
Method Article
Sıtma Anopheles Vektörlerinde SiteYe Yönelik φC31 Aracılı Entegrasyon ve Kaset Değişimi
Bu Makalede
Özet
Protokol, φC31 sistemini kullanarak Anopheles sıtma sivrisineklerinin genomunda bölgeye yönelik değişikliklerin nasıl eldeılacağını açıklar. Açıklanan değişiklikler, attP taşıyan yerleştirme hatlarının genomunda transgenik kasetlerin hem entegrasyonunu hem de değişimini içerir.
Özet
Fonksiyonel genomik analiz ve sıtmanın genetik kontrolü için ilgili stratejiler, Anopheles sivrisineklerinin genomlarını doğru bir şekilde değiştirmek için doğrulanmış ve tekrarlanabilir yöntemlere dayanır. Bu yöntemler arasında, φC31 sistemi transgenelerin hassas ve istikrarlı bir şekilde haline yönlendirilmesine veya entegre transgenik kasetlerin rekombinaz aracılı kaset değişimi (RMCE) yoluyla değiştirilmesine izin verir. Bu yöntem, attP (fajdan türetilmiştir) ve attB (konak bakteriden türetilmiştir) olarak belirlenen iki özel ek bölgesi arasındaki rekombinasyonu katalize etmek için Streptomyces φC31 bakteriyofaj integrazının etkisine dayanır. Sistem, donör şablonu DNA'sında sivrisinek genomuna ve attB bölgelerine daha önce entegre edilmiş bir veya iki ATTP sitesi kullanır. Burada, attP taşıyan Anopheles yerleştirme hatlarının genomunun iki plazmid kullanarak nasıl sağlam bir şekilde değiştirilerek değiştirilerek değiştiril şunları gösteriyoruz: entegrasyon veya değişim şablonunu taşıyan ATTB etiketli bir donör ve φC31 integrase'yi kodlayan bir yardımcı plazmid. φC31 aracılı siteye yönelik modifikasyonun iki temsili sonucu rapor ediyoruz: An. gambiae sivrisineklerinde An. stephensi ve RMCE'de transgenik bir kasetin tek entegrasyonu. φC31 aracılı genom manipülasyonu, doğrulanmış, fitness nötr genomik bölgelerden tekrarlanabilir transgene ekspresyon avantajı sunarak fenotiplerin karşılaştırmalı nitel ve nicel analizlerine olanak sağlar. Entegrasyonun siteye yönelik doğası, kararlı bir transgenik hat elde etmek için tek ekleme sitesinin ve çiftleşme düzeninin doğrulanmasını da önemli ölçüde basitleştirir. Bu ve diğer özellikler, φC31 sistemini sıtma sivrisineklerinin ve diğer böcek vektörlerinin transgenik manipülasyonu için genetik araç setinin önemli bir bileşeni haline getirir.
Giriş
Hastalıkların sivrisinek vektörlerinin genomlarını güvenilir ve tekrarlanabilir bir şekilde değiştirme yeteneği, genlerin in vivo fonksiyonel doğrulamasını güçlendirmiş ve sıtmayı ileten Anopheles sivrisineklerini hedef alanlar gibi gerçekleştirilebilir genetik vektör kontrol stratejilerine kapı açmıştır1.
Erken sivrisinek genom düzenlemesi sadece transpoze edilebilir eleman (TE) aracılı dönüşüme dayanıyordu ve piggyBac Anopheles2,3,4'te en sık kullanılan transposondu. Bununla birlikte, TE entegrasyonunun rastgele doğası, gen nakavtları (eklemeli mutagenezi) ve transgene ekspresyonu üzerinde önemli konum etkileri gibi istenmeyen değişikliklere yol açabilir5,6,7,8. Birden fazla ekleme, tek eklemelerle transgenik çizgilerin doğrulanmasını ve yalıtılmasını zahmetli hale getiren piggyBac5,9 kullanırken de yaygın bir durumdur. Diğer dezavantajları, bir domuzbac transposase10,11,12 kaynağı sağlarken Anopheles stephensi'nin germline'ında gözlemlendiği gibi potansiyel yeniden hareketlenmelerini ve donör plazmid13,14'ün artan boyutuyla azalan dönüşüm verimliliği ile sınırlı DNA kargo boyutlarını (10-15 kb uzunluğunda) içerir.
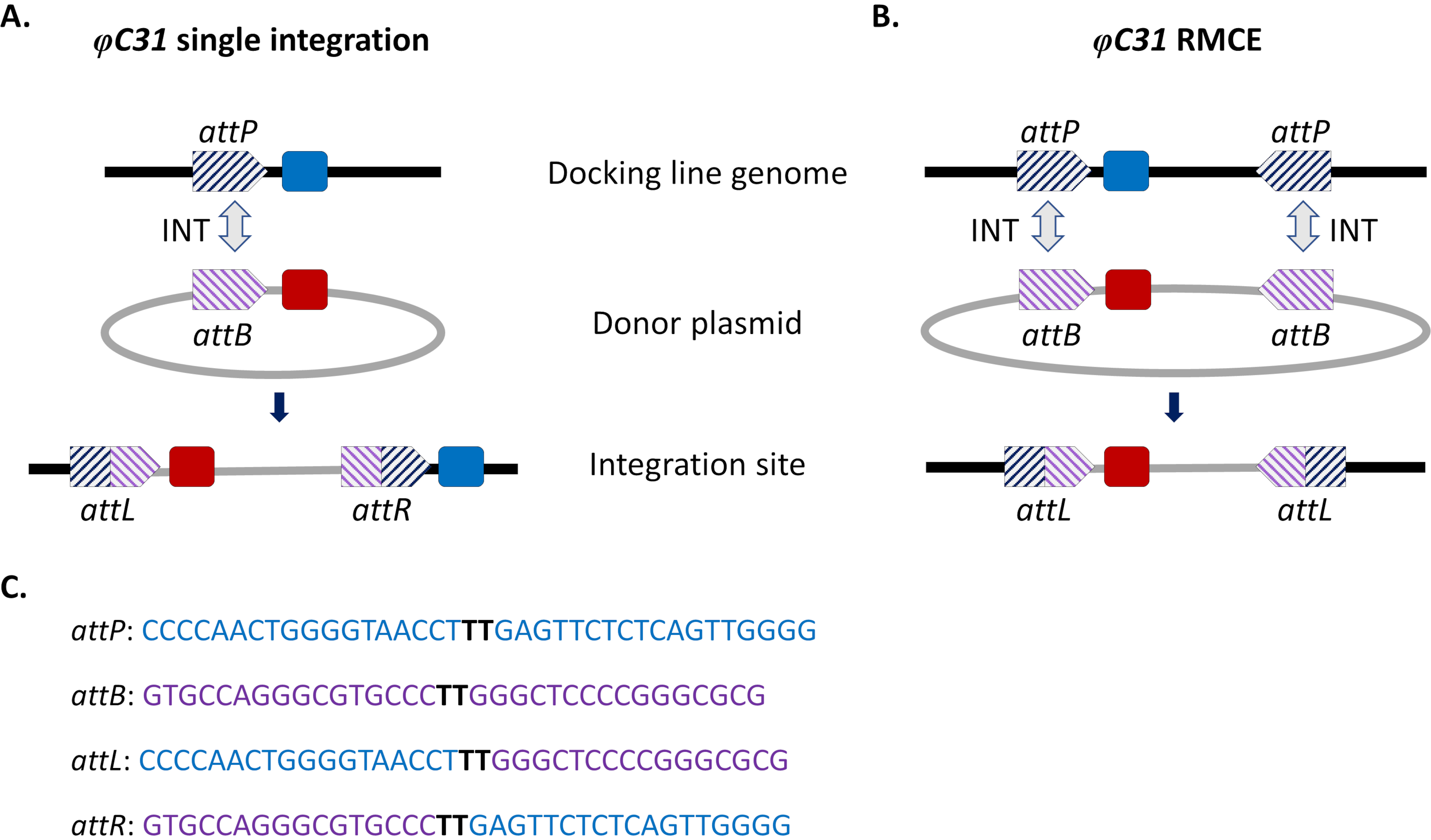
Bu sorunları atlatmak için siteye yönelik entegrasyon yaklaşımları getirilmiştir. Sivrisineklerde en yaygın bölgeye yönlendirilen genom modifikasyonu, φC31 sisteminin aracılık ettiğidir (Şekil 1a). Bu, bakteriyofaj φC31 (attP) genomunda ve Streptomices bakteri konağında (attB)15'te doğal olarak meydana gelen iki heterospesifik bağlanma (att) bölgesi arasındaki rekombinasyonu katalizleyen viral bir integrase tarafından yönlendirilir. İki bölgenin yeniden birleşmesi tek yönlüdür ve hibrit bölgelerin (attL ve attR) oluşumuyla sonuçlanır. Bu tür melez bölgelerin yeniden bireşmesi (DNA eksizyona yol açan) sadece aktif bir viral integrazın varlığını değil, aynı zamanda başka bir faj kodlu rekombinasyon faktörünü de gerektirecektir16,17. Böylece, potansiyel istenmeyen yeniden hareketlenme sorununu hafifleten kararlı bir entegrasyon sitesi oluşturulur15. Ayrıca, sistem büyük yüklerin entegrasyonuna izin verir (örneğin, D. melanogaster18'de >100 kb yapı entegrasyonu bildirilmiştir), taşıma kapasitelerini önemli ölçüde artırmaktadır. Entegrasyon, kararlı bir transgenik hat elde etmek için ekleme ve çiftleşme şemasının doğrulanmasını büyük ölçüde basitleştiren önceden tanımlanmış tek bir genomik lokusta gerçekleşir. Son olarak, entegrasyonun meydana gelen doğası, alternatif transgenler aynı çekirgede bulunduğundan ve bu nedenle aynı komşu genomik bağlamda düzenlendiğinden ifadenin normalleşmesine izin verir. Aslında, tekniğin ana uygulamalarından biri, farklı transgenler tarafından aynı lokusa yerleştirildiğinden sonra verilen fenotiplerin doğrudan karşılaştırılmasıdır.
φC31 aracılı entegrasyona ulaşmak iki aşamayı içerir: aşama I, ATTP sahalarını taşıyan transgenik yerleştirme hatlarının oluşturulmasıdır ve aşama II, attB tarafından kuşatılmış bir kargonun yerleştirme hattının genomuna siteye yönelik entegrasyonudur19. Faz I yerleştirme hatlarının oluşturulması, ATTP etiketli yapıların TE aracılı rastgele entegrasyonuna dayanmıştır ve bu nedenle benzersiz, transkripsiyonel olarak aktif ve fitness nötr genomik konumlarda tek bir entegrasyon olayı taşıyan transgenik çizgileri izole etmek ve doğrulamak için ilk zahmetli bir süreci (tek kadın soylar üzerinde güney lekesi ve ters PCR analizleri dahil) içermektedir. Bununla birlikte, An. gambiae19,20,21,22 ve An. stephensi23,24,25'te (Tablo 1) φC31 aracılı tek entegrasyon için birkaç yerleştirme hattı geliştirilmiş ve doğrulanmıştır. Bu çizgilerin her biri, yanaşma bölgesinin genomik konumu ve suşlara özgü genetik arka plan açısından değişir ve bunlardan çok çeşitli yeni transgenik çizgiler oluşturulabilir. Yerleştirme hatları üretmek için TE aracılı entegrasyonların karmaşık doğrulaması artık CRISPR/Cas9 teknolojisi tarafından atlatılabilir26; ancak bu, hedeflenecek nötr loci ve çevresindeki diziler hakkında a priori bilgisine dayanır.
φC31 aracılı entegrasyon, model organizma D. melanogaster27'den Aedes aegypti13,28, Ae. albopictus29, An. gambiae19 ve An. stephensi24'ün yanı sıra Ceratitis capitata30 ve Bombyx mori31 gibi diğer böceklere kadar böcek genom düzenlemesine kapsamlı bir şekilde uygulanmıştır.
φC31 aracılı entegrasyonun bir sınırlaması, özellikle vektör kontrolü için potansiyel alan salınımları göz önüne alınarak, antibiyotik direnci gen belirteçleri ve bakteri kökenli plazmid omurga bileşenleri gibi istenmeyen diziler de dahil olmak üzere tüm ATTB taşıyan donör plazmidinin sivrisinek genomuna entegrasyonudur. Bunu gidermek için, daha önce entegre edilmiş bir transgenik kasetin yeni bir donör DNA'sı ile kesin olarak değiştirilmesini sağlayan standart sistemin, rekombinazi aracılı kaset değişiminin (RMCE) bir modifikasyonu uygulanmıştır (Şekil 1b). Bu, her iki uçta donör ve alıcı kasetleri kuşatan iki ters att bölgesi kullanılarak elde edilir, bu da iki bağımsız rekombinasyon olayının aynı anda gerçekleşmesini sağlar ve plazmid omurgası entegre edilmeden kaset değişimi ile sonuçlanır. Bu geliştirilmiş tasarım, istenmeyen dizilerin entegrasyonunu atlatır ve φC31 sistemlerinin uygulamasını, örneğin daha önce entegre edilmiş bir floresan marker32'nin kaybını tarayarak işaretlenmemiş DNA kargolarının entegrasyonunu içerecek şekilde genişletir.
RMCE ilk olarak D. melanogaster32 ile elde edildi ve daha sonra An. gambiae9,26,33, Ae. aegypti34, Plutella xylostella34 ve B. mori35 dahil olmak üzere model olmayan böceklere başarıyla uygulandı. An. gambiae5,9,26 'da RMCE için çeşitli yerleştirme hatları geliştirilmiş ve doğrulanmıştır (Tablo 1). Bilgimize göre, RMCE henüz diğer Anopheles vektör türlerinde araştırılmamıştır.
Bugüne kadar, φC31 sistemi, antimalaria efektörleri19,24,36, GAL4/UAS sisteminin bileşenleri, insektisit direnci çalışmaları için genleri aşırı ifade etmek ve devirmek için çeşitli molekülleri tanıtmak ve incelemek için Anopheles sivrisineklerinde yaygın olarak kullanılmıştır9,33, düzenleyici elementler, muhabir genleri5,21,37 ve gen tahrik elemanları26 ,38.
Bu protokol, 1) ATTB tarafından kuşatılmış bir kargonun saha tarafından yönlendirilen entegrasyonunun ve 2) ters attB siteleri tarafından kuşatılmış bir yapının RMCE'nin Anopheles yerleştirme hatlarının genomuna nasıl gerçekleştirildiğini açıklar. Bu, iki plazmid kullanılarak elde edilir: ilginin transgenesini taşıyan bir donör attB etiketli plazmid ve φC31 integrazını ifade eden bir yardımcı plazmid. Başlıca sıtma vektörleri An. gambiae ve An. stephensi özel örnekler olarak kullanılır, ancak bu protokoller diğer Anopheles türleri için geçerlidir.

Şekil 1. φC31 sistemi kullanılarak raya yönlendirilmiş genom modifikasyonları, tek entegrasyon ve rekombinoz aracılı kaset değişimi (RMCE). φC31 integraz (INT, gri çift ok), donör plazmidinde bulunan ATTB siteleri (mor çizgili) ile alıcı yerleştirme hattında bulunan attP siteleri (mavi çizgili) arasındaki yeniden birleştirmeyi katalizörler ve bu da attL ve attR'de hibrit sitelerin oluşmasına neden olur. A) Entegrasyon, tek attB ve ATTP siteleri yeniden birleştirildiğinde ve iki entegre işaretleyicinin (mavi ve kırmızı) varlığıyla sonuçlandığında elde edilir. B) RMCE, iki attB/P bölgesi aynı anda yeniden birleşince ortaya çıkar ve kasetin yerleştirme hattının att bölgeleri (mavi işaretleyici) arasında donör plazmid (kırmızı işaretleyici) tarafından taşınanla değiştirilmesiyle sonuçlanır. C) AttP (mavi) ve attB (mor) ve hibrid siteler attL / R kısmi nükleotid dizileri. Yeniden birleştirme, koyu siyah renkle vurgulanan 'TT' çekirdek dizileri arasında gerçekleşir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Protokol
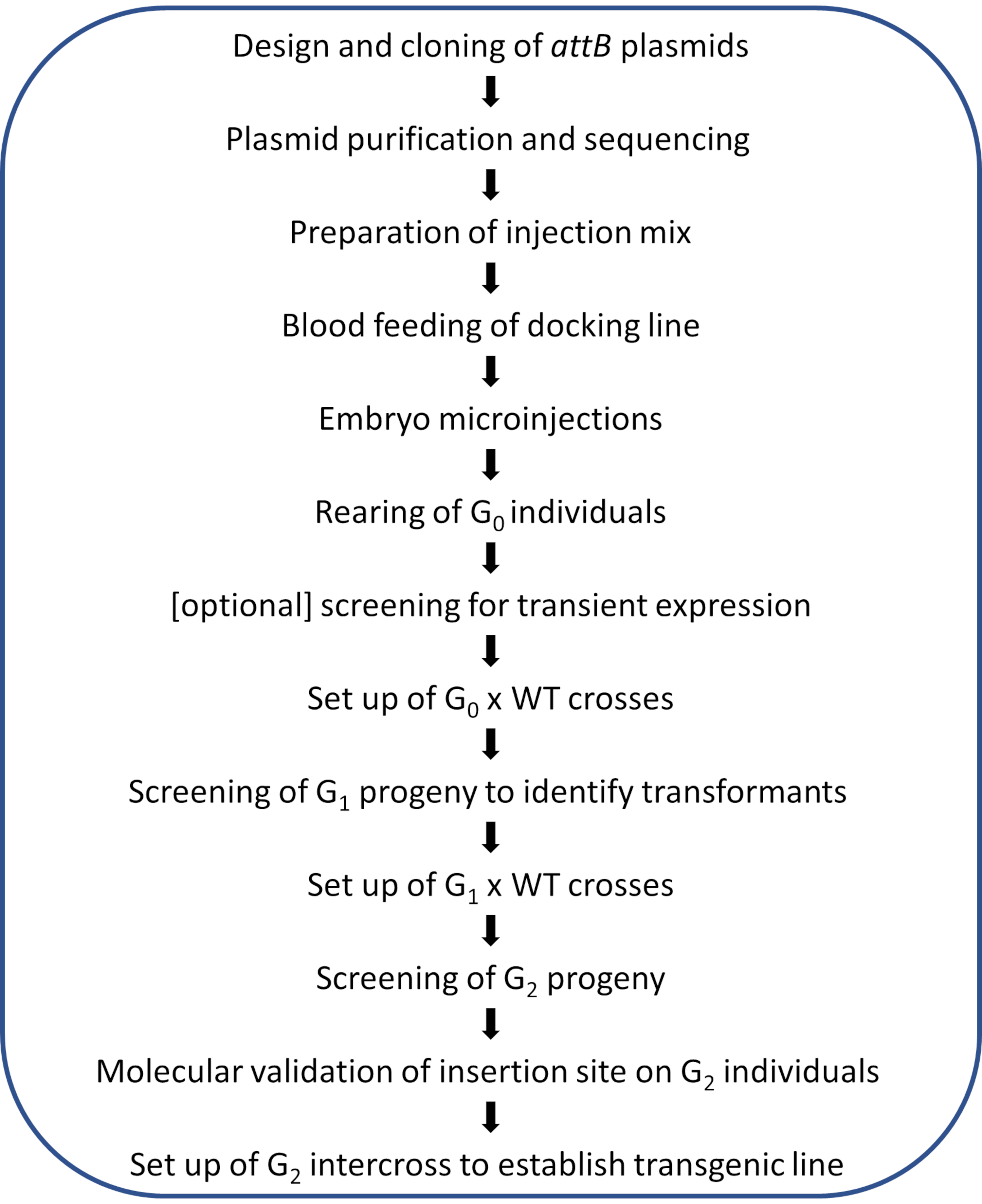
NOT: Şekil 2'de resimli protokolün şematik iş akışı gösterilmiştir.
1. φC31 attB etiketli plazmidlerin tasarımı (Şekil 3)
- Aşağıdaki temel bileşenleri taşıyan ATTB donör plazmidleri oluşturun
- Baskın floresan işaretleyici
- Floresan işaretleyicinin ifadesini yönlendirmek için bir organizatör seçin.
NOT: Anopheles transgenez için floresan belirteçler genellikle gözlerde ve sinir kordonunda ifadeyi yönlendiren 3xP3 promotör39'un düzenlemesi altındadır. Alternatif olarak, PUBc promotörü5 , birden fazla dokuda ifade istendiğinde kullanılabilir. Bu protokolde örnek olarak kullanılan donör plazmidleri ve yerleştirme hatları 3xP3 promotörü kullanılarak işaretlenir. - Kolayca ayırt edilebilmeleri için alıcı yerleştirme hattınınkiyle uyumlu bir floresan protein (FP) seçin.
NOT: Yerleştirme hattında zaten bulunan aynı işaretleyiciyi kullanmayın ve güvenilir bir şekilde ayırt etmek çok zor olduğu için GFP (yeşil)/YFP (sarı) ve GFP (yeşil)/CFP (siyan) aynı anda kullanılmasını önleyin. Bu protokolde örnek olarak kullanılan donör plazmidleri, CFP ile işaretlenmiş bir yerleştirme hattına entegre edilecekleri için DsRed veya YFP ile işaretlenir.
- Floresan işaretleyicinin ifadesini yönlendirmek için bir organizatör seçin.
-
attB yeniden birleştirme sahaları
- Transgenik kasetin entegrasyonu için tek bir attB sitesi kullanın (tek attB tasarımı) (Şekil 3A).
- RMCE (çift attB tasarımı) için iki ters attB sitesi kullanın, burada siteler birbirine göre ters çevrilmiş olarak uzanır ve donör DNA şablonünü çevreler (Şekil 3B).
NOT: ATTB sitelerinin yönü, yerleştirme satırında bulunan ATTP sitelerininkiyle uyumlu olmalıdır.
- İstenen transgene kargo
- Deneyin özel amacına dayanarak sivrisinek genomuna entegre olmak için istenen diğer özellikleri kullanın. Burada, bir antimalarial efektör molekülün An. stephensi'nin genomuna entegrasyonunu ve GAL4/UAS sisteminin bileşenlerinin An. gambiae sivrisineklerine entegrasyonunu anlatıyoruz.
- Plazmid omurga bileşenleri
- Bakterilerde plazmid replikasyon için diğer temel bileşenlerin yanı sıra, in vitro (yani bir antibiyotik direnç geni) plazmid seçimi için bir işaretleyici içerir.
NOT: Plazmid omurga, entegrasyon için tek attB tasarımında sivrisinek genomuna entegre edilecektir (Şekil 3A), RMCE için çift attB tasarımına eklenmeyecektir (Şekil 3B).
- Bakterilerde plazmid replikasyon için diğer temel bileşenlerin yanı sıra, in vitro (yani bir antibiyotik direnç geni) plazmid seçimi için bir işaretleyici içerir.
- Baskın floresan işaretleyici
2. Mikroenjeksiyon karışımı için plazmidlerin hazırlanması
NOT: Burada gösterilen protokol iki plazmid kullanımını içerir: ilgi transgene taşıyan attB etiketli donör plazmid ve Drosophila Hsp70 promotör40 düzenlemesi altında φC31 bütünlüğünü ifade eden bir yardımcı plazmid.
- Endotoksin içermeyen plazmid saflaştırma kiti kullanarak donör ve yardımcı plazmidleri saflaştırın.
NOT: Tüm bileşenlerin bütünlüğünü doğrulamak için enjeksiyon için kullanılan son plazmid hazırlığını sıralayın. - Enjeksiyon tamponunda yeniden sulandığında donör plazmidinin 350 ng/μL'si ve yardımcı plazmid'in 150 ng/μL'lik son konsantrasyonu ile bir karışım elde etmek için iki plazmidin uygun miktarlarını birleştirin.
NOT: Gerekli karışım hacmini hesaplarken, planlanan enjeksiyonların her günü için 10-15 μL'nin yeterli olduğunu ve DNA'nın önceden hazırlanabileceğini ve -20 °C'de saklanabileceğini düşünün. Integrase yardımcı plazmid konsantrasyonları 60-500 ng/μL ve donör plazmid konsantrasyonları 85-200 ng/μL de bildirilmiştir21,22,26,41. - 0.1 cilt 3 M sodyum asetat (pH 5.2) ve 2.5 hacim buz gibi 100% EtOH ve girdap ekleyerek DNA'yı çökeltin. Beyaz bir çökeltme hemen görülebilir olmalıdır. Yüksek yoğunluklu ilk plazmid preparatlarına (yani ~ 1 μg / μL) sahip olmak yağış verimliliğini artırır.
NOT: Durma noktası - Çökelt bir gecede -20 °C'de saklanabilir. - 4 °C'de 20 dakika boyunca 15.000 x g'da santrifüj, süpernatantı atın ve peletin 1 mL buz gibi% 70 EtOH ile yıkayın.
- Peletin 1 mL buz gibi %70 EtOH ve santrifüj ile 15.000 x g'da oda sıcaklığında 5 dakika yıkayın.
- Pelet ve hava kuru rahatsız etmeden süpernatant atın.
- Peleti 1x enjeksiyon tamponunda (0,1 mM Na3PO4, 5 mM KCl, pH 7,2, 0,22 μm filtre sterilize edilmiş) toplam 500 ng/μL nihai konsantrasyona ulaşmak için yeniden biriktirin.
NOT: Yağış işlemi sırasında bazı DNA'ların kaybolacağını varsayalım; Bu nedenle, önce daha küçük bir enjeksiyon tamponu ekleyin, spektrofotometredeki konsantrasyonu kontrol edin (örneğin, Nanodrop) ve ardından 500 ng / μL'ye ulaşmak için uygun bir kalan hacim ekleyin. - DNA'nın iyice yeniden depolanıp saklanmadığına emin olun, her biri 10-15 μL'lik aliquots hazırlayın ve -20 °C'de saklayın.
- Enjeksiyon gününde, partikül kalıntılarını gidermek için 5 dakika boyunca 15.000 x g'da bir aliquot ve santrifüj çözün.
NOT: Partikül giderme için alternatif bir yöntem, çözeltiyi 0,22 μm filtreden filtrelemektir. Embriyo mikroenjeksiyonu sırasında iğne tıkanmasına yol açan enjeksiyon karışımında partikül kalıntılarının bulunmasından kaçının.
3. Anopheles yanaşma hattından embriyoların mikroenjeksiyon
- Kan besleme 4-7 günlük sivrisinekler mikroenjeksiyondan önce istenen yanaşma hattından 72 saat (yani, Pazartesi ve Salı günü enjeksiyon için bir önceki Cuma günü dişileri besler; Perşembe ve Cuma günü enjeksiyon için aynı haftanın Pazartesi günü dişileri besler).
- Kan, vahşi tip (WT) sivrisinekleri (yani, yanaşma hattının aynı genomik arka planına sahip sivrisinekler) aynı gün besler; bunlar outcrossing için gerekli olacaktır.
NOT: Kan uzamasının büyüklüğü ve kalitesi yumurta kalitesini etkiler, bu nedenle her zaman taze kan kullanılması önerilir (yani, önceki 7 gün içinde çekilen kan). Farelerin kolla beslenmesi veya beslenmesi yumurtaların kalitesini ve miktarını artırabilir, ancak bu yöntemler teşvik edilir. İnsan ve hayvan kullanımı için belirli onaylanmış protokoller gerekli olacaktır. - Embriyo mikroenjections gerçekleştirin
- An. gambiae embriyo mikroenjeksiyonlarını 25 mM NaCl42'de embriyonun arka kutbuna 45 derecelik bir açıyla hedef alarak gerçekleştirin. Embriyo toplama, hizalama ve mikroenjeksiyon için ayrıntılı bir protokol için Pondeville ve ark.43 ve Lobo ve ark.44'e bakın.
- Halekkarbon yağı 700:27 (2:1) içinde an. stephensi embriyo mikroenjections gerçekleştirmek 30 derecelik bir açıyla embriyonun arka kutbu hedef. Embriyo toplama, hizalama ve mikroenjeksiyon için ayrıntılı bir protokol Terenius ve ark.45 ve Lobo ve ark.44'te bulunabilir.
- Yumurtaları steril damıtılmış su (pH 7.2) ile dolu bir Petri kabına enjeksiyondan hemen sonra aktarın ve böcek koşullarına geri döndürün.
- Kuluçkadan çıktıktan sonra, G0 larvalarını günlük tuzlu damıtılmış su (% 0.1 tonik tuz) içeren bir tepsiye aktarın ve pupalara arka.
- Kuluçka oranını kaydedin (yani, yumurtadan çıkan larva sayısı/enjekte edilen embriyo sayısı).
NOT: Embriyo hareketi kuluçkaya yardımcı olur, bu nedenle nazik girdap arzu edilir. Kuluçka enjeksiyondan sonra ~ 48 saat başlamalıdır. Enjeksiyon hafif bir gelişimsel gecikmeye neden olabileceğinden, geç kuluçkalayan larvaları 3-4 gün boyunca izlemeye devam etmeniz önerilir.
4. Dönüştürülmüş bireylerin geçişi ve taranır
- [İsteğE BAĞLI ADIM] Floresan işaretleyicinin geçici ekspresyoyu için ekran G0 (enjekte) 1. veya 2.
- G0 L1-L2 larvalarını kuyulu mikroskop slaytlarına aktarmak için ince uçlu bir cam pipet kullanın. Her kuyuya bir larva yerleştirin.
- Floresan işaretleyicinin geçici ifadesinin varlığını taramak için uygun filtreye sahip bir floresan stereoskop kullanın.
NOT: Geçici ifade deseni, kullanılan organizatör tarafından dikte edilir. 3xP3 promotörü kullanırken, anal papillada floresan işaretleyicinin geçici ifadesi görülebilir (Bkz. Pondeville ve ark.43'teki Şekil 6) - Arka G0 pozitif bireyler ayrı ayrı.
- G0 pupalarını stereoskop altında sekse göre sıralayın52.
- Erkeklerin 3-5 kişilik gruplar halinde ayrı kafeslerde ortaya çıkmasına izin verin (kurucu aileler) ve yaşla eşleşen WT kadınlarının 10 kat fazlalığını ekleyin.
NOT: Erkekler birden fazla kez çiftleştiğinden, her erkeğin çiftleşme şansını en üst düzeye çıkarmak için fazla miktarda WT dişisi sağlamak önemlidir. - Dişilerin 10-15 kişilik gruplar halinde ayrı kafeslerde ortaya çıkmasına izin verin (kurucu aileler) ve eşit sayıda yaş uyumlu WT erkeği ekleyin.
NOT: Böcekçide sınırlı alan varsa, dişiler tek bir kafeste birlikte ortaya çıkabilir. Kadın-erkek oranı 1 erkek ila 3 kadın kadar düşük olabilir. - Yetişkinlerin 4-5 gün çiftleşmelerine izin verin ve kadınlara bir kan unu sağlayın.
NOT: Kan, birden fazla gonotrofik döngüden dönüştürücü alma şansını en üst düzeye çıkarmak için G0 dişilerinden birçok kez yumurta besler ve toplar. - Kan besleme WT bireyleri aynı zamanda outcrossing için.
- Yumurtaları ve ortaya çıkan yeni nesil G1'leri toplayın.
- Dönüştürücüleri tanımlamak için uygun floresan için G1 L3-L4 larvalarını tarayın.
- Larvaları filtre kağıdı ile kaplı bir Petri kabında veya attB etiketli kargo ile tanıtılan işaretleyicinin varlığı için uygun filtrelere sahip floresan bir stereoskop kullanarak mikroskop slayt ve ekranında toplayın.
NOT: 3xP3 promotörü tarafından yönlendirilen floresan tüm postembriyonik aşamalarda görülebilir ve tarama daha genç larvalar üzerinde yapılabilir, ancak bunlar daha kırılgandır ve nispeten dikkatli bir şekilde ele alınmalıdır. Pupalar da taranabilir.- Yeni ve önceden var olan işaretleyicinin varlığı için entegrasyon ekranı için tek attB tasarımlar için; her ikisi de mevcut olmalıdır, çünkü yeni kaset orijinalinin yanına yerleştirilmiştir (Şekil 3A, Şekil 4).
NOT: Tek attB tasarımları için tarama istisnası: İşaretsiz yerleştirme çizgileri22 kullanırken, yalnızca yeni işaretçinin varlığı için ekran. Tümleştirmenin önceden varolan marker21'in devre dışı bırakılmasıyla sonuçlandığı yerleştirme çizgilerini kullanırken, yeni işaretçinin varlığını ve önceden varolanın kaybını tarayın. - RMCE için çift attB tasarımları için, yeni işaretleyicinin varlığı ve önceden var olanın kaybı için ekran, yeni kaset orijinalinin yerini aldığından yalnızca yeni tanıtılan işaretleyici bulunmalıdır (Şekil 3B, Şekil 5).
NOT: Yalnızca tek bir attP'nin yeniden birleştiği ve böylece her iki işaretin de bulunacağı RMCE denemelerinde zaman zaman tümleştirme olayları kurtarılabilir. G1 bireylerinin taranma işlemi de aynı prosedürü takip eden pupa aşamasında yapılabilir52.
- Yeni ve önceden var olan işaretleyicinin varlığı için entegrasyon ekranı için tek attB tasarımlar için; her ikisi de mevcut olmalıdır, çünkü yeni kaset orijinalinin yanına yerleştirilmiştir (Şekil 3A, Şekil 4).
- Larvaları filtre kağıdı ile kaplı bir Petri kabında veya attB etiketli kargo ile tanıtılan işaretleyicinin varlığı için uygun filtrelere sahip floresan bir stereoskop kullanarak mikroskop slayt ve ekranında toplayın.
- Transfer, G1 bireylerini bir larva tepsisine ve pupalara geri döndürür. Floresan olmayan bireyleri ve beklenmedik bir işaretleyici ifade deseni olan bireyleri atın.
- Dönüştürülmüş G1 pupalarını cinsiyete göre sıralayın ve karşı cins yaşla eşleşen WT bireyleri ile toplu olarak geçin.
- Yetişkinlerin 4-5 gün çiftleşmelerine, bir kan unu sağlamalarına, yumurtaları toplamalarına ve yeni nesil G2 soylarını yetiştirmelerine izin verin.
- Tek entegrasyon deneyleri için, entegrasyon sitesi tüm bireylerde aynı olduğundan, yumurtaları doğrudan toplu haçtan toplayın.
- RMCE deneyleri için, tek dişilerden yumurta toplayın ve iki alternatif kaset yönünün potansiyel varlığı nedeniyle moleküler değerlendirme tamamlanana kadar soyu ayrı tutun (Şekil 3B).
- Floresan işaretleyicinin varlığı için G2 soyunun (larva veya pupa aşamasında) taranması (bireylerin% 50'sinin pozitif olması beklenir), floresan olmayan soyları atın.
- Moleküler analiz için G2 pozitif bireylerin bir alt kümesini bir kenara bırakın, gerisini yetişkinliğe geri alın.
NOT: Tüm G2 bireylerinin hayatta tutulması gerekiyorsa, tek yetişkin bacaklarında moleküler analiz yapılabilir46 veya pupal vaka DNA ekstraksiyonları (L. Grigoraki kişisel iletişim). Alternatif olarak, tüm G2 bireyleri yumurtladıktan ve yumurtalar yumurtadan çıktıktan sonra moleküler analiz yapılabilir. - Yetişkin erkek ve dişilerin yeni transgenik hattı kurmak için aynı kafeste çapraz yapmalarına izin verin.
NOT: RMCE deneyleri için, moleküler analiz yoluyla ekleme oryantasyonu belirlenene kadar tek bir kadından türeyen kardeşler arasında yetişkin intercross meydana gelmelidir.
5. Ekleme bölgesinin DNA amplifikasyonu (PCR) ile moleküler olarak doğrulanması
- Dönüşümden sonra yerleştirme hattının genomunda öngörülen ekleme bölgesinin bir haritasını hazırlayın.
- Tek entegrasyon: Tahmin edilen ekleme bölgesinin orijinal yerleştirme yapısına ek olarak iki hibrit site attL ve attR arasındaki donör plazmid dizisinin tamamını taşıdığından emin olun (Şekil 3A).
- RMCE: Tahmin edilen ekleme sitesinin, karma ters çevrilmiş attL sitelerinin orijinal ters attP sitelerinin yerini aldığı ve değişim şablonunun aralarında orijinal olarak bulunan kasetin yerini aldığı yerleştirme satırıyla aynı olduğundan emin olun (Şekil 3B).
- Entegrasyon locusunun her iki tarafındaki ekleme birliğini güçlendirmek için oligonükleotid astarlar tasarlayın.
- Tek entegrasyon: AttR ve/veya attL sitelerine yayılan oligonükleotid astar çiftleri tasarlayın. Astarlardan biri önceden entegre edilmiş yerleştirme yapısına, diğeri de yeni entegre edilmiş transgeneye bağlanmalıdır (Şekil 3A).
- RMCE: Kaset değişimi kromozom (A ve B olarak belirlenmiştir) açısından iki farklı yönde gerçekleşebilir. 4 oligonükleotid astarın alternatif kombinasyonlarını tasarlayarak, bir çift A oryantasyonu için, diğeri ise B oryantasyonu için tanılama olmak üzere oryantasyonlardan sadece birinde ayrı bir ürün vermek için (Şekil 3B, Şekil 6).
- G2 pozitif bireylerden genomik DNA çıkarın ve öngörülen entegrasyon alanı haritalarından beklenen tanısal ampliconların varlığını görselleştirmek için tanısal PCR ve jel elektroforezi gerçekleştirin.
NOT: DNA alternatif olarak tek yetişkin bacaklarından 46 veya pupal vakalardan (L. Grigoraki kişisel iletişim) çıkarılabilir. - Beklenen dizileri onaylamak için PCR ürünlerini sıralayın.

Şekil 2. Anopheles sivrisineklerinde siteye yönelik φC31 genom modifikasyonu için iş akışı diyagramı. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3. φC31 aracılı tek entegrasyonun moleküler temeli (A) ve RMCE (B). A) Tek bir ATTP alanı taşıyan ve CFP (üstte), DsRed (ortada) ile işaretlenmiş tek attB tasarım donör plazmidi ve başarılı entegrasyondan sonra ortaya çıkan beklenen ekleme bölgesi (altta) ile işaretlenmiş bir An. stephensi yerleştirme hattındaki (80.9, Tablo 1) genomik yerleştirmenin şematik haritaları. B) İki ters ATTP bölgesi taşıyan ve CFP (üstte), YFP (ortada) ile işaretlenmiş çift attB tasarım donör plazmidi ve başarılı RMCE 'den (altta) sonra ortaya çıkan beklenen ekleme bölgesi ile işaretlenmiş bir An. gambiae yerleştirme hattındaki (A11, Tablo 1) genomik yerleştirmenin şematik haritaları. Dalgalı çizgi: sivrisinek genom; Çizgili oklar: piggyBac transposon kolları; 3xP3: floresan işaretleyicinin promotörü; SV40: viral sonlandırıcı; Ori: çoğaltmanın kaynağı; AmpR: ampisilin direnç geni. Geçiş çizgileri, attP ve attB siteleri arasındaki yeniden birleştirme alanlarını temsil eder. Numaralı siyah oklar, ekleme locusunun moleküler doğrulaması için astar bağlama bölgelerini temsil eder (protokolün 5. adımı). Tamamen açıklamalı tek ve çift attB etiketli plazmidler talep üzerine yazarlardan temin edilebilir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Sonuçlar
Burada gösterilen protokol, ~10 hafta içinde (21 günlük bir sivrisinek yaşam döngüsü varsayarak) kararlı bir Anopheles transgenik hattı oluşturulmasını sağlar.
An. gambiae'de enjeksiyon sonrası larva kuluçka oranlarının genellikle An. stephensi'den daha düşük olması beklenmektedir, ancak% 10-50 arasındaki kuluçka oranları 9,20,24,26,33,43,47 bildirilmiştir. Uygun enjeksiyon tekniği göz önüne alındığında, kuluçka oranları genellikle transformantları elde etmek için% ≥20 yeterlidir. Embriyolar tarafından DNA alımı, genç larvaların floresan belirtecin geçici ekspresyasyonu için taranarak değerlendirilebilir. Hayatta kalan G0 larvalarının% 50'sine kadar 3xP3 promotörü kullanılarak An. gambiae'deki başarılı RMCE deneylerinde, anal papillae48'deki işaretleyicinin epizomal ifadesini gösterdi.
Genelleştirilmiş dönüşüm verimliliği tahminlerinin laboratuvarlar arasında ve hatta deneyler arasında değerlendirilmesi zordur, çünkü dönüşüm, enjekte edilen DNA'nın saflığı, konsantrasyonu, büyüklüğü ve potansiyel toksisitesi, yumurtaların kalitesi, yumurtaların enjeksiyon öncesi ve sonrası kullanımı, sivrisinek yetiştirme ve en önemlisi operatörün deneyimi dahil olmak üzere değişkenlerin karmaşık bir etkileşimine bağlıdır. An. gambiae'deki RMCE için %7'ye kadar dönüşüm oranları elde edilmiştir (toplam G0 bireylerdeki bağımsız dönüşüm olaylarının sayısı olarak hesaplanır)9,26,33 ve An. stephensi'ye entegrasyon için %2,2'ye kadar dönüşüm oranı. En az 100 G0 larvasının kuluçkalanmasına ve saplanarak dönüştürülmüş soy elde edilebilen 2-7 G0 yetişkin kurucuya yol açması gereken en az 500 embriyo enjekte etmeyi öneriyoruz. G0 larvalarında geçici ekspresyon taraması varsa, 40'a kadar pozitif larva beklenebilir.
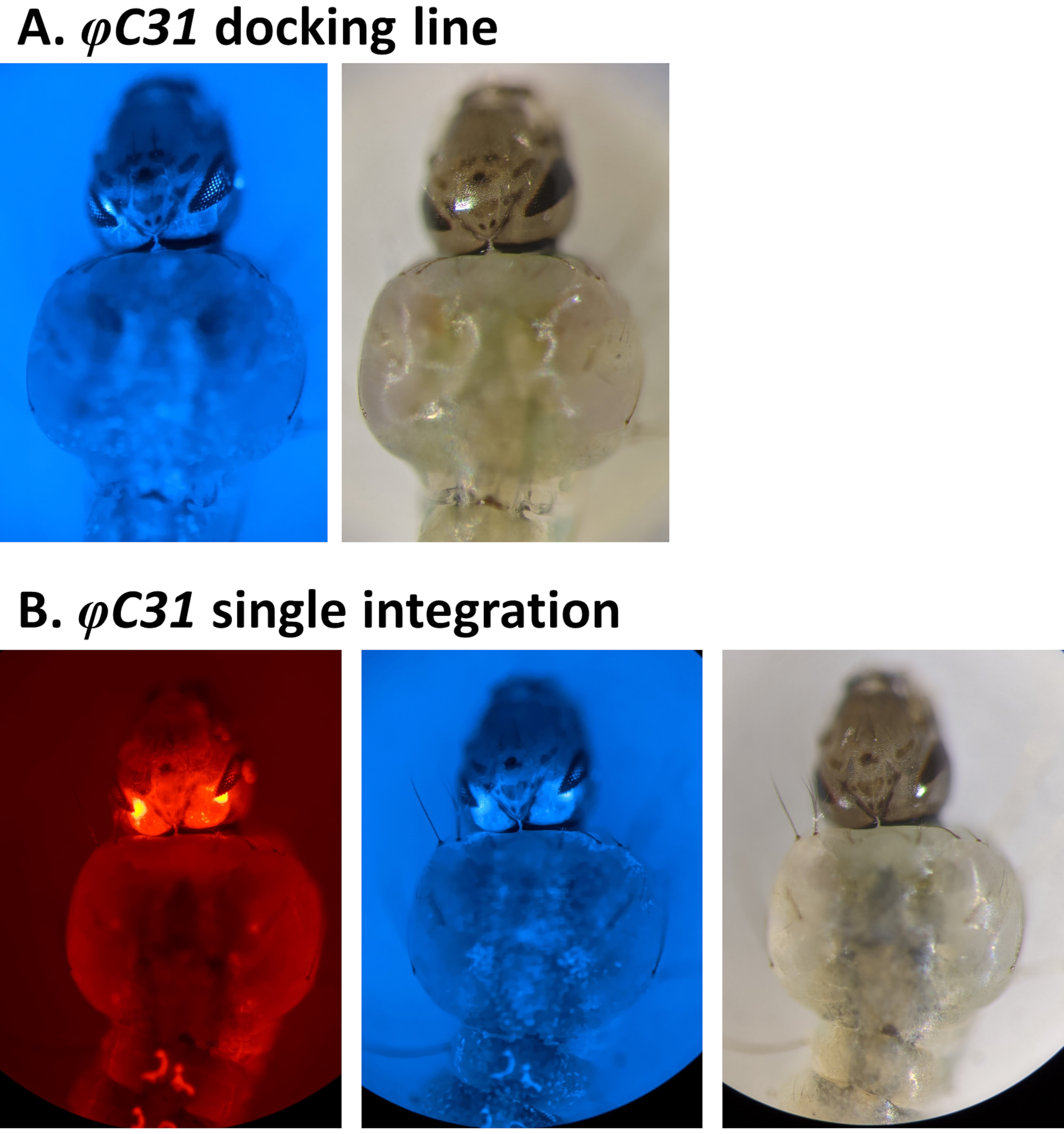
3xP3 promotörü tarafından düzenlenen floresan belirteçlerin taranarak dönüşümün fenotipik doğrulanması örnekleri Şekil 4 veFigure 5'te bildirilmiştir. Şekil 4, DsRed işaretli bir kasetin CFP (80.9, Tablo 1) ile işaretlenmiş bir yerleştirme hattına yerleştirilmesiyle elde edilen yeni bir An. stephensi çizgisini gösterir ve bu da gözlerde tespit edilen kırmızı ve mavi floresan tarafından belirtilen her iki işareti de ifade eden G1 soyunu gösterir.
Bunun yerine RMCE tasarımlarının, başlangıçta yerleştirme hattına yerleştirilen işaretleyicinin donör plazmid ile değiştirilmesine neden olması beklenir. Şekil 5A veFigure B, bu işaretleyici değişimini CFP (A11, Tablo 1) ile işaretlenmiş bir An. gambiae yerleştirme hattında göstermektedir, burada başarılı RMCE'den sonra CFP işaretçisi kaybolur ve YFP işaretçisi sarı (ancak mavi değil) göz ve sinir kordonu floresan33 ile sonuçlanır. Bazen, RMCE, hem orijinal CFP hem de yeni YFP işaretleyicileri ile işaretlenmiş bir larvanın gösterildiği Şekil 5C'de gösterildiği gibi istenen transgenik kasetin değişimi yerine tek bir entegrasyon olayıyla sonuçlanabilir. Toplam dönüşüm olayı sayısının %50'ye kadarının tek entegrasyon olduğu bildirilmektedir9,33.
Floresan bir işaretleyicinin varlığını tadırken, sinyalini olası arka plan otofluoresansından ayırt etmek çok önemlidir. Bu, CFP'yi Anopheles larvaları doğal mavi otofluoresans gösterirken özellikle önemlidir (Şekil 6A). Büyütmeyi artırmak ve floresanın organizatör tarafından yönlendirilmesi beklenen doku ve organlara odaklanmak, 3xP3-CFP işaretleyicisini kullanarak Şekil 6B'de gösterildiği gibi gerçek CFP pozitif bireyleri tanımlamak için gereklidir.
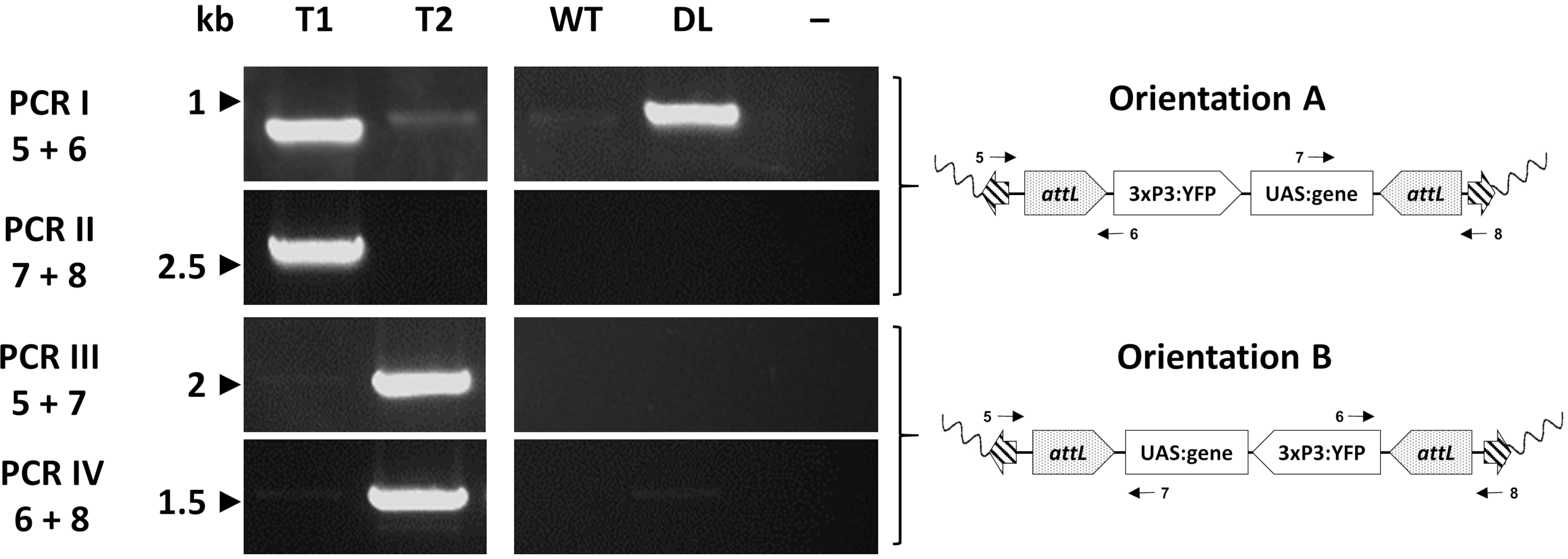
Bireysel transformantlar sonunda beklenen ekleme alanını doğrulamak için PCR aracılığıyla moleküler olarak değerlendirilir. Şekil 7 , sivrisinek genomuna eklemenin iki potansiyel yönelimini gösteren bir değişim An. gambiae hattından bireylerde PCR doğrulamasını rapor eder33.

Şekil 4. An. stephensi larvalarında φC31 tek entegrasyonunun doğrulanması (dorsal görünüm). A) Yerleştirme hattı (80.9, Tablo 1), 3xP3 organizatörünün düzenlemesi altında gözlerdeki CFP'yi ifade eder. B) Başarılı entegrasyon, yeni alınan DsRed'in yanı sıra gözlerdeki orijinal CFP işaretleyicisinin ifadesinde sonuçlanır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 5. An. gambiae larvalarında φC31 RMCE doğrulaması (ventral görünüm). A) Yerleştirme hattı (A10, Tablo 1), gözlerdeki (e) ve sinir kordonu (nc)5'teki 3xP3 promotörünün düzenlenmesi kapsamında CFP'yi ifade eder. B) Başarılı RMCE, floresan işaretleyicinin CFP'den YFP33'e takası ile sonuçlanır. C) Dönüştürücü larvaların hem CFP hem de YFP işaretleyicilerini ifade ettiği RMCE deneyi sırasında tek entegrasyon olayı meydana geldi. Bu larva, özellikle karın kaslarında (am) güçlü olan geniş bir YFP ekspresyon kalıbına neden olan GAL4 / UAS bileşenlerini taşır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 6. An. gambiae larvalarında CFP otofluoresans (dorsal görünüm). A) CFP filtresini kullanan pozitif (CFP+) ve negatif (CFP-) L4 larvasının yan yana görüntüsü. B) CfP+ vs CFP-birey ortaya çıkan larva gözlerinin yakın çekim görüntüsü. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 7. φC31 RMCE tarafından oluşturulan temsili transgenik An. gambiyada kaset ekleme yönünün moleküler doğrulaması. Transgenik kaset, ekleme bölgesine göre iki alternatif yönden (A veya B) birine eklenebilir. Her PCR reaksiyonu (I - IV), şematik plazmid haritalarında belirtildiği gibi her oryantasyon için bir tanısal amplifikasyon parçası vermek üzere tasarlanmış astarların (5-8)33 kombinasyonunu kullanır. T1: A ekleme oryantasyonunu taşıyan temsili transgenik birey; T2: B ekleme oryantasyonunu temsili transgenik birey; WT: vahşi tip; DL: yerleştirme hattı; -: suyun şablon olarak kullanıldığı reaksiyon negatif kontrolü. Bu rakam Adolfi ve ark. (2019)33'ten değiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
| Tür | Soy | Ad | attP'ler | Kromo-some | Organizatör işaretçisi | Menşe kurumu | Referans |
| An. stephensi | Indiana | 26.10b | Tek | 2R | 3xP3-eCFP | Kaliforniya Irvine'in Univ. | 25 |
| An. stephensi | Indiana | 44Cb | Tek | X | 3xP3-eCFP | Kaliforniya Irvine'in Univ. | 23, 24 |
| An. stephensi | Indiana | 80,9b | Tek | 2L | 3xP3-eCFP | Kaliforniya Irvine'in Univ. | Bu çalışma |
| An. gambiae | G3 | 113 | Tek | 2R | 3xP3-eCFP | Kaliforniya Irvine'in Univ. | Bu çalışma |
| An. gambiae | KIL | Ec | Tek | 3R | 3xP3-eCFP | Keele Üniv. | 19, 43 |
| An. gambiae | G3 | X1 | Tek | 2L | İşaretçi yok | Strazburg Üniversitesi | 22 |
| An. gambiae | G3 | YAttP | Tek | Y | 3xP3-RFP | Imperial College Londra | 21 |
| An. gambiae | G3 | A10b | Çift | 2R | 3xP3-eCFP | Liverpool Okulu Trop. Med. | 5 |
| An. gambiae | G3 | A11b | Çift | 2R | 3xP3-eCFP | Liverpool Okulu Trop. Med. | 9 |
| a. Johns Hopkins Üniversitesi'nden (M. Jacobs-Lorena'nın hediyesi) ve Kaliforniya Irvine'in Univ.'inde kültürde >20 yıl boyunca suş. | |||||||
| b. Bu satırlar makul talep üzerine yazarlardan temin edilebilir. | |||||||
| c. Bu hat, MRA-1163 olarak www.beiresources.org BEI deposunda mevcuttur. | |||||||
Tablo 1. Anopheles attP yerleştirme hatları.
Tartışmalar
Tercih edilen yerleştirme hattıyla uyumlu attB etiketli plazmidlerin doğru tasarımı, deneyin başarısı için çok önemlidir. Floresan rengi ve yerleştirme hattında zaten mevcut olan desene tabi olacak ifade deseni de dahil olmak üzere dönüştürücülerin taranması için kullanılan işaretleyicinin seçimine dikkat edilmelidir. Kolayca ayırt edilebilen floresan belirteçler kullanmak gerekir: iyi işaret kombinasyonları RFP (kırmızı)/CFP (siyan), RFP (kırmızı)/GFP (yeşil), RFP (kırmızı)/'yı içerir YFP (sarı) ve YFP (sarı)/CFP (siyan), kaçınılması gereken kombinasyonlar ise YFP (sarı)/GFP (yeşil) ve CFP (siyan)/GFP (yeşil). Gözlere ve sinir kordonuna özgü 3xP3 promotör39, sivrisinek transgenez için floresan belirteçlerin ekspresyonunun yönlendirilmede en sık kullanılandır. Gerçekten de, şu anda mevcut olan tüm Anopheles yerleştirme hatları bu organizatörü kullanır. Alternatif düzenleyici bölgeler, An. gambiae polyubiquitin geni (PUBc)5 veya birden fazla dokuda ifadeyi yönlendiren viral promotör IE120'dir. 3xP3 ile birlikte kullanıldığında, bu organizatörler olası renk kombinasyonlarını ve hatta aynı floroforun kullanımını genişletir. Belirtilen organizatörler, sivrisinek yaşam döngüsü boyunca aktiftir ve tüm yaşam aşamalarında tarama ve floresan izleme sağlar. Plazmid tasarımı sırasında ek bir husus, entegre edilecek veya değiştirilecek kargonun boyutudur. φC31 sistemi dikkat çekici taşıma kapasitelerine sahipken18, donör plazmid boyutunun genellikle dönüşüm verimliliği ile negatif korelasyona sahip olduğu düşünülmelidir22.
Açıklanan protokolde integraz kaynağı, enzimi her yerde ifade eden bir yardımcı plazmiddir40. Mikroenjeksiyonlar mikromline'ın oluştuğu bölgeye tam olarak yönlendirilmezse, bütünlüğün her yerde bulunması somatik hücrelerin dönüşümüne yol açabilir. Bu tür dönüşüm olayları kalıtsal olmadıkları için kaybolacak olsa da, somatik etkiler enjekte edilen bireylerin zindeliğini azaltabilir. Bunu önlemek ve dönüşüm verimliliğini artırmak için, örneğin vasa promotörü22,26 kullanılarak, integrase ifadesi mikrop çizgisiyle sınırlanabilir. Diğer protokoller, in vitro transkriptli haberci RNA'nın (mRNA) kullanımını φC31 integrase19,24,43 kaynağı olarak tanımlar. Bununla birlikte, bu mRNA'nın zahmetli bir şekilde hazırlanmasını içerir ve bozulmayı önlemek için enjeksiyon karışımının dikkatli bir şekilde işlenmesini ve RNaz serbest reaktiflerin kullanılmasını gerektirir. Plazmid integrase kaynakları hem An. gambiae9,21,22,26,33,37 hem de An. stephensi (A.A. kişisel iletişim) güvenilir ve verimli dönüşüme yol açarak gösterilmiştir ve bu nedenle tercih edilen seçeneğimizdir. Bütünleştirici teslimat için bir diğer seçenek de kendinden yerleştirmeli yardımcı hatlarda in vivo üretimidir. Bu tür çizgiler, Germline özgü promotör nanolarının düzenlenmesi altında φC31 bütünlüğünü ifade eden An. gambiae'de oluşturulmuş ve daha iyi bir hayatta kalma ve dönüşüm verimliliğine yol açtıkları bulunmuştur20. Bununla birlikte, integraz enziminin yardımcı hatta in vivo üretimi tarafından dayatılan potansiyel fitness yükleri göz önünde bulundurulmalıdır.
Diğer transgenik tekniklerde olduğu gibi, dönüştürücüleri kurtarma şansını en üst düzeye çıkarmak için enjekte edilen embriyolardan türeyen bireylerin yetiştirilmesine ve geçişine özel bakım ayrılmalıdır. Transjeni sapan miras alan bireyler öncelikle G1 soyunda geri kazanılabilir. Bununla birlikte, potansiyel dönüşümün erken belirtileri, 3xP3 promotörü43 kullanırken G0 birinci ve ikinci instar larvalarının anal papilla ve/veya sinir kordonunda floresan belirtecin geçici epizomal ekspresyonunun varlığı ile değerlendirilebilir. Geçici floresan varlığı başarılı plazmid doğumu düşündürmekle birlikte, kalıtsal germline dönüşümünü garanti etmez. Benzer şekilde, geçici ifade eksikliği başarılı dönüşümü dışlamaz. Bununla birlikte, geçici olarak pozitif bireylerin geçici olarak negatif olanlara kıyasla transgenik soy verme olasılığının daha yüksek olduğu görülmüştür43,48. Uzman ellerde, sadece pozitif bireylerin yetiştirilmesi ve geçilmesi sivrisinek sayılarını azaltmak için bir seçenek olabilir. Bununla birlikte, küçük G0 larvalarının önemi ve kırılganlığı göz önüne alındığında, en az manipülasyon hala tavsiye edilir ve tüm G0 bireylerinin yetiştirülmeleri her zaman tavsiye edilir.
Bu protokolde bildirilen çiftleşme şeması, çiftleşme şansını en üst düzeye çıkarmak ve bağımsız dönüşüm olaylarını yalıtmak için tasarlanmıştır. Bununla birlikte, böcek alanı veya personel mevcudiyeti bir sorunsa, yeterli karşı cins bireyler sağlanırsa G0 yetişkinleri tek kafeslerde seks tarafından birikebilir. Böyle bir kurulum, aynı kafesten bireylerde meydana gelen birden fazla dönüşüm olayı arasında ayrımcılığa izin vermez. Deneysel kuruluma bağlı olarak, tarama işlemi sırasında çift (tek entegrasyon) veya tek (RMCE) işaretleyicinin bulunması beklenir. Tek tümleştirme denemelerinde, orijinal işaretçinin varlığını yerleştirme satırından doğrulamak önemlidir, RMCE'de ise daha önce tümleşik işaretleyicinin kaybını doğrulamak önemlidir. Gerçekten de, RMCE tasarımlarında, tek bir attP sitesinin yeniden birleştirilmesi nedeniyle değişim yerine tek bir entegrasyonun gerçekleştiği dönüştürücüleri kurtarmak nadir değildir9,33. Bu tür bireylerde her iki floresan belirteç de mevcuttur ve tüm donör plazmid omurgası, her iki floresan belirteç için kapsamlı bir tarama yapılmasının önemini vurgulamaktadır.
Beklenen floresan desenlerinin varlığı başarılı bir dönüşüme işaret ederken, ekleme bölgesinin moleküler karakterizasyonu yapılmalıdır. Bunu yapmak için, kenetlenme hattının yan genomik bölgeleri de dahil olmak üzere tahmin edilen ekleme locusunun doğru haritalarının hazırlanması, gen amplifikasyon analizleri için yeterli tanısal oligonükleotid astarların tasarımı için çok önemlidir. Tek entegrasyon olayları, yeni entegre edilmiş DNA ile daha önce yerleştirilmiş kaset arasındaki kavşakta attR ve attL hibrid sahalarının oluşmasına neden oldu. Bu siteler ekleme sitesi doğrulaması için hedeflenebilir. RMCE tasarımlarında, donör kasetin yerleştirilmesi genomik lokus ile ilgili olarak iki alternatif yönde gerçekleşebilir, böylece hattın hangi oryantasyonu taşıdığını tespit etmek için alternatif PCR kombinasyonlarında dört astar kullanılabilir. Kaset eklemenin yönü transgene ekspresyonunu etkileyebileceğinden, karşılaştırmalı gen ekspresyon analizinde aynı ekleme yönünü taşıyan çizgilerin kullanılması önemlidir.
Düşük sayıda dönüştürücü ile çalışırken, tüm bireyleri moleküler analiz için feda etmek isndenmeyebilir. Bunun bir seçeneği, bacak kaybı yetişkin bir kadın çiftleşme yeteneğini etkilemediği ve oviposit49'a etkilemediği için tek yetişkin bacaklarından çıkarılan DNA üzerinde moleküler analiz yapmaktır46. Bununla birlikte, bacak çıkarma sürecinde bireye zarar verme riski vardır. Başarı, atılan pupal vakalar (L. Grigoraki kişisel iletişim) kullanılarak elde edilmiştir, ancak en güvenli yaklaşım, uygulanabilir G3 soyu elde ettikten sonra G2 ebeveynleri üzerinde moleküler analiz yapmaktır.
Son yıllarda CRISPR/Cas9, bölgeye özgü genom düzenleme yönteminde devrim yaratmada devrim yaptı26,41,50,51. Raya yönelik RMCE'nin aksine, CRISPR/Cas9 aracılı gen entegrasyonları (knock-in'ler), yalnızca tek adımlı bir dönüşüm olayına ihtiyaç duyulan önceden eklenmiş yeniden birleştirme alanlarının varlığından bağımsızdır. Bununla birlikte, CRISPR /Cas9 sistemi, başarılı homoloji yönlendirilmiş onarım için istenen ekleme bölgesini kuşatır ve rehber RNA'ların aracılık ettiği verimli saha tanımada bilinen büyük genomik dizilerin varlığına dayanır. Bu koşullar her zaman karşılanamaz veya sorun gidermek için zahmetli olabilir ve An. gambiae ve An. stephensi'deki birden fazla yerleştirme hattının ve bunlardan türetilen çizgilerin mevcudiyeti göz önüne alındığında, φC31 sistemi aynı genomik konumlardaki transgenler arasında doğrudan fenotipik karşılaştırmalar yapmak için çok değerli bir araç olmaya devam etmektedir.
Açıklamalar
Yazarların açıklayacak bir şeyi yok.
Teşekkürler
Transgenik An. stephensi larvalarının görüntülerini sağladığı için Kiona Parker'a (UCI) ve transgenik An. gambiae larvalarını sağladığı için Fraser Colman (LSTM) ve Beth Poulton'a (LSTM) minnettarız. Beth Poulton (LSTM) ayrıca An. gambiae larvalarının görüntülenmesi sırasında değerli yardım sağladı. Bu çalışma Tata Genetik ve Toplum Enstitüsü (TIGS) ve LSTM'nin A.A.'ya (DCF2014AA) verilen Direktör Katalizör Fonu tarafından finanse edildi. A.A.J. Kaliforniya Üniversitesi, Irvine'de Donald Bren profesörüdür.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 1.5 mL eppendorf tubes | |||
| 8-well microslides | VWR | MARI1216690 | |
| DNeasy Blood & Tissue Kit | Qiagen | 69504 | |
| EndoFree Plasmid Maxi Kit (10) | Qiagen | 12362 | |
| Ethanol, Absolute, Molecular Biology Grade | |||
| Filter set CFP for Leica MZ FLIII Excitation 436/20 nm, extinction 480/40 nm | Leica | 10446363 | |
| Filter set dsRED for Leica MZ FLIII Excitation 545/30 nm, extinction 620/60 nm | Leica | 10447079 | |
| Filter set YFP customised for Leica MZ FLIII | Omega Optical | 500QM25, 500QM35 | |
| Halocarbon oil 27 | Sigma | H8773 | |
| Halocarbon oil 700 | Sigma | H8898 | |
| Petri dishes | |||
| Potassium chloride | |||
| Sodium Chloride | |||
| Sodium phosphate dibasic | |||
| Sodium Acetate Solution (3 M), pH 5.2 | Thermo Fisher Scientific (Life Technologies) | R1181 | |
| Stable brush Size 0 |
Referanslar
- Adolfi, A., Lycett, G. J. Opening the toolkit for genetic analysis and control of Anopheles mosquito vectors. Current Opinion in Insect Science. 30, 8-18 (2018).
- Grossman, G. L., Rafferty, C. S., Clayton, J. R., Stevens, T. K., Mukabayire, O., Benedict, M. Q. Germline transformation of the malaria vector, Anopheles gambiae, with the piggyBac transposable element. Insect Molecular Biology. 10 (6), 597-604 (2001).
- Nolan, T., Bower, T. M., Brown, A. E., Crisanti, A., Catteruccia, F. piggyBac-mediated germline transformation of the malaria mosquito Anopheles stephensi using the red fluorescent protein dsRED as a selectable marker. Journal of Biological Chemistry. 277 (11), 8759-8762 (2002).
- Perera, O. P., Harrell, R. A., Handler, A. M. Germ-line transformation of the South American malaria vector, Anopheles albimanus, with a piggyBac/EGFP transposon vector is routine and highly efficient. Insect Molecular Biology. 11 (4), 291-297 (2002).
- Adolfi, A., Pondeville, E., Lynd, A., Bourgouin, C., Lycett, G. J. Multi-tissue GAL4-mediated gene expression in all Anopheles gambiae life stages using an endogenous polyubiquitin promoter. Insect Biochemistry and Molecular Biology. 96, 1-9 (2018).
- Carballar-Lejarazú, R., Jasinskiene, N., James, A. Exogenous gypsy insulator sequences modulate transgene expression in the malaria vector mosquito, Anopheles stephensi. Proceedings of the National Academy of Sciences of the United States of America. 110 (18), 7176-7181 (2013).
- Galizi, R., et al. A synthetic sex ratio distortion system for the control of the human malaria mosquito. Nature Communications. 5, 3977(2014).
- Nolan, T., Petris, E., Müller, H. M., Cronin, A., Catteruccia, F., Crisanti, A. Analysis of two novel midgut-specific promoters driving transgene expression in Anopheles stephensi mosquitoes. PLoS ONE. 6 (2), 16471(2011).
- Lynd, A., Balabanidou, V., Vontas, J., Lycett, G. J. Development of a functional genetic tool for Anopheles gambiae oenocyte characterisation: appliction to cuticular hydrocarbon synthesis. BioRxiv. , (2019).
- O'Brochta, D. A., Alford, R. T., Pilitt, K. L., Aluvihare, C. U., Harrell, R. A., Harrell, R. A. piggyBac transposon remobilization and enhancer detection in Anopheles mosquitoes. Proceedings of the National Academy of Sciences of the United States of America. 108 (39), 16339-16344 (2011).
- O'Brochta, D. A., Pilitt, K. L., Harrell, R. A., Aluvihare, C., Alford, R. T. Gal4-based enhancer-trapping in the malaria mosquito Anopheles stephensi. G3. 2 (11), Bethesda. 1305-1315 (2012).
- Macias, V. M., et al. nanos-Driven expression of piggyBac transposase induces mobilization of a synthetic autonomous transposon in the malaria vector mosquito, Anopheles stephensi. Insect Biochemistry and Molecular Biology. 87, 81-89 (2017).
- Nimmo, D. D., Alphey, L., Meredith, J. M., Eggleston, P. High efficiency site-specific genetic engineering of the mosquito genome. Insect Molecular Biology. 15 (2), 129-136 (2006).
- Kim, A., Pyykko, I. Size matters: Versatile use of PiggyBac transposons as a genetic manipulation tool. Molecular and Cellular Biochemistry. 354, 301-309 (2011).
- Thorpe, H. M., Smith, M. C. M. In vitro site-specific integration of bacteriophage DNA catalyzed by a recombinase of the resolvase/invertase family. Proceedings of the National Academy of Sciences of the United States of America. 95 (10), 5505-5510 (1998).
- Khaleel, T., Younger, E., Mcewan, A. R., Varghese, A. S., Smith, M. C. M. A phage protein that binds φC31 integrase to switch its directionality. Molecular Microbiology. 80 (6), 1450-1463 (2011).
- Farruggio, A. P., Chavez, C. L., Mikell, C. L., Calos, M. P. Efficient reversal of phiC31 integrase recombination in mammalian cells. Biotechnology Journal. 7 (11), 1332-1336 (2012).
- Venken, K. J. T., He, Y., Hoskins, R. A., Bellen, H. J. P[acman]: A BAC Transgenic Platform for Targeted Insertion of Large DNA Fragments in D. melanogaster. Science. 314 (5806), 1747-1751 (2006).
- Meredith, J. M., et al. Site-specific integration and expression of an anti-malarial gene in transgenic Anopheles gambiae significantly reduces Plasmodium infections. PLoS ONE. 6 (1), 14587(2011).
- Meredith, J. M., Underhill, A., McArthur, C. C., Eggleston, P. Next-Generation Site-Directed Transgenesis in the Malaria Vector Mosquito Anopheles gambiae: Self-Docking Strains Expressing Germline-Specific phiC31 Integrase. PLoS ONE. 8 (3), 59264(2013).
- Bernardini, F., et al. Site-specific genetic engineering of the Anopheles gambiae Y chromosome. Proceedings of the National Academy of Sciences of the United States of America. 111 (21), 7600-7605 (2014).
- Volohonsky, G., et al. Tools for Anopheles gambiae Transgenesis. G3. 5 (6), Bethesda. 1151-1163 (2015).
- Amenya, D. A., et al. Comparative fitness assessment of Anopheles stephensi transgenic lines receptive to site-specific integration. Insect Molecular Biology. 19 (2), 263-269 (2010).
- Isaacs, A. T., et al. Transgenic Anopheles stephensi coexpressing single-chain antibodies resist Plasmodium falciparum development. Proceedings of the National Academy of Sciences of the United States of America. 109 (28), 1922-1930 (2012).
- Pham, T. B., et al. Experimental population modification of the malaria vector mosquito, Anopheles stephensi. PLoS Genetics. 15 (12), 1008440(2019).
- Hammond, A., et al. A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nature Biotechnology. 34 (1), 78-83 (2015).
- Groth, A. C., Fish, M., Nusse, R., Calos, M. P. Construction of Transgenic Drosophila by Using the Site-Specific Integrase from Phage phiC31. Genetics. 166 (4), 1775-1782 (2004).
- Franz, A. W. E., et al. Comparison of transgene expression in Aedes aegypti generated by mariner Mos1 transposition and site-directed recombination. Insect Molecular Biology. 20 (5), 587-598 (2011).
- Labbé, G., Nimmo, D., Alphey, L. piggybac-and PhiC31-Mediated Genetic Transformation of the Asian Tiger Mosquito, Aedes albopictus (Skuse). PLoS Neglected Tropical Diseases. 4 (8), 788(2010).
- Schetelig, M. F., Scolaric, F., Handler, A. M., Kittelmann, S., Gasperi, G., Wimmer, E. A. Site-specific recombination for the modification of transgenic strains of the Mediterranean fruit fly Ceratitis capitata. Proceedings of the National Academy of Sciences of the United States of America. 106 (43), 18171-18176 (2009).
- Yonemura, N., et al. phiC31-integrase-mediated, site-specific integration of transgenes in the silkworm, Bombyx mori (Lepidoptera: Bombycidae). Applied Entomology and Zoology. 43 (11), 997-1008 (2013).
- Bateman, J. R., Lee, A. M., Wu, C. T. Site-specific transformation of Drosophila via phiC31 integrase-mediated cassette exchange. Genetics. 173 (2), 769-777 (2006).
- Adolfi, A., Poulton, B., Anthousi, A., Macilwee, S., Ranson, H., Lycett, G. J. Functional genetic validation of key genes conferring insecticide resistance in the major African malaria vector, Anopheles gambiae. Proceedings of the National Academy of Sciences of the United States of America. 116 (51), 25764-25772 (2019).
- Haghighat-Khah, R. E., et al. Site-specific cassette exchange systems in the aedes aegypti mosquito and the Plutella xylostella moth. PLoS ONE. 10 (4), 0121097(2015).
- Long, D., Lu, W., Zhang, Y., Bi, L., Xiang, Z., Zhao, A. An efficient strategy for producing a stable, replaceable, highly efficient transgene expression system in silkworm, Bombyx mori. Scientific Reports. 5 (1), 8802(2015).
- Volohonsky, G., et al. Transgenic Expression of the Anti-parasitic Factor TEP1 in the Malaria Mosquito Anopheles gambiae. PLoS Pathogens. 13 (1), 1006113(2017).
- Grigoraki, L., Grau-Bové, X., Yates, H. C., Lycett, G. J., Ranson, H. Isolation and transcriptomic analysis of anopheles gambiae oenocytes enables the delineation of hydrocarbon biosynthesis. eLife. 9, 58019(2020).
- Kyrou, K., et al. A CRISPR-Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nature Biotechnology. 36 (11), 1062-1066 (2018).
- Berghammer, A. J., Klingler, M., Wimmer, E. A. A universal marker for transgenic insects. Nature. 402 (6760), 370-371 (1999).
- Ringrose, L. Transgenesis in Drosophila melanogaster. Methods in Molecular Biology. 561, 3-19 (2009).
- Dong, Y., Simões, M. L., Marois, E., Dimopoulos, G. CRISPR/Cas9 -mediated gene knockout of Anopheles gambiae FREP1 suppresses malaria parasite infection. PLoS Pathogens. 14 (3), 1006898(2018).
- Lombardo, F., Lycett, G. J., Lanfrancotti, A., Coluzzi, M., Arcà, B. Analysis of apyrase 5' upstream region validates improved Anopheles gambiae transformation technique. BMC research notes. 2, 24(2009).
- Pondeville, E., et al. Efficient ΦC31 integrase-mediated site-specific germline transformation of Anopheles gambiae. Nature Protocols. 9 (7), 1698-1712 (2014).
- Lobo, N. F., Clayton, J. R., Fraser, M. J., Kafatos, F. C., Collins, F. H. High efficiency germ-line transformation of mosquitoes. Nature protocols. 1 (3), 1312-1317 (2006).
- Terenius, O., Juhn, J., James, A. A. Injection of An. stephensi embryos to generate malaria-resistant mosquitoes. Journal of Visualized Experiments. 5, 216(2007).
- Lynd, A., et al. Insecticide resistance in Anopheles gambiae from the northern Democratic Republic of Congo, with extreme knockdown resistance (kdr) mutation frequencies revealed by a new diagnostic assay. Malaria Journal. 17 (1), 412(2018).
- Marinotti, O., et al. Development of a population suppression strain of the human malaria vector mosquito, Anopheles stephensi. Malaria Journal. 12 (1), 142(2013).
- Adolfi, A. In vivo functional genetic analysis of insecticide resistance in the malaria mosquito Anopheles gambiae. University of Liverpool. , PhD thesis (2017).
- Isaacs, A. T., Lynd, A., Donnelly, M. J. Insecticide-induced leg loss does not eliminate biting and reproduction in Anopheles gambiae mosquitoes. Scientific Reports. 7, 46674(2017).
- Gantz, V. M., et al. Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. Proceedings of the National Academy of Sciences of the United States of America. 112 (49), 6736-6743 (2015).
- Li, M., Akbari, O. S., White, B. J. Highly efficient site-specific mutagenesis in malaria mosquitoes using CRISPR. G3: Genes, Genomes, Genetics. 8 (2), 653-658 (2018).
- Poulton, B. C., et al. Using the GAL4-UAS System for Functional Genetics in Anopheles gambiae. J. Vis. Exp. , (2021).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır