
Method Article
Lagria villosa Böcekleri burkholderia gladioli Symbiont Bakteriyel Kolonizasyon Faktörlerini Elucidate için Bir Araç Olarak Transposon-ekleme Dizileme
Bu Makalede
Özet
Bu, Burkholderia faydalı bir symbiont'taki aday böcek kolonizasyon faktörlerini tanımlamak için uyarlanmış bir yöntemdir. Böcek konağı transposon mutajensis yoluyla oluşturulan rastgele bir mutant kütüphanesi ile enfektedir ve kolonileşmeden sonra kütüphane karmaşıklığı in vitroyetiştirilen bir kontrol ile karşılaştırılır.
Özet
Genlerin aktivitelerini manipüle ederek işlevini çıkarıcı olmak, çoğu biyolojik işlemin genetik temellerini anlamak için önemli bir araçtır. Moleküler mikrobiyolojideki gelişmeler, genlerin manipülasyonu için çeşitli mutajensis tekniklerinin ortaya çıkışını görmüştür. Bunlar arasında, transposon ekleme dizilimi (Tn-seq), birçok aday genin işlevselliğini aynı anda hedefsiz bir şekilde değerlendirmek için değerli bir araçtır. Teknik, çeşitli patojenik mikroplarda ve birkaç faydalı simpiontlarda ökaryotik konakların kolonizasyonu için moleküler mekanizmaları tanımlamak için anahtar olmuştur.
Burada, Tn-seq, lagria villosaböceğinin karşılıklı burkholderia gladioli ortakyaşamında kolonileşme faktörlerini tanımlamak için bir yöntem olarak kurulmuştur. Konjugasyon ile, Tn5 transposon aracılı antibiyotik direnci kasetinin yerleştirilmesi B. gladioli'derastgele genomik yerlerde gerçekleştirilir. Gen bozulmalarının bakterilerin böcek konağını kolonileştirme yeteneği üzerindeki etkisini tanımlamak için, üretilen B. gladioli transposon-mutant kütüphanesi böcek yumurtaları üzerinde aşılanırken, sıvı kültür ortamında bir kontrol in vitro olarak yetiştirilir. Kolonizasyon için yeterli zaman sağladıktan sonra, in vivo ve in vitro yetiştirilen kütüphanelerden DNA çıkarılır. DNA kütüphanesi hazırlama protokolünün ardından, DNA örnekleri transposon ekleme dizilimi için hazırlanır. Transposon-insert kenarını ve yan bakteriyel DNA'yı içeren DNA parçaları seçilir ve mutasyon bölgeleri transposon-insert kenarından uzağa dizilerek belirlenir. Son olarak, in vivo ve in vitro kütüphaneler arasındaki her bir mutantın frekanslarını analiz ederek ve karşılaştırarak, böcek kolonizasyonu sırasında spesifik symbiont genlerinin önemi tahmin edilebilir.
Giriş
Burkholderia gladioli, Lagria villosa böcekleri ile simbiyotik bir ilişkiye girebilir ve böcekkonağınınmikrobiyal antagonistlerine karşı savunmada önemli bir rol oynar 4,5,6. Dişi böcekler, üreme sistemine özel bez aksesuarında B. gladioli'nin çeşitli suşlarını barındırır. Yumurtlama üzerine dişiler, B. gladioli tarafından üretilen antimikrobiyal bileşiklerin entomopatojenik mantarlar tarafından enfeksiyonları inhibe ettiği yumurta yüzeyinde B. gladioli hücrelerini lekeler4,6. Geç embriyonik gelişim sırasında veya larvalar yumurtadan çıktıktan sonra, bakteri larvaların dorsal yüzeyindeki kbüler invaginasyonları kolonizasyona eder. Bu özel lokalizasyona ve symbiontların dikey iletim rotasına rağmen, L. villosa muhtemelen B. gladioli'yi ortamdan yatay olarak da edinebilir4. Ayrıca, L. villosa4,6ile ilişkili olarak en az üç B. gladioli türü bulunmuştur. Bunlar arasında, B. gladioli Lv-StA in vitroekimi için uygun olan tek kişidir.
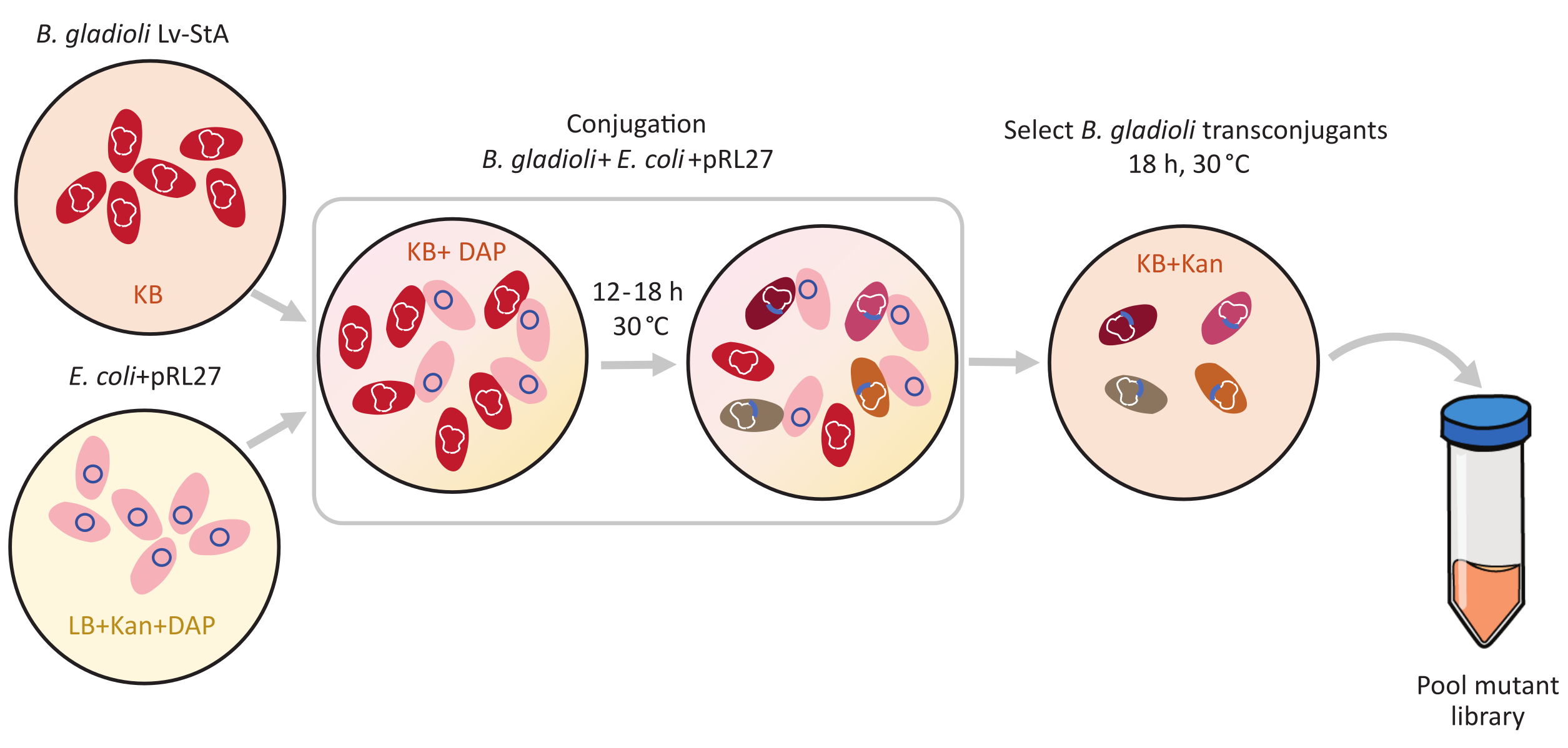
B. gladioli Lv-StA 8.56 Mb 6 genom boyutuna sahiptir ve 7.468 gen içerir. Bu genlerden hangisi B. gladioli bakterilerinin böcek konağını kolonileştirmesi için önemlidir? Busoruyucevaplamak için, koşullu olarak gerekli mikrobiyal genleri tanımlamak için keşif yöntemi olan transposon ekleme dizilimlerini (Tn-seq) kullandık 1,2,3. Bir Tn5 transposon kullanılarak B. gladioli Lv-StA'dan oluşan bir mutant kütüphanesi oluşturuldu. Escherichia coli donör hücrelerinden B. gladioli Lv-StA'ya konjugasyon yoluyla, Tn5 transposonunu taşıyan bir pRL27 plazmid ve ters tekrarlarla çevrili bir antibiyotik direnç kaseti aktarılmıştır (Şekil 1). Böylece, 3.736 symbiont geninin bozulmalarını ayrı ayrı taşıyan bir dizi mutant üretilmiştir (Şekil 2).
Mutant havuzu, kolonizasyon faktörlerini tanımlamak için böcek yumurtalarına bulaştı ve kontrol olarak King's B (KB) ortamında in vitro olarak da yetiştirildi. Kolonizasyon için yeterli zaman sağladıktan sonra, yumurtadan çıkmış larvalar toplandı ve DNA ekstraksiyonu için birikti. Transposon kesici ucu ve B. gladioli Lv-StA'nın yan genomik bölgesini içeren DNA parçaları, dizileme için değiştirilmiş bir DNA kütüphanesi hazırlama protokolü kullanılarak seçildi. B. gladioli Lv-StA'nın yumurta yüzeyi üzerinden bulaştığında L. villosa larvalarını kolonileştirmesi için çok önemli olan belirli genleri tanımlamak için DESeq2 ile analizlerin ardından kalite işlemeyi okuyun.
Protokol
1. Medya ve tampon hazırlama
- Tablo 1'deverilen KB ve LB ortam ve agar plakalarını hazırlayın ve 121 °C, 15 psi, 20 dk'da otoklavlayın.
- E.coli WM3064 + pRL27'yi kültlemeden önce otomatik kapatılmış LB ortamına 50 μg/mL filtre sterilize kanamycin ve 300 μM filtre sterilize edilmiş 2,6-diaminopimelik asit (DAP) ekleyin.
- Başarılı B. gladioli Lv-StA transkonjugantlarını seçmek için gerekli plakaları dökmek için otomatik kapatılmış KB agar'a 50 μg/mL filtre sterilize kanamycin ekleyin.
- Aşağıdaki bileşenleri karıştırarak 1x fosfat tamponlu salin (PBS) hazırlayın: NaCl 8 g/L, KCl 0.201 g/L, Na2HPO4 1.42 g/L ve KH2PO4 0.272 g/L. Tuzları damıtılmış suda çözün ve karışımı kullanımdan önce 121 °C, 15 psi, 20 dk'da otoklavlayın. Oda sıcaklığında saklayın.
- Aşağıdaki bileşenleri çözerek 2x Bind ve yıkama tamponu hazırlayın: damıtılmış suda 10 mM Tris-HCl (pH 7.5), 1 mM etilenediamin tetraasetik asit (EDTA) ve 2 M NaCl. Kullanmadan önce karışımı filtreleyin. Oda sıcaklığında saklayın.
- 10 mM Tris-HCl (pH 8.0) ve 0.1 mM EDTA'yı çift damıtılmış suda eriterek 1x Düşük TE hazırlayın. 121 °C, 15 psi, 20 dk'da otoklavlayarak sterilize edin. Oda sıcaklığında saklayın.
2. Transposon mutant kütüphanesini oluşturmak için konjugasyon

Şekil 1: Konjugasyon protokolü adımları. Konjugasyon alıcısı Burkholderia gladioli Lv-StA (kırmızı) ve pRL27 plazmidini (pembe) içeren donör Escherichia coli, kanamycin ve DAP ile desteklenmiş sırasıyla KB agar ve LB'de yetiştirilir. Plazmidin 30 °C'de 12-18 saat konjugatif transferden sonra, transkonjugant B. gladioli hücreleri KANAMYCIN içeren KB'de seçilir ve bir araya getirilir. Kısaltmalar: DAP = 2,6-diaminopimelik asit; Kan = kanamycin. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
- Steril bir davlumbaz altında, kanamycin ve DAP ile desteklenmiş 10 mL LB ortamda Escherichia coli WM3064 + pRL27'nin taze bir donör kültürünü aşıla. Burkholderia gladioli Lv-StA alıcı hücrelerini 5 mL KB ortamda aşıla. Kültürleri bir gecede 30 °C'de 250 rpm'de bir çalkalayıcıda kuluçkaya yatırın.
- Bir gecede büyümeden sonra, hücreleri peletmek için 6 dakika boyunca 9.600 × g'da kültürlerin her birinin 4 mL'sini santrifüj edin. Üstnatant atın.
- Steril bir davlumbaz altında, peletlenmiş hücre kültürlerini DAP içeren KB ortamında yıkayın ve son olarak kültürleri 4 mL KB + DAP ortamında ayrı ayrı yeniden diriltin.
- Taze bir 15 mL tüpte, yıkanmış E. coli donör hücrelerinin 250 μL'sini yıkanmış B. gladioli Lv-StA alıcı hücrelerinin 1 mL'si ile karıştırın.
- Bu konjugasyon hücresi karışımının 10 μL'yi DAP içeren KB agar plakalarında tespit edin. Plakanın steril kaputta 1 saat boyunca oda sıcaklığında bozulmadan dinlenmesine izin verin. Daha sonra, plakaları 30 °C'de 12-18 saat boyunca konjugasyon lekeleriyle kuluçkaya yatırın.
NOT: Konjugasyon süresi hedef türlere göre ayarlanabilir. Bununla birlikte, uzun bir konjugasyon süresi, genoma çift ekleme veya plazmid entegrasyonu riskini arttırır. Yavaş büyüyen bakteriler için daha uzun konjugasyon sürelerine izin verin. - Kuluçkadan sonra, steril bir kaputun altındaki plakalara 2-4 mL 1x PBS ekleyin ve büyüyen bakteriyel konjugasyon noktalarını agardan serbest bırakmak için bir hücre kazıyıcı kullanın. Konjuge hücre karışımını 2 mL mikrofuge tüplerine pipetlayın.
- Hücreleri 2 dakika boyunca 9.600 × g'da santrifüjleme yaparak pelet. Süpernatantı atın ve peleti yukarı ve aşağı pipetleme yaparak 1 mL 1x PBS'de iki kez yıkayın. Son peleti 1200 μL 1x PBS'de yeniden sunun. Karışımdaki hücre sayısı 1 × 104'ünüzerindeyse kaplamadan önce seyreltmeler yapın.
- İyice karıştırın ve kanamycin ile desteklenmiş büyük KB agar plakaları (gerekirse 6 veya daha fazla) üzerine 200 μL hücre karışımını yayın ve gece boyunca 30 °C'de kuluçkaya yayın.
NOT: Hedef mutant kolonileri seçici agar plakalarında 30 saat içinde görünür. Antibiyotik direnç belirteci nedeniyle, seçici agar plakasında sadece mutasyona uğramış koloniler görülür. Bu nedenle, tüm kolonilerin başarılı transkonjuganlar olması beklenir. - Üç plaka üzerinde transkonjugant kolonilerinin toplam sayısını sayın ve tüm plakalarda elde edilen yaklaşık mutant sayısını hesaplamak için tahminde bulunma. Temsili bir kütüphane elde etme şansını artırmak için, toplam koloni sayısının genomdaki toplam gen sayısından birkaç kat daha fazla olduğundan emin olun. Konjugasyonun başarısını doğrulamak için, bölüm 3'te açıklandığı gibi 10-20 örnek koloni kullanarak ekleme kasetini hedefleyen bir PCR gerçekleştirin.
NOT: Amaç, koloni sayısının tüm genomdaki gen sayısının en az 10 kat olmasını sağlamaktır, bu durumda 75.000 > mutanttır. Bununla birlikte, tam temsili bir kütüphaneye karşılık gelecek kolonilerin sayısını doğru bir şekilde tahmin etmek genellikle zordur. Mutasyona uğrayan benzersiz genlerin sayısı bu noktada belirgin değildir, temel genlerdeki bozulmaların yakalanmadığı göz önüne alındığında, aynı gen için genellikle birden fazla farklı mutasyon bölgesi vardır ve Tn5 transposonları ile oluşturulan mutasyonlar tamamen rastgele değildir. - Steril bir davlumbazın altında, ağar üzerine 1-2 mL 1x PBS ekleyerek tabaklardan kolonileri kazıyın. Plakalardan kazınmış hücre karışımını 50 mL tüpler halinde bir araya alın. Kütüphaneyi iyice karıştırmak ve daha sonra havuza alınmış mutant kütüphanesinin 4 mL'lik kısmını birkaç kriyotüse bölmek için Vortex. Tüplere 1 mL% 70 gliserol ekleyin ve -80 ° C'de saklayın.
3. B. gladioli Lv-StA'da başarılı eklemeleri onaylamak için PCR ve jel elektroforezi
- Eklemenin varlığını onaylamak için, adım 2.9'daki seçim plakalarından tek tek mutant kolonileri seçin ve Tablo 2'delistelenen astarları kullanarak ekleme kasetini hedefleyen bir PCR gerçekleştirin. PCR ana karışımını Tablo 3'e göre hazırlayın ve Tablo 4'teaçıklandığı gibi termal çevrimde koşulları ayarlayın.
- Güçlendirilmiş DNA parçalarının beklenen 1580 bp uzunluğunda olup olmadığını kontrol etmek için PCR ürünlerini elektroforezi (250 V, 40 dk) ile% 1,6 agarose jel üzerinde çalıştırın.
4. Böcek yumurtalarında mutant havuz enfeksiyonu
- Kütüphane yıkama adımları
- Hazırlanan mutant kütüphanesinin bir aliquot'ını buzun üzerinde çözün. 2.683 × g'da 10 dakika santrifüj ve süpernatantı çıkarın. Steril bir kaputun altında, kalan herhangi bir ortamı hücrelerden çıkarmak için hücreleri 4 mL 1x PBS ile yıkayın. Hücreleri 1x PBS'nin 4 mL'sinde yeniden biriktirin.
- Bir hücre sayma odası kullanarak kitaplığın bir aliquot'undaki hücre sayısını sayın. Kütüphanenin bir bölümünü 1x PBS'de 2 ×10 6 hücre/μL'ye seyreltin.
- Gerekli ses seviyesini almadan önce tüm kütüphaneyi homojen bir şekilde karıştırmak için kütüphane aliquot'unu iyice vorteks edin.
- Yumurta debriyaj sterilizasyonu ve in vivo enfeksiyon
- Bir L. villosa yumurta debriyajı seçin. Yumurta sayısını sayın ve debriyaj 100'den fazla yumurta içeriyorsa devam edin.
- Tüm yumurta debriyajını sterilize edin.
- 200 μL% 70 etanol ekleyin ve yumurtaları 5 dakika boyunca hafifçe yıkayın. Etanolleri çıkarın ve yumurtaları iki kez otoklavlı suyla yıkayın.
- 200 μL% 12 çamaşır suyu (NaOCl) ekleyin ve yumurtaları 30 sn boyunca hafifçe yıkayın. Çamaşır suyunu hemen çıkarın ve yumurtaları 200 μL otomatik kapalı su ile üç kez tekrar yıkayın.
- Sterilize edilmiş yumurta debriyajında yıkanmış mutant kütüphanesinin 2 ×10 6 hücresini /μL'sini enfekte edin (yumurta başına 2,5 μL).
- Enfekte böcek larvaları yumurtadan çıktıktan iki gün sonra, 1,5 mL mikrofuge tüpü başına 100 2nd başlangıç larvaları toplayın ve -80 ° C'de depolayın.
-
Tüp bebek mutant kütüphanesi kontrolü
- Steril bir davlumbaz altında, kanamycin içeren 10 mL KB ortamda yıkanmış mutant kütüphanesinin 2 × 106 hücre / μL'nin 250 μL'sini aşılayın.
- 30 °C'de in vitro mutant kültürünü 20 saat kuluçkaya yatırın.
NOT: Koloniizasyon sırasında in vivo WT B. gladioli Lv-StA'nın yaklaşık nesil sayısıyla eşleşecek şekilde inkübasyon süresini hesaplayın. - 20 saat inkübasyondan sonra, in vitro mutant kültürüne eşit miktarda% 70 gliserol ekleyin ve -80 ° C'de saklayın.
5. Enfekte böcekler ve in vitro mutant kütüphanesi DNA ekstraksiyonu
NOT: DNA ekstraksiyonları, üreticinin aşağıda kısaca özetlenen protokolüne göre bir DNA ve RNA saflaştırma kiti kullanılarak gerçeklendirilmiştir.
- 1-2 mL sıvı azot ekleyerek ve bir pestle ile ezerek havuzlanmış larvaları (mikrofuge tüpü başına maksimum 4 mg) homojenize edin.
- Tüp bebekte yetişen mutant kültürlerini buzdaki gliserol stoklarından çözün. Hücre lizizden önce 10 dakika boyunca 9.600 × g'da santrifüj yaparak hücreleri pelet.
- In vitro ve in vivo örneklere 300 μL Doku ve Hücre liziz çözeltisi ekleyin. 5 μL 10 mg/mL Proteinaz K ekleyin, karışımı 60 °C'de 15 dakika kuluçkaya yatırın ve ardından 3-5 dakika boyunca buza yerleştirin.
- Lysates ve girdaplara iyice 150 μL protein çökeltme reaktifi ekleyin. 10 dakika boyunca 9.600 × g'da santrifüjleme yaparak protein kalıntılarını pelet.
- Süpernatantı 1,5 mL mikrobuj tüpüne aktarın. Süpernatanta 500 μL izopropanol ekleyin ve 1 saat veya bir gece boyunca -20 °C'de kuluçkaya yatırmadan önce tüpleri en az 40 kez hafifçe ters çevirin.
- 10 dakika boyunca 9.600 × g'da santrifüjleme yaparak çökemiş DNA'yı peletle. Süpernatant atın ve DNA peletine buz gibi% 70 etanol ekleyin.
- 5 dakika boyunca 10.000 ≥ × g'de santrifüj. Üstnatant atın ve numuneleri en az 1 saat boyunca hava kurumaya bırakın.
- 100 μL Low-TE tamponunda in vitro ve in vivo örneklerden ALıNAN DNA'yı yeniden dürt.
- Numuneleri -20 °C'de saklayın.
6. Kütüphane hazırlığının sıralanıyor
NOT: DNA kütüphanesi hazırlama protokolü ve reaktifleri, DNA kütüphanesi hazırlama kitinin üreticisi tarafından sağlanan talimatlardan uyarlanır ve değiştirilir.

Şekil 2: DNA kütüphanesi hazırlama adımlarının şeması. Kesme ve adaptör ligasyonundan sonra, değiştirilen protokol, ekleme kasetini içeren DNA parçalarını zenginleştirmek için bir streptavidin boncuk seçimi adımı içerir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
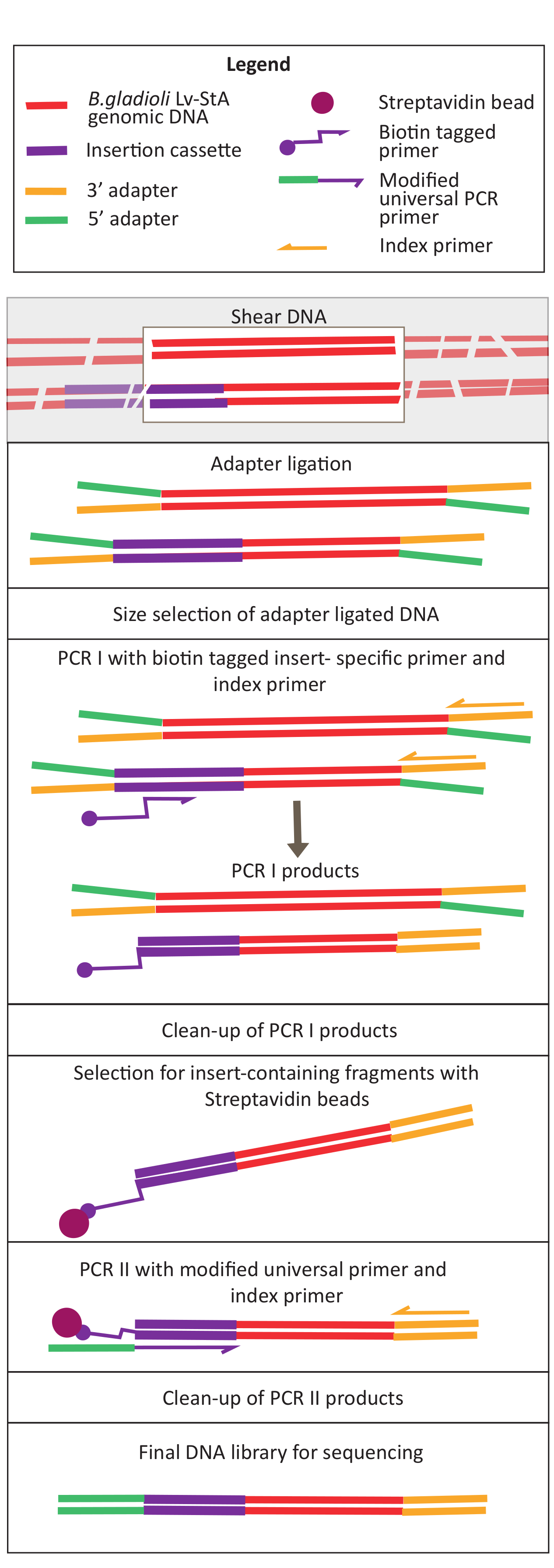{kind=link}
- Numuneleri 20 ng/μL konsantrasyona ve 100 μL hacme seyreltin ve buzda tutun.
- Ultrasonicator kullanarak in vivo ve in vitro örnek DNA'yı yamuma. Ultrasonicator'ı % 70 güçte ayarlayın. Örnekleri kısaca girdap ve 1 dakika 30 sn için makas.
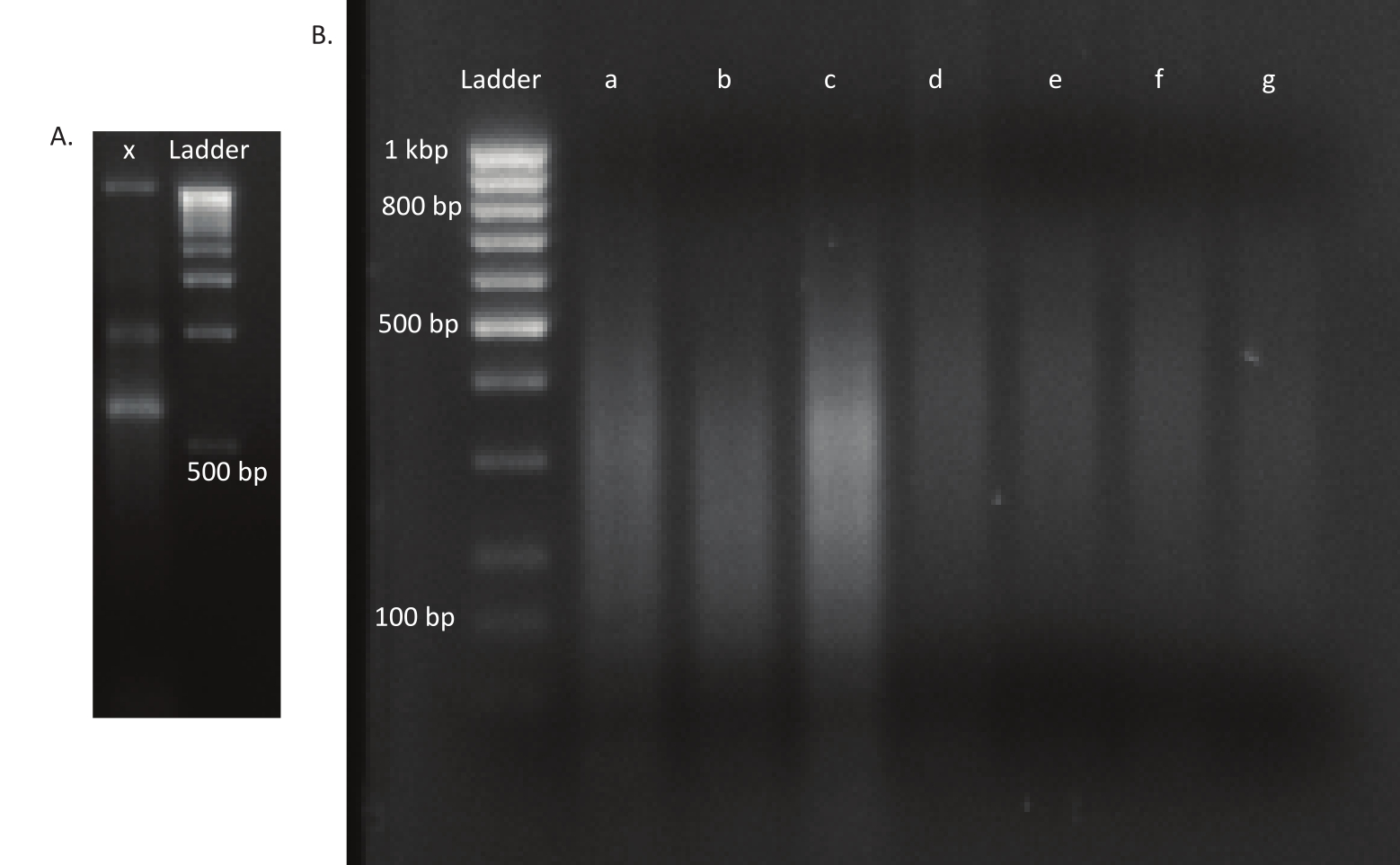
NOT: Ultrasonicator ayarları enstrümanlar arasında farklılık gösterecektir. Bu durumda, parça boyutu 200-400 bp idi, bu da 150 bp'lik bu sıralama yaklaşımı için uygundur, eşleştirilmiş uçlu (bkz. adım. 9.1). Kesme parametreleri deneycinin gereksinimlerine göre ayarlanabilir. - DNA'nın istenen boyut aralığına yamup sildiği kontrol edin (bu durumda, 200-400 bp). 40 dakika boyunca 250 V'de çalışan% 1,6 agarose jel üzerinde 1:1 oranında jel yükleme boyası ile karıştırıldıktan sonra duyulmamış ve yamulmuş DNA'nın 5 μL'siniyükleyin (Şekil 3A,B).
- Adaptör ligasyonu için gereken parça uçlarının hazırlanması
- Yamüle DNA'nın 50 μL'sine, kütüphane hazırlama kitinde verilen uç hazırlama reaktiflerini ekleyin: enzim karışımının 3 μL'si ve 7 μL reaksiyon tamponu ve pipetleme ile iyice karıştırın. 75 °C'≥ ısıtmalı kapaklı bir termal çevrimleyici ayarlayın ve numuneleri 20 °C'de 30 dakika ve 65 °C'de 30 dakika kuluçkaya yatırın. 4 °C'de tutun.
- Bağdaştırıcı ligasyonu
- Adaptör ligasyonu için, son hazırlık adımının ürünlerine aşağıdaki reaktifleri ekleyin: 30 μL Ligation Master Mix, 1 μL Ligation Enhancer ve 2,5 μL seyreltilmiş Adaptör. Pipetleme ile iyice karıştırın ve ısıtılan kapak kapalıyken termal çevrimde numuneyi 20 °C'de 15 dakika kuluçkaya yatırın.
- 15 dakika sonra, enzimin 3 μL'sini ekleyin (urasil DNA glikosylaz + DNA glikosilaz-lyaz Endonucleaz VIII) (bkz. Malzeme Tablosu). Pipetleme ile iyice karıştırın ve numuneyi 37 °C'de 15 dakika boyunca kapağı 47 °C'de ısıtılmış bir termal çevrimde ≥.
NOT: Protokol bu adımda duraklatılabilir ve numuneler -20 °C'de saklanabilir.
- 250 bp'lik adaptörle bağlı DNA hedefleme parçalarının boyut seçimi
- Manyetik boncuk çözeltisini vorteks edin (Malzeme Tablosunabakın) ve kullanmadan önce 30 dakika boyunca oda sıcaklığına yerleştirin.
- Liglenmiş DNA karışımının 96,5 μL'sine 0,3 kat boncuk ekleyin ve iyice pipetleyarak karıştırın. Boncuk karışımını 5 dakika kuluçkaya yatırın.
NOT: Boncuk karışımında tuzların ve polietilen glikolün bulunması, BONCUKlar üzerinde DNA parçalarının çökeltilini kolaylaştırır. Boncukların DNA moleküllerine oranının düşük olması, sadece daha büyük DNA parçalarının boncuklara bağlanmasına yol açar. Bu durumda, 250 bp'nin üzerindeki DNA parçaları boncuklara bağlanır. - Boncukları aşağı çekmek ve istenmeyen boyuttaki DNA parçalarını çıkarmak için tüpleri manyetik bir standa yerleştirin. Boncukların 5 dakika yerleşmesine izin verin ve ardından açık süpernatantı yeni bir mikrofuge tüpüne aktarın (süpernatantı saklayın).
- Süpernatant içine 0,15x taze boncuk ekleyin ve iyice pipetleme yaparak karıştırın. Boncuk karışımını 5 dakika kuluçkaya yatırın ve ardından hedef DNA'ya bağlı boncukları aşağı çekmek için tüpleri manyetik bir standa yerleştirin. 5 dakika bekleyin ve ardından süpernatant atın (boncukları saklayın).
NOT: Boncukların DNA'ya olan bu oranı, istenen 250 bp büyüklüğündeki parçaların bağlanmasına yol açar. - Manyetik standdaki boncuklarla, % 80 etanol 200 μL ekleyin (taze hazırlanmış) ve 30 s bekleyin. Pipet dışarı ve manyetik standdaki boncukları rahatsız etmeden etanol yıkama dikkatlice atın. Bu adımı yineleyin.
- Son yıkamadan sonra, boncuklardan etanol izlerini çıkarın ve daha sonra boncukları parlak görünene kadar 2 dakika boyunca havayla kurulayın, ancak tamamen kurumadılar. Boncukları fazla kurutmayın.
- Tüpleri manyetik standdan çıkarın ve 10 mM Tris-HCl veya 0,1x TE (Düşük TE) 17 μL ekleyin. ~10 kez pipetleme ile karıştırın ve karışımı oda sıcaklığında 2 dakika kuluçkaya yatırın.
- Tüpleri manyetik standa geri yerleştirin ve 5 dakika bekleyin. Boncuklar yerleştikten sonra DNA'nın üstünü yeni bir tüpe aktarın.
- PCR I, ekleme kasetini içeren DNA parçalarına biotin etiketi eklemek için
- Transposon spesifik biyotinilasyonlu astarı (Tablo 5) ve bir indeks astarı kullanarak Tn5 ekleme kasetini içeren DNA parçalarına biyotinillenmiş astar etiketi ekleyin. PCR ana karışımını Tablo 6'ya göre hazırlayın ve Tablo 7'delistelenen termal çevrimleyici için PCR koşullarını izleyin.
- Boyut seçimi olmadan PCR I'in temizlenmesi
- Vortex 0.9x boncukları temizleyip temizlemeden önce en az 30 dakika oda sıcaklığına yerleştirin.
- PCR ürünlerine 0,9x boncuk ekleyin ve iyice karıştırın.
- Boncukları aşağı çekmek için boncukları manyetik bir standa yerleştirin.
- Şeffaf süpernatant çıkarın ve boncuk bağlı DNA'yı 200 μL taze hazırlanmış% 80 etanol ile iki kez yıkayın.
- Yıkama adımlarından sonra etanolleri çıkarın ve boncukları parlak görünene kadar kurutun, ancak çok kurumasın.
- 32 μL 10 mM Tris-HCl veya 0,1X TE (Düşük TE) ekleyin ve boncukları 5 dakika kuluçkaya yatırın. Karışımı manyetik standa geri yerleştirin ve süpernatantı taze bir mikroyakıt tüpüne aktarın.
- Biyotinillenmiş DNA parçalarını streptavidin boncuklarına bağlama
- 1x Bind ve-wash tamponunda 32 μL streptavidin boncuklarını yeniden depola. Manyetik bir standa yerleştirilirken boncukları tamponla üç kez yıkayın.
- 32 μL 2x Bağla ve yıka tamponu ekleyin ve boncukları yeniden depoleyin. Buna, temizlenmiş PCR 1 ürünlerinin 32 μL'sini ekleyin. İyice karıştırın ve oda sıcaklığında 30 dakika kuluçkaya yatırarak.
- Boncuk-DNA karışımını 2 dakika boyunca manyetik bir standa yerleştirin. Pipet, takma kenarını içeren biotin etiketli DNA olarak süpernatantı boncuklardaki streptavidin'e bağlar.
- Boncukları 500 μL 1x Bağla ve yıka tamponu ile yıkayın ve ardından boncukları 200 μL Düşük TE ile yıkayın. DNA'ya bağlı boncukları 17 μL Düşük TE'de yeniden dirilt.
- Ekleme kaseti kenarını içeren parçalara bağdaştırıcı eklemek için PCR II
- Tablo 8'degösterildiği gibi, Tablo 5 'te listelenen dizin astarlarını ve değiştirilmiş evrensel PCR astarlarını kullanarak bir ana karışım hazırlayın. PCR karışımına önceki adımdan 15 μL DNA'ya bağlı streptavidin boncuk ekleyin. Termal çevrimleyici koşulları için Tablo 7'ye bakın.
- Bu protokolün 6.8. Son DNA ürünlerini 30 μL moleküler dereceli suda kurut.
- Numuneleri -20 °C'de saklayın ve sıralama için kullanın.
7. Sıralama ve analiz
- Yüksek aktarım hızı sıralama teknolojisini kullanarak kitaplığı sıralayın. Aşağıda belirtildiği gibi, transposon kitaplığı boyutuna bağlı olarak sıralama derinliğini ayarlayın. FastQC7ile okuma kalitesini değerlendirin. Okumanın 5' ucundaki Tn5 ekleme kenarını içeren okumaları seçin ve Cutadapt8 ve/veya Trimmomatic9kullanarak ekleme kenarı sırasını kaldırın.
NOT: Burada, okuma başına 150 bp hedeflemek için eşleştirilmiş uçlu bir sıralama yaklaşımı kullanıldı ve toplam 8 Mio okuma. Temsili bir veri kümesi elde etmek için, toplam sıralı okuma sayısının kitaplıktaki mümkün olan en fazla mutant sayısını, yani 2.9 adımındaki toplam tahmini koloni sayısını aştığından emin olun. Referans olarak, bu protokol mümkün olan en büyük kütüphane boyutunun 40 katını amaçlaydı. Benzer bir amaç için Tn-seq kullanan diğer başarılı çalışmalar, ilgili mutant kütüphanesindeki gerçek benzersiz ekleme sayısının 25 katına yakın toplam okuma sayısınısıraladı 22,23. - Genlerin uçlarındaki mutasyonların işlevsel olarak bozucu olmadığını göz önünde bulundurarak, referans genom GFF dosyasının gen ek açıklamalarının her iki ucundan% 5 indirim. Bowtie2 10 kullanarak kırpılan okumaları referans genomunaeşleyin.
- Bam hizalama dosyasındaki benzersiz 5' konum sayısından ekleme sayısını hesaplayın.
- FeatureCounts11kullanarak, her çoğaltma örneği için isabet genlerinin sayısını elde edin.
- RStudio'daki DESeq212 paketini kullanarak, farklı koşullar arasındaki mutant bolluklarındaki farkı hesaplayın.
Sonuçlar
Konakçı ile ilişkili bakteriler, yapışkanlık, hareketlilik, kemotaksi, stres yanıtları veya belirli taşıyıcılar da dahil olmak üzere bir ilişki kurmak için çeşitli faktörler kullanabilir. Patojen konak etkileşimleri için önemli faktörler rapor edilmiş olsa da, burkholderia19,20cinsinin üyeleri de dahil olmak üzere çeşitli bakteriler13, 14 ,15,16,17 ,18, daha az çalışma koloniizasyon için faydalı symbiontlar tarafından kullanılan moleküler mekanizmaları araştırdı21,22,23 . Transposon ekleme dizilimini kullanarak amaç, B. gladioli'nin L. villosa böceklerini kolonileştirmesini sağlayan moleküler faktörleri tanımlamaktı.
Transposon aracılı mutajensis, bir Tn5 transposon ve ters tekrar bölgeleri tarafından kuşatılmış bir kanamycin direnç kaseti taşıyan pRL27 plazmid kullanılarak gerçekleştirildi. Plazmid, plazmid donör E. coli WM3064 suşu ile konjugasyon yapılarak hedef B. gladioli Lv-StA hücrelerine sokuldu (Şekil 1'degösterildiği gibi). Konjugasyondan sonra B. gladioli alıcısı ve E. coli donör hücrelerini içeren konjugasyon karışımı kanamycin içeren seçici agar plakaları üzerine kaplandı. Plakalarda DAP bulunmaması donör E. coli hücrelerini ve başarılı B. gladioli Lv-StA transkonjugantları için seçilen kanamycin varlığını ortadan kaldırdı. 100.000 transkonjugant kolonisinin hasatından elde edilen havuzlu B. gladioli Lv-StA mutant kütüphanesi, değiştirilmiş bir DNA kütüphanesi hazırlama kiti ve özel astarlar kullanılarak sıralama için hazırlandı. Şekil 2, DNA kütüphanesi hazırlama adımlarını vurgulamaktadır. Sıralama 4 Mio eşleştirilmiş okuma sağladı; B. gladioli Lv-StA'daki 7.468 genden 3.736'sı bozuldu.
Konakta kolonizasyon kusurlu olan mutantları tanımlamak için, B. gladioli Lv-StA mutant kütüphanesi böcek yumurtalarına bulaştı ve kontrol olarak KB ortamında in vitro olarak yetiştirildi. In vivo kolonizasyon darboğaz boyutu deneyden önce hesaplandı. Bilinen sayıda B. gladioli Lv-StA hücresi böcek yumurtasına bulaştı ve taze yumurtadan çıkan ilk başlangıç larvalarındaki kolonileşen hücrelerin sayısı, her larvadan bir süspansiyonun kaplaılması ve kişi başına koloni oluşturan birimlerin sayılmasıyla elde edildi. Bu hesaplamalar, kolonileşen hücrelerin sayısının, kütüphanedeki mutantların tamamını veya yüksek bir yüzdesini konak sahibini kolonileştirme yetenekleri için değerlendirmek için yeterli olduğundan emin olmak için yapılmıştır. Ek olarak, in vitro ve in vivo koşullar arasındaki büyüme süresi, bu örnekleri karşılaştırılabilir hale getirmek için bakteri nesillerinin sayısına göre normalleştirildi.
Yumurtalar yumurtadan çıktıktan sonra 13 havuzda 1.296 larva toplandı. İlgili in vitro mutant kültürleri yetiştirildi ve gliserol stokları olarak depolandı. In vivo ve in vitro yetiştirilen mutant kütüphanelerinin DNA'sı bir ultrasonicator'da çıkarıldı ve parçalandı. Şekil 3, parçaların çoğunluğunun beklendiği gibi 100 ila 400 bp arasında yayıldığı yamyulan DNA'nın boyut dağılımını göstermektedir. Bu adımı sıralama için değiştirilmiş DNA kütüphanesi hazırlama protokolü izledi. Protokolün her adımında, adımların doğru bir şekilde gerçekleştirilmesini sağlamak ve DNA kayıplarını izlemek için kalan DNA konsantrasyonu kontrol edildi. Sıralamadan önce yapılan bir kalite kontrolü (Bkz. Malzeme Tablosu),DNA kütüphanelerinin beklenmedik şekilde büyük (>800 bp) DNA parçaları içerdiğini ve bunun in vivo kütüphanelerde daha belirgin olduğunu ortaya koydu. Sıralama şeritlerindeki parçaların kümelenmesindeki zorluk göz önüne alındığında, istenen okuma sayısına elde etmek için sıralama derinliğini in vivo kütüphanelerde 10 Mio eşleştirilmiş okumalara çıkarmak gerekiyordu. Sıralama sonuçlarının analizi, in vivo kütüphanelerde ortalama 4 Mio ve in vitro kütüphanelerinde 3.1 Mio okumasının, bu deney için tatmin edici olan 5' Read-1(Tablo 9)ucundaki Transposon kenarını içerdiğini ortaya koydu. Orijinal kütüphanedeki B. gladioli genomuna 24.224 benzersiz eklemenin dağılımı Şekil 4'te gösterilmiştir. DESeq2 kullanılarak yapılan bir analiz, 271 mutantın bolluğunun in vivo ve in vitro koşullar arasında önemli ölçüde farklı olduğunu ortaya koydu.

Şekil 3: Bir mutant ve DNA kütüphanelerinin agarose jelleri. (A) X şeridindeki bir mutantın duyulmamış DNA'sı ve ölçek için 1 kbp merdiveni olan agarose jel. (B) Yamlu DNA kütüphanesi ile jel. İlk şeritteki merdivenin bant boyutları sol tarafta belirtilir. A, b ve c'nin ilk üç şeridi in vivo kütüphanelerin yamlu DNA parçalarınıiçerir. Lanes d, e, f ve g, in vitro kütüphanelerin yamuğunda DNA parçaları içerir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 4: Burkholderia gladioli Lv-StA genomunda yer alan dört replicon genelinde orijinal kütüphanedeki benzersiz ekleme alanlarının konumu. X ekseni boyunca her çubuk bir ekleme yerinde bulunur. y ekseni boyunca bir çubuğun yüksekliği, bu siteyle ilişkili okuma sayısına karşılık gelir. İki kromozom ve iki plazmidin tam uzunlukta gösterildiğini ve bu nedenle x ekseninde farklı ölçeklere sahip olduğunu unutmayın. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
| Kralın B orta / agar | |
| Pepton (soya fasulyesi) | 20 g/M |
| K2HPO4 | 1,5 g/M |
| MgSO4.7H2O | 1,5 g/M |
| Agar | 15 g/M |
| Damıtılmış suda çözünmüş | |
| LB orta/agar | |
| Tryptone | 10 g/M |
| Maya özü | 5 g/M |
| NaCl | 10 g/M |
| Damıtılmış suda çözünmüş | |
Tablo 1: Ortam bileşenleri.
| Hayır. | Astar | Sıra | PCR tavlama sıcaklığı( °C) | |
| 1 | tpnRL17–1RC | 5'-CGTTACATCCCTGGCTTGTT-3' | 58.2 | |
| 2 | tpnRL13–2RC | 5'-TCGTGAAGAAGGTGTTGCTG-3' | ||
Tablo 2: Konjugasyonun başarısını onaylamak için primerler.
| Parça | Hacim (μL) |
| HPLC arıtılmış su | 4.92 |
| 10x Tampon S (yüksek özgüllük) | 1 |
| MgCl2 (25 mM) | 0.2 |
| dNTP'ler (2 mM) | 1.2 |
| Astar 1 (10 pmol/μL) | 0.8 |
| Astar 2 (10 pmol/μL) | 0.8 |
| Taq (5 U/μL) | 0.08 |
| Mastermix toplamı | 9 |
| Şablon | 1 |
Tablo 3: Konjugasyonun başarısını onaylamak için PCR master mix. Kısaltmalar: HPLC = yüksek performanslı sıvı kromatografisi; dNTP'ler = deoksinükleozit trifosfat.
| Adım -ları | Sıcaklık °C | Saat | Döngü |
| İlk Denatürasyon | 95 | 3 dk | 1 |
| Denatürasyon | 95 | 40 sn | |
| Tavlama | 58.2 | 40 sn | 30 ila 35 |
| Uzantı | 72 | 1-2 dk | |
| Son Uzantı | 72 | 4 dk | 1 |
| Tutmak | 4 | ∞ | |
Tablo 4: Konjugasyonun başarısını onaylamak için PCR koşulları.
| Astar | Sıra | Tm °C | Kullanmak | Kaynak | |
| Transposon spesifik biyotinilasyonlu astar | 5'-Biotin-ACAGGAACACTTAACGGCTGACATG -3' | 63.5 | 6.7.1. PCR I | Töre | |
| Değiştirilmiş Evrensel PCR astarı | 5'- AATGATACGGCGACCACCGAGATC TACACTCTTTCCCTACACGACGCTCC TTCCGATCTGAATTCATCGATGAT GGTTGAGATGTGT – 3' | 62 | 6.10.1. PCR II | Töre | |
| Dizin astarı | Üretici kılavuzuna bakın | 6.7.1. PCR I ve 6.10.1. PCR II | Illumina için NEBNext Multiplex Oligos (İndeks primer seti 1) | ||
| Bağdaştırıcı | Üretici kılavuzuna bakın | 6.5. Adaptör ligasyonu | Illumina için NEBNext Ultra II DNA kütüphanesi hazırlık kiti | ||
Tablo 5: DNA kütüphanesi hazırlığı sırasında PCR I ve II için astarlar ve adaptör.
| PCR karışımı | (μL) |
| Adaptörle liglenmiş DNA parçaları | 15 |
| NEBNext Ultra II Q5 ana karışımı | 25 |
| İndeks astarı (10 pmol/ μL) | 5 |
| Transposon spesifik biyotinillenmiş astar (10 pmol/ μL) | 5 |
| Toplam birim | 50 |
Tablo 6: DNA kütüphanesi hazırlığı-PCR I master mix.
| Adım -ları | Sıcaklık | Saat | Döngü |
| İlk Denatürasyon | 98 °C | 30 sn | 1 |
| Denatürasyon | 98 °C | 10 sn | 6'dan 12'ye |
| Tavlama | 65 °C | 30 sn | |
| Uzantı | 72 °C | 30 sn | |
| Son Uzantı | 72 °C | 2 dk | 1 |
| Tutmak | 16 °C | ∞ | |
Tablo 7: DNA kütüphanesi hazırlama-PCR I ve II koşulları.
| PCR karışımı | (μL) |
| Boncuk tarafından seçilen DNA | 15 |
| NEBNext Ultra II Q5 ana karışımı | 25 |
| Dizin astarı | 5 |
| Değiştirilmiş evrensel PCR astarı | 5 |
| Toplam birim | 50 |
Tablo 8: DNA kütüphanesi hazırlama-PCR II ana karışımı.
| Kitaplık | Invivo-1 | Invivo-2 | Invivo-3 | Invitro-1 | Invitro-2 | Invitro-3 | Özgün kitaplık | |
| Hayır. okuma sayısı (PE) | 56,57,710 | 39,19,051 | 30,65,849 | 35,73,494 | 28,83,440 | 36,61,956 | 46,09,410 | |
| Hayır. Tn içeren okumaların sayısı – 5' Read-1'in sonundaki kenar | 54,15,880 | 37,31,169 | 29,36,247 | 33,00,499 | 27,35,705 | 33,50,402 | 41,53,270 | |
| Bowtie2 genel hizalama oranı (%) (yalnızca Okuma-1) | 95.53% | 83.71% | 89.87% | 80.79% | 78.00% | 73.06% | 74.92% | |
| Benzersiz ekleme sayısı | 8,539 | 4,134 | 7,183 | 18,930 | 18,421 | 20,438 | 24,224 | |
| Vurulan gen sayısı | 1575 | 993 | 1450 | 2793 | 2597 | 3037 | 3736 | |
Tablo 9: Kitaplık başına sıralama çıktısı ve transposon ekleme sıklığının özeti. Kısaltma: PE = eşleştirilmiş uç.
Tartışmalar
L. villosa böcekleri ve B. gladioli bakterileri arasındaki simbiyotik etkileşimde önemli konak kolonizasyon faktörlerini belirlemek için bir B. gladioli transposon mutant kütüphanesi oluşturulmuştır. Protokoldeki başlıca adımlar konjugasyon, konakçı enfeksiyonu, DNA kütüphanesi hazırlama ve sıralamaydı.
Burkholderia'nın birçok türü konjugasyon24,25ile genetik modifikasyona elverişliolduğundan,transposon ve antibiyotik ekleme kasetini taşıyan plazmid, E. coli'denhedef B. gladioli Lv-StA suşuna başarıyla konjuge edildi. Elektroporasyon ile önceki dönüşüm girişimleri çok düşük bir şekilde neredeyse hiç B. gladioli transformantı vermedi. Hedef organizmanın çok sayıda dönüştürücüyu verimli bir şekilde elde etmesi için dönüşüm tekniğini optimize etmek tavsiye edilir.
B. gladioli Lv-StA'da bir tur konjugasyon ve 40 konjugasyon noktası 3.736 geni bozdu. Geçmişe bakıldığında, 7.468 genin çoğunu bozmak ve doymuş bir kütüphane elde etmek için birden fazla konjugasyon turu gerekli olacaktır. Özellikle, konjugasyon sırasında kuluçka süresinin B. gladioli'ninüstel büyüme aşamasının sonu olan 12-18 saati aşmasına izin verilmedi. Bakteri hücrelerinin üstel büyüme evresinin ötesinde konjugasyona izin vermek, transkonjugant elde etme şansını azaltır26. Bu nedenle, konjugasyon süresi bakteri türlerinin büyümesine göre ayarlanmalıdır.
Bir konakta mutant kütüphanelerinin enfeksiyonunu içeren bir deneyi başarıyla gerçekleştirmek için, kolonileşme sırasında bakteriyel popülasyon darboğaz boyutunu ve enfeksiyondan önce kütüphanedeki mutantların çeşitliliğini değerlendirmek önemlidir1,2,27. Deneye hazırlanırken, kütüphanedeki her bir mutantın örneklenme ve kolonileştirmesine izin verme şansının yüksek olması için enfekte olması gereken minimum böcek sayısını tahmin ettik. Deney süresince yaklaşık in vivo bakteriyel üretim süresi ve nesil sayısı da hesaplandı. İn vitro kültür daha sonra kuluçka süresi ayarlanarak benzer sayıda nesile kadar büyüdü. Model olmayan diğer konaklarda benzer bir enfeksiyon deneyi için, bir laboratuvar kültürünü ve konak organizmaların sabit bir kaynağını koruma yeteneği arzu edilir.
Mutant kütüphane in vivo ve in vitro ve örnek toplamanın büyümesinin ardından transposon ekleme dizilimi için değiştirilmiş bir DNA kütüphanesi hazırlama protokolü gerçekleştirildi. Protokoldeki değişiklik, özel PCR astarları tasarlamayı ve ekleme kasetini içeren DNA parçalarını seçmek için PCR adımları eklemeyi içeriyordu. Protokol özelleştirildiğinden, protokoldeki ek PCR döngüleri, aşırı örnekleme ve son kitaplıklarda karma bağdaştırıcı bağdaştırıcısı parçaları elde etme riskini artırdı. Bu nedenle, bu parçaların çıkarılmasına yardımcı olduğu için iki PCR'den sonra son bir temizleme adımı (boyut seçimi olmadan) önerilir. DNA kütüphanelerinin boyut dağılımı hala beklenenden daha genişti. Bununla birlikte, sıralama derinliğinin artırılması biyoinformatik analizi sırasında filtrelenen yeterli veriyi sağlayarak tatmin edici sonuçlar elde etti.
Transposon aracılı mutajensis tek bir deneyde binlerce rastgele ekleme ürettiğinden, bakteri üremesi için gerekli genlerin bozulduğu mutantlar hariç hepsini içeren doymuş bir mutant kütüphanesi oluşturmak mümkündür. Burkholderia sp ile ilgili diğer çalışmalarda temel genlerin tahminleri göz önüne alındığında, büyük olasılıkla doymuş bir mutant kütüphanesi ile çalışmadık. 28,29. Doymayan bir kütüphane, hedefli mutajensis kullanarak daha fazla çalışma için çeşitli aday genlerin araştırılmasına yardımcı olur. Deneylerden önce, bazı transposonların genom30'dakibelirli locilerde mutantların bolluğunu artıran belirli ekleme hedef bölgelerine sahip olduğunu hatırlamak da önemlidir. Mariner transposons ekleme için AT siteleri hedef bilinmektedir31, ve Tn5 transposons bir GC önyargı vardır32,33. Transposon eklemeleri için sıcak noktaları tanımak için biyoinformatik analizi sırasındaki adımların dahil etmesi, herhangi bir dağıtım önyargısını değerlendirmeye yardımcı olacaktır.
Aksiliklere eğilimli olsa da, iyi tasarlanmış bir transposon ekleme dizileme deneyi, bakterilerdeki birçok koşullu önemli geni tek bir deneyde tanımlamak için güçlü bir araç olabilir. Örneğin, Burkholderia seminalis'te orkide yaprağı nekrozunun bastırılması için önemli olan bir düzine gen transposon mutajensis ve genomik34birleştirilerek tanımlanmıştır. Burkholderia'nınötesinde, çeşitli yapışma ve hareketlilik genleri ve taşıyıcıları, Apis mellifera (Honeybee)22'nin Snodgrassella alvi symbionts'unda ve Euprymna scolopes'un Vibrio fischerii payandalarında (Hawaii bobtail kalamar)23 transposon eklemeli mutagenesis yaklaşımını kullanarak önemli kolonileşme faktörleri olarak tanımlanmıştır.
Alternatif bir yaklaşım olarak transposon mutajensis, sıralama yerine seçici ortam kullanılarak bireysel mutantların taranarak takip edilebilir. Hareketlilik, biyoaktif ikincil metabolitlerin üretimi veya spesifik auxotrophies gibi eksiklikleri tanımlamak için fenotipik tarama veya biyoassaylar mümkündür. Örneğin, burkholderia insecticola (Caballeronia35cinsine yeniden atanmıştır) transposon mutant kütüphanesinin taranmasının, symbiontların Riptortus pedestris'ikolonileştirmek için hareketlilik genleri kullandığı, böcek konakçılarının36. Ayrıca, transposon mutajensis ve fenotipik tarama kullanılarak, burkholderia caryophylli37'debiyoaktif ikincil metabolit karyosenentin için biyosentetik gen kümesi tanımlanmıştır. Transposon mutajensis ve taramadan sonra Burkholderia psödomallei'nin bir auxotrophic mutant'ı tanımlanmıştır ve insanlarda ve hayvanlarda tehlikeli bir hastalık olan melioidoza karşı olası bir zayıflatılmış aşı adayıdır38. Bu nedenle, transposon mutajensis ve dizileme, patojenik veya karşılıklı ilişkilerde ilgili konakçılarla etkileşimleri için önemli olan bakterilerin moleküler özelliklerinin incelenmesinde değerli bir yaklaşımdır.
Açıklamalar
Yazarlar, çalışmayla ilgili herhangi bir çıkar çatışması olmadığını beyan ederler.
Teşekkürler
Prosedürde konjugasyon ve rehberlik için E. coli WM3064 +pRL27 suşu sağladığı için Junbeom Lee'ye, mutant kütüphane üretimi sırasında sorun gidermeye yardımcı olduğu için Kathrin Hüffmeier'e ve böcek toplama ve izin alımını desteklediği için Prof. André Rodrigues'e minnettarız. Ayrıca Rebekka Janke ve Dagmar Klebsch'e böceklerin toplanması ve yetiştirisinde destekleri için teşekkür ediyoruz. Brezilyalı yetkilileri böcek örneklerine erişim, toplama ve ihracat için aşağıdaki izinleri verdiği için kabul ediyoruz: SISBIO yetkilendirme Nr. 45742-1, 45742-7 ve 45742-10, CNPq süreci nº 01300.004320/2014-21 ve 01300.0013848/2017-33, IBAMA Nr. 14BR016151DF ve 20BR035212/DF). Bu araştırma, Alman Bilim Vakfı (DFG) Araştırma Hibeleri FL1051/1-1 ve KA2846/6-1 tarafından finanse edildi.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 2,6- Diaminopimelic Acid | Alfa Aesar | B22391 | For E.coli WM3064+ pRL27 |
| Agar - Agar | Roth | 5210 | |
| Agarose | Biozym | 840004 | |
| AMPure beads XP (magentic beads + polyethylene glycol + salts) | Beckman Coulter | A63880 | Size selection in step 6.6 |
| Bleach (NaOCl) 12% | Roth | 9062 | |
| Bowtie2 v.2.4.2 | Bioinfromatic tool for read mapping. Reference 10 in main manuscript. | ||
| Buffer-S | Peqlab | PEQL01-1020 | For PCRs |
| Cell scraper | Sarstedt | 83.1830 | |
| Cutadapt v.2.10 | Bioinformatic tool for removing specific adapter sequences from the reads. Reference 8 in main manuscript. | ||
| DESeq2 | RStudio package for assessing differential mutant abundance. Usually used for RNAseq analysis. Reference 12 in main manuscript. | ||
| DNA ladder 100 bp | Roth | T834.1 | |
| dNTPs | Life Technology | R0182 | PCR for confirming success of conjugation |
| EDTA, Di-Sodium salt | Roth | 8043 | |
| Epicentre MasterPure Complete DNA and RNA Purification Kit | Lucigen | MC85200 | |
| Ethidium bromide | Roth | 2218.1 | |
| FastQC v.0.11.8 | Bioinformatic tool for assessing the quality of sequencing data. Reference 7 in main manuscript. | ||
| FeatureCounts v.2.0.1 | Bioinformatic tool to obtain read counts per genomic feature. Reference 11 in main manuscript. | ||
| Glycerol | Roth | 7530 | |
| K2HPO4 | Roth | P749 | |
| Kanamycin sulfate | Serva | 26899 | |
| KCl | Merck | 4936 | |
| KH2PO4 | Roth | 3904 | |
| MgSO4.7H2O | Roth | PO27 | |
| Na2HPO4 | Roth | P030 | |
| NaCl | Merck | 6404 | |
| NEBNext Multiplex Oligos for Illumina (Index primers set 1) | New England Biolabs | E7335S | |
| NEBNext Ultra II DNA library prep kit for Illumina | New England Biolabs | E7645S | |
| Peptone (soybean) | Roth | 2365 | For Burkholderia gladioli Lv-StA KB-medium |
| peqGOLD 'Hot' Taq- DNA Polymerase | VWR | PEQL01-1020 | PCR for confirming success of conjugation |
| Petri plates - 145 x 20 mm | Roth | XH90.1 | For selecting transconjugants |
| Petri plates - 90 x 16 mm | Roth | N221.2 | |
| Qiaxcel (StarSEQ GmbH, Germany) | Quality check after DNA library preparation | ||
| Streptavidin beads | Roth | HP57.1 | |
| Taq DNA polymerase | VWR | 01-1020 | |
| Trimmomatic v.0.36 | Bioinformatic tool for trimming low quality reads and also adapter sequences. Reference 9 in main manuscript. | ||
| Tris -HCl | Roth | 9090.1 | |
| Tryptone | Roth | 2366 | For Escherichia coli WM3064+pRL27 LB medium |
| Ultrasonicator | Bandelin | GM 70 HD | For shearing |
| USER enzyme (uracil DNA glycosylase + DNA glycosylase- lyase Endonuclease VIII) | New England Biolabs | E7645S | Ligation step 6.5.2 |
| Yeast extract | Roth | 2363 |
Referanslar
- Cain, A. K., et al. A decade of advances in transposon-insertion sequencing. Nature Reviews Genetics. 21 (9), 526-540 (2020).
- Chao, M. C., Abel, S., Davis, B. M., Waldor, M. K. The design and analysis of transposon insertion sequencing experiments. Nature Reviews Microbiology. 14 (2), 119-128 (2016).
- Barquist, L., Boinett, C. J., Cain, A. K. Approaches to querying bacterial genomes with transposon-insertion sequencing. RNA Biology. 10 (7), 1161-1169 (2013).
- Flórez, L. V., et al. Antibiotic-producing symbionts dynamically transition between plant pathogenicity and insect-defensive mutualism. Nature Communications. 8 (1), 15172 (2017).
- Flórez, L. V., Kaltenpoth, M. Symbiont dynamics and strain diversity in the defensive mutualism between Lagria beetles and Burkholderia. Environmental Microbiology. 19 (9), 3674-3688 (2017).
- Flórez, L. V., et al. An antifungal polyketide associated with horizontally acquired genes supports symbiont-mediated defense in Lagria villosa beetles. Nature Communications. 9 (1), 2478 (2018).
- . FastQC A quality control tool for high throughput sequence data Available from: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (2012)
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal. 17 (1), 10-12 (2011).
- Bolger, A. M., Lohse, M., Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics. 30 (15), 2114-2120 (2014).
- Langmead, B., Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nature Methods. 9 (4), 357-359 (2012).
- Liao, Y., Smyth, G. K., Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics. 30 (7), 923-930 (2014).
- Love, M. I., Huber, W., Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biology. 15 (12), 550 (2014).
- Gaytán, M. O., Martínez-Santos, V. I., Soto, E., González-Pedrajo, B. Type three secretion system in attaching and effacing pathogens. Frontiers in Cellular and Infection Microbiology. 6, 129 (2016).
- Hachani, A., Wood, T. E., Filloux, A. Type VI secretion and anti-host effectors. Current Opinion in Microbiology. 29, 81-93 (2016).
- Deep, A., Chaudhary, U., Gupta, V. Quorum sensing and bacterial pathogenicity: From molecules to disease. Journal of Laboratory Physicians. 3 (1), 4-11 (2011).
- Silva, A. J., Benitez, J. A. Vibrio cholerae biofilms and cholera pathogenesis. PLOS Neglected Tropical Diseases. 10 (2), 0004330 (2016).
- Navarro-Garcia, F., Ruiz-Perez, F., Cataldi, &. #. 1. 9. 3. ;., Larzábal, M. Type VI secretion system in pathogenic Escherichia coli: structure, role in virulence, and acquisition. Frontiers in Microbiology. 10, 1965 (2019).
- Ribet, D., Cossart, P. How bacterial pathogens colonize their hosts and invade deeper tissues. Microbes and Infection. 17 (3), 173-183 (2015).
- Schwarz, S., et al. Burkholderia Type VI secretion systems have distinct roles in eukaryotic and bacterial cell interactions. PLoS Pathogens. 6 (8), 1001068 (2010).
- Jones, C., et al. Kill and cure: genomic phylogeny and bioactivity of Burkholderia gladioli bacteria capable of pathogenic and beneficial lifestyles. Microbial Genomics. 7 (1), 000515 (2021).
- Takeshita, K., Kikuchi, Y. Riptortuspedestris and Burkholderia symbiont: an ideal model system for insect-microbe symbiotic associations. Research in Microbiology. 168 (3), 175-187 (2017).
- Powell, J. E., et al. Genome-wide screen identifies host colonization determinants in a bacterial gut symbiont. Proceedings of the National Academy of Sciences of the United States of America. 113 (48), 13887-13892 (2016).
- Brooks, J. F., et al. Global discovery of colonization determinants in the squid symbiont Vibrio fischeri. Proceedings of the National Academy of Sciences of the United States of America. 111 (48), 17284-17289 (2014).
- Somprasong, N., McMillan, I., Karkhoff-Schweizer, R. R., Mongkolsuk, S., Schweizer, H. P. Methods for genetic manipulation of Burkholderia gladioli pathovar cocovenenans. BMC Research Notes. 3 (308), (2010).
- Garcia, E. C. Burkholderia thailandensis: Genetic manipulation. Current Protocols in Microbiology. 45, 1-15 (2017).
- Headd, B., Bradford, S. A. The conjugation window in an Escherichia coli K-12 strain with an IncFII plasmid. Applied and Environmental Microbiology. 86 (17), 00948 (2020).
- Van Opijnen, T., Camilli, A. Transposon insertion sequencing: A new tool for systems-level analysis of microorganisms. Nature Reviews Microbiology. 11 (7), 435-442 (2013).
- Gallagher, L. A., Ramage, E., Patrapuvich, R., Weiss, E., Brittnacher, M., Manoil, C. Sequence-defined transposon mutant library of Burkholderia thailandensis. mBio. 4 (6), 00604-00613 (2013).
- Wong, Y. -. C., et al. Candidate essential genes in Burkholderia cenocepacia J2315 identified by genome-wide TraDIS. Frontiers in Microbiology. 7, 1288 (2016).
- Moule, M. G., et al. Genome-wide saturation mutagenesis of Burkholderia pseudomallei K96243 predicts essential genes and novel targets for antimicrobial development. mBio. 5 (1), 00926 (2014).
- Ding, Q., Tan, K. S. Himar1 transposon for efficient random mutagenesis in Aggregatibacter actinomycetemcomitans. Frontiers in Microbiology. 8, 1842 (2017).
- Green, B., Bouchier, C., Fairhead, C., Craig, N. L., Cormack, B. P. Insertion site preference of Mu, Tn5, and Tn7 transposons. Mobile DNA. 3, 3 (2012).
- Lodge, J. K., Weston-Hafer, K., Berg, D. E. Transposon Tn5 target specificity: Preference for insertion at G/C pairs. Genetics. 120 (3), 645-650 (1988).
- Ará Ujo, W. L., et al. Genome sequencing and transposon mutagenesis of Burkholderia seminalis TC3.4.2R3 identify genes contributing to suppression of orchid necrosis caused by B. gladioli. Molecular Plant-microbe Interactions: MPMI. 29 (6), 435-446 (2016).
- Dobritsa, A. P., Samadpour, M. Reclassification of Burkholderiainsecticola as Caballeroniainsecticola comb. nov. and reliability of conserved signature indels as molecular synapomorphies. International Journal of Systematic and Evolutionary Microbiology. 69 (7), 2057-2063 (2019).
- Ohbayashi, T., et al. Insect's intestinal organ for symbiont sorting. Proceedings of the National Academy of Sciences of the United States of America. 112 (37), 5179-5188 (2015).
- Ross, C., Scherlach, K., Kloss, F., Hertweck, C. The molecular basis of conjugated polyyne biosynthesis in phytopathogenic bacteria. Angewandte Chemie International Edition. 53 (30), 7794-7798 (2014).
- Atkins, T., et al. A mutant of Burkholderia pseudomallei, auxotrophic in the branched chain amino acid biosynthetic pathway, is attenuated and protective in a murine model of melioidosis. Infection and Immunity. 70 (9), 5290-5294 (2002).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır