
Method Article
Integração mediada pelo site φC31 e troca de em vetores anofelinos da malária
Neste Artigo
Resumo
O protocolo descreve como alcançar modificações direcionadas ao local no genoma dos mosquitos anofelinos da malária usando o sistema φC31 . As modificações descritas incluem tanto a integração quanto a troca de fitas transgênicas no genoma das linhas de acoplamento de rolamento attP.
Resumo
A análise genômica funcional e estratégias relacionadas para o controle genético da malária dependem de métodos validados e reprodutíveis para modificar com precisão o genoma dos mosquitos Anopheles. Entre esses métodos, o sistema φC31 permite uma integração precisa e estável direcionada ao local de transgenes, ou a substituição de fitas transgênicas integradas por troca de mediadas por recombinase (RMCE). Este método baseia-se na ação do Streptomyces φC31 bacteriophage integrase para catalisar a recombinação entre dois locais específicos de fixação designados attP (derivado da praga) e attB (derivado da bactéria hospedeira). O sistema usa um ou dois locais attP que foram integrados anteriormente ao genoma do mosquito e ao site attB no DNA do modelo de doador. Aqui ilustramos como modificar o genoma das linhas de acoplamento Anopheles portadores de attP usando dois plasmídeos: um doador com etiqueta attB carregando o modelo de integração ou troca e um ajudante plasmid codificando o integrase φC31. Relatamos dois resultados representativos de modificação mediada pelo site φC31: a integração única de uma fita transgênica em An. stephensi e RMCE em mosquitos An. gambiae. A manipulação de genoma mediada por φC31 oferece a vantagem da expressão transgênica reprodutível de locais genômicos validados e neutros em aptidão, permitindo análises qualitativas e quantitativas comparativas de fenótipos. A natureza direcionada ao local da integração também simplifica substancialmente a validação do local de inserção única e do esquema de acasalamento para obter uma linha transgênica estável. Essas e outras características fazem do sistema φC31 um componente essencial do kit de ferramentas genéticas para a manipulação transgênica de mosquitos da malária e outros vetores de insetos.
Introdução
A capacidade de modificar o genoma dos mosquitos vetores de doenças de forma confiável e reprodutível reforçou a validação funcional in vivo de genes e abriu as portas para estratégias de controle genético de vetores, como as que visam mosquitos Anopheles que transmitem malária1.
A edição precoce do genoma do mosquito dependia apenas da transformação transposável (TE) mediada por piggyBac, sendo o co-co-mosquito o transposon mais usado em Anopheles2,3,4. No entanto, a natureza aleatória da integração do TE pode levar a modificações indesejáveis, como nocautes genéticos (mutagênese insercional) e efeitos significativos de posição na expressão transgênese5,6,7,8. Múltiplas inserções também são comuns ao usar o piggyBac5,9, o que torna a validação e o isolamento de linhas transgênicas com inserções únicas laboriosas. Outras desvantagens incluem sua potencial remobilização, como observado na linha germinal de Anopheles stephensi ao fornecer uma fonte de transposase 10,11,12, e seu tamanho limitado de carga de DNA (10-15 kb de comprimento) com a eficiência de transformação diminuindo com o tamanho crescente do doador plasmid13,14.
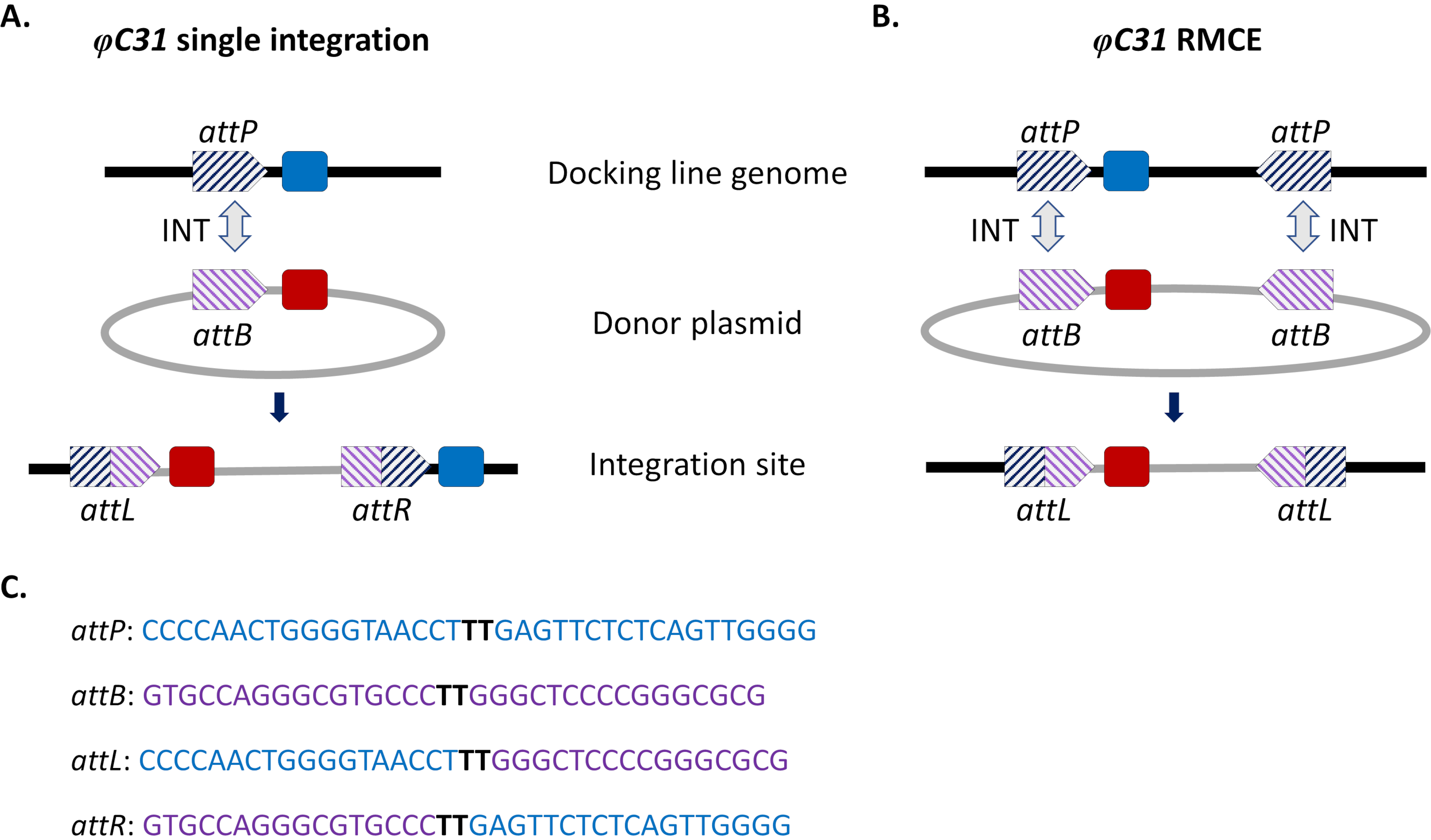
Abordagens de integração direcionadas ao site foram introduzidas para contornar esses problemas. A modificação de genoma mais comum em mosquitos é a mediada pelo sistema φC31 (Figura 1a). Isso é impulsionado por uma integração viral que catalisa a recombinação entre dois sítios de apego heteroespecífico (att) que ocorrem naturalmente no genoma do bacteriófago φC31 (attP) e no hospedeiro de bactérias estreptomíceos (attB)15. A recombinação dos dois locais é unidirecional e resulta na formação de sítios híbridos (attL e attR). A recombinação desses locais híbridos (levando à excisão de DNA) exigiria não apenas a presença de uma integração viral ativa, mas também outro fator de recombinação codificado por phage16,17. Assim, é gerado um local de integração estável que alivia a questão da remobilização indesejada em potencial15. Além disso, o sistema permite a integração de grandes cargas (por exemplo, a integração de construções de >100 kb foi relatada em D. melanogaster18), aumentando significativamente as capacidades de transporte. A integração ocorre em um único lócus genômico predefinido que simplifica muito a validação da inserção e o esquema de acasalamento para obter uma linha transgênica estável. Finalmente, a natureza direcionada ao local da integração permite a normalização da expressão, pois transgenes alternativos estão localizados no mesmo lócus e, portanto, são regulados dentro do mesmo contexto genômico vizinho. De fato, uma das principais aplicações da técnica é a comparação direta dos fenótipos conferidos por diferentes transgênicos após a inserção em um lócus idêntico.
Alcançar a integração mediada pelo ΦC31 envolve duas fases: a fase I é a criação de linhas transgênicas de acoplamento que transportam attP(s), e a fase II é a integração direcionada ao local de uma carga ladeada por ATTB no genoma da linha de acoplamento19. A criação de linhas de acoplamento de fase I baseou-se na integração aleatória mediada pelo TE de construções marcadas por ATTP e, portanto, envolveu um processo inicial trabalhoso (incluindo análises de PCR sulistas e inversas em prole unimpresa feminina) para isolar e validar linhas transgênicas que carregam um único evento de integração em locais genômicos neutros únicos, transcricionistas e neutros em aptidão. No entanto, várias linhas de acoplamento para integração única mediada por φC31 foram desenvolvidas e validadas em An. gambiae19,20,21,22 e em An. stephensi23,24,25 (Tabela 1). Cada uma dessas linhas varia em termos da localização genômica do local de atracação e do fundo genético específico da cepa e delas uma grande variedade de novas linhas transgênicas podem ser criadas. A complexa validação de integrações mediadas pelo TE para a produção de linhas de acoplamento pode agora ser contornada pela tecnologia CRISPR/Cas926; no entanto, isso se baseia no conhecimento a priori de loci neutro a ser direcionado e suas sequências circundantes.
A integração mediada pelo φC31 tem sido aplicada extensivamente à edição do genoma dos insetos do organismo modelo D. melanogaster27, aos mosquitos Aedes aegypti13,28, Ae. albopictus29, An. gambiae19 e An. stephensi24, bem como outros insetos, incluindo Ceratitis capitata30 e Bombyx mori31.
Uma limitação da integração mediada pelo ΦC31, especialmente tendo em vista possíveis liberações de campo para controle vetorial, é a integração no genoma do mosquito de todo o plasmídeo doador portador de ATTB, incluindo sequências indesejáveis, como marcadores genéticos de resistência a antibióticos e componentes de espinha dorsal plasmida de origem bacteriana. Para lidar com isso, foi implementada uma modificação do sistema padrão, a troca de mediada por recombinase (RMCE), que permite a substituição precisa de um transgênico previamente integrado por um novo DNA doador (Figura 1b). Isso é conseguido usando dois locais de att invertidos flanqueando as fitas de doador e receptor em cada extremidade, o que leva dois eventos independentes de recombinação a ocorrer simultaneamente resultando em troca de sem integração da espinha dorsal plasmida. Este projeto melhorado contorna a integração de sequências indesejadas e expande a aplicação de sistemas φC31 para incluir, por exemplo, a integração de cargas de DNA não marcadas por triagem para a perda de um marcador fluorescente anteriormente integrado32.
A RMCE foi alcançada primeiro com D. melanogaster32 e posteriormente aplicada com sucesso a insetos não-modelo, incluindo An. gambiae9,26,33, Ae. aegypti34, Plutella xylostella34 e B. mori35. Várias linhas de acoplamento para RMCE foram desenvolvidas e validadas em An. gambiae5,9,26 (Tabela 1). Para nosso conhecimento, a RMCE ainda está para ser explorada em outras espécies de vetores Anopheles.
Até o momento, o sistema φC31 tem sido amplamente utilizado em mosquitos Anopheles para introduzir e estudar uma variedade de moléculas, incluindo efeitos antimalárias19,24,36, componentes do sistema GAL4/UAS para superexpressar e derrubar genes para estudos de resistência a inseticidas9,33, elementos regulatórios, genes repórteres5,21,37, e elementos de unidade genética26 38.
Este protocolo descreve como executar 1) integração direcionada ao local de uma carga flanqueada attB e 2) RMCE de uma construção ladeada por locais attB invertidos no genoma das linhas de acoplamento anopheles . Isso é conseguido usando dois plasmídeos: um plasmídeo marcado por attB com um doador com etiqueta de transgene de interesse, e um plasmid auxiliar expressando a integração φC31 . Os principais vetores de malária An. gambiae e An. stephensi são usados como exemplos específicos, no entanto esses protocolos são aplicáveis a outras espécies de Anopheles .

Figura 1. Modificações de genoma dirigidas pelo local, integração única e troca de mediada por recombinase (RMCE), utilizando o sistema φC31. O φC31 integrase (INT, seta dupla cinza) catalisa a recombinação entre o local attB (listrado roxo) presente em um plasmídeo doador e o site attP(s) (listrado azul) presente em uma linha de acoplamento receptor, o que resulta na formação de locais híbridos attL e attR. A) A integração é alcançada quando os locais attB e attP únicos se recombinam e resulta na presença de dois marcadores integrados (azul e vermelho). B) A RMCE ocorre quando dois locais attB/P se recombinam simultaneamente e resultam na substituição do entre os locais att da linha de acoplamento (marcador azul) com o transportado pelo plasmídeo doador (marcador vermelho). C) Sequências parciais de nucleotídeos de attP (azul) e attB (roxo) e os locais híbridos attL/R. A recombinação ocorre entre as sequências do núcleo 'TT' destacadas em preto negrito. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Protocolo
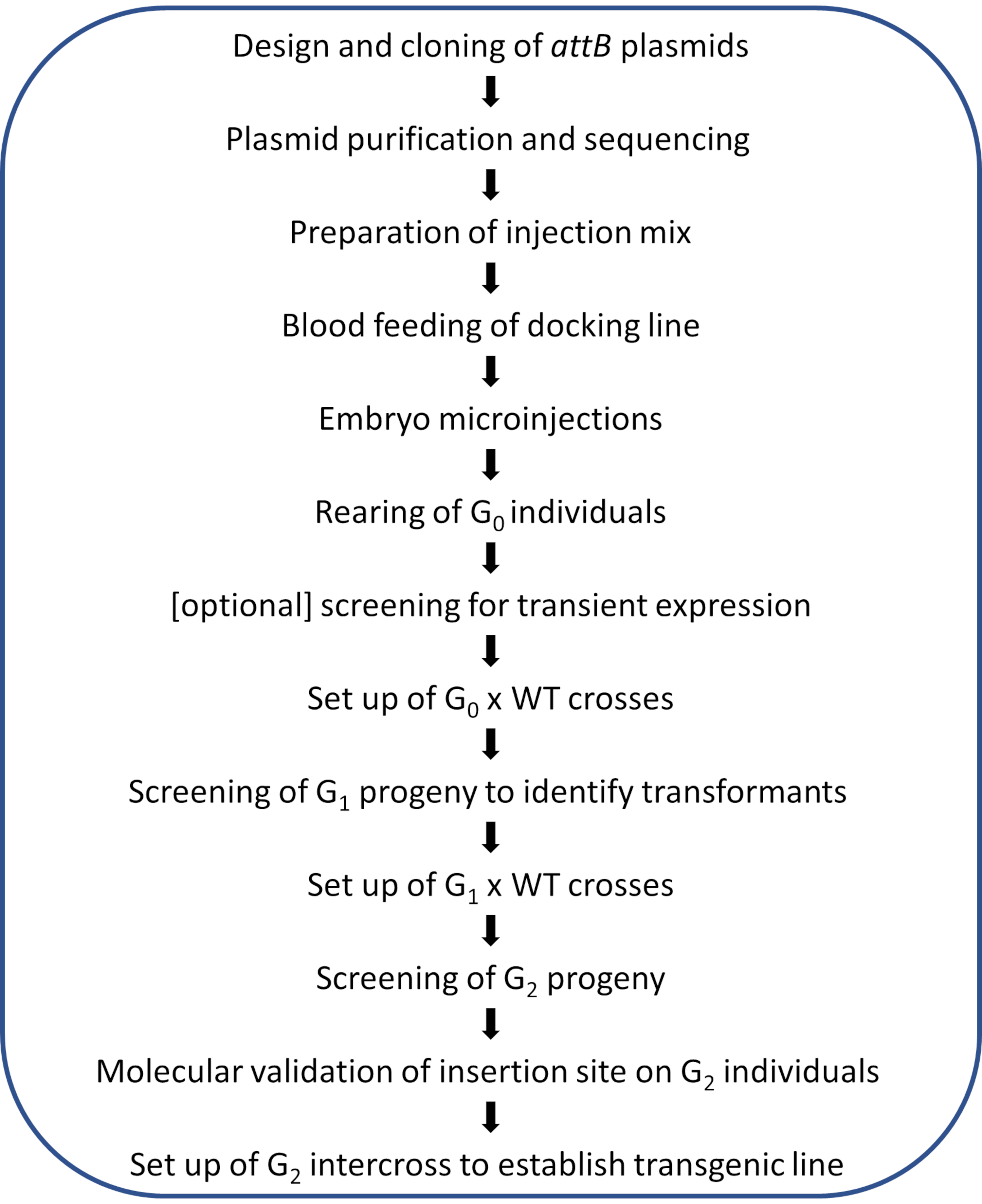
NOTA: Um fluxo de trabalho esquemático do protocolo ilustrado é mostrado na Figura 2.
1. Design de plasmídeos marcados por φC31 attB (Figura 3)
- Criar plasmídeos doadores attB carregando os seguintes componentes essenciais
- Marcador fluorescente dominante
- Escolha um promotor para conduzir a expressão do marcador fluorescente.
NOTA: Para a transgênese anofeeles, os marcadores fluorescentes geralmente estão sob a regulamentação do promotor 3xP3339, que impulsiona a expressão nos olhos e no fio nervoso. Alternativamente, o promotor PUBc5 pode ser usado quando a expressão em múltiplos tecidos é desejada. Plasmids doadores e linhas de acoplamento usadas como exemplos neste protocolo são marcados usando o promotor 3xP3. - Escolha uma proteína fluorescente (FP) compatível com a linha de acoplamento receptora para que elas sejam prontamente distinguíveis.
NOTA: Não utilize o mesmo marcador que já esteja presente na linha de acoplamento e evite o uso simultâneo de GFP (verde)/YFP (amarelo) e GFP (verde)/CFP (ciano), pois são muito difíceis de diferenciar de forma confiável. Os plasmídeos doadores usados como exemplos neste protocolo são marcados com DsRed ou YFP, pois devem ser integrados em uma linha de acoplamento marcada com CFP.
- Escolha um promotor para conduzir a expressão do marcador fluorescente.
-
site de recombinação attB(s)
- Use um único site attB para integração de um transgênico (design single-attB ) (Figura 3A).
- Use dois locais de attB invertidos para RMCE (design de attB duplo) onde os locais estavam invertidos entre si e incluem o modelo de DNA do doador (Figura 3B).
NOTA: A orientação do site attB deve ser compatível com a dos sites attP presentes na linha de acoplamento.
- Carga transgênica desejada
- Use quaisquer outros recursos desejados para serem integrados no genoma do mosquito com base no propósito específico do experimento. Aqui, descrevemos a integração de uma molécula efeitousa antimalárica no genoma de An. stephensi e a integração dos componentes do sistema GAL4/UAS em mosquitos An. gambiae .
- Componentes da coluna vertebral plasmid
- Inclua, entre outros componentes essenciais para a replicação plasmida em bactérias, um marcador para seleção de plasmídeos in vitro (ou seja, um gene de resistência a antibióticos).
NOTA: A espinha dorsal plasmida será integrada no genoma do mosquito no projeto single-attB para integração (Figura 3A), enquanto não será inserida no design de duplo attB para RMCE (Figura 3B).
- Inclua, entre outros componentes essenciais para a replicação plasmida em bactérias, um marcador para seleção de plasmídeos in vitro (ou seja, um gene de resistência a antibióticos).
- Marcador fluorescente dominante
2. Preparação de plasmídeos para a mistura de microinjeção
NOTA: O protocolo ilustrado aqui envolve o uso de dois plasmídeos: um plasmídeo doador marcado por attB carregando o transgene de interesse, e um plasmídeo auxiliar que expressa a integração φC31 sob a regulamentação do promotor Drosophila Hsp7040.
- Purificar plasmídeos doadores e ajudantes usando um kit de purificação plasmida sem endotoxina.
NOTA: Sequencie a preparação final do plasmídeo usada para injeção para verificar a integridade de todos os componentes. - Combine quantidades apropriadas dos dois plasmídeos para obter uma mistura com uma concentração final de 350 ng/μL do plasmídeo doador e 150 ng/μL do plasmídeo auxiliar quando resuspendido em tampão de injeção.
NOTA: Ao calcular o volume necessário de mistura, considere que 10-15 μL são suficientes para cada dia de injeções planejadas e o DNA pode ser preparado com antecedência e armazenado a -20 °C. Também foram relatadas concentrações plasmid de 60-500 ng/μL e de doadores plasmídeos de 85-200 ng/μL. - Precipitar o DNA adicionando 0,1 volumes de acetato de sódio de 3 M (pH 5.2) e 2,5 volumes de gelo frio 100% EtOH e vórtice. Um precipitado branco deve ser imediatamente visível. Ter preparações pládidas iniciais altamente concentradas (ou seja, ~1 μg/μL) melhora a eficiência da precipitação.
NOTA: Ponto de parada - O precipitado pode ser armazenado a -20 °C durante a noite. - Centrífuga a 15.000 x g por 20 min a 4 °C, descarte o supernasce e lave a pelota com 1 mL de gelo frio 70% EtOH.
- Lave a pelota com 1 mL de gelo frio 70% EtOH e centrífuga a 15.000 x g por 5 min à temperatura ambiente.
- Descarte o supernatante sem perturbar a pelota e o ar seco.
- Resuspende a pelota em tampão de injeção de 1x (0,1 mM Na3PO4, 5 mM KCl, pH 7,2, filtro de 0,22 μm esterilizado) para atingir uma concentração final total de 500 ng/μL.
NOTA: Suponha que algum DNA será perdido durante o processo de precipitação; portanto, adicione um volume menor de tampão de injeção primeiro, verifique a concentração em um espectrômetro (por exemplo, Nanodrop) e, em seguida, adicione um volume restante apropriado para atingir 500 ng/μL. - Certifique-se de que o DNA está completamente resuspentado, prepare alíquotas de 10-15 μL cada e armazene-as a -20 °C.
- No dia da injeção, descongele uma alíquota e centrífuga a 15.000 x g por 5 min para remover quaisquer resíduos de partículas.
NOTA: Um método alternativo para a remoção de partículas é filtrar a solução através de um filtro de 0,22 μm. Evite a presença de resíduos de partículas na mistura de injeção, pois levam ao bloqueio da agulha durante a microinjeção do embrião.
3. Microinjeção de embriões de uma linha de ancoragem Anopheles
- Alimentação sanguínea mosquitos de 4 a 7 dias de idade da linha de acoplamento desejada 72 h antes da microinjeção (ou seja, para injeção na segunda e terça-feira alimentar fêmeas na sexta-feira anterior; para injeção na quinta e sexta-feira alimentar fêmeas na segunda-feira da mesma semana).
- Os mosquitos do tipo wild (WT) de alimentação sanguínea (WT) (ou seja, mosquitos com o mesmo fundo genômico da linha de acoplamento) no mesmo dia; estes serão necessários para o outcrossing.
NOTA: O tamanho e a qualidade da refeição sanguínea afetam a qualidade do ovo, por isso recomenda-se sempre usar sangue fresco (ou seja, sangue extraído nos últimos 7 dias). A alimentação ou alimentação de braço em camundongos pode aumentar a qualidade e a quantidade de ovos, no entanto, esses métodos não são incentivados. Serão necessários protocolos aprovados específicos para uso humano e animal. - Realizar microinjeções de embriões
- Realize microinjeções de embriões de An. gambiae em 25 mM NaCl42 mirando o polo posterior do embrião em um ângulo de 45 graus. Para um protocolo detalhado para coleta, alinhamento e microinjeção de embriões, consulte Pondeville et al.43 e Lobo et al.44.
- Realize microinjeções de embrião an. stephensi em óleo de halocarbono 700:27 (2:1) mirando o polo posterior do embrião em um ângulo de 30 graus. Um protocolo detalhado para coleta, alinhamento e microinjeção de embriões pode ser encontrado em Terenius et al.45 e Lobo et al.44.
- Transfira os ovos imediatamente após a injeção em uma placa de Petri cheia de água destilada estéril (pH 7.2) e devolva-os a condições insetídas.
- Após a eclosão, transfira larvas G0 para uma bandeja com água salgada destilada (0,1% de sal tônico) diariamente e volte para pupas.
- Taxa de incubação recorde (ou seja, número de larvas eclodidas/número de embriões injetados).
NOTA: O movimento do embrião ajuda a eclodir, por isso o redemoinho suave é desejável. A eclosão deve começar ~48 h após a injeção. Uma vez que a injeção pode causar um pequeno atraso no desenvolvimento, é aconselhável continuar monitorando larvas de eclosão tardia por 3-4 dias.
4. Travessia e triagem de indivíduos transformados
- [PASSO OPCIONAL] Tela G0 (injetada) 1ª ou 2ª larvas instar (L1-L2) para expressão transitória do marcador fluorescente.
- Use uma pipeta de vidro de ponta fina para transferir larvas G0 L1-L2 para um microscópio desliza com poços. Coloque uma larva em cada poço.
- Use um estereoscópio de fluorescência com o filtro apropriado para tela para a presença de expressão transitória do marcador fluorescente.
NOTA: O padrão de expressão transitória é ditado pelo promotor utilizado. Ao usar o promotor 3xP3, a expressão transitória do marcador fluorescente é visível nas papilas anais (ver Figura 6 em Pondeville et al.43) - Indivíduos positivos do G0 traseiro separadamente.
- Classificar G0 pupae por sexo sob um estereoscópio52.
- Deixe os machos emergirem em gaiolas separadas em grupos de 3-5 (famílias fundadoras) e adicione um excesso de 10 vezes de fêmeas WT com correspondência etária.
NOTA: Uma vez que os machos acasalam várias vezes, é importante fornecer um excesso de fêmeas WT para maximizar as chances de acasalamento de cada macho. - Que as fêmeas emerjam em gaiolas separadas em grupos de 10-15 (famílias fundadoras) e adicione um número igual de machos WT com idade compatível.
NOTA: Se houver espaço limitado no inseto, as fêmeas podem emergir todas juntas em uma única gaiola. A proporção feminina para masculina pode ser tão baixa quanto 1 macho para 3 fêmeas. - Permita que os adultos acasalem por 4-5 dias e forneçam às fêmeas uma refeição de sangue.
NOTA: O sangue alimenta e coleta ovos de fêmeas G0 várias vezes para maximizar as chances de obter transformadores de múltiplos ciclos gonotróficos. - Sangue alimenta indivíduos WT ao mesmo tempo para outcrossing.
- Coletar ovos e criar g1s emergentes na próxima geração.
- Tela G1 L3-L4 larvas para fluorescência apropriada para identificar transformadores.
- Colete larvas em uma placa de Petri forrada com papel filtro ou em um slide de microscópio e tela usando um estereoscópio fluorescente com filtros apropriados para a presença do marcador introduzido com a carga marcada por attB.
NOTA: A fluorescência impulsionada pelo promotor 3xP3 é visível em todas as etapas pós-mbrínicas e a triagem pode ser realizada em larvas mais jovens, porém estas são mais frágeis e devem ser tratadas com relativamente cuidado. Pupae também pode ser exibido.- Para projetos single-attB para tela de integração para a presença do marcador novo e pré-existente; ambos devem estar presentes, uma vez que o novo está inserido ao lado do original (Figura 3A,Figura 4).
NOTA: Exceção de triagem para designs attB únicos: Ao usar linhas de acoplamento sem marcador22, tela apenas para a presença do novo marcador. Ao usar linhas de acoplamento onde a integração resulta na inativação do marcador pré-existente21, tela para a presença do novo marcador e a perda do pré-existente. - Para projetos de attB duplo para RMCE, tela para a presença do novo marcador e a perda do marcador pré-existente, apenas o marcador recém-introduzido deve estar presente, uma vez que o novo substitui o original (Figura 3B, Figura 5).
NOTA: Eventos de integração ocasionais podem ser recuperados em experimentos RMCE onde apenas um único attP recombinado e, portanto, ambos os marcadores estarão presentes. A triagem de indivíduos do G1 também pode ser realizada na fase pupa após o mesmo procedimento52.
- Para projetos single-attB para tela de integração para a presença do marcador novo e pré-existente; ambos devem estar presentes, uma vez que o novo está inserido ao lado do original (Figura 3A,Figura 4).
- Colete larvas em uma placa de Petri forrada com papel filtro ou em um slide de microscópio e tela usando um estereoscópio fluorescente com filtros apropriados para a presença do marcador introduzido com a carga marcada por attB.
- A transferência transformou os indivíduos do G1 em uma bandeja de larvas e traseira para pupas. Descarte indivíduos e indivíduos não fluorescentes com um padrão de expressão de marcador inesperado.
- Sort transformou g1 pupae por sexo e cruzá-los em massa com indivíduos com idade oposta combinada com WT.
- Permita que os adultos acasalem por 4-5 dias, forneçam uma refeição de sangue, recolham os ovos e criem a prole G2 da próxima geração.
- Para experimentos de integração única, colete ovos diretamente da cruz em massa , pois o local de integração é idêntico em todos os indivíduos.
- Para experimentos de RMCE, colete ovos de fêmeas solteiras e mantenha a prole separada até que a avaliação molecular seja completa devido à presença potencial de duas orientações alternativas de (Figura 3B).
- Tela a progêneria G2 (na fase larva ou pupa) para a presença do marcador fluorescente (50% dos indivíduos devem ser positivos), descartar progênero não fluorescente.
- Reserve um subconjunto de indivíduos positivos do G2 para análise molecular, e crie o resto até a idade adulta.
NOTA: Se todos os indivíduos do G2 devem ser mantidos vivos, a análise molecular pode ser realizada nas pernas de um único adulto46 ou extrações de DNA de caso pupal (comunicação pessoal L. Grigoraki). Alternativamente, a análise molecular pode ser realizada depois que todos os indivíduos do G2 ovipositados e ovos eclodiram. - Permitir que machos e fêmeas adultos intercruzem na mesma gaiola para estabelecer a nova linha transgênica.
NOTA: Para experimentos de RMCE, o intercross adulto deve ocorrer entre irmãos provenientes de uma única fêmea até que a orientação da inserção seja determinada através da análise molecular.
5. Validação molecular do local de inserção por amplificação de DNA (PCR)
- Prepare um mapa do local de inserção previsto no genoma da linha de acoplamento após a transformação.
- Integração única: Certifique-se de que o local de inserção previsto carregue a construção original de acoplamento mais toda a sequência do plasmídeo do doador entre os dois locais híbridos attL e attR (Figura 3A).
- RMCE: Certifique-se de que o local de inserção previsto seja idêntico ao da linha de acoplamento onde os locais híbridos invertidos de ATTL substituem os locais invertidos originais e o modelo de troca substitui o originalmente presente entre eles (Figura 3B).
- Projete primers de oligonucleotídeos para amplificar a junção insercional em ambos os lados do lócus de integração.
- Integração única: Projetar pares de primer oligonucleotídeos que se estendem pelos locais attR e/ou attL . Um primer deve vincular-se à construção de acoplamento previamente integrada e o outro ao transgene recém-integrado (Figura 3A).
- RMCE: A substituição do pode ocorrer em duas orientações diferentes em relação ao cromossomo (designado A e B). Projete combinações alternativas de 4 primers de oligonucleotídeos para dar um produto discreto em apenas uma das orientações, com um par sendo diagnóstico para orientação A, e o outro para orientação B (Figura 3B, Figura 6).
- Extrair DNA genômico de indivíduos positivos do G2 e realizar o diagnóstico de ELETROforese pcr e gel para visualizar a presença de amplicons diagnósticos esperados a partir dos mapas previstos do local de integração.
NOTA: O DNA pode ser extraído de casos de pernas de um adulto solteiro46 ou pupal (comunicação pessoal L. Grigoraki). - Sequenciar produtos PCR para confirmar sequências esperadas.

Figura 2. Diagrama de fluxo de trabalho para modificação do genoma φC31 direcionada ao local em mosquitos Anopheles . Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3. Base molecular de integração única mediada por φC31 (A) e RMCE (B). A) Mapas esquemáticos da inserção genômica em uma linha de acoplamento An. stephensi (80.9, Tabela 1) carregando um único local attP e marcado com CFP (topo), um plasmídeo de design de um único attB marcado com DsRed (meio), e o local de inserção esperado resultando após integração bem sucedida (inferior). B) Mapas esquemáticos da inserção genômica em uma linha de acoplamento An. gambiae (A11, Tabela 1) carregando dois locais de attP invertidos e marcados com CFP (superior), um plasmídeo de design de duplo attB marcado com YFP (meio), e o local de inserção esperado resultando após o sucesso de RMCE (inferior). Linha ondulada: genoma do mosquito; Flechas listradas: braços transposon piggyBac; 3xP3: promotor do marcador fluorescente; SV40: exterminador viral; Ori: origem da replicação; AmpR: gene de resistência à ampicilina. As linhas de travessia representam o local de recombinação entre os locais attP e attB. As setas pretas numeradas representam locais de ligação primer para a validação molecular do lócus de inserção (etapa 5 do protocolo). Plasmids totalmente anotados com marca attB estão disponíveis nos autores mediante solicitação. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Resultados
O protocolo ilustrado aqui permite gerar uma linha transgênica Anopheles estável em ~10 semanas (assumindo um ciclo de vida de mosquito de 21 dias).
Espera-se que as taxas de eclosão de larvas pós-injeção em An. gambiae sejam geralmente mais baixas que a de An. stephensi, porém as taxas de incubação entre 10-50% foram relatadas9,20,24,26,33,43,47. Dada a técnica de injeção adequada, as taxas de eclosão de ≥20% são geralmente suficientes para produzir transformadores. A captação de DNA pelos embriões pode ser avaliada pela triagem de larvas jovens para expressão transitória do marcador fluorescente. Em experimentos de RMCE bem sucedidos em An. gambiae usando o promotor 3xP3 até 50% das larvas G0 sobreviventes mostraram expressão episômica do marcador na papila anal48.
Estimativas generalizadas de eficiência de transformação são difíceis de avaliar entre laboratórios e até mesmo entre experimentos, pois a transformação depende de uma complexa interação de variáveis, incluindo pureza, concentração, tamanho e toxicidade potencial do DNA injetado, qualidade dos ovos, manuseio pré e pós-injeção de ovos, criação de mosquitos e, mais importante, a experiência do operador. Foram obtidas taxas de transformação de até 7% para RMCE em An. gambiae (calculada como o número de eventos de transformação independente no total de G0 indivíduos)9,26,33, e até 2,2% de taxa de transformação para integração em An. stephensi. Sugerimos injetar pelo menos 500 embriões, o que deve levar à eclosão de pelo menos 100 G0 de larvas e a 2-7 G0 fundadores adultos dos quais a prole transformada a facada pode ser obtida. Se a triagem para expressão transitória em larvas G0, até 40 larvas positivas podem ser esperadas.
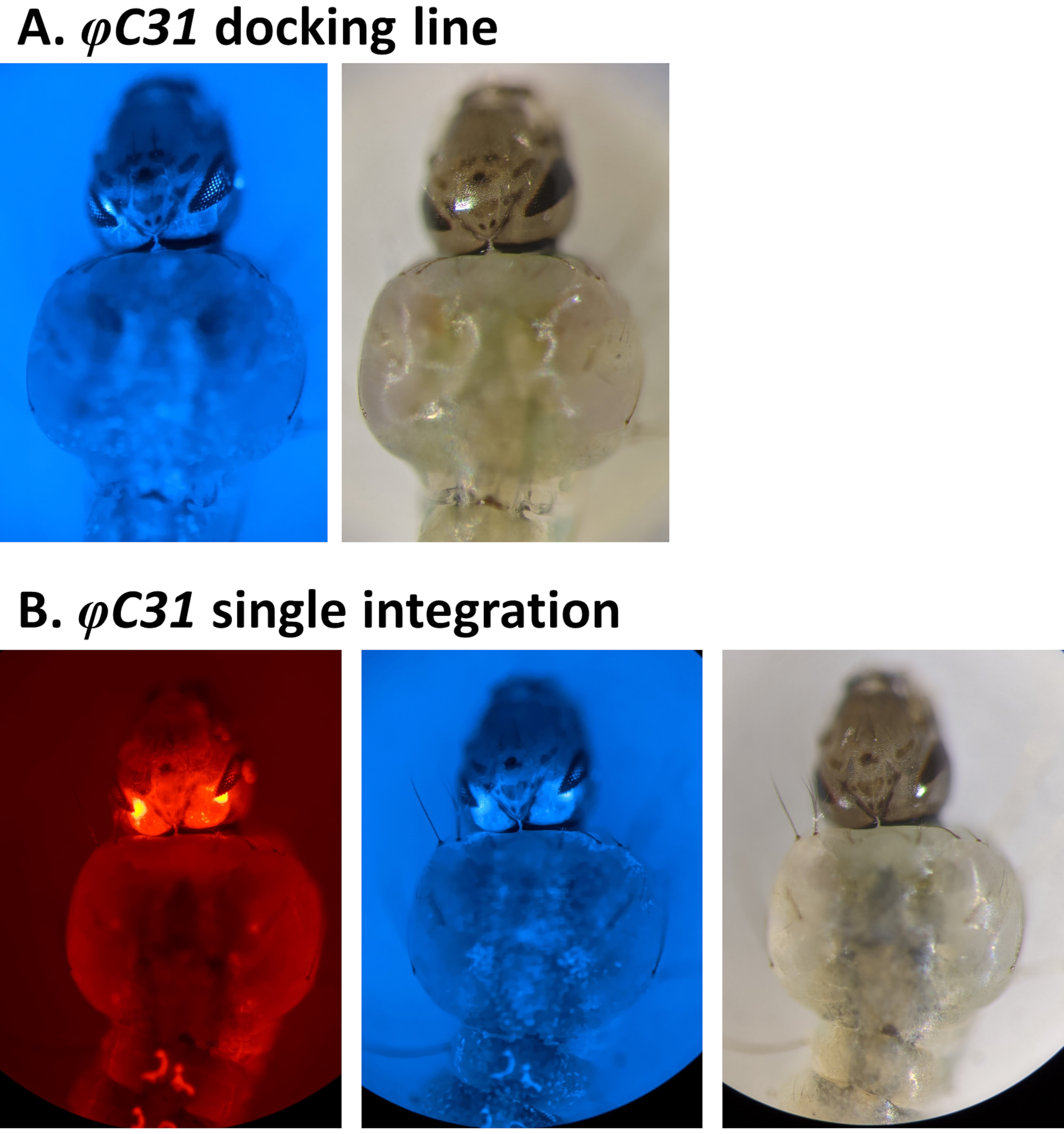
Exemplos de validação fenotípica da transformação através da triagem de marcadores fluorescentes regulados pelo promotor 3xP3 são relatados na Figura 4 e NaFigure 5. A Figura 4 mostra uma nova linha An. stephensi obtida pela inserção de um marcado por DsRed em uma linha de acoplamento marcada com CFP (80.9, Tabela 1), resultando em prole G1 expressando ambos os marcadores como indicado pela fluorescência vermelha e azul detectada nos olhos.
Espera-se que os projetos RMCE resultem na substituição do marcador originalmente inserido na linha de acoplamento com o do plasmídeo doador. Figura 5A eFigure B ilustram esta troca de marcadores em uma linha de acoplamento An. gambiae marcada com CFP (A11, Tabela 1) onde após o sucesso RMCE o marcador CFP é perdido e o marcador YFP é adquirido resultando em fluorescência amarela (mas não azul) nos olhos e cordões nervosos33. Ocasionalmente, o RMCE pode resultar em um único evento de integração em vez da troca do transgênico desejado como ilustrado na Figura 5C, onde uma larva marcada com o CFP original e os novos marcadores YFP é mostrada. Relata-se que até 50% do total de eventos de transformação são integrações únicas9,33.
Ao analisar a presença de um marcador fluorescente é crucial distinguir seu sinal de uma possível autofluorescência de fundo. Isso é particularmente importante quando o uso de CFP como larvas anofeeles exibem autofluorescência azul natural (Figura 6A). Aumentar a ampliação e focar nos tecidos e órgãos onde se espera que a fluorescência seja impulsionada pelo promotor é necessário para identificar indivíduos verdadeiros cfp positivos como ilustrado na Figura 6B usando o marcador 3xP3-CFP.
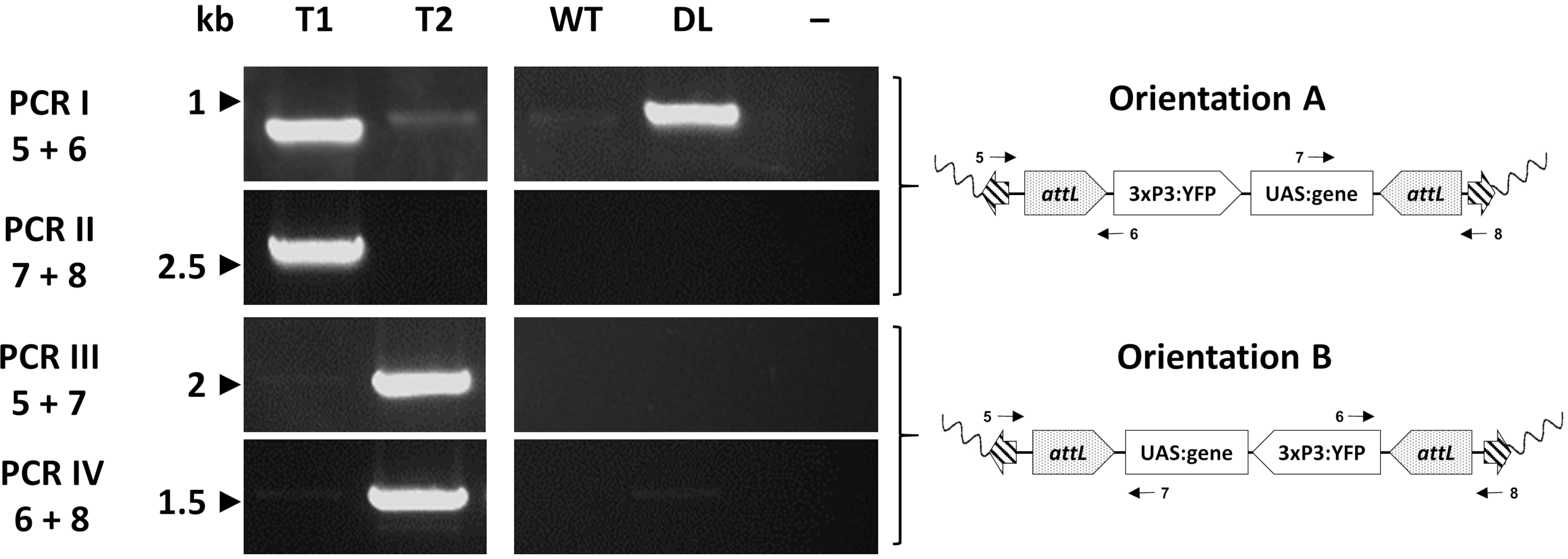
Transformadores individuais são finalmente avaliados molecularmente via PCR para confirmar o local de inserção esperado. Figura 7 relata a validação do PCR em indivíduos de uma linha de troca An. gambiae mostrando as duas orientações potenciais de inserção no genoma do mosquito33.

Figura 4. Validação da integração única de φC31 em larvas an. stephensi (visão dorsal). A) A linha de acoplamento (80,9, Tabela 1) expressa CFP aos olhos sob a regulamentação do promotor 3xP3 . B) A integração bem-sucedida resulta na expressão do DsRed recém-adquirido, bem como no marcador CFP original nos olhos. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5. Validação de φC31 RMCE em An. larvas de gambias (visão ventral). A) A linha de acoplamento (A10, Tabela 1) expressa CFP sob a regulamentação do promotor 3xP3 nos olhos (e) e do cabo nervoso (nc)5. B) O RMCE bem-sucedido resulta na troca de marcador fluorescente de CFP para YFP33. C) O único evento de integração ocorreu durante o experimento RMCE, onde a larva transformadora expressa os marcadores CFP e YFP. Esta larva carrega componentes GAL4/UAS que causam um amplo padrão de expressão de YFP, particularmente forte nos músculos abdominais (am). Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 6. Autofluorescência CFP em An. larvas de gambiae (visão dorsal). A) Imagem lado a lado de uma larva L4 positiva (CFP+) e uma larva L4 negativa (CFP-) utilizando o filtro CFP. B) Imagem de close-up dos olhos larvais que revela um CFP+ vs CFP-individual. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 7. Validação molecular da orientação da inserção de em an. gambiae representativo criado por φC31 RMCE. O transgênico pode ser inserido em uma das duas orientações alternativas (A ou B) em relação ao local de inserção. Cada reação pcr (I - IV) usa uma combinação de primers (5-8)33 projetados para dar um fragmento de amplificação diagnóstica para cada orientação, conforme indicado nos mapas plasmídeos esquemáticos. T1: indivíduo transgênico representativo que carrega orientação de inserção A; T2: indivíduo transgênico representativo que carrega orientação de inserção B; WT: tipo selvagem; DL: linha de acoplamento; -: controle negativo de reação onde a água foi usada como modelo. Este número foi modificado a partir de Adolfi et al. (2019)33. Clique aqui para ver uma versão maior desta figura.
{kind=link}
| Espécie | Coar | Nome | attP(s) | Chromo-some | Marcador de promotor | Instituição de origem | Referência |
| An. stephensi | Indiana | 26.10b | Único | 2R | 3xP3-eCFP | Univ. da Califórnia Irvine | 25 |
| An. stephensi | Indiana | 44Cb | Único | X | 3xP3-eCFP | Univ. da Califórnia Irvine | 23, 24 |
| An. stephensi | Indiana | 80,9 bilhões | Único | 2L | 3xP3-eCFP | Univ. da Califórnia Irvine | Este estudo |
| An. gambiae | G3 | 113 | Único | 2R | 3xP3-eCFP | Univ. da Califórnia Irvine | Este estudo |
| An. gambiae | KIL | Ec | Único | 3R | 3xP3-eCFP | Keele Univ. | 19, 43 |
| An. gambiae | G3 | X1 | Único | 2L | Sem marcador | Univ. de Estrasburgo | 22 |
| An. gambiae | G3 | YAttP | Único | Y | 3xP3-RFP | Imperial College London | 21 |
| An. gambiae | G3 | A10b | Dobrar | 2R | 3xP3-eCFP | Liverpool School Trop. Med. | 5 |
| An. gambiae | G3 | A11b | Dobrar | 2R | 3xP3-eCFP | Liverpool School Trop. Med. | 9 |
| um. Cepa da Universidade Johns Hopkins (presente de M. Jacobs-Lorena) e da cultura na Univ. da Califórnia Irvine por >20 anos. | |||||||
| b. Estas linhas estão disponíveis dos autores mediante solicitação razoável. | |||||||
| c. Esta linha está disponível no repositório BEI www.beiresources.org como MRA-1163. | |||||||
Mesa 1. Linhas de acoplamento anofeeles attP .
Discussão
O design preciso de plasmídeos marcados por attB compatíveis com a linha de acoplamento de escolha é primordial para o sucesso do experimento. Deve-se considerar cuidadosamente a escolha do marcador utilizado para a triagem de transformadores, incluindo a cor da fluorescência e seu padrão de expressão, que estará sujeito ao padrão já presente na linha de acoplamento. É necessário usar marcadores fluorescentes facilmente distinguíveis: boas combinações de marcadores incluem RFP (vermelho)/CFP (ciano), RFP (vermelho)/GFP (verde), RFP (vermelho)/YFP (amarelo) e YFP (amarelo)/CFP (ciano), enquanto as combinações a evitar são YFP (amarelo)/GFP (verde) e CFP (ciano)/GFP (verde). O promotor 3xP33, específico para os olhos e o cabo nervoso, é o mais usado para conduzir a expressão de marcadores fluorescentes para transgênese de mosquitos. De fato, todas as linhas de acoplamento Anopheles atualmente disponíveis utilizam este promotor. Regiões regulatórias alternativas são as do gene an. gambiae poliubiquitin (PUBc)5 ou o promotor viral IE120, que impulsionam a expressão em múltiplos tecidos. Quando usados junto com o 3xP3, esses promotores expandiriam as possíveis combinações de cores e até mesmo o uso do mesmo fluoróforo. Os promotores indicados estão ativos durante todo o ciclo de vida do mosquito, permitindo o rastreamento e o monitoramento da fluorescência em todas as fases da vida. Uma consideração adicional durante o projeto plasmídeo é o tamanho da carga a ser integrada ou trocada. Embora o sistema φC31 tenha capacidades notáveis de transporte18, deve-se considerar que o tamanho do plasmídeo doador geralmente se correlaciona negativamente com a eficiência de transformação22.
No protocolo descrito, a fonte de integração é um plasmídeo auxiliar expressando a enzima onipresentemente40. A presença onipresente da integrase pode levar à transformação de células somáticas se as microinjeções não forem precisamente direcionadas para a área onde a germina se forma. Embora tais eventos de transformação sejam perdidos por não serem herdáveis, efeitos somáticos podem diminuir a aptidão dos indivíduos injetados. Para evitar isso e aumentar a eficiência da transformação, a expressão integrase pode ser restrita à germinação, por exemplo, utilizando o promotor vasa22,26. Outros protocolos descrevem o uso do RNA mensageiro transcrito in vitro (mRNA) como fonte de φC31 integra19,24,43. No entanto, isso envolve a preparação laboriosa do mRNA e requer um manuseio cuidadoso da mistura de injeção e o uso de reagentes livres de RNase para evitar a degradação. Fontes plasmidas de integração foram demonstradas tanto em An. gambiae9,21,22,26,33,37 quanto em An. stephensi (Comunicação Pessoal A.A.) para serem confiáveis e levarem a uma transformação eficiente, e portanto são nossa opção preferida. Outra opção para entrega integrase é sua produção in vivo em linhas auxiliares de auto-acoplamento. Tais linhas foram criadas em An. gambiae que expressam a integração φC31 sob a regulamentação dos nanos promotores específicos de germina e foram encontradas para levar a uma melhor eficiência de sobrevivência e transformação20. No entanto, devem ser consideradas cargas potenciais de condicionamento físico impostas pela produção in vivo da enzima integrase na linha auxiliar.
Como em outras técnicas transgênicas, devem ser reservados cuidados especiais à criação e cruzamento de indivíduos provenientes de embriões injetados para maximizar as chances de recuperar transformadores. Indivíduos que herdaram o transgene podem ser recuperados em primeiro lugar na prole do G1. No entanto, os primeiros sinais de transformação potencial podem ser avaliados pela presença de expressão epissomal transitória do marcador fluorescente nas papilas anais e/ou cordão nervoso de G0 primeira e segunda larvas instar ao usar o promotor 3xP343. Embora a presença de fluorescência transitória sugira o sucesso da entrega plasmida, não garante a transformação herdável da germinação. Da mesma forma, a falta de expressão transitória não exclui a transformação bem sucedida. No entanto, observou-se que indivíduos transitoriamente positivos são mais propensos a produzir progênero transgênico em comparação com os transitoriamente negativos43,48. Em mãos especializadas, a criação e o cruzamento de apenas indivíduos positivos podem ser uma opção para reduzir o número de mosquitos. No entanto, dada a importância e fragilidade das pequenas larvas G0, a menor quantidade de manipulação ainda é aconselhável e a criação de todos os indivíduos G0 é sempre recomendada.
O esquema de acasalamento relatado neste protocolo foi projetado para maximizar a chance de acasalamento e isolar eventos de transformação independentes. No entanto, se o espaço inseto ou a disponibilidade de pessoal for um problema, os adultos G0 podem ser agrupados por sexo em gaiolas individuais se forem fornecidos indivíduos de sexo oposto suficientes. Tal configuração não permitirá discriminação entre múltiplos eventos de transformação que ocorrem em indivíduos da mesma gaiola. Dependendo da configuração experimental, espera-se a presença de um marcador duplo (integração única) ou único (RMCE) durante o processo de triagem. Em experimentos de integração única é importante verificar a presença do marcador original da linha de acoplamento, enquanto na RMCE é importante verificar a perda do marcador previamente integrado. De fato, não é incomum nos projetos RMCE recuperar transformadores nos quais a integração única em vez de troca ocorreu devido à recombinação de um único site attP9,33. Nesses indivíduos, ambos os marcadores fluorescentes estão presentes, bem como toda a espinha dorsal plasmida do doador, destacando a importância de realizar uma triagem minuciosa para ambos os marcadores fluorescentes.
Enquanto a presença de padrões esperados de fluorescência indica uma transformação bem sucedida, a caracterização molecular do local de inserção deve ser realizada. Para isso, a elaboração de mapas precisos do lócus de inserção previsto, incluindo as regiões genômicas de flanqueamento da linha de acoplamento, é crucial para o projeto de primers de oligonucleotídeos diagnósticos adequados para análises de amplificação genética. Eventos de integração único resultam na formação de sítios híbridos attR e attL na junção entre o DNA recém-integrado e o anteriormente inserido. Esses sites podem ser direcionados para validação do local de inserção. Nos desenhos RMCE, a inserção do doador pode ocorrer em duas orientações alternativas em relação ao lócus genômico, assim quatro primers podem ser usados em combinações alternativas de PCR para detectar qual orientação a linha carrega. Como a orientação da inserção de pode afetar a expressão transgênica, na análise comparativa da expressão genética é importante utilizar linhas com a mesma orientação de inserção.
Ao trabalhar com baixo número de transformadores pode não ser desejável sacrificar indivíduos inteiros para análise molecular. Uma opção para isso é a realização de análises moleculares no DNA extraído das pernas de um único adulto46, pois a perda de perna não afeta uma habilidade feminina adulta de acasalar e oviposit49. No entanto, há o risco de danificar o indivíduo no processo de remoção da perna. O sucesso foi obtido usando casos de pupal descartados (comunicação pessoal L. Grigoraki), no entanto, a abordagem mais segura é realizar análise molecular em pais G2 após obter prole G3 viável.
Nos últimos anos, o CRISPR/Cas9 revolucionou a maneira de realizar a edição de genomas específicos do local26,41,50,51. Ao contrário do RMCE dirigido pelo site, as integrações genéticas mediadas pelo CRISPR/Cas9 (knock-ins) são independentes da presença de locais de recombinação pré-inseridos com apenas um evento de transformação de uma etapa necessário. No entanto, o sistema CRISPR/Cas9 conta com a presença de grandes sequências genômicas conhecidas flanqueando o local de inserção desejado para reparo direcionado à hologia bem sucedida, bem como no eficiente reconhecimento do local mediado pelo guia RNAs. Essas condições nem sempre podem ser atendidas ou podem ser laboriosas para solucionar problemas e, dada a disponibilidade de múltiplas linhas de acoplamento em An. gambiae e An. stephensi e linhas derivadas delas, o sistema φC31 continua sendo uma ferramenta muito valiosa para realizar comparações fenotípicas diretas entre transgenes nos mesmos locais genômicos.
Divulgações
Os autores não têm nada a revelar.
Agradecimentos
Somos gratos a Kiona Parker (UCI) por fornecer imagens de larvas transgênicas An. stephensi , e a Fraser Colman (LSTM) e Beth Poulton (LSTM) por fornecer larvas transgênicas an. gambiae . Beth Poulton (LSTM) também prestou assistência preciosa durante a imagem de larvas de An. gambiae . Este trabalho foi financiado pelo Instituto Tata de Genética e Sociedade (TIGS) e pelo Fundo Catalyst diretor do LSTM concedido à A.A. (DCF2014AA). A.A.J. é professor donald bren na Universidade da Califórnia, Irvine.
Materiais
| Name | Company | Catalog Number | Comments |
| 1.5 mL eppendorf tubes | |||
| 8-well microslides | VWR | MARI1216690 | |
| DNeasy Blood & Tissue Kit | Qiagen | 69504 | |
| EndoFree Plasmid Maxi Kit (10) | Qiagen | 12362 | |
| Ethanol, Absolute, Molecular Biology Grade | |||
| Filter set CFP for Leica MZ FLIII Excitation 436/20 nm, extinction 480/40 nm | Leica | 10446363 | |
| Filter set dsRED for Leica MZ FLIII Excitation 545/30 nm, extinction 620/60 nm | Leica | 10447079 | |
| Filter set YFP customised for Leica MZ FLIII | Omega Optical | 500QM25, 500QM35 | |
| Halocarbon oil 27 | Sigma | H8773 | |
| Halocarbon oil 700 | Sigma | H8898 | |
| Petri dishes | |||
| Potassium chloride | |||
| Sodium Chloride | |||
| Sodium phosphate dibasic | |||
| Sodium Acetate Solution (3 M), pH 5.2 | Thermo Fisher Scientific (Life Technologies) | R1181 | |
| Stable brush Size 0 |
Referências
- Adolfi, A., Lycett, G. J. Opening the toolkit for genetic analysis and control of Anopheles mosquito vectors. Current Opinion in Insect Science. 30, 8-18 (2018).
- Grossman, G. L., Rafferty, C. S., Clayton, J. R., Stevens, T. K., Mukabayire, O., Benedict, M. Q. Germline transformation of the malaria vector, Anopheles gambiae, with the piggyBac transposable element. Insect Molecular Biology. 10 (6), 597-604 (2001).
- Nolan, T., Bower, T. M., Brown, A. E., Crisanti, A., Catteruccia, F. piggyBac-mediated germline transformation of the malaria mosquito Anopheles stephensi using the red fluorescent protein dsRED as a selectable marker. Journal of Biological Chemistry. 277 (11), 8759-8762 (2002).
- Perera, O. P., Harrell, R. A., Handler, A. M. Germ-line transformation of the South American malaria vector, Anopheles albimanus, with a piggyBac/EGFP transposon vector is routine and highly efficient. Insect Molecular Biology. 11 (4), 291-297 (2002).
- Adolfi, A., Pondeville, E., Lynd, A., Bourgouin, C., Lycett, G. J. Multi-tissue GAL4-mediated gene expression in all Anopheles gambiae life stages using an endogenous polyubiquitin promoter. Insect Biochemistry and Molecular Biology. 96, 1-9 (2018).
- Carballar-Lejarazú, R., Jasinskiene, N., James, A. Exogenous gypsy insulator sequences modulate transgene expression in the malaria vector mosquito, Anopheles stephensi. Proceedings of the National Academy of Sciences of the United States of America. 110 (18), 7176-7181 (2013).
- Galizi, R., et al. A synthetic sex ratio distortion system for the control of the human malaria mosquito. Nature Communications. 5, 3977 (2014).
- Nolan, T., Petris, E., Müller, H. M., Cronin, A., Catteruccia, F., Crisanti, A. Analysis of two novel midgut-specific promoters driving transgene expression in Anopheles stephensi mosquitoes. PLoS ONE. 6 (2), 16471 (2011).
- Lynd, A., Balabanidou, V., Vontas, J., Lycett, G. J. Development of a functional genetic tool for Anopheles gambiae oenocyte characterisation: appliction to cuticular hydrocarbon synthesis. BioRxiv. , (2019).
- O'Brochta, D. A., Alford, R. T., Pilitt, K. L., Aluvihare, C. U., Harrell, R. A., Harrell, R. A. piggyBac transposon remobilization and enhancer detection in Anopheles mosquitoes. Proceedings of the National Academy of Sciences of the United States of America. 108 (39), 16339-16344 (2011).
- O'Brochta, D. A., Pilitt, K. L., Harrell, R. A., Aluvihare, C., Alford, R. T. Gal4-based enhancer-trapping in the malaria mosquito Anopheles stephensi. G3. 2 (11), 1305-1315 (2012).
- Macias, V. M., et al. nanos-Driven expression of piggyBac transposase induces mobilization of a synthetic autonomous transposon in the malaria vector mosquito, Anopheles stephensi. Insect Biochemistry and Molecular Biology. 87, 81-89 (2017).
- Nimmo, D. D., Alphey, L., Meredith, J. M., Eggleston, P. High efficiency site-specific genetic engineering of the mosquito genome. Insect Molecular Biology. 15 (2), 129-136 (2006).
- Kim, A., Pyykko, I. Size matters: Versatile use of PiggyBac transposons as a genetic manipulation tool. Molecular and Cellular Biochemistry. 354, 301-309 (2011).
- Thorpe, H. M., Smith, M. C. M. In vitro site-specific integration of bacteriophage DNA catalyzed by a recombinase of the resolvase/invertase family. Proceedings of the National Academy of Sciences of the United States of America. 95 (10), 5505-5510 (1998).
- Khaleel, T., Younger, E., Mcewan, A. R., Varghese, A. S., Smith, M. C. M. A phage protein that binds φC31 integrase to switch its directionality. Molecular Microbiology. 80 (6), 1450-1463 (2011).
- Farruggio, A. P., Chavez, C. L., Mikell, C. L., Calos, M. P. Efficient reversal of phiC31 integrase recombination in mammalian cells. Biotechnology Journal. 7 (11), 1332-1336 (2012).
- Venken, K. J. T., He, Y., Hoskins, R. A., Bellen, H. J. P[acman]: A BAC Transgenic Platform for Targeted Insertion of Large DNA Fragments in D. melanogaster. Science. 314 (5806), 1747-1751 (2006).
- Meredith, J. M., et al. Site-specific integration and expression of an anti-malarial gene in transgenic Anopheles gambiae significantly reduces Plasmodium infections. PLoS ONE. 6 (1), 14587 (2011).
- Meredith, J. M., Underhill, A., McArthur, C. C., Eggleston, P. Next-Generation Site-Directed Transgenesis in the Malaria Vector Mosquito Anopheles gambiae: Self-Docking Strains Expressing Germline-Specific phiC31 Integrase. PLoS ONE. 8 (3), 59264 (2013).
- Bernardini, F., et al. Site-specific genetic engineering of the Anopheles gambiae Y chromosome. Proceedings of the National Academy of Sciences of the United States of America. 111 (21), 7600-7605 (2014).
- Volohonsky, G., et al. Tools for Anopheles gambiae Transgenesis. G3. 5 (6), 1151-1163 (2015).
- Amenya, D. A., et al. Comparative fitness assessment of Anopheles stephensi transgenic lines receptive to site-specific integration. Insect Molecular Biology. 19 (2), 263-269 (2010).
- Isaacs, A. T., et al. Transgenic Anopheles stephensi coexpressing single-chain antibodies resist Plasmodium falciparum development. Proceedings of the National Academy of Sciences of the United States of America. 109 (28), 1922-1930 (2012).
- Pham, T. B., et al. Experimental population modification of the malaria vector mosquito, Anopheles stephensi. PLoS Genetics. 15 (12), 1008440 (2019).
- Hammond, A., et al. A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nature Biotechnology. 34 (1), 78-83 (2015).
- Groth, A. C., Fish, M., Nusse, R., Calos, M. P. Construction of Transgenic Drosophila by Using the Site-Specific Integrase from Phage phiC31. Genetics. 166 (4), 1775-1782 (2004).
- Franz, A. W. E., et al. Comparison of transgene expression in Aedes aegypti generated by mariner Mos1 transposition and site-directed recombination. Insect Molecular Biology. 20 (5), 587-598 (2011).
- Labbé, G., Nimmo, D., Alphey, L. piggybac-and PhiC31-Mediated Genetic Transformation of the Asian Tiger Mosquito, Aedes albopictus (Skuse). PLoS Neglected Tropical Diseases. 4 (8), 788 (2010).
- Schetelig, M. F., Scolaric, F., Handler, A. M., Kittelmann, S., Gasperi, G., Wimmer, E. A. Site-specific recombination for the modification of transgenic strains of the Mediterranean fruit fly Ceratitis capitata. Proceedings of the National Academy of Sciences of the United States of America. 106 (43), 18171-18176 (2009).
- Yonemura, N., et al. phiC31-integrase-mediated, site-specific integration of transgenes in the silkworm, Bombyx mori (Lepidoptera: Bombycidae). Applied Entomology and Zoology. 43 (11), 997-1008 (2013).
- Bateman, J. R., Lee, A. M., Wu, C. T. Site-specific transformation of Drosophila via phiC31 integrase-mediated cassette exchange. Genetics. 173 (2), 769-777 (2006).
- Adolfi, A., Poulton, B., Anthousi, A., Macilwee, S., Ranson, H., Lycett, G. J. Functional genetic validation of key genes conferring insecticide resistance in the major African malaria vector, Anopheles gambiae. Proceedings of the National Academy of Sciences of the United States of America. 116 (51), 25764-25772 (2019).
- Haghighat-Khah, R. E., et al. Site-specific cassette exchange systems in the aedes aegypti mosquito and the Plutella xylostella moth. PLoS ONE. 10 (4), 0121097 (2015).
- Long, D., Lu, W., Zhang, Y., Bi, L., Xiang, Z., Zhao, A. An efficient strategy for producing a stable, replaceable, highly efficient transgene expression system in silkworm, Bombyx mori. Scientific Reports. 5 (1), 8802 (2015).
- Volohonsky, G., et al. Transgenic Expression of the Anti-parasitic Factor TEP1 in the Malaria Mosquito Anopheles gambiae. PLoS Pathogens. 13 (1), 1006113 (2017).
- Grigoraki, L., Grau-Bové, X., Yates, H. C., Lycett, G. J., Ranson, H. Isolation and transcriptomic analysis of anopheles gambiae oenocytes enables the delineation of hydrocarbon biosynthesis. eLife. 9, 58019 (2020).
- Kyrou, K., et al. A CRISPR-Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nature Biotechnology. 36 (11), 1062-1066 (2018).
- Berghammer, A. J., Klingler, M., Wimmer, E. A. A universal marker for transgenic insects. Nature. 402 (6760), 370-371 (1999).
- Ringrose, L. Transgenesis in Drosophila melanogaster. Methods in Molecular Biology. 561, 3-19 (2009).
- Dong, Y., Simões, M. L., Marois, E., Dimopoulos, G. CRISPR/Cas9 -mediated gene knockout of Anopheles gambiae FREP1 suppresses malaria parasite infection. PLoS Pathogens. 14 (3), 1006898 (2018).
- Lombardo, F., Lycett, G. J., Lanfrancotti, A., Coluzzi, M., Arcà, B. Analysis of apyrase 5' upstream region validates improved Anopheles gambiae transformation technique. BMC research notes. 2, 24 (2009).
- Pondeville, E., et al. Efficient ΦC31 integrase-mediated site-specific germline transformation of Anopheles gambiae. Nature Protocols. 9 (7), 1698-1712 (2014).
- Lobo, N. F., Clayton, J. R., Fraser, M. J., Kafatos, F. C., Collins, F. H. High efficiency germ-line transformation of mosquitoes. Nature protocols. 1 (3), 1312-1317 (2006).
- Terenius, O., Juhn, J., James, A. A. Injection of An. stephensi embryos to generate malaria-resistant mosquitoes. Journal of Visualized Experiments. 5, 216 (2007).
- Lynd, A., et al. Insecticide resistance in Anopheles gambiae from the northern Democratic Republic of Congo, with extreme knockdown resistance (kdr) mutation frequencies revealed by a new diagnostic assay. Malaria Journal. 17 (1), 412 (2018).
- Marinotti, O., et al. Development of a population suppression strain of the human malaria vector mosquito, Anopheles stephensi. Malaria Journal. 12 (1), 142 (2013).
- Adolfi, A. In vivo functional genetic analysis of insecticide resistance in the malaria mosquito Anopheles gambiae. University of Liverpool. , (2017).
- Isaacs, A. T., Lynd, A., Donnelly, M. J. Insecticide-induced leg loss does not eliminate biting and reproduction in Anopheles gambiae mosquitoes. Scientific Reports. 7, 46674 (2017).
- Gantz, V. M., et al. Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. Proceedings of the National Academy of Sciences of the United States of America. 112 (49), 6736-6743 (2015).
- Li, M., Akbari, O. S., White, B. J. Highly efficient site-specific mutagenesis in malaria mosquitoes using CRISPR. G3: Genes, Genomes, Genetics. 8 (2), 653-658 (2018).
- Poulton, B. C., et al. Using the GAL4-UAS System for Functional Genetics in Anopheles gambiae. J. Vis. Exp. , (2021).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados