
Method Article
Ориентированная на сайт φC31-опосредованная интеграция и кассетный обмен в переносчиках малярии Anopheles
В этой статье
Резюме
Протокол описывает, как достичь направленных на сайт модификаций в геноме малярийных комаров Anopheles с использованием системы φC31 . Описанные модификации включают как интеграцию, так и обмен трансгенными кассетами в геноме attP-несущих стыковочных линий.
Аннотация
Функциональный геномный анализ и связанные с ним стратегии генетического контроля малярии основаны на проверенных и воспроизводимых методах для точной модификации генома комаров Anopheles. Среди этих методов система φC31 обеспечивает точную и стабильную интеграцию трансгенов, направленную на сайт, или замену интегрированных трансгенных кассет с помощью опосредованного рекомбиназой кассетного обмена (RMCE). Этот метод опирается на действие бактериофагов Streptomyces φC31 integrase для катализа рекомбинации между двумя специфическими сайтами присоединения, обозначенными attP (полученным из фага) и attB (полученным из бактерии-хозяина). Система использует один или два участка attP, которые были ранее интегрированы в геном комара, и сайт (сайты) attB в ДНК донорского шаблона. Здесь мы иллюстрируем, как стабильно модифицировать геном attP-несущих линий стыковки Anopheles с использованием двух плазмид: донора с меткой attB, несущего шаблон интеграции или обмена, и вспомогательной плазмиды, кодирующей интегразу φC31. Мы сообщаем о двух репрезентативных результатах модификации, направленной на сайт φC31: единая интеграция трансгенной кассеты у комаров An. stephensi и RMCE у комаров An. gambiae. φC31-опосредованная манипуляция геномом предлагает преимущество воспроизводимой экспрессии трансгенов из проверенных, нейтральных к приспособленности геномных сайтов, что позволяет проводить сравнительный качественный и количественный анализ фенотипов. Ориентированный на участок характер интеграции также существенно упрощает валидацию единого места вставки и схемы сопряжения для получения стабильной трансгенной линии. Эти и другие характеристики делают систему φC31 важным компонентом генетического инструментария для трансгенного манипулирования малярийными комарами и другими насекомыми-переносчиками.
Введение
Способность надежно и воспроизводимо модифицировать геном комаров-переносчиков болезней укрепила функциональную валидацию генов in vivo и открыла двери для реализуемых стратегий борьбы с генетическими переносчиками, таких как стратегии борьбы с комарами Anopheles, передающими малярию1.
Раннее редактирование генома комара основывалось исключительно на транспонируемой трансформации, опосредованной элементами (TE), причем piggyBac является наиболее часто используемым транспозоном в Anopheles2,3,4. Однако случайный характер интеграции ТЭ может привести к нежелательным модификациям, таким как нокауты генов (вставочный мутагенез) и значительное влияние позиции на экспрессию трансгенов5,6,7,8. Множественные вставки также являются распространенным явлением при использовании piggyBac5,9, что делает валидацию и изоляцию трансгенных линий с одиночными вставками трудоемкими. Другие недостатки включают их потенциальную ремобилизацию, наблюдаемую в зародышевой линии Anopheles stephensi при обеспечении источника транспозазы piggyBac10,11,12, и их ограниченный размер груза ДНК (10-15 кб в длину) с уменьшением эффективности трансформации с увеличением размера донорской плазмиды13,14.
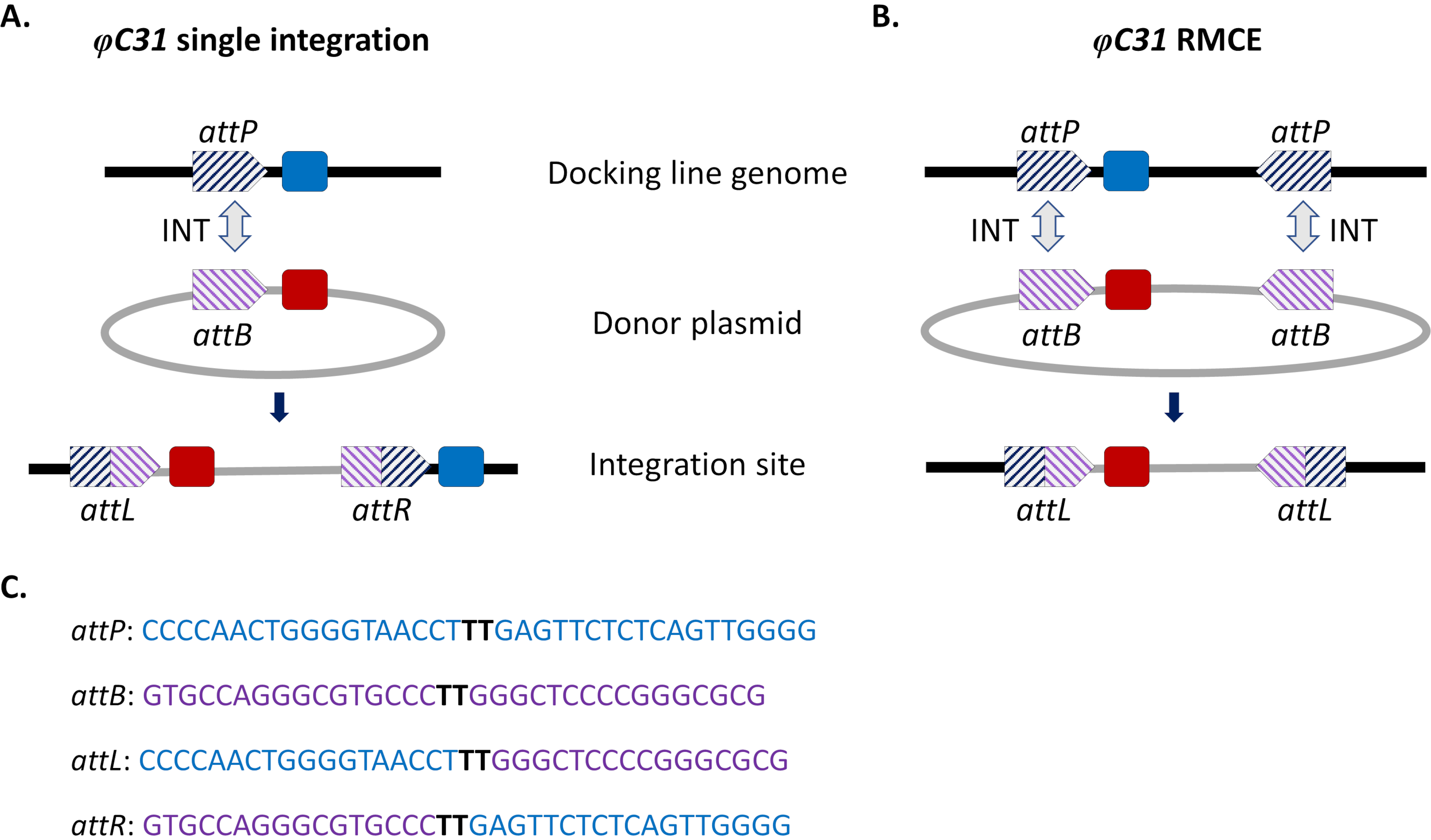
Для обхода этих проблем были внедрены подходы к интеграции, ориентированные на сайты. Наиболее распространенной сайт-направленной модификацией генома у комаров является модификация, опосредованная системой φC31 (рисунок 1a). Это обусловлено вирусной интегразой, которая катализирует рекомбинацию между двумя гетероспецифическими участками присоединения (att), встречающимися естественным образом в геноме бактериофага φC31 (attP) и в хозяине бактерии Streptomyces (attB)15. Рекомбинация двух участков является однонаправленной и приводит к образованию гибридных сайтов (attL и attR). Рекомбинация таких гибридных участков (приводящая к иссечению ДНК) потребует не только наличия активной вирусной интегразы, но и другого фагового фактора рекомбинации16,17. Таким образом, создается стабильный сайт интеграции, который снимает проблему потенциальной нежелательной ремобилизации15. Кроме того, система позволяет интегрировать крупногабаритные грузы (например, в D. melanogaster18 сообщалось об интеграции конструкций >100 кб), что значительно увеличивает грузоподъемность. Интеграция происходит в одном предопределенном геномном локусе, что значительно упрощает валидацию вставки и схемы спаривания для получения стабильной трансгенной линии. Наконец, сайт-ориентированный характер интеграции позволяет нормализовать экспрессию, поскольку альтернативные трансгены расположены в одном локусе и, следовательно, регулируются в одном и том же соседнем геномном контексте. Действительно, одним из основных применений метода является прямое сравнение фенотипов, присвоенных различными трансгенами после введения в идентичный локус.
Достижение φC31-опосредованной интеграции включает в себя две фазы: фаза I - создание трансгенных стыковочных линий, несущих attP-сайт(ы), а фаза II - это направленная на сайт интеграция attB-флангового груза в геном стыковочной линии19. Создание стыковочных линий фазы I основывалось на TE-опосредованной случайной интеграции конструкций, помеченных attP, и, таким образом, включало начальный трудоемкий процесс (включая анализ южного пятна и обратный ПЦР на потомстве одной самки) для выделения и проверки трансгенных линий, несущих одно событие интеграции в уникальных, транскрипционно активных и нейтральных к фитнесу геномных местах. Тем не менее, несколько стыковочных линий для φC31-опосредованной одиночной интеграции были разработаны и проверены в An. gambiae19,20,21,22 и в An. stephensi23,24,25 (таблица 1). Каждая из этих линий варьируется с точки зрения геномного местоположения места стыковки и генетического фона, специфичного для штамма, и из них может быть создано большое разнообразие новых трансгенных линий. Сложная валидация ИНТЕГРАЦИЙ, опосредованных TE, для создания стыковочных линий теперь может быть обойдена технологией CRISPR/Cas926; однако это опирается на априорное знание нейтральных локусов, которые должны быть нацелены, и окружающих их последовательностей.
φC31-опосредованная интеграция широко применяется для редактирования генома насекомых от модельного организма D. melanogaster27, до комаров Aedes aegypti13,28, Ae. albopictus29, An. gambiae19 и An. stephensi24, а также других насекомых, включая Ceratitis capitata30 и Bombyx mori31.
Ограничением φC31-опосредованной интеграции, особенно с учетом потенциальных полевых высвобождений для борьбы с переносчиками, является интеграция в геном комара всей донорской плазмиды, несущей attB, включая нежелательные последовательности, такие как маркеры генов устойчивости к антибиотикам и компоненты позвоночника плазмиды бактериального происхождения. Для решения этой проблемы была реализована модификация стандартной системы, рекомбиназно-опосредованный кассетный обмен (RMCE), которая позволяет точно заменить ранее интегрированную трансгенную кассету новой донорской ДНК (рисунок 1b). Это достигается путем использования двух перевернутых атт-сайтов , фланкирующих донорские и реципиентные кассеты на каждом конце, что приводит к одновременному проведению двух независимых событий рекомбинации, что приводит к кассетному обмену без интеграции плазмидной магистрали. Эта улучшенная конструкция обходит интеграцию нежелательных последовательностей и расширяет применение систем φC31 , включая, например, интеграцию немаркированных грузов ДНК путем скрининга на потерю ранее интегрированного флуоресцентного маркера32.
RMCE был достигнут сначала с D. melanogaster32, а затем успешно применен к немодельным насекомым, включая An. gambiae9,26,33, Ae. aegypti34, Plutella xylostella34 и B. mori35. Несколько стыковочных линий для RMCE были разработаны и проверены в An. gambiae5,9,26 (таблица 1). Насколько нам известно, RMCE еще предстоит исследовать на других видах переносчиков Anopheles.
На сегодняшний день система φC31 широко используется у комаров Anopheles для введения и изучения различных молекул, включая противомалярийные эффекторы19,24,36, компоненты системы GAL4/UAS для сверхэкспрессии и нокдауна генов для исследований устойчивости к инсектицидам9,33, регуляторные элементы, гены-репортеры5,21,37 и элементы генного драйва26 ,38.
Этот протокол описывает, как выполнить 1) направленную на сайт интеграцию attB-флангового груза и 2) RMCE конструкции, окруженной перевернутыми участками attB , в геном стыковочных линий Anopheles . Это достигается с помощью двух плазмид: донорской плазмиды с меткой attB, несущей интересующий трансген, и вспомогательной плазмиды, экспрессирующей интегразу φC31 . Основные переносчики малярии An. gambiae и An. stephensi используются в качестве конкретных примеров, однако эти протоколы применимы к другим видам Anopheles .

Рисунок 1. Site-направленные модификации генома, единичная интеграция и рекомбиназно-опосредованный кассетный обмен (RMCE) с использованием системы φC31. Интеграза φC31 (INT, серая двойная стрелка) катализирует рекомбинацию между сайтом (сайтами) attB (фиолетовым полосатым), присутствующим в донорской плазмиде, и сайтом (сайтами) attP (синими полосатыми), присутствующими в приемной стыковочной линии, что приводит к образованию гибридных сайтов attL и attR. A) Интеграция достигается, когда одиночные участки attB и attP рекомбинируют и приводят к наличию двух интегрированных маркеров (синего и красного). B) RMCE возникает, когда два сайта attB/P рекомбинируют одновременно и приводит к замене кассеты между att-участками стыковочной линии (синий маркер) на сайт, переносимый плазмидой донора (красный маркер). C) Частичные нуклеотидные последовательности attP (синий) и attB (фиолетовый) и гибридные сайты attL/R. Рекомбинация происходит между последовательностями ядра «TT», выделенными жирным черным шрифтом. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
протокол
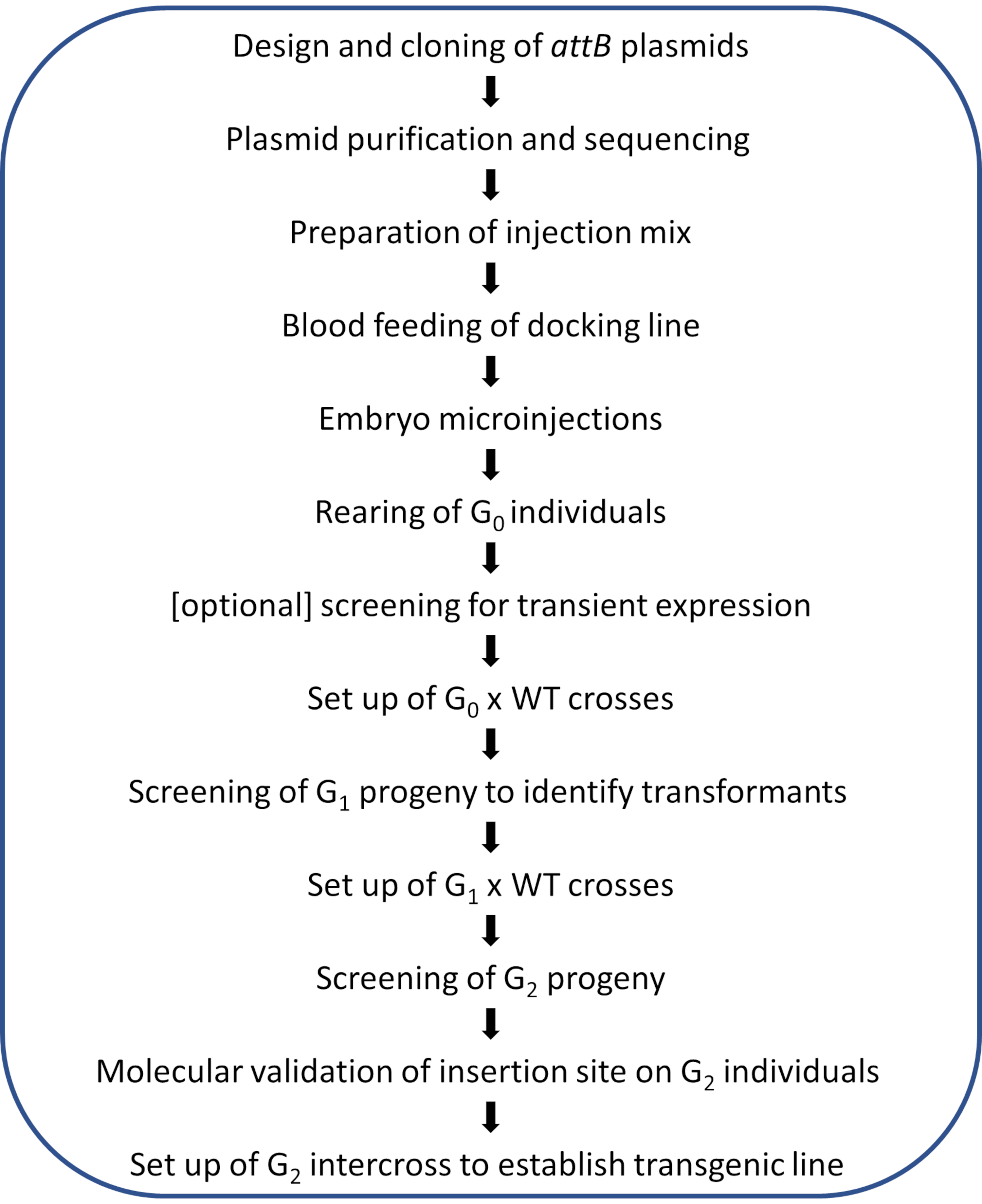
ПРИМЕЧАНИЕ: Схематический рабочий процесс проиллюстрированного протокола показан на рисунке 2.
1. Проектирование плазмид с метками φC31 attB (рисунок 3)
- Создание донорских плазмид attB, несущих следующие основные компоненты
- Доминирующий флуоресцентный маркер
- Выберите промоутер, чтобы управлять выражением флуоресцентного маркера.
ПРИМЕЧАНИЕ: Для трансгенеза Anopheles флуоресцентные маркеры обычно находятся под регуляцией промотора 3xP39, который управляет экспрессией в глазах и нервном канатике. Альтернативно, промотор PUBc5 может быть использован, когда желательна экспрессия в нескольких тканях. Донорские плазмиды и стыковочные линии, используемые в качестве примеров в этом протоколе, маркируются с помощью промотора 3xP3. - Выберите флуоресцентный белок (FP), совместимый с приемной стыковочной линией, чтобы они были легко различимы.
ПРИМЕЧАНИЕ: Не используйте тот же маркер, который уже присутствует в стыковочной линии, и избегайте одновременного использования GFP (зеленый) / YFP (желтый) и GFP (зеленый) / CFP (голубой), поскольку их очень трудно надежно дифференцировать. Донорские плазмиды, используемые в качестве примеров в этом протоколе, помечены либо DsRed, либо YFP, поскольку они должны быть интегрированы в стыковочную линию, помеченную CFP.
- Выберите промоутер, чтобы управлять выражением флуоресцентного маркера.
-
Сайт(ы) рекомбинации attB
- Используйте один сайт attB для интеграции трансгенной кассеты (конструкция с одной аттB ) (рисунок 3A).
- Используйте два инвертированных участка attB для RMCE (конструкция с двойным аттВ ), где участки лежат перевернутыми по отношению друг к другу, и заключайте шаблон донорской ДНК (рисунок 3B).
ПРИМЕЧАНИЕ: Ориентация площадки (точек) attB должна быть совместима с ориентацией площадки (точек) attP , присутствующей в стыковочной линии.
- Желаемый трансгенный груз
- Используйте любые другие желаемые функции, чтобы быть интегрированными в геном комара на основе конкретной цели эксперимента. Здесь мы описываем интеграцию противомалярийной эффекторной молекулы в геном An. stephensi и интеграцию компонентов системы GAL4/UAS в комаров An. gambiae .
- Плазмидные компоненты позвоночника
- Включают, среди других важных компонентов для репликации плазмид у бактерий, маркер отбора плазмид in vitro (т.е. ген устойчивости к антибиотикам).
ПРИМЕЧАНИЕ: Плазмидная основа будет интегрирована в геном комара в одноаттб-конструкции для интеграции (рисунок 3A), в то время как она не будет вставлена в конструкцию с двойным аттБ для RMCE (рисунок 3B).
- Включают, среди других важных компонентов для репликации плазмид у бактерий, маркер отбора плазмид in vitro (т.е. ген устойчивости к антибиотикам).
- Доминирующий флуоресцентный маркер
2. Подготовка плазмид для микроинъекционной смеси
ПРИМЕЧАНИЕ: Протокол, проиллюстрированный здесь, включает использование двух плазмид: донорской плазмиды с меткой attB, несущей интересующий трансген, и вспомогательной плазмиды, которая экспрессирует интегразу φC31 под регуляцией промотора Drosophila Hsp7040.
- Очистите донорские и вспомогательные плазмиды с помощью набора для очистки плазмид без эндотоксинов.
ПРИМЕЧАНИЕ: Последовательность окончательного препарата плазмиды, используемого для инъекций, для проверки целостности всех компонентов. - Соедините соответствующие количества двух плазмид для получения смеси с конечной концентрацией 350 нг/мкл донорской плазмиды и 150 нг/мкл вспомогательной плазмиды при повторном суспендировании в инъекционном буфере.
ПРИМЕЧАНИЕ: При расчете необходимого объема смеси учитывайте, что 10-15 мкл достаточно для каждого дня плановых инъекций и ДНК может быть подготовлена заранее и сохранена при -20 °C. Сообщалось также о концентрациях плазмид интеграз-хелперов 60-500 нг/мкл и донорских плазмид 85-200 нг/мкл21,22,26,41. - Осаждение ДНК путем добавления 0,1 объема 3 М ацетата натрия (рН 5,2) и 2,5 объема ледяного 100% EtOH и вихря. Белый осадок должен быть сразу виден. Наличие высококонцентрированных исходных плазмидных препаратов (т.е. ~1 мкг/мкл) повышает эффективность осаждения.
ПРИМЕЧАНИЕ: Точка остановки - Осадок может храниться при температуре -20 °C в течение ночи. - Центрифугу при 15 000 х г в течение 20 мин при 4 °C, выбросьте супернатант и промыть гранулу 1 мл ледяного 70% EtOH.
- Промыть гранулу 1 мл ледяного 70% EtOH и центрифугой при 15 000 х г в течение 5 мин при комнатной температуре.
- Выбросьте супернатант, не нарушая гранулу, и высушите на воздухе.
- Повторно суспендировать гранулу в 1x инжекционном буфере (0,1 мМ Na3PO4, 5 мМ KCl, рН 7,2, 0,22 мкм фильтра стерилизованного) для достижения общей конечной концентрации 500 нг/мкл.
ПРИМЕЧАНИЕ: Предположим, что часть ДНК будет потеряна в процессе осаждения; поэтому сначала добавьте меньший объем инжекционного буфера, проверьте концентрацию на спектрофотометре (например, Nanodrop), а затем добавьте соответствующий оставшийся объем, чтобы достичь 500 нг/мкл. - Убедитесь, что ДНК тщательно суспендирована, подготовьте аликвоты по 10-15 мкл каждая и храните их при -20 °C.
- В день инъекции разморозьте одну аликвоту и центрифугу при 15 000 х г в течение 5 минут, чтобы удалить остатки твердых частиц.
ПРИМЕЧАНИЕ: Альтернативным методом удаления твердых частиц является фильтрация раствора через фильтр 0,22 мкм. Избегайте присутствия остатков твердых частиц в инъекционной смеси, поскольку они приводят к закупорке иглы во время микроинъекции эмбриона.
3. Микроинъекция эмбрионов из линии стыковки Anopheles
- Кровью кормят 4-7-дневных комаров из нужной стыковочной линии за 72 ч до микроинъекции (т.е. для инъекций в понедельник и вторник кормят самок в предыдущую пятницу; для инъекций в четверг и пятницу кормят самок в понедельник той же недели).
- Кровь питается комарами дикого типа (WT) (т.е. комарами с одинаковым геномным фоном стыковочной линии) в тот же день; они понадобятся для ауткроссинга.
ПРИМЕЧАНИЕ: Размер и качество кровяной муки влияют на качество яйцеклеток, поэтому рекомендуется всегда использовать свежую кровь (т.е. кровь, взятую в течение предыдущих 7 дней). Кормление руками или кормление мышей может увеличить качество и количество яиц, однако эти методы не поощряются. Конкретные утвержденные протоколы будут необходимы для использования человеком и животными. - Выполнение микроинъекций эмбриона
- Выполняйте микроинъекции эмбриона An. gambiae в 25 мМ NaCl42 , нацеливаясь на задний полюс эмбриона под углом 45 градусов. Подробный протокол сбора, выравнивания и микроинъекции эмбрионов см. в Pondeville et al.43 и Lobo et al.44.
- Выполняйте микроинъекции эмбриона An. stephensi в галогенуглеродном масле 700:27 (2:1), нацеливаясь на задний полюс эмбриона под углом 30 градусов. Подробный протокол сбора, выравнивания и микроинъекции эмбрионов можно найти в Terenius et al.45 и Lobo et al.44.
- Перенесите яйца сразу после инъекции в чашку Петри, наполненную стерильной дистиллированной водой (рН 7,2) и верните их в инсектарные условия.
- После вылупления ежедневно перекладывайте личинок G0 в лоток с подсоленной дистиллированной водой (0,1% тонизирующей соли) и задним ходом к куколкам.
- Регистрируйте скорость вылупления (т.е. количество вылупившихся личинок/количество введенных эмбрионов).
ПРИМЕЧАНИЕ: Движение эмбриона способствует вылуплению, поэтому желательно мягкое закручивание. Штриховка должна начинаться ~48 ч после впрыска. Поскольку инъекция может вызвать небольшую задержку развития, желательно вести наблюдение за поздно вылупляющимися личинками в течение 3-4 дней.
4. Скрещивание и скрининг трансформированных особей
- [НЕОБЯЗАТЕЛЬНЫЙ ШАГ] Экран G0 (инъекционный) 1-й или 2-й звездчатый личинок (L1-L2) для переходной экспрессии флуоресцентного маркера.
- Используйте стеклянную пипетку с тонким наконечником для переноса личинок G0 L1-L2 на предметные стекла микроскопа с лунками. Поместите по одной личинке в каждый колодец.
- Используйте флуоресцентный стереоскоп с соответствующим фильтром для скрининга на наличие переходной экспрессии флуоресцентного маркера.
ПРИМЕЧАНИЕ: Шаблон переходного выражения диктуется используемым промоутером. При использовании промотора 3xP3 преходящая экспрессия флуоресцентного маркера видна в анальных сосочках (см. Рисунок 6 в Pondeville et al.43) - Задние G0 положительные особи отдельно.
- Сортировка куколок G0 по полу под стереоскопом52.
- Пусть самцы появляются в отдельных клетках в группах по 3-5 (семьи основателей) и добавляют 10-кратное превышение соответствовавших возрасту самок WT.
ПРИМЕЧАНИЕ: Поскольку самцы спариваются несколько раз, важно обеспечить избыток самок WT, чтобы максимизировать шансы на спаривание каждого самца. - Пусть самки появляются в отдельных клетках в группах по 10-15 человек (семьи основателей) и добавляют равное количество соответствующих возрасту самцов WT.
ПРИМЕЧАНИЕ: Если в инсектарии ограниченное пространство, самки могут появиться все вместе в одной клетке. Соотношение женщин и мужчин может составлять всего 1 самец к 3 самкам. - Дайте взрослым спариваться в течение 4-5 дней и обеспечьте самок кровяной пищей.
ПРИМЕЧАНИЕ: Кровь питает и собирает яйца у самок G0 несколько раз, чтобы максимизировать шансы на получение трансформантов из нескольких гонотрофных циклов. - Кровь питается WT особей одновременно для ауткроссинга.
- Собирайте яйца и выращивайте G1 следующего поколения.
- Экран G1 L3-L4 личинок для соответствующей флуоресценции для идентификации трансформантов.
- Собирайте личинок в чашке Петри, выстланной фильтровальной бумагой, или на слайде микроскопа и экране с помощью флуоресцентного стереоскопа с соответствующими фильтрами для присутствия маркера, введенного с грузом, помеченным attB.
ПРИМЕЧАНИЕ: Флуоресценция, управляемая промотором 3xP3 , видна на всех постэмбриональных стадиях, и скрининг может быть выполнен на молодых личинках, однако они более хрупкие и должны обрабатываться относительно осторожно. Куколки также могут быть проверены.- Для одноаттб конструкций для интеграции экрана на наличие нового и уже существующего маркера; они оба должны присутствовать, так как новая кассета вставляется рядом с исходной (рисунок 3A, рисунок 4).
ПРИМЕЧАНИЕ: Исключение для экранирования для одиночных конструкций attB : При использовании стыковочных линий без маркера22 экран только для присутствия нового маркера. При использовании стыковочных линий, интеграция которых приводит к инактивации ранее существовавшего маркера21, экран на наличие нового маркера и потерю ранее существовавшего. - Для конструкций с двойным аттБ для RMCE, экрана для присутствия нового маркера и потери ранее существовавшего маркера должен присутствовать только вновь введенный маркер, поскольку новая кассета заменяет оригинальный (рисунок 3B, рисунок 5).
ПРИМЕЧАНИЕ: Случайные события интеграции могут быть восстановлены в экспериментах RMCE, где рекомбинируется только один attP и, таким образом, будут присутствовать оба маркера. Скрининг особей G1 может проводиться также на стадии куколки по той же процедуре52.
- Для одноаттб конструкций для интеграции экрана на наличие нового и уже существующего маркера; они оба должны присутствовать, так как новая кассета вставляется рядом с исходной (рисунок 3A, рисунок 4).
- Собирайте личинок в чашке Петри, выстланной фильтровальной бумагой, или на слайде микроскопа и экране с помощью флуоресцентного стереоскопа с соответствующими фильтрами для присутствия маркера, введенного с грузом, помеченным attB.
- Перенос превратил особей G1 в личиночный лоток и задний в куколок. Отбросьте нефлуоресцентных людей и людей с неожиданным маркерным паттерном выражения.
- Сортируйте преобразованных куколок G1 по полу и массово скрещивайте их с разнополыми по возрасту особями WT.
- Позвольте взрослым спариваться в течение 4-5 дней, обеспечьте кровяную пищу, соберите яйца и вырастите потомство следующего поколения G2 .
- Для одиночных интеграционных экспериментов собирайте яйца непосредственно из массового кросса , так как сайт интеграции идентичен у всех особей.
- Для экспериментов RMCE собирайте яйца у одиночных самок и поддерживайте потомство отдельно до тех пор, пока молекулярная оценка не будет завершена из-за потенциального присутствия двух альтернативных ориентаций кассет (рисунок 3B).
- Скрининг потомства G2 (либо на стадии личинки, либо на стадии куколки) на наличие флуоресцентного маркера (ожидается, что 50% особей будут положительными), отбросьте нефлуоресцентное потомство.
- Отложите подмножество G2-положительных людей для молекулярного анализа, остальных доведите до взрослой жизни.
ПРИМЕЧАНИЕ: Если все особи G2 должны быть сохранены в живых, молекулярный анализ может быть проведен на ногах одного взрослого человека46 или извлечении ДНК куколки (личное общение Л. Григораки). В качестве альтернативы, молекулярный анализ может быть выполнен после того, как все особи G2 яйцекладутся и яйца вылупятся. - Позвольте взрослым самцам и самкам скрещиваться в одной клетке, чтобы установить новую трансгенную линию.
ПРИМЕЧАНИЕ: Для экспериментов RMCE взрослое скрещивание должно происходить между братьями и сестрами, происходящими от одной самки, до тех пор, пока ориентация вставки не будет определена с помощью молекулярного анализа.
5. Молекулярная валидация сайта вставки методом амплификации ДНК (ПЦР)
- Подготовьте карту прогнозируемого места вставки в геном стыковочной линии после преобразования.
- Одиночная интеграция: убедитесь, что прогнозируемый сайт вставки несет исходную конструкцию стыковки плюс всю последовательность плазмиды донора между двумя гибридными сайтами attL и attR (рисунок 3A).
- RMCE: Убедитесь, что прогнозируемый сайт вставки идентичен сайту стыковочной линии, где гибридные инвертированные сайты attL заменяют исходные инвертированные сайты attP , а шаблон обмена заменяет кассету, первоначально присутствующую между ними (рисунок 3B).
- Проектирование олигонуклеотидных грунтовок для усиления инсерционного перехода по обе стороны от локуса интеграции.
- Единая интеграция: Проектирование пар олигонуклеотидных праймеров, которые охватывают участки attR и/или attL . Одна грунтовка должна связываться с ранее интегрированной стыковочной конструкцией, а другая — с вновь интегрированным трансгеном (рисунок 3А).
- RMCE: Замена кассеты может происходить в двух различных ориентациях по отношению к хромосоме (обозначенной A и B). Спроектируйте альтернативные комбинации из 4 олигонуклеотидных праймеров для получения дискретного продукта только в одной из ориентаций, причем одна пара является диагностической для ориентации A, а другая для ориентации B (рисунок 3B, рисунок 6).
- Извлеките геномную ДНК из G2-положительных лиц и выполните диагностическую ПЦР и гель-электрофорез для визуализации присутствия ожидаемых диагностических ампликонов из предсказанных карт сайта интеграции.
ПРИМЕЧАНИЕ: ДНК может быть альтернативно извлечена из ног одного взрослого человека46 или куколок (личное общение Л. Григораки). - Последовательность продуктов ПЦР для подтверждения ожидаемых последовательностей.

Рисунок 2. Диаграмма рабочего процесса для локальной модификации генома φC31 у комаров Anopheles . Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3. Молекулярная основа φC31-опосредованной одиночной интеграции (A) и RMCE (B). A) Схематические карты геномной вставки в стыковочной линии An. stephensi (80.9, таблица 1), несущей один участок attP и помеченной CFP (вверху), одноатт-аттБ-донорской плазмиды, помеченной DsRed (посередине), и ожидаемого сайта вставки, полученного после успешной интеграции (внизу). B) Схематические карты геномной вставки в стыковочной линии An. gambiae (A11, таблица 1), несущей два перевернутых участка attP и помеченной CFP (вверху), донорской плазмиды с двойным аттБ, помеченной YFP (посередине), и ожидаемого места вставки, полученного после успешного RMCE (внизу). Волнистая линия: геном комара; Полосатые стрелы: мешковые транспозонные руки; 3xP3: промоутер флуоресцентного маркера; SV40: вирусный терминатор; Ори: происхождение репликации; AmpR: ген устойчивости к ампициллину. Пересекающиеся линии представляют собой участок (участки) рекомбинации между участками attP и attB. Пронумерованные черные стрелки представляют собой места связывания грунтовки для молекулярной валидации локуса вставки (этап 5 протокола). Полностью аннотированные одинарные и двойные плазмиды с метками attB доступны у авторов по запросу. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Результаты
Протокол, проиллюстрированный здесь, позволяет генерировать стабильную трансгенную линию Anopheles за ~ 10 недель (при условии 21-дневного жизненного цикла комара).
Ожидается, что показатели вылупления личинок после инъекции у An. gambiae будут в целом ниже, чем у An. stephensi, однако сообщалось о показателях вылупления между 10-50%, 9,20,24,26,33,43,47. При соответствующей технике впрыска скорость вылупления ≥20%, как правило, достаточна для получения трансформаторов. Поглощение ДНК эмбрионами может быть оценено путем скрининга молодых личинок на преходящую экспрессию флуоресцентного маркера. В успешных экспериментах RMCE в An. gambiae с использованием промотора 3xP3 до 50% выживших личинок G0 показали эписомальную экспрессию маркера в анальных сосочках48.
Обобщенные оценки эффективности трансформации трудно оценить среди лабораторий и даже среди экспериментов, поскольку трансформация зависит от сложного взаимодействия переменных, включая чистоту, концентрацию, размер и потенциальную токсичность вводимой ДНК, качество яйцеклеток, обработку яиц до и после инъекции, выращивание комаров и, самое главное, опыт оператора. Коэффициенты трансформации до 7% были получены для RMCE в An. gambiae (рассчитывается как количество независимых трансформационных событий в общем количестве G0 индивидов)9,26,33, и до 2,2% коэффициент трансформации для интеграции в An. stephensi. Мы предлагаем ввести не менее 500 эмбрионов, что должно привести к вылуплению не менее 100 личинок G0 и к 2-7 взрослым основателям G0, из которых можно получить стабильно трансформированное потомство. При скрининге на преходящую экспрессию у личинок G0 можно ожидать до 40 положительных личинок.
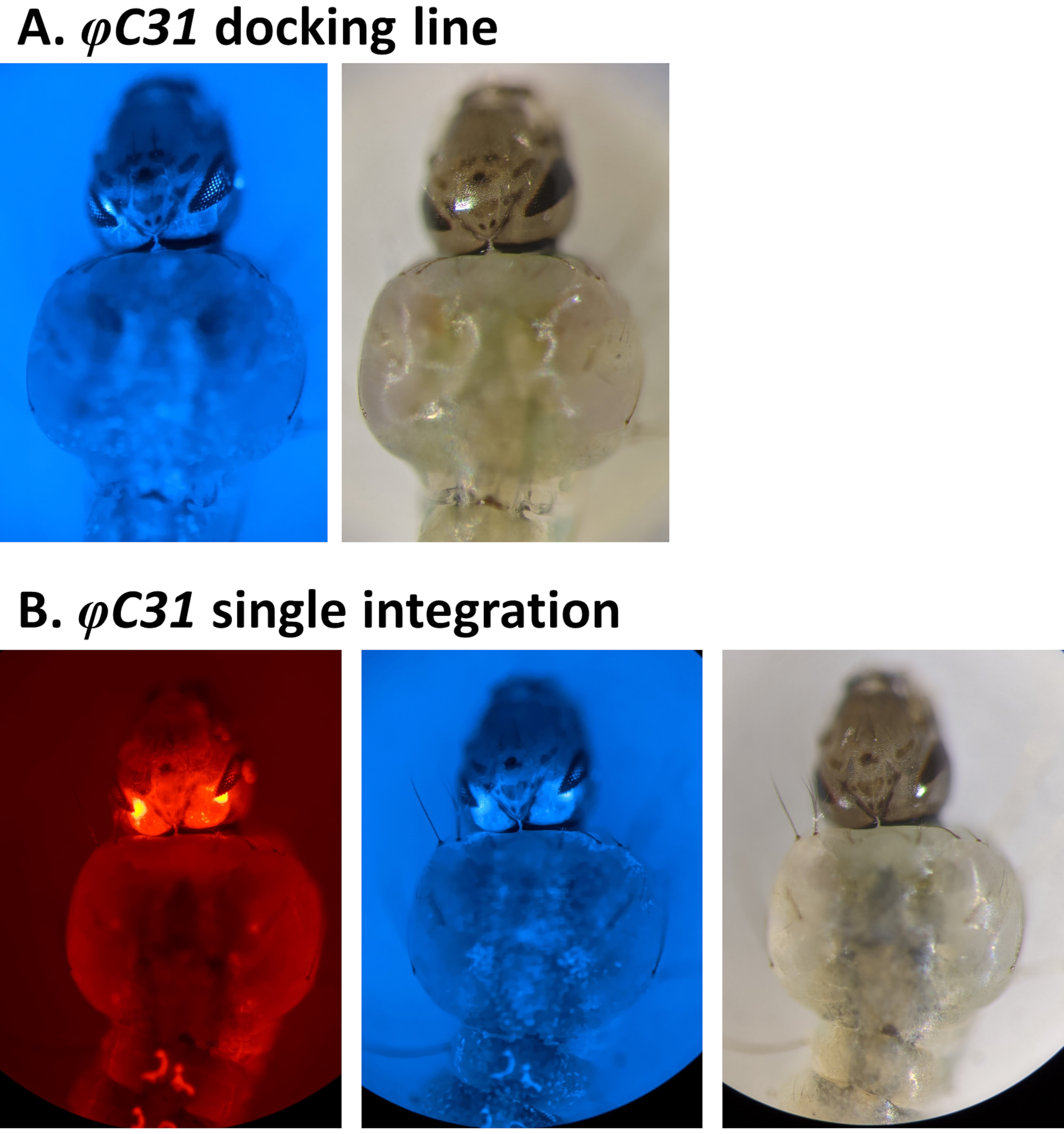
Примеры фенотипической валидации трансформации посредством скрининга флуоресцентных маркеров, регулируемых промотором 3xP3, приведены на рисунке 4 и рисунке 5. На рисунке 4 показана новая линия An. stephensi, полученная путем вставки кассеты с маркировкой DsRed в стыковочную линию, помеченную CFP (80.9, таблица 1), в результате чего потомство G1 выражает оба маркера, о чем свидетельствует красная и синяя флуоресценция, обнаруженная в глазах.
Вместо этого ожидается, что конструкции RMCE приведут к замене маркера, первоначально вставленного в линию стыковки, на маркер донорской плазмиды. Рисунок 5A и рисунок B иллюстрируют этот обмен маркерами в стыковочной линии An. gambiae, обозначенной CFP (A11, таблица 1), где после успешного RMCE маркер CFP теряется и приобретается маркер YFP, что приводит к флуоресценции желтого (но не синего) глаз и нервного канатика33. Иногда RMCE может привести к одному событию интеграции вместо обмена желаемой трансгенной кассетой, как показано на рисунке 5C, где показана личинка, помеченная как исходным CFP, так и новыми маркерами YFP. Сообщается, что до 50% от общего числа событий трансформации составляют единичные интеграции9,33.
При скрининге на наличие флуоресцентного маркера крайне важно отличить его сигнал от возможной фоновой автофлуоресценции. Это особенно важно при использовании CFP в качестве личинок Anopheles , демонстрирующих естественную синюю автофлуоресценцию (рисунок 6A). Увеличение и фокусировка на тканях и органах, где флуоресценция, как ожидается, будет управляться промотором, необходимы для идентификации истинных CFP-положительных людей, как показано на рисунке 6B с использованием маркера 3xP3-CFP.
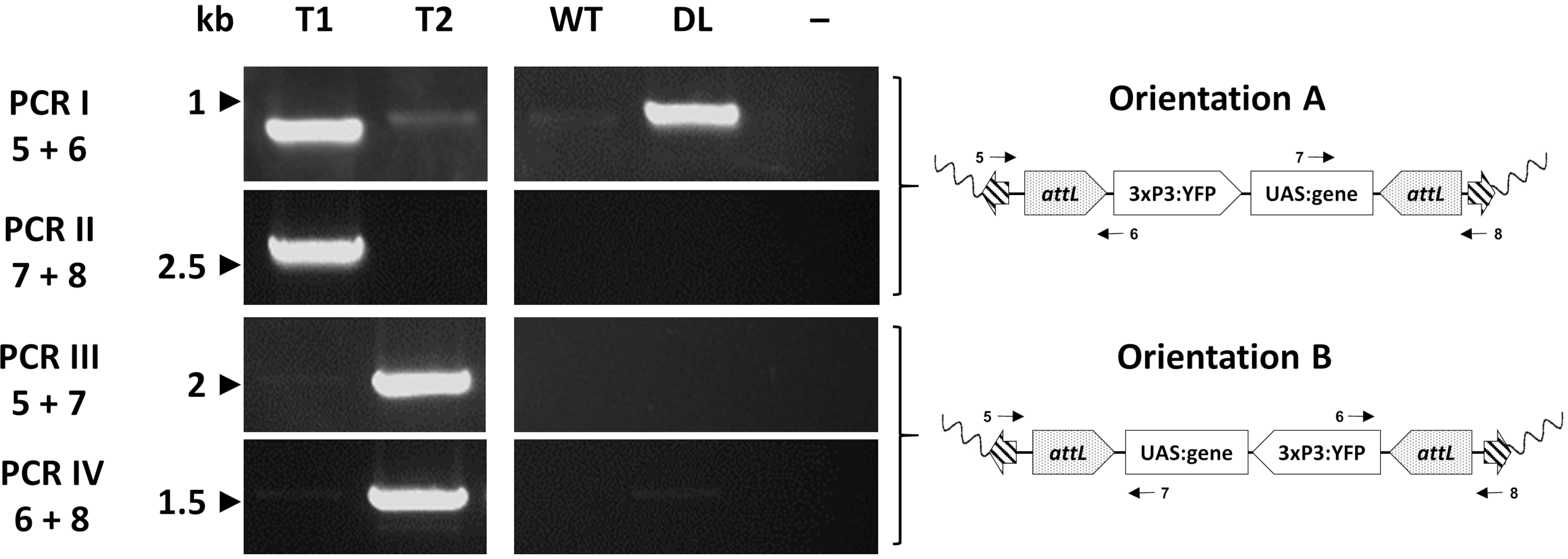
Отдельные трансформанты, наконец, оцениваются молекулярно с помощью ПЦР для подтверждения ожидаемого места введения. На рисунке 7 показана валидация ПЦР у отдельных лиц из обменной линии An. gambiae , показывающей две потенциальные ориентации вставки в геном комара33.

Рисунок 4. Валидация одиночной интеграции φC31 в личинках An. stephensi (дорсальный вид). A) Стыковочная линия (80.9, таблица 1) выражает CFP в глазах в соответствии с регулированием промоутера 3xP3 . B) Успешная интеграция приводит к выражению недавно приобретенного DsRed, а также оригинального маркера CFP в глазах. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 5. Валидация φC31 RMCE у личинок An. gambiae (вентральный вид). A) Стыковочная линия (A10, таблица 1) экспрессирует CFP в соответствии с регулированием промотора 3xP3 в глазах (e) и нервном канатике (nc)5. B) Успешный RMCE приводит к обмену флуоресцентного маркера с CFP на YFP33. C) Единичное событие интеграции произошло во время эксперимента RMCE, когда личинка трансформанта экспрессирует маркеры CFP и YFP. Эта личинка несет компоненты GAL4 / UAS, которые вызывают широкую экспрессию YFP, особенно сильную в мышцах живота (am). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 6. Автофлуоресценция CFP у личинок An. gambiae (дорсальный вид). A) Параллельное изображение положительной (CFP+) и отрицательной (CFP-) личинки L4 с использованием фильтра CFP. Б) Крупное изображение личиночных глаз, которое показывает CFP+ против CFP- индивидуума. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 7. Молекулярная валидация ориентации вставки кассеты в репрезентативном трансгенном An. gambiae , созданная φC31 RMCE. Трансгенная кассета может быть вставлена в одну из двух альтернативных ориентаций (A или B) по отношению к месту вставки. Каждая реакция ПЦР (I - IV) использует комбинацию праймеров (5-8)33 , предназначенных для получения фрагмента диагностической амплификации для каждой ориентации, как указано в схематических плазмидных картах. Т1: репрезентативная трансгенная индивидуальная несущая ориентация вставки А; Т2: репрезентативная трансгенная индивидуальная несущая ориентация вставки В; WT: дикий тип; DL: стыковочная линия; -: реакция отрицательного контроля, где вода использовалась в качестве шаблона. Эта цифра была изменена с Adolfi et al. (2019)33. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
| Вид | Напряжение | Имя | аттП(ы) | Хромо-некоторые | Промоутер-маркер | Учреждение происхождения | Ссылка |
| Ан. Стивенси | Индиана | 26.10б | Единственный | 2Р | 3xP3-eCFP | Калифорнийский университет в Ирвине | 25 |
| Ан. Стивенси | Индиана | 44Сб | Единственный | X | 3xP3-eCFP | Калифорнийский университет в Ирвине | 23, 24 |
| Ан. Стивенси | Индиана | 80.9б | Единственный | 2л | 3xP3-eCFP | Калифорнийский университет в Ирвине | Это исследование |
| An. gambiae | Г3 | 113 | Единственный | 2Р | 3xP3-eCFP | Калифорнийский университет в Ирвине | Это исследование |
| An. gambiae | КИЛЬ | Эк | Единственный | 3Р | 3xP3-eCFP | Килский университет. | 19, 43 |
| An. gambiae | Г3 | Х1 | Единственный | 2л | Нет маркера | Страсбургский университет | 22 |
| An. gambiae | Г3 | ЙаттП | Единственный | Y | 3xP3-RFP | Имперский колледж Лондона | 21 |
| An. gambiae | Г3 | А10б | Двойной | 2Р | 3xP3-eCFP | Ливерпульская школа Троп. Мед. | 5 |
| An. gambiae | Г3 | А11б | Двойной | 2Р | 3xP3-eCFP | Ливерпульская школа Троп. Мед. | 9 |
| a. Штамм из Университета Джона Хопкинса (подарок М. Джейкобса-Лорены) и в культуре Калифорнийского университета в Ирвине в течение >20 лет. | |||||||
| b. Эти строки доступны от авторов по разумному запросу. | |||||||
| c. Эта линия доступна в репозитории BEI www.beiresources.org как MRA-1163. | |||||||
Таблица 1. Стыковочные линии Anopheles attP .
Обсуждение
Точный дизайн плазмид с метками attB, совместимых с линией стыковки, имеет первостепенное значение для успеха эксперимента. Необходимо тщательно рассмотреть выбор маркера, используемого для экранирования трансформантов, включая цвет флуоресценции и его рисунок выражения, который будет зависеть от рисунка, уже присутствующего в стыковочной линии. Необходимо использовать флуоресцентные маркеры, которые легко различимы: хорошие комбинации маркеров включают RFP (красный) / CFP (голубой), RFP (красный) / GFP (зеленый), RFP (красный) / YFP (желтый) и YFP (желтый) / CFP (голубой), в то время как комбинации, которых следует избегать, - это YFP (желтый) / GFP (зеленый) и CFP (голубой) / GFP (зеленый). Промотор 3xP39, специфичный для глаз и нервного канатика, наиболее часто используется для управления экспрессией флуоресцентных маркеров для трансгенеза комаров. Действительно, все доступные в настоящее время стыковочные линии Anopheles используют этот промоутер. Альтернативными регуляторными областями являются ген полиубиквитина An. gambiae (PUBc)5 или вирусный промотор IE120, которые стимулируют экспрессию в нескольких тканях. При использовании вместе с 3xP3 эти промоторы расширят возможные цветовые комбинации и даже использование одного и того же флуорофора. Указанные промоторы активны на протяжении всего жизненного цикла комара, что позволяет проводить скрининг и мониторинг флуоресценции на всех этапах жизни. Дополнительным соображением при проектировании плазмид является размер груза, который должен быть интегрирован или обменен. Хотя система φC31 обладает замечательной несущей способностью18, следует учитывать, что размер донорской плазмиды обычно отрицательно коррелирует с эффективностью трансформации22.
В описанном протоколе источником интегразы является плазмида-помощник, экспрессирующая фермент повсеместно40. Повсеместное присутствие интегразы может привести к трансформации соматических клеток, если микроинъекции не направлены точно в область, где образуется зародышевая линия. Хотя такие трансформационные события будут потеряны, поскольку они не наследуются, соматические эффекты могут снизить пригодность инъекционных людей. Чтобы избежать этого и повысить эффективность трансформации, экспрессия интегразы может быть ограничена зародышевой линией, например, с помощью промотора ваза22,26. Другие протоколы описывают использование in vitro транскрибированной матричной РНК (мРНК) в качестве источника интегразы φC311111111. Однако это связано с трудоемким приготовлением мРНК и требует осторожного обращения с инъекционной смесью и использования реагентов, свободных от РНКазы, чтобы избежать деградации. Плазмидные источники интегразы были продемонстрированы как в An. gambiae9,21,22,26,33,37, так и в An. stephensi (A.A. personal communication) как надежные и ведущие к эффективной трансформации, и, таким образом, являются нашим предпочтительным вариантом. Еще одним вариантом доставки интегразы является ее производство in vivo в самостыковывающихся вспомогательных линиях. Такие линии были созданы в An. gambiae, которые экспрессируют интегразу φC31 под регуляцией специфического для зародышевой линии промоторных нано и, как было установлено, приводят к улучшению выживаемости и эффективности трансформации20. Однако необходимо учитывать потенциальные нагрузки на приспособленность, налагаемые выработкой in vivo фермента интегразы на хелперную линию.
Как и в случае с другими трансгенными методами, особое внимание должно быть уделено выращиванию и скрещиванию лиц, полученных из инъекционных эмбрионов, чтобы максимизировать шансы на восстановление трансформантов. Лица, которые стабильно унаследовали трансген, могут быть сначала восстановлены в потомстве G1. Однако ранние признаки потенциальной трансформации могут быть оценены по наличию преходящей эписомальной экспрессии флуоресцентного маркера в анальных сосочках и/или нервном канатике G0 первой и второй звезды при использовании промотора 3xP343. Хотя наличие переходной флуоресценции предполагает успешную доставку плазмид, это не гарантирует наследственную трансформацию зародышевой линии. Точно так же отсутствие переходного выражения не исключает успешного преобразования. Тем не менее, было замечено, что транзиторно положительные индивидуумы с большей вероятностью дают трансгенное потомство по сравнению с транзиторно отрицательными43,48. В руках экспертов выращивание и скрещивание только положительных особей может быть вариантом сокращения численности комаров. Однако, учитывая важность и хрупкость мелких личинок G0, наименьшее количество манипуляций все же целесообразно, и всегда рекомендуется выращивание всех особей G0.
Схема спаривания, описанная в этом протоколе, предназначена для максимизации вероятности спаривания и изоляции независимых событий трансформации. Однако, если наличие инсектарного пространства или персонала является проблемой, взрослые особи G0 могут быть объединены по полу в одиночные клетки, если предусмотрено достаточное количество разнополых особей. Такая установка не позволит различать множественные события трансформации, происходящие у людей из одной клетки. В зависимости от экспериментальной установки во время процесса скрининга ожидается наличие двойного (одинарная интеграция) или одинарного (RMCE) маркера. В экспериментах по одиночной интеграции важно проверить наличие исходного маркера из стыковочной линии, в то время как в RMCE важно проверить потерю ранее интегрированного маркера. Действительно, в конструкциях RMCE нередко восстанавливается трансформаторы, в которых единичная интеграция вместо обмена произошла из-за рекомбинации одного сайта attP9,33. У таких людей присутствуют как флуоресцентные маркеры, так и вся донорская плазмидная основа, что подчеркивает важность проведения тщательного скрининга на оба флуоресцентных маркера.
Хотя наличие ожидаемых флуоресцентных паттернов указывает на успешное преобразование, необходимо провести молекулярную характеристику места вставки. Для этого подготовка точных карт прогнозируемого локуса вставки, включая фланговые геномные области стыковочной линии, имеет решающее значение для разработки адекватных диагностических олигонуклеотидных праймеров для анализа амплификации генов. Одиночные события интеграции приводят к образованию гибридных сайтов attR и attL на стыке между вновь интегрированной ДНК и ранее вставленной кассетой. Эти сайты могут быть предназначены для проверки сайта вставки. В конструкциях RMCE вставка донорской кассеты может происходить в двух альтернативных ориентациях по отношению к геномному локусу, таким образом, четыре праймера могут быть использованы в альтернативных комбинациях ПЦР для определения того, какую ориентацию несет линия. Поскольку ориентация вставки кассеты может влиять на экспрессию трансгенов, в сравнительном анализе экспрессии генов важно использовать линии, несущие одинаковую ориентацию вставки.
При работе с низким количеством трансформантов может быть нежелательно жертвовать целыми особями для молекулярного анализа. Вариантом этого является проведение молекулярного анализа ДНК, извлеченной из ног одного взрослого человека46, поскольку потеря ног не влияет на способность взрослой женщины спариваться и яйцекладу49. Тем не менее, существует риск повреждения человека в процессе удаления ноги. Успех был достигнут с использованием выброшенных случаев куколки (личное общение Л. Григораки), однако самым безопасным подходом является проведение молекулярного анализа родителей G2 после получения жизнеспособного потомства G3.
В последние годы CRISPR/Cas9 произвел революцию в способе выполнения редактирования генома для конкретного сайта26,41,50,51. В отличие от RMCE, направленных на сайт, CRISPR/Cas9-опосредованные интеграции генов (knock-ins) не зависят от наличия предварительно вставленных сайтов рекомбинации, при этом требуется только одноэтапное событие трансформации. Тем не менее, система CRISPR/Cas9 опирается на наличие больших известных геномных последовательностей, фланкирующих желаемый сайт вставки, для успешного гомологического направленного восстановления, а также на эффективное распознавание сайта, опосредованное направляющими РНК. Эти условия не всегда могут быть выполнены или могут быть трудоемкими для устранения неполадок, и, учитывая наличие нескольких стыковочных линий в An. gambiae и An. stephensi и линий, полученных из них, система φC31 остается очень ценным инструментом для выполнения прямых фенотипических сравнений между трансгенами в одних и тех же геномных местоположениях.
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
Мы благодарны Кионе Паркер (UCI) за предоставление изображений трансгенных личинок An. stephensi , а также Фрейзеру Колману (LSTM) и Бет Полтон (LSTM) за предоставление трансгенных личинок An. gambiae . Бет Полтон (LSTM) также оказала ценную помощь во время визуализации личинок An. gambiae . Эта работа финансировалась Институтом генетики и общества Тата (TIGS) и Фондом катализаторов LSTM, присужденным A.A. (DCF2014AA). A.A.J. является профессором Дональда Брена в Калифорнийском университете в Ирвине.
Материалы
| Name | Company | Catalog Number | Comments |
| 1.5 mL eppendorf tubes | |||
| 8-well microslides | VWR | MARI1216690 | |
| DNeasy Blood & Tissue Kit | Qiagen | 69504 | |
| EndoFree Plasmid Maxi Kit (10) | Qiagen | 12362 | |
| Ethanol, Absolute, Molecular Biology Grade | |||
| Filter set CFP for Leica MZ FLIII Excitation 436/20 nm, extinction 480/40 nm | Leica | 10446363 | |
| Filter set dsRED for Leica MZ FLIII Excitation 545/30 nm, extinction 620/60 nm | Leica | 10447079 | |
| Filter set YFP customised for Leica MZ FLIII | Omega Optical | 500QM25, 500QM35 | |
| Halocarbon oil 27 | Sigma | H8773 | |
| Halocarbon oil 700 | Sigma | H8898 | |
| Petri dishes | |||
| Potassium chloride | |||
| Sodium Chloride | |||
| Sodium phosphate dibasic | |||
| Sodium Acetate Solution (3 M), pH 5.2 | Thermo Fisher Scientific (Life Technologies) | R1181 | |
| Stable brush Size 0 |
Ссылки
- Adolfi, A., Lycett, G. J. Opening the toolkit for genetic analysis and control of Anopheles mosquito vectors. Current Opinion in Insect Science. 30, 8-18 (2018).
- Grossman, G. L., Rafferty, C. S., Clayton, J. R., Stevens, T. K., Mukabayire, O., Benedict, M. Q. Germline transformation of the malaria vector, Anopheles gambiae, with the piggyBac transposable element. Insect Molecular Biology. 10 (6), 597-604 (2001).
- Nolan, T., Bower, T. M., Brown, A. E., Crisanti, A., Catteruccia, F. piggyBac-mediated germline transformation of the malaria mosquito Anopheles stephensi using the red fluorescent protein dsRED as a selectable marker. Journal of Biological Chemistry. 277 (11), 8759-8762 (2002).
- Perera, O. P., Harrell, R. A., Handler, A. M. Germ-line transformation of the South American malaria vector, Anopheles albimanus, with a piggyBac/EGFP transposon vector is routine and highly efficient. Insect Molecular Biology. 11 (4), 291-297 (2002).
- Adolfi, A., Pondeville, E., Lynd, A., Bourgouin, C., Lycett, G. J. Multi-tissue GAL4-mediated gene expression in all Anopheles gambiae life stages using an endogenous polyubiquitin promoter. Insect Biochemistry and Molecular Biology. 96, 1-9 (2018).
- Carballar-Lejarazú, R., Jasinskiene, N., James, A. Exogenous gypsy insulator sequences modulate transgene expression in the malaria vector mosquito, Anopheles stephensi. Proceedings of the National Academy of Sciences of the United States of America. 110 (18), 7176-7181 (2013).
- Galizi, R., et al. A synthetic sex ratio distortion system for the control of the human malaria mosquito. Nature Communications. 5, 3977(2014).
- Nolan, T., Petris, E., Müller, H. M., Cronin, A., Catteruccia, F., Crisanti, A. Analysis of two novel midgut-specific promoters driving transgene expression in Anopheles stephensi mosquitoes. PLoS ONE. 6 (2), 16471(2011).
- Lynd, A., Balabanidou, V., Vontas, J., Lycett, G. J. Development of a functional genetic tool for Anopheles gambiae oenocyte characterisation: appliction to cuticular hydrocarbon synthesis. BioRxiv. , (2019).
- O'Brochta, D. A., Alford, R. T., Pilitt, K. L., Aluvihare, C. U., Harrell, R. A., Harrell, R. A. piggyBac transposon remobilization and enhancer detection in Anopheles mosquitoes. Proceedings of the National Academy of Sciences of the United States of America. 108 (39), 16339-16344 (2011).
- O'Brochta, D. A., Pilitt, K. L., Harrell, R. A., Aluvihare, C., Alford, R. T. Gal4-based enhancer-trapping in the malaria mosquito Anopheles stephensi. G3. 2 (11), Bethesda. 1305-1315 (2012).
- Macias, V. M., et al. nanos-Driven expression of piggyBac transposase induces mobilization of a synthetic autonomous transposon in the malaria vector mosquito, Anopheles stephensi. Insect Biochemistry and Molecular Biology. 87, 81-89 (2017).
- Nimmo, D. D., Alphey, L., Meredith, J. M., Eggleston, P. High efficiency site-specific genetic engineering of the mosquito genome. Insect Molecular Biology. 15 (2), 129-136 (2006).
- Kim, A., Pyykko, I. Size matters: Versatile use of PiggyBac transposons as a genetic manipulation tool. Molecular and Cellular Biochemistry. 354, 301-309 (2011).
- Thorpe, H. M., Smith, M. C. M. In vitro site-specific integration of bacteriophage DNA catalyzed by a recombinase of the resolvase/invertase family. Proceedings of the National Academy of Sciences of the United States of America. 95 (10), 5505-5510 (1998).
- Khaleel, T., Younger, E., Mcewan, A. R., Varghese, A. S., Smith, M. C. M. A phage protein that binds φC31 integrase to switch its directionality. Molecular Microbiology. 80 (6), 1450-1463 (2011).
- Farruggio, A. P., Chavez, C. L., Mikell, C. L., Calos, M. P. Efficient reversal of phiC31 integrase recombination in mammalian cells. Biotechnology Journal. 7 (11), 1332-1336 (2012).
- Venken, K. J. T., He, Y., Hoskins, R. A., Bellen, H. J. P[acman]: A BAC Transgenic Platform for Targeted Insertion of Large DNA Fragments in D. melanogaster. Science. 314 (5806), 1747-1751 (2006).
- Meredith, J. M., et al. Site-specific integration and expression of an anti-malarial gene in transgenic Anopheles gambiae significantly reduces Plasmodium infections. PLoS ONE. 6 (1), 14587(2011).
- Meredith, J. M., Underhill, A., McArthur, C. C., Eggleston, P. Next-Generation Site-Directed Transgenesis in the Malaria Vector Mosquito Anopheles gambiae: Self-Docking Strains Expressing Germline-Specific phiC31 Integrase. PLoS ONE. 8 (3), 59264(2013).
- Bernardini, F., et al. Site-specific genetic engineering of the Anopheles gambiae Y chromosome. Proceedings of the National Academy of Sciences of the United States of America. 111 (21), 7600-7605 (2014).
- Volohonsky, G., et al. Tools for Anopheles gambiae Transgenesis. G3. 5 (6), Bethesda. 1151-1163 (2015).
- Amenya, D. A., et al. Comparative fitness assessment of Anopheles stephensi transgenic lines receptive to site-specific integration. Insect Molecular Biology. 19 (2), 263-269 (2010).
- Isaacs, A. T., et al. Transgenic Anopheles stephensi coexpressing single-chain antibodies resist Plasmodium falciparum development. Proceedings of the National Academy of Sciences of the United States of America. 109 (28), 1922-1930 (2012).
- Pham, T. B., et al. Experimental population modification of the malaria vector mosquito, Anopheles stephensi. PLoS Genetics. 15 (12), 1008440(2019).
- Hammond, A., et al. A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nature Biotechnology. 34 (1), 78-83 (2015).
- Groth, A. C., Fish, M., Nusse, R., Calos, M. P. Construction of Transgenic Drosophila by Using the Site-Specific Integrase from Phage phiC31. Genetics. 166 (4), 1775-1782 (2004).
- Franz, A. W. E., et al. Comparison of transgene expression in Aedes aegypti generated by mariner Mos1 transposition and site-directed recombination. Insect Molecular Biology. 20 (5), 587-598 (2011).
- Labbé, G., Nimmo, D., Alphey, L. piggybac-and PhiC31-Mediated Genetic Transformation of the Asian Tiger Mosquito, Aedes albopictus (Skuse). PLoS Neglected Tropical Diseases. 4 (8), 788(2010).
- Schetelig, M. F., Scolaric, F., Handler, A. M., Kittelmann, S., Gasperi, G., Wimmer, E. A. Site-specific recombination for the modification of transgenic strains of the Mediterranean fruit fly Ceratitis capitata. Proceedings of the National Academy of Sciences of the United States of America. 106 (43), 18171-18176 (2009).
- Yonemura, N., et al. phiC31-integrase-mediated, site-specific integration of transgenes in the silkworm, Bombyx mori (Lepidoptera: Bombycidae). Applied Entomology and Zoology. 43 (11), 997-1008 (2013).
- Bateman, J. R., Lee, A. M., Wu, C. T. Site-specific transformation of Drosophila via phiC31 integrase-mediated cassette exchange. Genetics. 173 (2), 769-777 (2006).
- Adolfi, A., Poulton, B., Anthousi, A., Macilwee, S., Ranson, H., Lycett, G. J. Functional genetic validation of key genes conferring insecticide resistance in the major African malaria vector, Anopheles gambiae. Proceedings of the National Academy of Sciences of the United States of America. 116 (51), 25764-25772 (2019).
- Haghighat-Khah, R. E., et al. Site-specific cassette exchange systems in the aedes aegypti mosquito and the Plutella xylostella moth. PLoS ONE. 10 (4), 0121097(2015).
- Long, D., Lu, W., Zhang, Y., Bi, L., Xiang, Z., Zhao, A. An efficient strategy for producing a stable, replaceable, highly efficient transgene expression system in silkworm, Bombyx mori. Scientific Reports. 5 (1), 8802(2015).
- Volohonsky, G., et al. Transgenic Expression of the Anti-parasitic Factor TEP1 in the Malaria Mosquito Anopheles gambiae. PLoS Pathogens. 13 (1), 1006113(2017).
- Grigoraki, L., Grau-Bové, X., Yates, H. C., Lycett, G. J., Ranson, H. Isolation and transcriptomic analysis of anopheles gambiae oenocytes enables the delineation of hydrocarbon biosynthesis. eLife. 9, 58019(2020).
- Kyrou, K., et al. A CRISPR-Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nature Biotechnology. 36 (11), 1062-1066 (2018).
- Berghammer, A. J., Klingler, M., Wimmer, E. A. A universal marker for transgenic insects. Nature. 402 (6760), 370-371 (1999).
- Ringrose, L. Transgenesis in Drosophila melanogaster. Methods in Molecular Biology. 561, 3-19 (2009).
- Dong, Y., Simões, M. L., Marois, E., Dimopoulos, G. CRISPR/Cas9 -mediated gene knockout of Anopheles gambiae FREP1 suppresses malaria parasite infection. PLoS Pathogens. 14 (3), 1006898(2018).
- Lombardo, F., Lycett, G. J., Lanfrancotti, A., Coluzzi, M., Arcà, B. Analysis of apyrase 5' upstream region validates improved Anopheles gambiae transformation technique. BMC research notes. 2, 24(2009).
- Pondeville, E., et al. Efficient ΦC31 integrase-mediated site-specific germline transformation of Anopheles gambiae. Nature Protocols. 9 (7), 1698-1712 (2014).
- Lobo, N. F., Clayton, J. R., Fraser, M. J., Kafatos, F. C., Collins, F. H. High efficiency germ-line transformation of mosquitoes. Nature protocols. 1 (3), 1312-1317 (2006).
- Terenius, O., Juhn, J., James, A. A. Injection of An. stephensi embryos to generate malaria-resistant mosquitoes. Journal of Visualized Experiments. 5, 216(2007).
- Lynd, A., et al. Insecticide resistance in Anopheles gambiae from the northern Democratic Republic of Congo, with extreme knockdown resistance (kdr) mutation frequencies revealed by a new diagnostic assay. Malaria Journal. 17 (1), 412(2018).
- Marinotti, O., et al. Development of a population suppression strain of the human malaria vector mosquito, Anopheles stephensi. Malaria Journal. 12 (1), 142(2013).
- Adolfi, A. In vivo functional genetic analysis of insecticide resistance in the malaria mosquito Anopheles gambiae. University of Liverpool. , PhD thesis (2017).
- Isaacs, A. T., Lynd, A., Donnelly, M. J. Insecticide-induced leg loss does not eliminate biting and reproduction in Anopheles gambiae mosquitoes. Scientific Reports. 7, 46674(2017).
- Gantz, V. M., et al. Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. Proceedings of the National Academy of Sciences of the United States of America. 112 (49), 6736-6743 (2015).
- Li, M., Akbari, O. S., White, B. J. Highly efficient site-specific mutagenesis in malaria mosquitoes using CRISPR. G3: Genes, Genomes, Genetics. 8 (2), 653-658 (2018).
- Poulton, B. C., et al. Using the GAL4-UAS System for Functional Genetics in Anopheles gambiae. J. Vis. Exp. , (2021).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены