокой. Разрешение флуоресцентной микроскопии, обратитесь к разделу обсуждения.
- Установите параметры отображения, чтобы обнаружить наиболее, в идеале все ди-8-ANEPPS окрашивали внутриклеточный MEMбран внутри данного изображения миоцитов плоскости. Используйте следующие параметры, как отправной точки для конфокальной лазерной сканирующей микроскопии: возбуждения 458 нм например, в размере 3% от максимальной мощности лазера; обнаружить излучаемого сигнала между 550 нм и 740 нм; усиления детектора (например, мастер команда 800); и обскуры 1 а.е. для оптической толщины среза 900 нм. Настройте эти параметры для оптимизации соотношения сигнал-шум.
- Использование режима светлое поле, чтобы выбрать интактный АМ или VM ячейку, если необходимо (фиг.4А и 4В). Обратитесь к точке 1,12 для соответствующих критериев, как судить целостности клеток выделить ячейки, как кратко: сотовые всей регулярных страт и равноотстояния саркомера, острых поверхностных краев и зазубрин на всех четырех сторон элемента, непрерывной целостности боковой поверхности мембраны и отсутствие любые мембранные пузырьки.
- Возьмите пример изображения центрального внутриклеточного разделе миоцитов. Для регулировки ROI использовать "урожай" функции →; Настроить окно культур → конечный размер пиксела измеряет 100 нм х 100 нм. Отрегулируйте ось х окна урожая переписываться с основной (с осевой / продольной) оси миоцита.
- Выберите окончательный самолет изображений. Используйте одинарные кадров изображения, чтобы вручную выбрать соответствующий самолет изображений в Z-направлении. Убедитесь, что таты мембраны, в том числе Т-канальцев и компонентов А-трубочка, визуально проявляется в фокальной плоскости. Следует отметить, что типичный внутриклеточного плоскости отображения может включать в себя ядро в качестве внутриклеточного опорной точки. Обратитесь к примерам в фиг.4А и 4В.
ПРИМЕЧАНИЕ: В общем, сохранять концентрацию клеток к лазерным светом как можно короче. Если возможно, используйте отдельные кадры изображения, чтобы определить оптимальный фокальной плоскости в кардиомиоциты. - Отрегулируйте время пикселей ожидания на приблизительно 0,5 мкс. Выберите 16x усреднение и записывать изображение в виде снимков. Повторите шаг изображения снимков для создания соответствующей визуализации плоскость аы изложил для Таты мембранных структур в 3,5 мере необходимости.
- Сохранить окончательное изображение и убедитесь, что файл был сохранен в его целевой папке. В общем, сохранить все файлы изображений в том же формате (например, LSM) для единообразного применения программного обеспечения для анализа. До любого анализа изображений, еще раз подтверждают достаточную целостность клеток офф-лайн (рассмотреть критерии, перечисленные в шаге 1.12.1) и исключить любые поврежденные клетки от анализа. Обращайтесь к примерам в цифрах 4C и 4D.
4 Анализ Таты мембраны Сети и ее компонентов
Следующие шаги обработки изображений для прямого анализа мембранных компонентов таты суммированы сверху вниз диаграмме рабочего процесса в рисунках 5А и 5В.
- Откройте файл образ ди-8-ANEPPS окрашенных миоцита в Фиджи (http://fiji.sc/), в свободном доступе вариант ImageJ который содержит анализ плагины, необходимые дляобработки изображений. Для получения дополнительной информации, пожалуйста, обратитесь к Schindelin соавт 29.
- Сохраните изображение → Файл → Сохранить как → TIF.
- Для анализа мембранные компоненты татов, выберите соответствующий ROI за исключением внешней сигнала поверхности мембраны, а затем использовать "полигон выбор" инструмент, чтобы очертить границы ROI за исключением внешней поверхности мембраны (сарколеммы) и в том числе внутриклеточных участков Таты мембран, как показано на 5А (ROI). Добавить выбранный ROI в "Диспетчере ROI", применяя Проанализируйте → Средства → Диспетчер ROI.
- Для выбора конкретного ROI для анализа ориентации компонентов татов, выровнять основной продольную ось клеток и изображения ось х параллельно. Если ячейка является слегка изогнутой, выберите несколько трансформирования и выровнять каждую ROI индивидуально. Исключите все ядра из анализа. Исключите все чрезмерно изогнутые клетки из анализа, поскольку точное выравнивание RОИ будет становиться все труднее и увеличить ошибки ориентации во время анализа.
- Удалить любую нежелательную информацию сигнала от внешней поверхности мембраны: Правка → Очистить Вне генерировать "выбранный ROI" (Рисунок 5A). Убедитесь, что выбранный ROI содержит только внутриклеточные мембранные части, которые соотносятся с Таты сети.
- Выполните следующую цепочку шагов обработки изображений (команд) до проведения последующих количественного анализа, как описано в рисунке 5А.
- Нажмите на → процесса → вычитания фона. Установите радиус катящийся шар 5 пикселей.
ПРИМЕЧАНИЕ: Установите радиус катящийся шар 5 пикселей, если проанализировать изображение имеет размер пикселя 100 нм х 100 нм. Для других размеров пикселя устанавливается на катящийся шар радиуса к количеству пикселей, которые примерно соответствуют физическим радиусом 500 нм. - Нажмите на → процесса → Усилить локальный контраст (CLAHЕ). Установка размера блока до 49, гистограммы бункеров до 256, максимальный наклон к 3, и маску "Нет".
- Нажмите на → процесса → Гладкий.
- Нажмите на → Плагины → Сегментация → Статистические области Слияние. Установите параметры для Q100 → нажмите на Показать средних.
- Подтверждение полной обработкой статистической области слияния указывается автоматической презентации нового кадра изображения и подтвердить, что появляется метка "SRM Q = 100". Продолжить следующие меры с файл этого изображения. Нажмите на → Изображение → Тип → 8-битной.
- Нажмите на → Image → Adjust → Порог. Выберите порог достаточно низкую, чтобы обнаружить наиболее, в идеале все Таты структур, в частности, избегайте исключения компонентов татов с низкой интенсивностью сигнала (используют порог 40 как отправной точки). Обратитесь к "Представитель Результаты" раздел для детального вывода данных и примеры в рисунке 6(Верхний порог 255). Документ окончательный выбор пороговых параметров. Обратите внимание, что правильный выбор порога следует производить только определенные TATS мембранные структуры, но не ложные положительные сигналы в связи с фоновым шумом.
- Подтверждение правильности наложения татов деталей изображения по сравнению с извлеченными данными скелета, в частности для уровней сигнала средней и высокой флуоресценции, который должен соответствовать (путем размыкания) непрерывность структуры скелета. После того, как соответствующий порог был определен, применить этот же порог ко всем изображениям в процессе анализа и повторите сравнение наложения исходного сигнала по сравнению с каркасно данные последовательно минимизировать этот потенциальный источник смещения.
- Нажмите на → Применить: данные изображения становится двоичный, как показано на рисунке 5 в разделе "Threshold".
- Нажмите на → Плагины → Скелет → скелетируют (2D / 3D). Сохраните скелетонизированный 2D-изображения (как показано на рисунке 5), как Tif файл понажав на → Файл → Сохранить как → TIF. Анализ скелетонизированный файл изображения для количественного вывода данных, нажав на → Плагины → Анализ скелета (2D / 3D). Выберите усечения метод цикла: нет. Подтвердите автоматическую генерацию результирующей таблицы данных.
- Сохраните автоматически сгенерированный таблицу данных как текстовый файл. Выберите соответствующие количественные параметры, например, общее число точек ветвления или средняя длина ветви, как показано на примерах данных на рисунке 7. Рассмотрим дальнейший анализ данных с комплементарными / поддерживающих программных инструментов, таких как Excel и в соответствующих случаях.
- Нажмите на → процесса → вычитания фона. Установите радиус катящийся шар 5 пикселей.
- Рассмотрим использовать автоматизированные процедуры обработки изображений, когда это возможно, включая все необходимые шаги, описанные в 4,5 гармонизировать анализ между отдельными изображениями и / или изображения партий. Используйте пример дополнительном файле кода, содержащего обработки изображений Макро запрограммированный для Фиджи (отрегулировать программирования по мере необходимости).
- Анализ инdividual ориентации всех или некоторых TATS сетевых компонентов из данных скелетонированными изображения с помощью Фиджи плагин "Направленность". Генерирование гистограмму направленности, где соответствующие ориентированные в осевом направлении, поперечном канальцев или ориентированные T-канальцев компоненты представлены в 0 ° и 90 ° бункеров. Обратите внимание на правильное обращение ориентации изображения и, что важно для оси Х изображение, чтобы соответствовать в тесном сотрудничестве с основными (продольными) 0 ° оси клетки В.М., как показано на рисунке 8 и Представитель Результаты.
- Нажмите на → Анализ → Направленность → указать метод: компоненты Фурье, Nbins 180, Гистограмма начать -45 → нажмите на Показать таблице.
- Сохраните только генерировать и выводить результаты таблицу, включающую связанных данных гистограммы как текстовый файл. Рассмотрим дальнейший анализ данных TXT файлов на бесплатные программные инструменты, такие как Excel и по мере необходимости.
- Для создания подклассовДанные как сгруппированные наборы данных, например, для всех клеток, обработанных в тех же условиях (и потенциально других условиях), повторить анализ шаги 4.1-4.7 для всех соответствующих изображений по мере необходимости. Импорт Скелетонированный параметры данных из всех изображений в одном сочетании файл Excel с целью извлечения средние значения. Кроме того, импортировать все данные направленность гистограммы из того же набора данных, сгруппированные в объединенный файл Excel для расчета и генерировать средний гистограмму направленности. Рассмотрим дальнейшую обработку сгруппированных наборов данных для анализа дополнительных TATS сетевых параметров, представляющих интерес для отдельных или между различными группами лечения по мере необходимости.
Результаты
Кроме того, в TATS мембрана анализа сети ряд широко используемых методов клеточной биологии как внутриклеточного Са 2 + изображений, патч-зажим электрофизиологии, или фармакологических исследований доза-реакция в решающей степени зависеть высококачественной изоляции первичной ячейки от предсердий или желудочков или выберите части сердечной ткани для того, чтобы охарактеризовать зрелых дифференцированных, структурно и физиологических интактных кардиомиоцитов. Таким образом, изоляция и оценка качества для АМ и ВМ клеток, описанных в разделе 1, в конечном счете полезны для многих различных вопросов, включая сюда описанной анализа сети TATS, которая зависит главным образом от неповрежденной мембраны и целостности клеток.
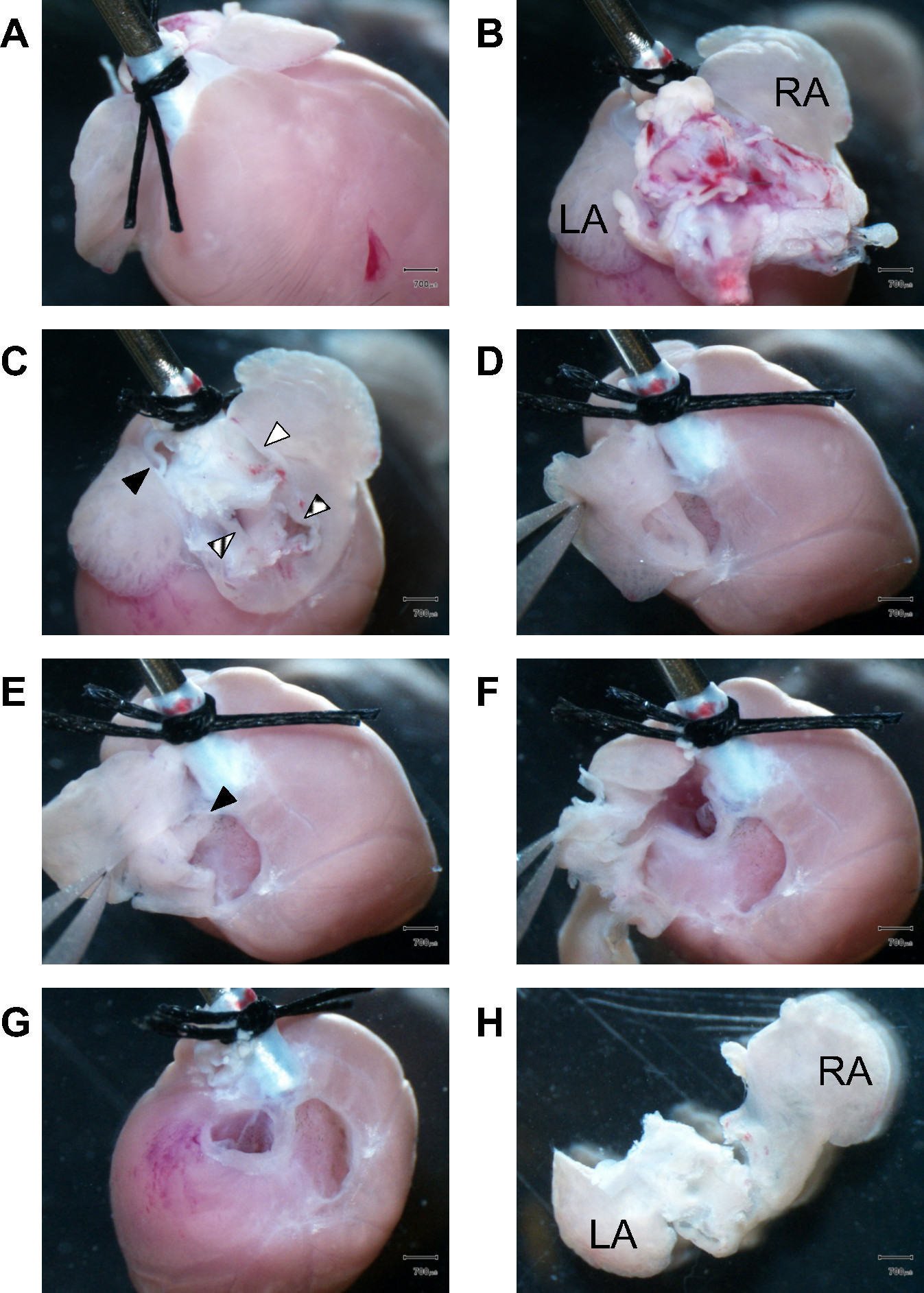
На рисунке 1 представлена поэтапного руководства изображений, как приступить к ткани рассечение сердечная начиная с мерцательной камер в мыши сердце. Впоследствии желудочковая камеры и перегородки готовятсяD расчлененный по мере необходимости. Точная отбор и подготовка правильных частей ткани важно достоверно установить AM и ВМ обособления с достаточной чистотой клеток. После коллагеназой это может быть довольно трудно определить правильную линию вскрытия между предсердий и желудочков ткани, еще раз AM и В.М. клетки смешивают неконтролируемое в клеточной суспензии, невозможно повернуть вспять смешанную популяцию клеток. Поэтому, анатомической ориентации, визуализация ткани 3D, достаточный опыт с расщепленной обработки тканей, правильная идентификация отдельных частей ткани и их рассечение линий будет все это способствует успеху выделения клеток.

Рисунок 1.Dissection мерцательной ткани. (A) Облицовка передней части сердца, два хирургических секutures исправить проксимального аорты до конца культи 21 G стали канюли. (B) вид на сердце базисных шоу оставшихся легочной ткани воспрепятствование предсердия вид камеры во время вскрытия. LA, левого предсердия; РА, правое предсердие. (C) Оставшееся lungtissue и магистральных сосудов были удалены, чтобы получить доступ к предсердий камеры. Заполненный черный треугольник, легочная артерия; поперечно-полосатой треугольник, легочные вены; заполнено белый треугольник, верхняя полая вена; В штучной упаковке треугольник, нижней полой вены. (D)-первых, стенки правого предсердия рассекают в то время как щипцы провести ушка предсердия. (Е) Вид в правом предсердии полости камеры. Черный треугольник обозначает интактный межпредсердной перегородки. (F), рассечение продолжают входить в полость левого предсердия камеры. (G) После полного рассечение левого и правого предсердия, желудочкового клапанов становятся видимыми. Аппарат волокнистый клапан рассекали и отбрасываются к после-uently урожай только желудочка мышечной ткани. (H) задней вид изолированных левого и правого предсердия. LA, левого предсердия; RA, правый atrium.Scale бары: 700 мкм.
Чтобы определить качество клеточных выделений, на фиг.2 представлены типичные примеры клеток при оценке урожайности и жизнеспособности типичной стержня или кирпича в форме поперечнополосатые интактные миоциты, как для АМС и виртуальных машин. Маленький повреждены, визуально незаметна, а также строго поврежденные клетки с аномальными чрезмерно изогнутыми морфологии, или аномальных клеток сферической формы, могут быть легко идентифицированы в том числе трипанового синего, как описано в разделе 1.11. В то время как неповрежденные миоциты остаются яркими и равномерно поперечно-полосатой при воздействии внеклеточной трипанового синего, поврежденные клетки, как правило, показывают несколько волдыри мембранные и / или быстро накапливаются трипанового синего внутриклеточно, указывающий повреждения мембраны. Тем не менее, трипанового синего само по себе может нанести вред клеткам через unnecessariлы Длинный инкубационный и непосредственная оценка качества клеток, следовательно, обязательным. Примеры наиболее очевидных форм повреждения клеток, как myofilament контрактуры или повреждения поверхности нарушения целостности валового клеток показаны на рис 4С и 4D. Пожалуйста, нажмите здесь, чтобы посмотреть увеличенную версию этой фигуры.
{kind=link}

Рисунок 2 трипанового синего окрашивания клеток исключение. Изолированный (А) AM и (Б) ВМ клетки смешивают в виде суспензии с трипановым синим и визуализировали с помощью перевернутой оптического микроскопа, показанной на 40Xmagnification. Примечание thatabnormal сферические клетки (А) и (В) с до трипановым синим, которые indicatesmembrane leakaGE и структурные повреждения. В противоположность этому, клетки centralAM и VM с интактными мембран исключить трипановым синим, как показано. Кроме того, обратите внимание, что нетронутыми AM и В.М. клетки показывают саркомера страты на протяжении всей их объема клеток, nomembrane волдыри, и острые края на обоих боковых сторон и оба интеркалированные диски. Шкала баров: 20 мкм.
После успешных урожайности изоляция, сотовые виртуальных машин от 5 х 10 5 до 10 6 можно ожидать от одной мыши сердца пищеварения. Выход АМН значительно ниже в порядке 3 х 10 3 до 3 × 10 4 в форме стержня, трипановым синим, за исключением клеток. В отличие от VM, AM изоляция иногда неудачно, даже в опытных руках. Шаг 1,11 суммирует процедуры, как оценить Выход выделенного здоровых клеток в суспензии. Кроме того, определить средние размеры клеток через шаг 1,13, как показано на рисунке 3 для индивидуального AM или клеточных штаммов VM или сравнить AM против населения В.М. клеток бок о бок (при необходимости). Пожалуйста, нажмите здесь, чтобы посмотреть увеличенную версию этой фигуры.
{kind=link}

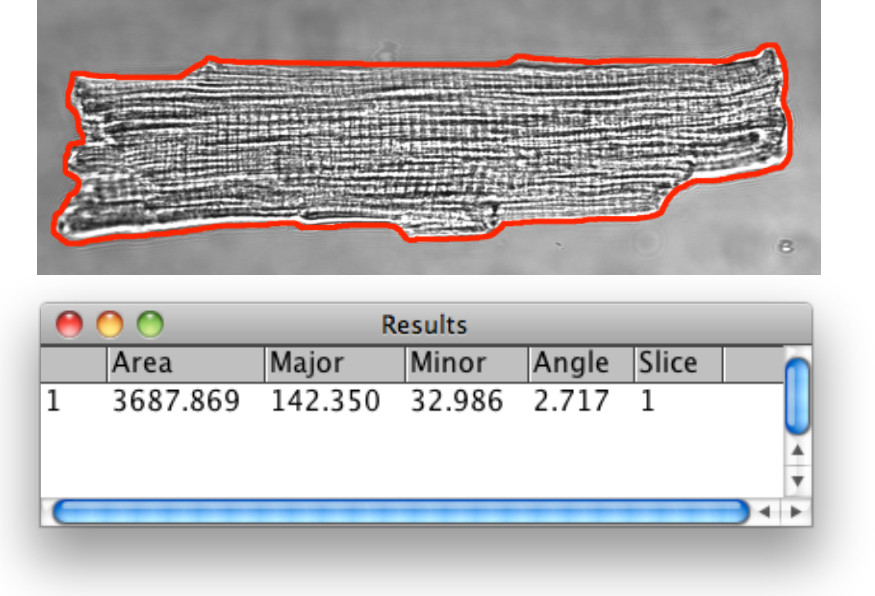
Рисунок 3.Bright поле морфометрический анализ кардиомиоцитов. Контур В.М. был обнаружен инструментами анализа изображений, описанных в шаге 1,13. С помощью инструмента выделения полигонов визуально след и определить границу внеклеточный Электролизер по наружной поверхности мембраны для анализа. Объявление выбранный область интереса (ROI) для менеджера ROI затем измерения расстояний 1D. Для сравнительных исследований AM против размеров В.М., это полезно для документирования длину ячейки, ширину и площадь, и вычислить длину: к ширине.
{kind=link}

Рисунок 4.Live мембрана окрашивание неповрежденной предсердий и желудочков миоциты. соответствующих проходящем свете и конфокальной образы живой ди-8-ANEPPS окрашивали нетронутыми (A) AM и (B) В.М. клетки. В отличие от этого, частично по контракту и, возможно, повреждены М. с длиной саркомера 1,2 мкм показано в (С). Контрактные миоциты обычно показывают аномально сокращен и искаженные Таты структуры, поэтому исключены из дальнейшего анализа. Еще один важный показатель для клеточных мембран дефектов мембранных волдыри (красные треугольники), как показано на виртуальной машине в (D). Мембранные везикулы представляют поврежденные поверхности мембранные структуры и клетки с пузырьков должны быть исключены из дальнейшего анализа татов. Кроме того, В. М. показывает валовую повреждения явно недостает целую часть его нижней левой части (отмечены звездочкой). Таким образом, при сравнении проходящего света и конфокальной изображения, клетки морфологии и поверхности неиспользованный документировано и в сочетании с флуоресцентным информационного сигнала. 'N' знаки нюCLEI исключена из анализа мембранного пятно татов. Желтые полоски показывают увеличивается трансформирования из того же конфокальной изображения, представленного выше. Шкала баров:. 10 мкм Пожалуйста, нажмите здесь, чтобы посмотреть увеличенную версию этой фигуры.
{kind=link}
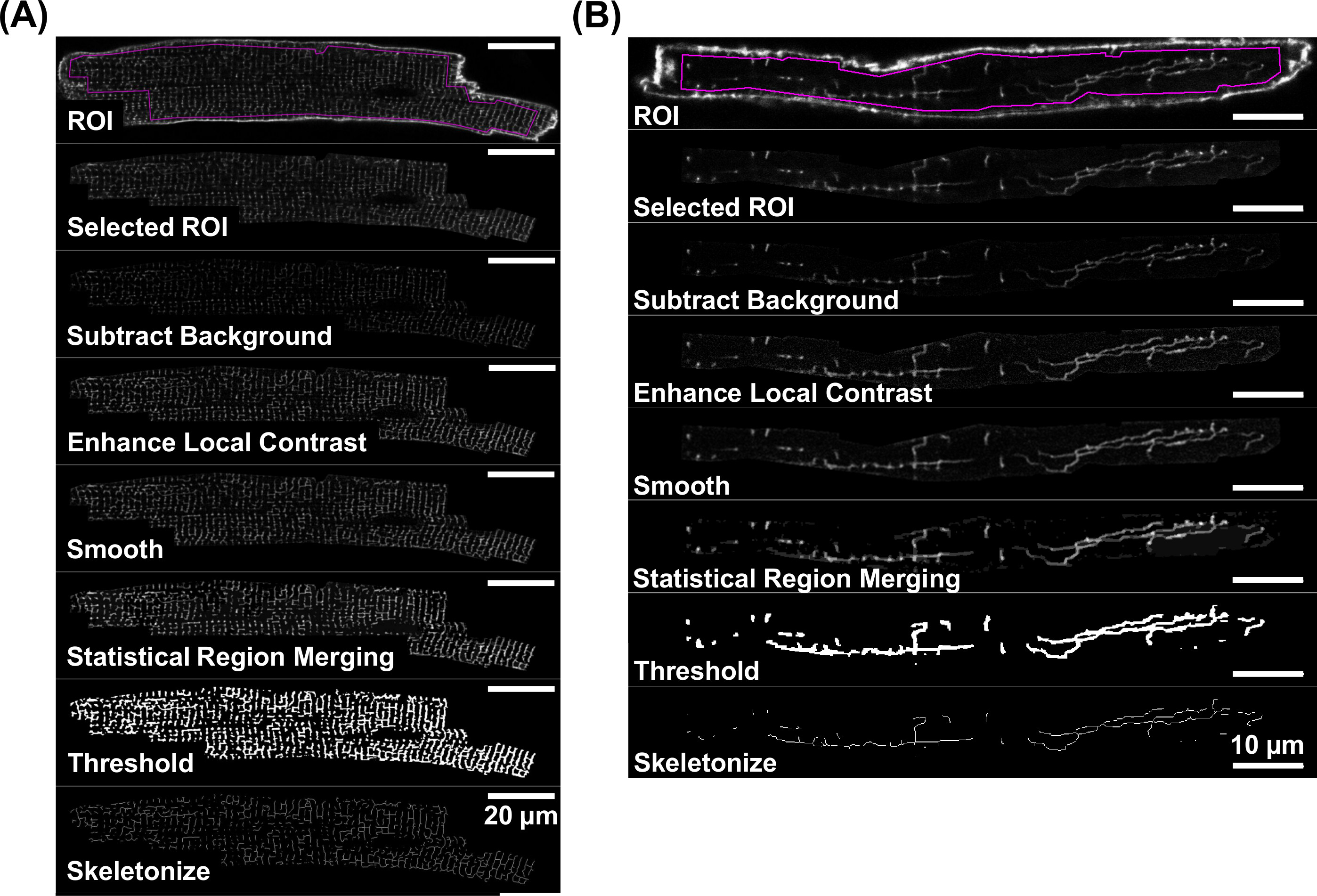
Конфокальные образы Таты мембран с достаточным отношением сигнал-шум, как показано на рисунках 4А и 4В принимаются для дальнейшего количественного анализа. Анализ мембраны TATS основан на скелетонированными данных, полученных из флуоресцентных прямолинейных компонент сигнала. Рисунок 5 показывает схему технологического процесса в отдельных этапов обработки изображений, которые подробно описаны шаги от 4,3 до 4,5. Эти шаги производить скелетонированными изображения, представляющие прямолинейных Таты мембранные сетей, как показано на каждой изолированной В.М. (рисунке 5А) и AM клетки (Fiфигура 5В).

Фигурки 5.Workflow для скелетизации флуоресцентных Таты изображений. Этапы обработки изображения, которые приводят к скелетной образа Таты сети представлены индивидуальными шаг за шагом примеры изображений как для ди-8-ANEPPS окрашивали VM (А) и AM (Б). Для отдельного шага обработки изображений, пожалуйста, обратитесь к разделу 4. Примечание различия в масштабные линейки:. 20мкм (А) и 10 мкм (B) Пожалуйста, нажмите здесь, чтобы посмотреть увеличенную версию этой фигуры.
{kind=link}
Критическим моментом обработки изображений, определить соответствующий порог бинаризации данных, как описано в разделе 4.5.6. Полученный бинарный образ должен включать только истинные мембранные сигналы от Таты сети, но не ложные структуры, получаемые ошибочной порога от шума фонового сигнала. Тем не менее, важно, что порог является достаточно низким, чтобы обнаружить все истинные Таты структуры, такие, что истинные компоненты Таты не ошибочно, потерянные во время анализа изображений. Рисунок 6 иллюстрирует процесс Как выбрать порог при бинаризации данных. В то время как порог 40, как показано на рис 6В и 6С представляется целесообразным выявить все истинные Таты структуры индивидуально, выбирая более высокий порог, например, из 60, как показано на рисунке 6A не обнаруживает слабее осевые структуры мембранных (ATS), как указано желтых треугольников . В отличие от этого, выбирают более низкий порог, например, из 20, как показано на рисунке 6D приводит к ошибочному обнаружению фонового шума, как ложноположительных структур TATS как указаножелтые треугольники.

Рисунок 6.How для определения порога сигнала во время скелетируя данных изображения татов. Примеры показывают Таты скелеты друг сгенерированные для разных порогов во бинаризации данных (описанных в шаге 4,5). Верхние изображения: показать пороговые настройки с использованием Фиджи. Нижние изображения: наложение скелетонированными изображений с соответствующей входной флуоресценции изображения и увеличенных областей, как указано. Высокий порог например, 60 применяется в (А), по-видимому, не подходит для обнаружения всех истинных TATS структуры, как показано желтых треугольников в увеличенном сечении. Порог 40, применяемой в (В) и (С) определяет все TATS структуры и правильно не обнаруживает фоновый шум, в то время как низкое пороговое напр., 20 в (D) ошибочно идентифицирует фоновый шум, как таты структур и тем самым производит ложноположительные сигналы несуществующих мембранных структур. Ложно-позитивные сигналы обозначены желтыми треугольниками в увеличенном вставке. Пожалуйста, нажмите здесь, чтобы посмотреть увеличенную версию этой фигуры.
{kind=link}
"Анализ скелета (2D / 3D)" Плагин поддерживает детальный анализ скелетонированными Таты структур. После выполнения плагин создает таблицу данных со следующими параметрами скелета: #branches, #junctions, # конечных точек вокселей, средней длины филиала, #triple пунктов, #quadruple пунктов, и максимальная длина ветви. Для подробного описания всех возможных выходных параметров приведен на сайтеhttp: //fiji.sc/wiki/index.php/AnalyzeSkeleton и соответствующих статей 29-31. Типичная выходная таблица данных показана на рисунке 7А.
Параметры могут быть дополнительно использованы для получения общей длины скелета на площадь или количество переходов на площади. Пример расчета количества филиалов, умноженных на среднюю длину филиала дает общую длину непрерывного скелета в 2D:Общая длина скелета за ROI:
Σ (#branches х средняя длина филиал) = 5155 PX = 515,5 мкм
Общая протяженность скелета может быть нормированы на области изображения. Для примера, показанного на фиг.7, нормированная длина скелет 0.64μm / мкм 2, а сумма всех переходов вычисляются, как показано ниже:
Нормированная длина скелета:
515,5 мкм / 803,6 мкм 2 = 0,64 мкм / мкм 2
Нормализованная количество перекрестков:
155 переходов / 803,6 мкм 2 = 0,19 переходы / мкм 2

Рисунок 7.Automated данные, выводимые из скелетонированными изображений. (А) Типичный таблицы данных, порожденная "Анализ скелета (2D / 3D)" плагин от скелетной изображения, показанного на (B). Для подробное описание возможных выходных параметров обратитесь к http://fiji.sc/wiki/index.php/AnalyzeSkeleton . Пожалуйста, нажмите здесь, чтобы посмотреть увеличенную версию этой фигуры.
{kind=link}
Этапы обработки изображений, описанные в 4,5 и показанные на рисунке 5 можно автоматизировать с помощью макроса Фиджи представили как Дополнительный код файла . Команды определить повторы обработки изображений через повторами шагов. Макро могут быть применены для завершения стеки входных изображений, полученных с помощью шагов 4,3 и 4,4. Макрос может быть выгодным для анализа полных наборов данных групп, например путем подготовки отдельных стеков входного изображения для каждой независимой группы лечения для автоматизированного анализа с использованием макрокоманд.
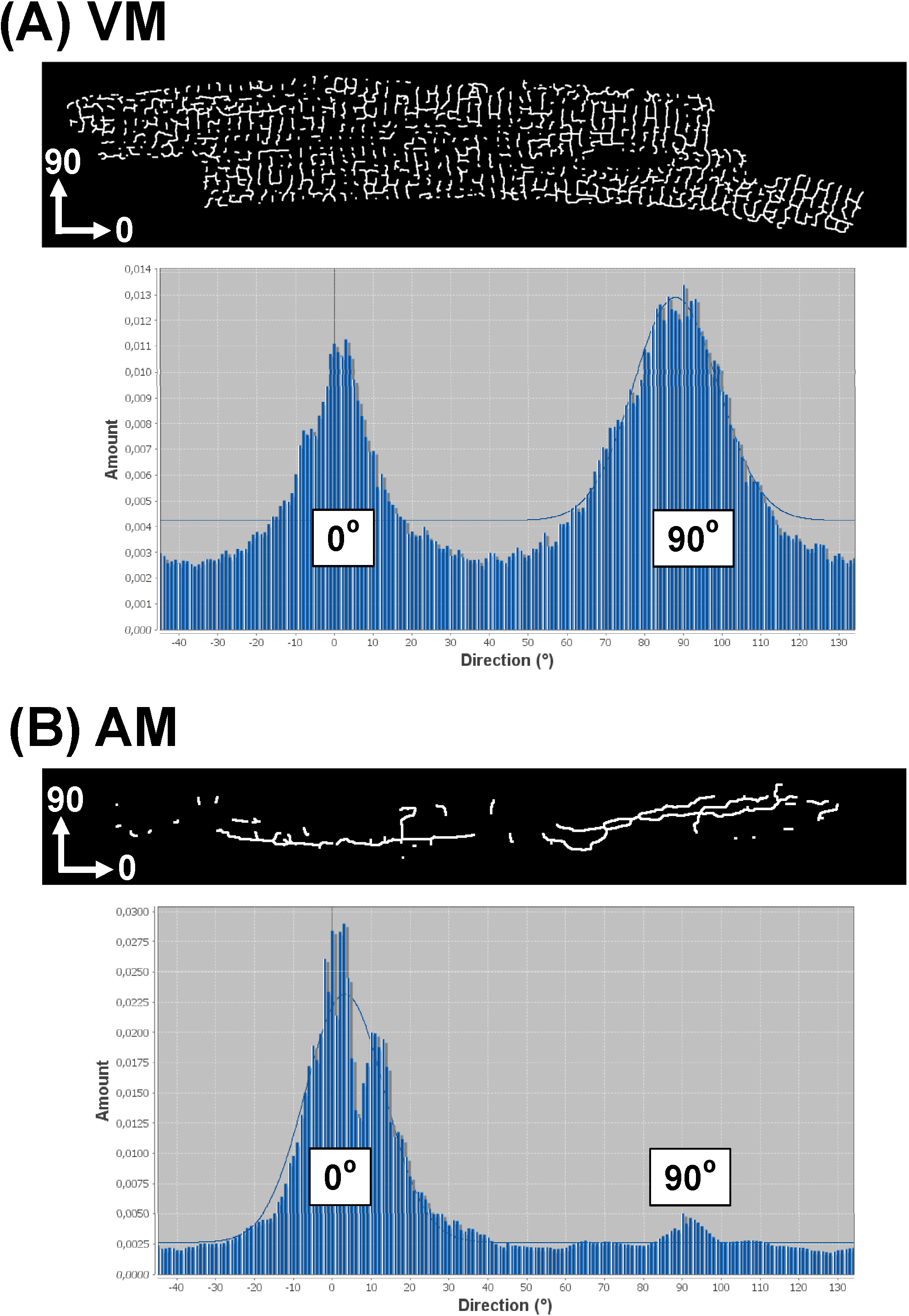
Изложил стратегию программного обеспечения дополнительно позволяет анализ TATS сети ориентации для всех компонентов. Для этого используйте "направленности" плагин (http://fiji.sc/Directionality) 29,31, которая производит данные гистограммы с ориентации всех татов компонентов ориентаций. Если X-ось входного изображения соответствует главной оси данного AM или VM клетки, осевая (продольная) TATS компоненты будут представлены в 0 ° бункера, в то время как поперечные компоненты будут представлены 90 и# 176; бен. Рисунок 8 показывает примерный направленность гистограмм от скелетонированными Таты изображений для виртуальной машины (8а) по сравнению с АМ (8b) клетки. В то время как направленность гистограмма типичном VM показывает двойное распределение пик при 0 ° и 90 °, гистограмма М. показан один доминирующий пик при 0 °. Эти примеры находятся в согласии с более ранними наблюдениями, что отдельные компоненты таты из виртуальных машин будут почти одинаково распределенных между Т-трубочек и-трубочек, в то время как компоненты Таты в AMs может быть преимущественно состоит из-трубочек.

Рисунок 8.Representative гистограмму направленности от Таты сетей отдельных клеток. Направленности гистограммы были получены из скелетонированными изображений индивидуального VM (A) по сравнению с AM (B ) Таты сети. Учитывая, что ось х анализируемого изображения соответствует главной (продольной) оси миоцита, как показано, компоненты А-канальцев представлены 0 ° бункера, в то время как Т-канальцев компоненты представлены в бункер 90 °. Гаусс подходит только к крупной гистограммы peakasshown в качестве фона графика. Пожалуйста, нажмите здесь, чтобы посмотреть увеличенную версию этой фигуры.
{kind=link}
| Перфузии буфер | мМ |
| NaCl | 120,4 |
| KCl | 14.7 |
| KH 2 PO 4 | 0.6 |
| Na 2 HPO 4 | 0.6 |
| MgSO 4 | 1.2 |
| HEPES | 10 |
| NaHCO 3 | 4.6 |
| Таурин | 30 |
| 2,3-бутандион-моноксима | 10 |
| Глюкоза | 5.5 |
| рН 7,4 | |
| Пищеварение буфера | мМ (если не указано) |
| NaCl | 120,4 |
| KCl | 14.7 |
| KH 2 PO 4 | 0.6 |
| Na 2 HPO 4 | 0.6 |
| MgSO 4 | 1.2 |
| HEPES | 10 |
| NaHCO 3 | 4.6 |
| Таурин | 30 |
| 2,3-бутандион-моноксима | 10 |
| Глюкоза | 5.5 |
| Коллагеназы типа II | 600 ед / мл |
| рН 7,4 | |
| Упорный буфер | мМ (если не указано) |
| NaCl | 120,4 |
| KCl | 14.7 |
| KH 2 PO 4 | 0.6 |
| Na 2 HPO 4 | 0.6 |
| MgSO 4 | 1.2 |
| HEPES | 10 |
| NaHCO 3 | 4.6 |
| Таурин | 30 |
| 2,3-бутандион-моноксима | 10 |
| Глюкоза | 5.5 |
| CaCl 2 | 0,0125 |
| телячьей сыворотки | 10% |
| рН 7,4 | |
Таблица 1.Buffer Solutионы. Содержание трех различных физиологических буферных растворов для выделения клеток и визуализации суммированы.
Обсуждение
Хотя кардиомиоцитов были выделены и изучены в течение десятилетий 32, недавний обзор к выводу, что последовательные высококачественные изоляция миоцитов клеток останется сложной 27. Это отражает относительно сложные протоколы для изоляции первичных кардиомиоцитов по отношению отношению отсутствия общих стандартных подходов, общей метаданных, и прозрачной документации качества клеток. Протоколы изоляторе, как правило, настроены на отдельные группы, произвести переменные результаты из клеток изолирует, зависит от индивидуальных настроек модели (например, вид, возраст, сосуществующие болезней сердца), и, как правило, с поправкой на конкретных экспериментальных условиях. В контексте количественного Таты мембранные исследования и протоколы, представленной здесь, существенном уровне оценки качества и документации касается конфокальной или сверхразрешения микроскопии отдельных клеточных мембран структур, склонных к метаболическим и протокольных изоляция зависимых изменений, какв AM или VM. Важно отметить, что даже если высокие урожаи клеток изолятов предложить здоровые неповрежденные миоциты, следователи должны задокументировать и критически судить каждый отдельный элемент тщательно против морфологических критериев поверхности и Таты целостности мембраны против неспецифической повреждения в результате процедур выделения против конкретных изменений вследствие различных типов вмешательств по сравнению с контрольными условий. Важной переменной в процессе выделения клеток сердечной является удельная активность коллагеназы данной партии. Для выбора нового много коллагеназы, ферментативная активность нескольких коллагеназы образцов должны оцениваться в сравнении друг с другом путем оценки сердечной выход миоцитов и качество, и в соответствии с инструкциями производителя. В идеале, новая много коллагеназы отождествляется с активности коллагеназы, аналогичном ранее успешно используются большим (для расширенной оценки возможных мероприятий ферментных относятся к "коллагеназа партии выбора инструмента" в материалах и методах таблице). ТАкен вместе, количественные подходы Таты мембраны визуализации решающей степени зависеть от качества изоляции клеток и, наоборот, выделений клеток сердца, ведущих к неспецифической повреждения мембраны, как документально Таты микроскопии должен вызывать критический обзор и коррекцию изоляции процедур. Поскольку качество изолятор и TATS мембрана визуализация и количественное неразрывно связаны, протоколы, обсуждаемые в этой статье, охватывают все основные аспекты, как непрерывной стратегии.
Дополнительной проблемой и общей проблемой кардиологических исследований, повреждения клеток и / или гибели клеток происходит за счет метаболически компрометирующих вмешательств например, после инфаркта миокарда 9, еще нужно судить против потенциального ущерба, например, случайного, после незаметно эмболии воздуха во время выделения клеток. Выделение кардиомиоцитов из больных сердец может привести к дополнительной, существенной потери клеток и снижение урожайности клеток. Поэтому, сравнимыисоном от общего числа изолированных интактных клеток между контролем и больных сердец может быть значимым, если изолятор и подсчета последовательно применяются через стандартизованных протоколов. Следовательно, очень важно, чтобы судить целостности клеток с помощью соответствующего контрольной группе, что отражает лучшее качество изолятор можно миоцитов. Важно отметить, что индивидуальный качество клеток и живых клеток микроскопия здоровым по сравнению с больными против миоцитов случайно повреждены процедуры выделения могут существенно повлиять на анализ Таты мембранных сетей. Протоколы представленные здесь связи с этим подчеркиваем целостности и стабильности физиологических компонентов мембранных процессе выделения клеток и живых клеток микроскопии интактных мембран. Весь технологический процесс выполнен в виде непрерывной стратегии для достижения и сохранения интактные компоненты TATS мембраны, исключая поврежденные клетки, поскольку они будут демонстрировать изоляции зависимых артефактов, таких как мембранные разрушенных мембран канальцев, MEMBRANе пузырьки, и измененные Таты сети ошибочно условиях контроля и поставить под угрозу дальнейшее количественного анализа. И наоборот, те же стратегии имеют решающее значение для интервенционных исследований с потенциалом, чтобы сорвать Таты мембраны, которые зависят главным образом от значимых управления сравнения между истинным здоровым по сравнению с истинной больной клетки с Таты мембранных изменений.
Кроме того, мы обращаемся процедур для достижения технически гораздо более сложной задачей изоляцию АМ клеток. Несмотря на прогресс и улучшенных протоколов, важно подчеркнуть, что это не так просто воспроизвести обособления высококачественные сотовые виртуальных машин и еще менее надежной для AMS. Это связано с общим нижним выходом АМ клеток, где даже небольшие ошибки или вариации в процессе выделения клеток может привести к полному отказу изоляции AM клеток, в то время как легкой степени повреждения клеток VM может быть менее очевидны в клеточной суспензии в связи с относительно высокой клетки цифры по сравнению с AM. С AM клетки могут стать сurved после выделения, анализ через несколько трансформирования может быть выгодным, как указано в шаге 4.3. После подробного процедуры стадий выделения клеток, мы предоставляем протокол для прямого интегрального мембранного окрашивания и конфокальной или STED сверхразрешения визуализации Таты сетей как для виртуальных машин и AMS. Эти протоколы позволяют как количественный анализ и дифференцировку некоторых компонентов Таты мембран через ранее установленными параметрами. По сравнению с виртуальными машинами, 3D организация и функциональные поведение предсердий сети Таты в AMs в настоящее время менее понятно.
Процедуры в изображение Таты мембран в живых клетках (шаги 3,1 до 3,7) были разработаны с коммерческой конфокальной (таблица материаловедения / Оборудование) и заказных STED флуоресценции микроскопы 9. Для оптимизации настройки микроскопа для флуоресцентного формирования изображений и количественного анализа татов, следующие пункты имеют общее значение:
- Цель
Для решения мелкие детали Таты мембранных структур, эмпирически проверить, цель которого обеспечивает самое высокое качество изображения при фокусировке несколько микрометр глубоко в клетки. Некоторые конфокальной микроскопы могут работать лучше с целями воды или глицерина, в отличие от 63X 1,4 NA объектива масла, используемого здесь. Цели с 100-кратном увеличении используются для сверхразрешения STED микроскопии, компромисса меньшее поле зрения для нанометрического резолюции 34. - Возбуждение и усиления
Оптимальные настройки мощности возбуждения и усиления детектора зависит от пути в микроскоп света, характеристики лазера и примеров свойств. В идеале, мощность лазера и усиления корректируются использовать весь спектр детектора, пока избежать насыщения изображения. Коммерческие пакеты микроскопия программного обеспечения обычно предоставляют справочные таблицы, что визуализировать нижнюю и верхнюю границу динамического диапазона. Кроме того, чтобы свести к минимуму отбеливание красителя используют минимально возможную лAser сила, которая все еще обеспечивает достаточные структурные детали TATS мембранные. Кроме того, мощность возбуждения должна быть достаточно низкой, чтобы избежать повреждения накопительный просмотром приводит к миоцитов контрактур и смерти. - Размер пикселя
Используйте размер пикселя совместимый с отбором проб Найквиста, примерно половина разрешение достигнутый с заданными настройками. Для конфокальной микроскопии размер пиксела 100 нм х 100 нм совместим, которая также ограничит отбеливание. Для сверхразрешения микроскопии значительно меньших размеров пикселя используются, например,, 20 нм х 20 нм для STED микроскопии 9. - Время задержки
Конфокальные микроскопы обеспечивают функцию усреднения. В общем, использовать в кратчайшие сроки пиксель задержки, чтобы избежать отбеливания в сочетании с сигнала усреднения, например, линия-в среднем ≥ 8 для улучшения шума-отношение сигнал-. - Документ настроек, установленных микроскопии Through Мета-данных
После настройки, какдетали изображения татов мембранных структур были оптимизированы на определенном конфокальной микроскопии, сейф и / или документирования параметров (протокол мета-данных). Приобретать все изображения в пределах (или между) группы (групп) клеток с той же целью, мощности возбуждения, усиления размера пикселя, пиксель времени выдержки, и в среднем функции. Равные условия формирования изображения позволяет для прямого сравнения и количественного в пределах (или между ними) группой (ами) клеток. - Для общего руководства и более подробной информации о принципах и применения конфокальной микроскопии см Справочника биологическом конфокальной микроскопии (Pawley JB, 3-е издание, 2006 г., Springer Science + Business Media, LLC).
В отличие от прямых стратегий анализа представленных здесь, предыдущие публикаций, описывающих Таты мембран и изменения заболеваний, связанных, использовали региональные совокупные показания плотности Т-канальцев как количественное стратегии 16,17, или косвенных региональных стратегий на основе Transf Фурьеormation анализ поперечно-полосатых мембранных сигналов для оценки Т-канальцев компонентов закономерность 7. В отличие от этого, количественные подходы, описанные здесь, непосредственно связаны с отдельными компонентами таты и предоставить ряд дополнительных параметров, включая свойствах сетевого мембранных и конкретных компонентов, таких как процент A-трубочек. Кроме того, плотность TATS сеть может быть определена количественно, как нормированной длины всей добываемой скелета в области ROI. Число тройных стыках трех индивидуальных, непрерывно соединенных компонентов канальцев может быть использован в качестве меры разветвления сложности мембраны сети TATS. Отметим, что любой анализ мельчайших компонентов татов зависит от процедур окрашивания. По нашему опыту, 800 мкл 50 мкМ раствора ди-8-ANEPPS достаточно, чтобы окрасить полные Таты сетей в осадок клеток, содержащей 50 000 ВМ клетки 9. Однако, если клеточный осадок содержит меньшее количество кардиомиоцитов, Если мощные детекторы флуоресценции доступны, и если конфокальной микроскопии от общего распределения TATS сети, а не мелкие детали мембранные и количественные изменения представляют интерес, концентрации низкие красителя может быть использован на основе эмпирической проверки. Наконец, программное обеспечение макроса, написанного для описанного анализа может быть использован для автоматизации этапы обработки изображений для облегчения анализа больших массивов данных, что особенно полезно для сравнения различных группах (например, наркотики), типов клеток (например, AM против VM ), и патофизиологические вмешательства (например, обман против инфаркта миокарда).
Для анализа изображений Таты сетей, следующая последовательность основных шагов применяется: 1) катящийся шар фон-вычитание (4.5.1), чтобы удалить пространственные изменения в интенсивности фона; 2) повышение локальный контраст (4.5.2).; 3) сглаживание изображения (4.5.3); 4) статистическая область слияние (4.5.4); 5) определениеПорог изображения бинаризации (4.5.6); и 6) расчет скелета данных (4.5.8). Критическим моментом скелетизации флуоресцентных Таты изображений бинаризация изображение показано на рисунке 6. Соответствующие шаги порогов в конечном счете, определить, какие истинные мембранные структуры обнаруживаются представлять основные компоненты Таты против потенциально ложных структур, выявленных ошибок от фонового шума. Идентификация правильной порога для анализа бинарного изображения должны соответствовать истинных Таты мембранных структур, которая зависит от достаточно высокой (SNR) отношение сигнал-шум каждого для конфокальной и сверхразрешения подходов микроскопии. Поэтому, достаточно качество изображения должны быть установлены первый и впоследствии в сочетании с критического суждения индивидуального качества клеток, включая документацию по изображениям в светлом поле, как указано. Альтернативные варианты для адаптации протокол сегментации изображений для данного выхода и / или физ данных микроскопаiological вопросы включают деконволюцию изображения и другие процедуры пороговых значений, как "Оцу" или "Изо-данных", входящей в ImageJ плагинов. Независимо от окончательного процедуры сегментации, рассмотрим сравнение добытых и исходных данных по наложенных изображений обязательный этап контроля качества. Таким образом, морфологические и мембраны целостность отдельных изолированных миоцитов, достаточной окрашивания внутриклеточных Таты мембран, оптимизации параметров для визуализации флуоресценции, и наложения контроля извлеченных данных скелета будут способствовать повышению качества люминесцентных Таты изображений и количественных результатов.
Если более крупные виды, чем мыши, используются для выделения клеток, протоколы могут быть легко адаптированы по мере необходимости. В течение следующих крупных видов, сердцах крыс может быть канюлю с тупым канюли 14 г (внешний диаметр 2,1 мм) и перфузии при 8 мл / мин. Значительно более старые или больные сердца может потребовать даже больших размеров канюли. В генеRAL, сердечная перфузии может проводиться либо например, постоянное давление, с использованием колонки высокого воды 1 м между резервуаром и аорты или постоянным током с использованием перистальтического насоса. Для выделения клеток из небольших грызунов сердца, как мыши и крысы постоянный поток может быть выгодно, так как коллагеназа пищеварения, в конечном счете сорвать коронарные сосуды сопротивления, ведущие к чрезмерной скоростью перфузии от утечки сосудов кроватями, которые будут контролируемых до некоторой степени протоколов постоянных потоков. В отличие от этого, постоянное давление перфузии является предпочтительным, если это мониторинг скорости потока и правильной катетеризации приоритетом, что является преимуществом для моделей вмешательства с измененной крови поведения сопротивления судна, а также для подготовки процедур выделения клеток.
Как отмечалось выше, достаточно качество клеток очень важно для количественных исследований эндогенных мембранных систем. Тем не менее, во время сердечного перфузии и коллагеназой многочисленные факторы могут крitically влияет на качество изоляции клеток, которые никогда не должны недооценивать во оптимизация протоколов или устранения неисправностей 27. В частности, активность данного коллагеназы лота должна быть определена для конкретной ткани процентных например, предсердий или желудочков до выполнения экспериментальных добросовестных исследований, чтобы создать условия изоляции, которые будут поддерживаться на протяжении оставшейся части исследования. Кроме того, качество воды, рН, температура, оптимизация и чистка установки перфузии позволит свести к минимуму риск случайного повреждения от загрязнений и эмболии, и потенциально дополнительные факторы должны контролироваться, чтобы установить оптимальные гомеостатические условия во время выделения клеток. BDM (2,3-бутандион-моноксима) обратимый ингибитор миозина АТФазы поперечных мостиков обычно используется во время рассечения тканей и пищеварения для поддержания сердечной мышечной релаксации, которая увеличивает выход клеточных выделений. Тем не менее, следователи должны то знайте, что BDM могут оказывать неспецифические фосфатазы деятельность, направленную на офф-целевых эффектов, например, ингибирования Na + / Ca 2 + обменных токов при определенных условиях 33. Для некоторых экспериментах было бы выгодно, чтобы заменить BDM путем blebbistatin как кардиоплегической раствора ингибитора, с высоким сродством к миозина в микромолярных концентрациях, которые, однако, токсичными и относительно дорогой и может иметь другие мимо ворот эффекты. Покоя здоровые кардиомиоциты не должны показывать никаких сокращений в отсутствие электрической стимуляции и таких клеток должны быть исключены из дальнейшего анализа. С другой стороны, сердечной миоцитов сокращения и расслабления в ответ на электрическую стимуляцию при физиологических внеклеточных концентраций Са 2 + может быть использован для установления нормального поведения сократительную в качестве дополнительной меры для оценки функционального качества клеток и / или неправильное поведение в болезни сердца по сравнению здорового контроля клетки.
Таким образом, протоколы для изоляции одного клеток и количественного анализа изображений, описанной здесь были успешно применены для конфокальной и сверхразрешения микроскопии мембранного сети Таты в VM 9 и АМ клетки 21, а также для количественного анализа микротрубочек сетей в неподвижные кардиомиоцитов (данные не представлены). Эти и будущие заявления протоколов может открыть возможности для различных экспериментальных вопросов, таких как характеристике Таты мембран на разных стадиях развития или анализа мембранных связано белка или органелл структур, которые контактируют Таты сеть для оказания сильно локализованы, область определенный сигнализации функции в АМ и ВМ клеток.
Раскрытие информации
The authors declare that no conflict of interest exists.
Благодарности
This work received support through Deutsche Forschungsgemeinschaft SFB 1002 (subprojects A05 and B05 to S.E.L.) and KFO 155 (subproject 4 to S.E.L.), a Halbach Foundation award to S.E.L. supporting E.W.; a grant from the German Cardiac Society to S.B.; and a DAAD exchange program supporting T.K. as visitor at the University of Maryland. The research leading to these results has received funding from the European Community’s Seventh Framework Program FP7/2007-2013 under grant agreement No. HEALTH-F2-2009-241526, EUTrigTreat (to S.E.L.). S.E.L. is a principal investigator of the German Center of Cardiovascular Research (DZHK).
Материалы
| Name | Company | Catalog Number | Comments |
| Chemicals and Enzymes | |||
| 2,3-Butanedione monoxime | Sigma-Aldrich, Munich, Germany | B0753 | |
| Bovine calf serum | Thermo Scientific, Schwerte, Germany | SH30073 | Triple 0.1 µm sterile filtered. |
| CaCl2 | Sigma-Aldrich, Munich, Germany | 21115 | Diluted 1:10 in MQ water to obtain 100 mM CaCl2 stock concentration. |
| Collagenase type II | Worthington via Cell Systems, Troisdorf, Germany | on request | Enzymatic activity depends on individual collagenase batches. Collagenase II and other enzyme activities (Caseinase, Clostripain, Tryptic) can be assessed in the "collagenase lot selection tool". Determine cell yield and quality individually for each new lot of collagenase. |
| Glucose | Carl Roth, Karlsruhe, Germany | HN06.1 | |
| Heparin | Rotexmedica, Trittau, Germany | PZN-03862340 | Diluted in 0.9% NaCl and injected subcutaneuosly in abdominal skin. |
| HEPES | Carl Roth, Karlsruhe, Germany | 9105.4 | |
| Forene 100% (V/V) | Abbott, Libertyville, IL, USA | B506 | Active agent: isoflurane, 250 ml. Use approximately 2 Vol% in air/oxygen dispenser instrument. |
| KCl | Carl Roth, Karlsruhe, Germany | 6781.3 | |
| KH2PO4 | Carl Roth, Karlsruhe, Germany | 3904.2 | |
| Laminin (2 mg/ml) | BD Biosciences, Heidelberg, Germany | 354232 | Lamination is described under step 2.1. |
| MgCl2·6H2O | Carl Roth, Karlsruhe, Germany | 2189.2 | |
| MgSO4·7H2O | Carl Roth, Karlsruhe, Germany | 8283.2 | |
| Na2HPO4·2H2O | Carl Roth, Karlsruhe, Germany | 4984.2 | |
| NaHCO3 | Carl Roth, Karlsruhe, Germany | HN01.1 | |
| Taurin | Carl Roth, Karlsruhe, Germany | 4721.2 | |
| Dyes | |||
| Di-8-ANEPPS | Molecular Probes, Life Technologies, Darmstadt, Germany | D-3167 | Stock solution 2 mM in DMSO |
| Trypan blue | Sigma-Aldrich, Munich, Germany | T8154 | Trypan blue is gently mixed 1:1 via tip-cut 1 ml plastic pipette with cell suspension prior to cell counting in Neubauer cytometer. |
| Langendorff Perfusion Setup | |||
| Circulation thermostat | Lauda, Lauda-Königshofen, Germany | Please refer to Louch et al. (JMCC 2011). Heat up thermostat und buffers in perfusion tubing to 37 °C 15 min prior to use. | |
| Flexible silicone tubing Tygon for peristaltic pump | VWR, Darmstadt, Germany | 224-2252 | Tubing needs to be changed regularly. |
| Flexible silicone tubing Tygon for thermostat | VWR, Darmstadt, Germany | 228-4340 | |
| Heating coil surroundung perfusion tubing | Rettberg, Göttingen, Germany | custom-made | Heating coil and tubing needs to be cleaned thoroughly via MQ water after using. Do not use detergents. Glass components should be bathed regularly in 10 mM NaOH overnight. |
| Peristaltic pump | Ismatec, Wertheim, Germany | ISM830 | |
| Three way stop cock Discofix C Luer Lock 10 cm | Braun, Melsungen, Switzerland | 16500C | |
| Three way stop cock Discofix 3SC | Braun, Melsungen, Switzerland | 4095146 | |
| Instruments | |||
| 42 mm glass coverslips | Menzel Gläser via Thermo Scientific, Schwerte, Germany | on request | 0.13-0.16 mm thickness |
| Cannula 21 G | Becton, Dickinson and Company, Franklin Lake, NY, USA | 304432 | Cut to a length of ~5 mm, roughened with sandpaper. |
| Coverslips for Neubauer cytometer 24 x 24 mm | Menzel Gläser via Thermo Scientific, Schwerte, Germany | on request | 0.38-0.42 mm thickness |
| Graefe forceps, 0.5 mm tips, slight curve | Fine Science Tools, Heidelberg, Germany | 11151-10 | |
| LSM 710 NLO | Carl Zeiss, Jena, Germany | 63X 1.4 NA oil objective | |
| Neubauer improved cytometer | Labor Optik, Friedrichsdorf, Germany | 1100000 | Counting procedure: Wipe cytometer and coverslip provided with the counting chamber with 70 % ethanol. Press coverslip gently on the counting chamber so that the two glass surfaces are in contact and Newton's rings can be observed. Subsequently, 10-20 µl cell suspension can be applied to the edge of the coverslip to be sucked into the void by capillary action. Count the intact vs. defect myocytes using the squares of the cytometer grid which reflects 100 nl. Repeat counting procedure on the second grid provided on the cytometer. Calculate the density of cells in your original cell suspension by taking account of any dilutions and counting shortcuts. |
| POC-R2 Imaging Chamber | Pecon, Erbach, Germany | Cell suspension volume: 800 µl; desired plating density: ~1,000 AM and ~10,000 VM | |
| Spring scissors, 8 mm blades straight, blunt | Fine Science Tools, Heidelberg, Germany | 15025-10 | |
| Student dumont #7 forceps, inox | Fine Science Tools, Heidelberg, Germany | 91197-00 | |
| Student iris scissors, curved, 11.5 cm | Fine Science Tools, Heidelberg, Germany | 91461-11 | |
| Student iris scissors, straight, 11.5 cm | Fine Science Tools, Heidelberg, Germany | 91460-11 | |
| Student surcigal scissors, straight, sharp, 12 cm | Fine Science Tools, Heidelberg, Germany | 91402-12 | |
| Tissue forceps, 1 x 2 teeth, slim, 10 cm | Fine Science Tools, Heidelberg, Germany | 11023-10 | |
Ссылки
- Prosser, B. L., Ward, C. W., Lederer, W. J. Subcellular Ca2+ signaling in the heart the role of ryanodine receptor sensitivity. J Gen Physiol. 136 (2), 135-142 (2010).
- Wehrens, X. H., Lehnart, S. E., Marks, A. R. Intracellular calcium release and cardiac disease. Annu Rev Physiol. 67, 69-98 (2005).
- Cheng, H., Cannell, M. B., Lederer, W. J. Propagation of excitation-contraction coupling into ventricular myocytes. Pflugers Arch. 428 (3-4), 415-417 (1994).
- Williams, G. S., Chikando, A. C., Tuan, H. T., Sobie, E. A., Lederer, W. J., Jafri, M. S. Dynamics of calcium sparks and calcium leak in the heart. Biophys J. 101 (6), 1287-1296 (2011).
- Sperelakis, N., Rubio, R. orderly lattice of axial tubules which interconnect adjacent transverse tubules in guinea-pig ventricular myocardium. J Mol Cell Cardiol. 2 (3), 211-220 (1971).
- Soeller, C., Cannell, M. B. of the transverse tubular system in living cardiac rat myocytes by 2-photon microscopy and digital image-processing techniques. Circ Res. 84 (3), 266-275 (1999).
- Song, L. S., et al. et al. ryanodine receptors in the failing heart. Proc Natl Acad Sci USA. 103 (11), 4305-4310 (2006).
- Oort, R. J., et al. Disrupted junctional membrane complexes and hyperactive ryanodine receptors after acute junctophilin knockdown in mice. Circulation. 123 (9), 979-988 (2011).
- Wagner, E., et al. Stimulated emission depletion live-cell super-resolution imaging shows proliferative remodeling of T-tubule membrane structures after myocardial infarction. Circ Res. 111 (4), 402-414 (2012).
- Asghari, P., Schulson, M., Scriven, D. R., Martens, G., Moore, E. D. Axial tubules of rat ventricular myocytes form multiple junctions with the sarcoplasmic reticulum. Biophys J. 96 (11), 4651-4660 (2009).
- Lukyanenko, V., Ziman, A., Lukyanenko, A., Salnikov, V., Lederer, W. J. Functional groups of ryanodine receptors in rat ventricular cells. J Physiol. 583 (Pt 1), 251-269 (2007).
- Shacklock, P. S., Wier, W. G., Balke, C. W. Local Ca2+ transients (Ca2+ sparks) originate at transverse tubules in rat heart cells. J Physiol. 487 (Pt 3), 601-608 (1995).
- Reynolds, J. O., et al. Junctophilin-2 is necessary for T-tubule maturation during mouse heart development. Cardiovasc Res. 100 (1), 44-53 (2013).
- Di Maio, A., Karko, K., Snopko, R. M., Mejia-Alvarez, R., Franzini-Armstrong, C. T-tubule formation in cardiac myocytes two possible mechanisms. J Muscle Res Cell Motil. 28 (4-5), 231-241 (2007).
- He, J., et al. Reduction in density of transverse tubules and L-type Ca(2+) channels in canine tachycardia-induced heart failure. Cardiovasc Res. 49 (2), 298-307 (2001).
- Heinzel, F. R., et al. Remodeling of T-tubules and reduced synchrony of Ca2+ release in myocytes from chronically ischemic myocardium. Circ Res. 102 (3), 338-346 (2008).
- Lyon, A. R., et al. Loss of T-tubules and other changes to surface topography in ventricular myocytes from failing human and rat heart. Proc Natl Acad Sci USA. 106 (16), 6854-6859 (2009).
- Crossman, D. J., Ruygrok, P. N., Soeller, C., Cannell, M. B. Changes in the organization of excitation-contraction coupling structures in failing human heart. PLoS One. 6 (3), e17901(2011).
- Kemi, O. J., et al. The effect of exercise training on transverse tubules in normal, remodeled, and reverse remodeled hearts. J Cell Physiol. 226 (9), 2235-2243 (2011).
- Sachse, F. B., et al. Subcellular structures and function of myocytes impaired during heart failure are restored by cardiac resynchronization therapy. Circ Res. 110 (4), 588-597 (2012).
- Arakel, E. C., et al. Tuning the electrical properties of the heart by differential trafficking of KATP ion channel complexes. J Cell Sciences. 127 (Pt 9), 2106-2119 (2014).
- Richards, M. A., et al. Transverse tubules are a common feature in large mammalian atrial myocytes including human. Am J Physiol Heart Circ Physiol. 301 (5), H1996-H2005 (2011).
- Trafford, A. W., Clarke, J. D., Richards, M. A., Eisner, D. A., Dibb, K. M. Calcium signalling microdomains and the t-tubular system in atrial mycoytes potential roles in cardiac disease and arrhythmias. Cardiovasc Res. 98 (2), 192-203 (2013).
- Greiser, M., Schotten, U. Dynamic remodeling of intracellular Ca2+ signaling during atrial fibrillation. J Mol Cell Cardiol. 58, 134-142 (2013).
- Voigt, N., Zhou, X. B., Dobrev, D. Isolation of human atrial myocytes for simultaneous measurements of Ca2+ transients and membrane currents. J Vis Exp. (77), 10-3791 (2013).
- Kaestner, L., et al. Isolation and genetic manipulation of adult cardiac myocytes for confocal imaging. J Vis Exp. (31), (2009).
- Louch, W. E., Sheehan, K. A., Wolska, B. M. Methods in cardiomyocyte isolation, culture, and gene transfer. J Mol Cell Cardiol. 51 (3), 288-298 (2011).
- King, N. M., et al. Mouse intact cardiac myocyte mechanics cross-bridge and titin-based stress in unactivated cells. J Gen Physiol. 137 (1), 81-91 (2011).
- Schindelin, J., et al. Fiji, an open-source platform for biological-image analysis. Nat Methods. 9 (7), 676-682 (2012).
- Arganda-Carreras, I., Fernandez-Gonzalez, R., Munoz-Barrutia, A., Ortiz-De-Solorzano, C. 3D reconstruction of histological sections Application to mammary gland tissue. Microsc Res Tech. 73 (11), 1019-1029 (2010).
- Liu, Z. Q. Scale space approach to directional analysis of images. Appl Opt. 30 (11), 1369-1373 (1991).
- Powell, T., Twist, V. W. A rapid technique for the isolation and purification of adult cardiac muscle cells having respiratory control and a tolerance to calcium. Biochem Biophys Res Commun. 72 (1), 327-333 (1976).
- Watanabe, Y., et al. Inhibitory effect of 2,3-butanedione monoxime (BDM) on Na(+)/Ca(2+) exchange current in guinea-pig cardiac ventricular myocytes. Br J Pharmacol. 132 (6), 1317-1325 (2001).
- Kohl, T., Westphal, V., Hell, S. W., Lehnart, S. E. Superresolution microscopy in heart - cardiac nanoscopy. J Mol Cell Cardiol. 58, 13-21 (2013).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены