
Method Article
الفرز السلبي للنوى المنشطة بالفلورة للخلايا العصبية جنبا إلى جنب مع تسلسل الحمض النووي الريبي أحادي النوى لدراسة مكانة الحصين العصبية
In This Article
Summary
تظهر هنا طريقة لتسلسل النوى المفردة المعزولة من التلفيف المسنن للفأر الذي يستبعد معظم الخلايا العصبية من خلال فرز النوى المنشطة بالتألق (FAN). يولد هذا النهج ملفات تعريف تعبير عالية الجودة ويسهل دراسة معظم أنواع الخلايا الأخرى الممثلة في مكانه ، بما في ذلك المجموعات السكانية النادرة مثل الخلايا الجذعية العصبية.
Abstract
تكوين الخلايا العصبية الحصينية للبالغين (AHN) ، والذي يتكون من صيانة مدى الحياة للخلايا الجذعية العصبية التكاثرية والهادئة (NSCs) داخل المنطقة شبه الحبيبية (SGZ) من التلفيف المسنن (DG) وتمايزها عن الخلايا العصبية المولودة حديثا إلى الخلايا الحبيبية في طبقة الخلايا الحبيبية ، تم التحقق من صحتها جيدا عبر العديد من الدراسات. يعد استخدام الحيوانات المعدلة وراثيا ، وخاصة القوارض ، أداة قيمة للتحقيق في مسارات الإشارات التي تنظم AHN ودراسة دور كل نوع من الخلايا التي تشكل مكانة الحصين العصبية. لمعالجة هذا الأخير ، كان للطرق التي تجمع بين عزل النواة المفردة وتسلسل الجيل التالي تأثير كبير في مجال AHN لتحديد التوقيعات الجينية لكل مجموعة خلية. ومع ذلك ، هناك حاجة إلى مزيد من التحسين لهذه التقنيات لتحديد ملامح مجموعات الخلايا النادرة ظاهريا داخل DG. هنا ، نقدم طريقة تستخدم فرز النوى المنشطة الفلورية (FANS) لاستبعاد معظم المجموعات العصبية من معلق نواة واحد معزول من DG الذي تم تشريحه حديثا ، عن طريق اختيار نوى غير ملوثة لمستضد NeuN ، من أجل إجراء تسلسل الحمض النووي الريبي أحادي النوى (snRNA-seq). هذه الطريقة هي نقطة انطلاق محتملة لمزيد من التحقيق في التنظيم بين الخلايا ل AHN والكشف عن علامات وآليات خلوية جديدة عبر الأنواع.
Introduction
يرتبط التوليد المستمر للخلايا العصبية الحصينية في مرحلة البلوغ ، والمعروف أيضا باسم تكوين الخلايا العصبية الحصينية للبالغين (AHN) ، بالوظائف المعرفية مثل التعلم واكتساب / إزالة الذاكرة وفصل الأنماط ويبدو أنه آلية مهمة للمرونة في الشيخوخة والأمراض التنكسية العصبية لمنع العجز المعرفي1،2،3 . كانت القوارض هي النموذج المفضل لدراسة AHN باستخدام عدة طرق ، بما في ذلك الكيمياء المناعية وطرق تسلسل الجيل التالي (NGS). لا تزال ترجمة هذه النتائج إلى الأنواع الأخرى مثيرة للجدل. في الواقع ، لوحظ AHN في معظم الأنواع ولكن مدى استمراره طوال الحياة ، وخاصة في البشر4،5،6،7،8 ، تتم مناقشته بانتظام.
حتى الآن ، تم تأكيد العديد من مسارات الإشارات الداخلية والخارجية لتعديل AHN1. ومع ذلك ، فإن تأثير الاتصال بين الخلايا على AHN هو فقطالناشئة 9. يمكن أن يعزى هذا أولا إلى عدم كفاية خصوصية علامات الخلايا المعروفة حاليا لإجراء تحليل في الجسم الحي مع الحيوانات المعدلة وراثيا. في الواقع ، اعتمدت العديد من الدراسات على علامات مثل الكورتين المزدوج أو البروتين الحمضي الليفي الدبقي (GFAP) التي يتم التعبير عنها في أنواع خلايا متعددة1. ثانيا ، يجلب التعقيد والدرجة العالية من تنوع الخلايا في مكانة الحصينالبالغة 10 تحديات تقنية لتعريف كل نوع من الخلايا. هذا هو الحال بشكل خاص بالنسبة للتحليل المعلوماتي الحيوي مع العلامات الخلوية المتداخلة المستخدمة في خطوط الأنابيب التحليلية لمختلف السكان ، مثل NSCs أو الخلايا الدبقية ، مما يؤدي إلى استنتاجات مثيرة للجدل عند تقييم AHN 7,11. ثالثا ، يقوض العدد الهائل من الخلايا العصبية التحقيق في مجموعات الخلايا الأقل وفرة ، مثل الخلايا النجمية أو الخلايا قليلة التغصن أو الخلايا العصبية ، على الرغم من أن دورها في تنظيم الضبط الدقيق ل AHN أصبح بارزا9. تؤثر هذه القيود معا على القدرة على ترجمة النتائج من القوارض إلى الأنواع الأخرى. يتضخم هذا بشكل خاص بسبب صعوبة تلخيص الأنسجة المعقدة في المختبر ، مثل مكانة الحصين العصبية ، والعقبات العديدة للوصول إلى الأنسجة عالية الجودة جنبا إلى جنب مع عدم وجود بروتوكولات موحدة لمعالجة الأنسجة في الدراسات التي تنطوي على الأنسجة البشرية12,13. لذلك من الأهمية بمكان تطوير مناهج جديدة لمجموعات الخلايا الجانبية وتحديد علامات خلوية جديدة داخل التلفيف المسنن (DG) والتي ستؤدي في النهاية إلى فهم أفضل للمساهمات المختلفة لكل نوع من أنواع الخلايا في تنظيم AHN.
لتحقيق ذلك ، أصبح عزل الخلية المفردة (sc) والنوى المفردة (sn) جنبا إلى جنب مع تسلسل الحمض النووي الريبي مفيدا للتحقيق في الأنسجة المعقدة مثل DG14. على هذا النحو ، تم تنفيذ استراتيجيات التخصيب الخلوي لعزل الخلايا المفردة من مكانة الحصين البالغة للفأر في الغالب لفحص NSCs15,16. تم تطبيق استراتيجية مثيرة للاهتمام لإثراء الخلايا غير العصبية من DG عن طريق تسلسل الخلايا المفردة المزدوجة السلبية GluR1 / Cd24 والتي أدت إلى تسلسل 1,408 خلية بدون مجموعات مميزة بين الخلايا النجمية و NSCs بعد التحليل المعلوماتي الحيوي17. قد يكون هذا بسبب الهضم الأنزيمي القاسي المطلوب لإعداد خلية واحدة يضر بسلامة الخلية والحمض النووي الريبي. لتجاوز هذه المشكلة الفنية ، تم تطوير عدة طرق تستخدم عزل النواة المفردة بدلا من ذلك وهي مناسبة بشكل خاص للأنسجة المعقدة11,18. ومع ذلك ، فإن غلبة الخلايا العصبية داخل DG أو على نطاق أوسع داخل نظام الحصين والأنف والأذن والحنجرة يولد تحيزا في أخذ العينات لدراسة كامل مجموعات الخلايا الموجودة داخل مناطق الدماغ هذه. بالإضافة إلى ذلك ، فإن العدد المحدود من الخلايا التي سيتم تحميلها لإعداد مكتبات الخلية المفردة يبرز وجود مجموعة الخلايا الرئيسية في خطوط الأنابيب التحليلية للنوى المفردة المتتابعة. في الواقع ، غالبا ما يتم شرح مجموعات الخلايا العصبية الكبيرة وتحليلها بينما يتم تمثيل مجموعات الخلايا الأخرى تمثيلا ناقصا أو تفويتها 5,11.
في محاولة للتغلب على هذه التحيزات والقدرة على تحديد أنواع الخلايا بخلاف الخلايا العصبية الموجودة في الفأر DG ، تم ابتكار طريقة في هذه الدراسة باستخدام مبدأ فرز النوى المنشطة الفلورية (FANS)18 الذي يستبعد معظم مجموعات الخلايا العصبية عن طريق الاختيار السلبي للنوى المفردة الملطخة مع المستضد النووي العصبي (NeuN, المعروف أيضا باسم Rbfox3). تم توجيه هذا الاختيار للمستضد من خلال الأدبيات التي تصف NeuN كعلامة عصبية موثوقة19 وضرورة استخدام بروتين نووي لهذا النهج. ثم تم تحضير الخلايا المصنفة من قبل FACS سلبية NeuN لتسلسل الحمض النووي الريبي على منصة الجينوم 10x. تظهر النتائج أن استبعاد الخلايا المعبرة عن NeuN يسمح بتنميط نسخي عالي الجودة من نوع الخلية لمجموعات الخلايا الدبقية والنادرة.
Protocol
تم تنفيذ رعاية الحيوان والإجراءات التجريبية وفقا للمبادئ التوجيهية لمعهد فرانسيس كريك ، وكذلك إرشادات وقوانين وزارة الداخلية في المملكة المتحدة.

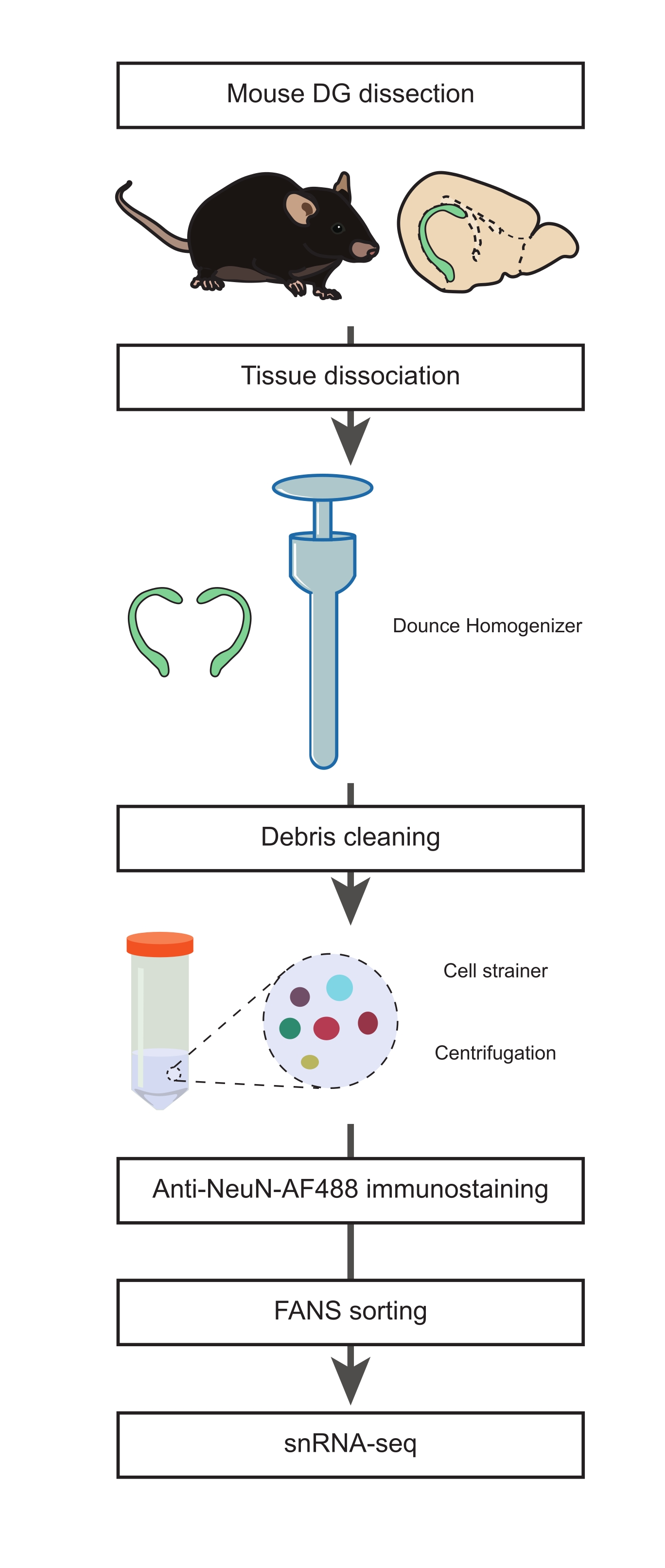
الشكل 1: تحضير معلق نواة واحدة من DG المقطع للفئران البالغة لتسلسل snRNA-seq للمجموعات غير العصبية. مخطط التدفق الذي يصف الخطوات الرئيسية للبروتوكول التي تشمل تشريح الماوس DG ، وإعداد تعليق النواة المفردة ، و NeuN المناعي ، وفرز NeuN-FANS السلبي قبل الشروع في snRNA-seq. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}
1. تشريح المدير العام (التوقيت: 15 دقيقة)
- تحضير وسائط عزل النوى 1 و 2 (NIM1 و NIM2) ، ومخزن التجانس (HB) ، ووسائط الغسيل (WM) (الجدول التكميلي 1). ضع جميع المخازن المؤقتة والوسائط والكواشف والأدوات على الثلج لحين الحاجة. ضع الخالط Dounce (انظر جدول المواد) على الثلج أثناء التحضير (1 ساعة على الأقل قبل خطوة التجانس).
ملاحظة: يمكن تحضير NIM1 وتخزينه في درجة حرارة 4 درجات مئوية لمدة تصل إلى 6 أشهر. يجب تحضير NIM2 و HB و WM حديثا.
تنبيه: تعامل مع DTT ومثبط الأنزيم البروتيني و Triton X-100 بعناية. هذه المركبات مزعجة للجلد والعين ، وسامة بشكل حاد ، وخطرة على البيئة المائية. أثناء استخدام هذه المواد الكيميائية ، ارتد قفازات واقية وملابس وحماية للعين والوجه ، واغسل يديك جيدا بعد المناولة ، وتجنب إطلاقها في البيئة. - القتل الرحيم لفأر C57Bl / 6J يبلغ من العمر 22 شهرا عن طريق خلع عنق الرحم باتباع إجراء الجدول 1 لوزارة الداخلية20.
ملاحظة: انظر المناقشة لمعرفة الأساس المنطقي فيما يتعلق باستخدام فأر يبلغ من العمر 22 شهرا في هذه الدراسة. ومع ذلك ، يمكن إجراء هذا البروتوكول في أي عمر عبر العمر. - قم بتشريح الدماغ من فأر القتل الرحيم ونقله إلى طبق بتري 10 سم مملوء بالثلج البارد 1x PBS (الشكل 1). ضع طبق بتري على الثلج. إزالة المخيخ باستخدام مشرط وقطع الدماغ في النصف بين نصفي الكرة الأرضية (على طول المحور السهمي).
- املأ طبق بتري جديد بطول 10 سم مع PBS المثلج وضعه على الثلج. انقل نصف الدماغ إلى طبق بتري الجديد. باستخدام مناظير ، قم بتشريح DG وكرر هذه الخطوة للحصول على DG الثاني من النصف الثاني من الدماغ.
ملاحظة: تم تكييف هذه الخطوات (الخطوات 1.2-1.4) من الإجراء21 الموصوف مسبقا. من المهم المضي قدما في أسرع وقت ممكن في هذه المرحلة للحفاظ على سلامة الخلايا. - انقل DGs إلى خالط Dounce المبرد مسبقا وأضف 1 مل من HB البارد.
2. تفكك الأنسجة ، وعزل النواة المفردة ، والتلطيخ المناعي المضاد ل NeuN (التوقيت: 2 ساعة)
- تجانس الأنسجة مع 10 ضربات من المدقة "A" فضفاضة ، تليها 15 السكتات الدماغية من المدقة "B" ضيقة.
ملاحظة: يجب إجراء التجانس باستخدام الهاون على الثلج بضربات لطيفة لتقليل الحرارة الناتجة عن الاحتكاك والرغوة. يجب تبريد جميع المخازن المؤقتة والمعدات مسبقا والاحتفاظ بها على الجليد أثناء العملية. - نقل التجانس إلى أنبوب 15 مل مبرد مسبقا ؛ شطف الخالط Dounce مع 1 مل من HB الباردة والجمع مع نفس الأنبوب. أضف 3 مل من HB إلى أنبوب 15 مل واحتضان 5 دقائق على الثلج. امزج 2x عن طريق قلب الأنبوب برفق.
- قم بترطيب غطاء مصفاة 70 ميكرومتر مسبقا مع 0.5 مل من HB على أنبوب اختبار سعة 50 مل. صفي معلق النوى من الخطوة 2.2 عن طريق قلب الأنبوب سعة 15 مل برفق في مصفاة الخلية. اغسل مصفاة الخلية ب 0.5 مل من HB.
- قم بإزالة مصفاة الخلية وأجهزة الطرد المركزي لأنبوب الاختبار عند 500 × جم لمدة 5 دقائق عند 4 درجات مئوية ، باستخدام جهاز طرد مركزي دلو متأرجح. تخلص من المادة الطافية.
ملاحظة: سيساعد إجهاد التجانس في تقليل الحطام ، وهو أمر بالغ الأهمية لقياس التدفق الخلوي وخطوات snRNA-seq في اتجاه مجرى النهر. - أعد تعليق الحبيبات برفق في 4 مل من HB باستخدام ماصة P1000. احتضان على الجليد لمدة 5 دقائق. تدور في 500 × غرام لمدة 10 دقائق عند 4 درجات مئوية. تخلص من المادة الطافية وأعد تعليق الحبيبات في 3 مل من WM.
- قم بترطيب غطاء مصفاة 35 ميكرومتر مسبقا فوق أنبوب اختبار سعة 15 مل مع 0.5 مل من WM. صفي معلق النوى من الخطوة 2.5 عبر مصفاة الخلية ، مع سحب 0.5 مل برفق في المرة الواحدة باستخدام ماصة P1000.
- اغسل غطاء المصفاة ب 0.5 مل من WM وضع الأنبوب على الثلج. انقل الراشح إلى أنبوب جديد سعة 15 مل وأجهزة طرد مركزي لمدة 5 دقائق و 4 درجات مئوية عند 500 × جم. تخلص من المادة الطافية وأعد تعليق الحبيبات في 3 مل من WM.
- تدور عند 500 × جم لمدة 5 دقائق عند 4 درجات مئوية. تخلص من المادة الطافية وأعد تعليق الحبيبات في 1 مل من WM باستخدام الجسم المضاد للفأر المضاد ل NeuN و Alexa Fluor 488 (anti-NeuN-AF488 ، 1: 32,000) و 1 ميكروغرام / مل DAPI. احتضان لمدة 45 دقيقة على الجليد في الظلام.
ملاحظة: لتحسين التلوين المناعي للنوى المعزولة ، يوصى بمعايرة الجسم المضاد لتحديد التخفيف الأمثل لتحليل قياس التدفق الخلوي وفرزه. بعد ذلك ، قم بتشغيل عناصر تحكم كافية للتأكد من أن ظروف التلوين هي الأمثل. على سبيل المثال ، مع الجسم المضاد المضاد ل NeuN-AF488 المترافق ، تم تشغيل عنصر تحكم سلبي (أي عدم إضافة الجسم المضاد ، الشكل التكميلي 1A) والتحكم الإيجابي (أي تلطيخ الجسم المضاد ، الشكل التكميلي 1B) لتقييم فصل السكان غير الملطخين والملطخين. عند البدء في العمل مع جسم مضاد مقترن AF488 ، يوصى بتشغيل عنصر تحكم في النمط المتماثل المقترن AF488 لتقييم النوعية. إذا تم استخدام جسم مضاد غير مقترن ، فقد يكون من الضروري التحكم الإضافي مثل إضافة جسم مضاد ثانوي فقط إلى مستحضر النوى لتقييم الارتباط غير المحدد للجسم المضاد الثانوي.
3. فرز النوى المنشط بالتألق (FANS) لاستبعاد مجموعات الخلايا العصبية (التوقيت: 45 دقيقة)
- انقل معلق النوى المصبوغ بالمناعة إلى أنبوب اختبار سعة 5 مل واحتفظ به على الجليد حتى بدء إجراء قياس التدفق الخلوي.
ملاحظة: إذا كنت تعمل مع قطع أكبر من الأنسجة من اثنين من الماوس DG ، فقد تكون هناك حاجة إلى مزيد من التخفيف باستخدام المخزن المؤقت WM لتجنب انسداد FACS إذا أصبحت كثافة النوى في المحلول عالية. - دوامة العينات لمدة 3 ثوان بسرعة منخفضة قبل وضع الأنابيب في أداة FACS (انظر جدول المواد).
ملاحظة: (إعداد FACS) يجب محاذاة آلات الفرز في بداية الإجراء مع جزيئات المعايرة وفقا لتوصيات الشركة المصنعة. تمت معايرة تأخير السقوط بالخرز أو الكريات المجهرية (انظر جدول المواد) وفقا لنموذج FACS. تم فرز العينات عند 4 درجات مئوية في وضع النقاء. لتقليل حجم التجميع ، تم فرز النوى من خلال فوهة 70 ميكرومتر عند الضغط الموصى به لمقياس التدفق الخلوي. تم فرز النوى إلى أنابيب ربط منخفضة سعة 1.5 مل (انظر جدول المواد) تحتوي على 50 ميكرولتر من WM. تم طلاء جميع أنابيب التجميع في PBS + 5٪ BSA عند 4 درجات مئوية طوال الليل لتقليل خطر التصاق النوى بجدران الأنبوب. - للحصول على البيانات من عينة من معلق النوى الملطخة ، اضبط البوابات في ارتفاع DAPI ومنطقة DAPI من أجل استبعاد حطام الخلية والنوى المجمعة (الشكل 2 أ). علاوة على ذلك ، افصل النوى المفردة عن أي مجاميع أو حطام خلوي ملطخ ب DAPI عن طريق وضع البوابات في منطقة التشتت الجانبي للسجل (SSC) ومنطقة التشتت الأمامي (FCS) (الشكل 2 ب).
- اضبط بوابات منطقة NeuN-AF488 المضادة ومنطقة FSC ، لعزل السكان السالبين NeuN-AF488 ، كما هو موضح في الشكل 2C.
- بعد التحليل ، باستخدام استراتيجية البوابة الموضحة أعلاه ، قم بفرز السكان سلبي NeuN-AF488 في أنبوب تجميع 1.5 مل مملوء ب 50 ميكرولتر من WM.
ملاحظة: باتباع استراتيجية البوابات الموضحة أعلاه وإجراء التشريح لعزل DG عن دماغ الفأر البالغ ، من المتوقع أن يمثل السكان سلبي NeuN-AF488 ~ 14٪ من النوى المفردة.

الشكل 2: العزل والتنميط النسخي لمجموعات الخلايا غير العصبية من استراتيجية بوابات DG. (A-C) لعزل نوى NeuN-AF488 المفردة السالبة واستبعاد حطام الخلية. (أ) مخطط نقطي لعينة تمثيلية من النوى المعزولة ، يصور إعداد البوابة لاختيار نوى DAPI + واستبعاد حطام الخلية والمجاميع. (ب) مزيد من اختيار النوى المفردة ذات الصلة باستخدام منطقة FSC ومنطقة SSC. ج: بوابات NeuN-AF488 لاستبعاد المجتمع الإحصائي الموجب وفرز النوى المفردة السالبة. (د) صورة مجهرية لمعلق أحادي النواة جيد مع الحد الأدنى من الحطام ونسبة أعلى من النوى ذات النوعية الجيدة (الشكل الدائري ، السهم الأسود) مقارنة بالنوى ذات النوعية الرديئة (السهم الأبيض). قضبان المقياس = 50 ميكرومتر ، 10 ميكرومتر (أقحم). (ه، واو) تحليل بيانات snRNA-seq وتوصيف مجموعات الخلايا المتميزة المعزولة من DG لذكور الفئران C57BL / 6J البالغة من العمر 22 شهرا. تقريب وإسقاط مشعب موحد لمخططات تقليل الأبعاد (UMAP) لملفات تعريف النواة المفردة من (E) الخلايا غير المصنفة FACS و (F) الخلايا المصنفة من FACS سلبية NeuN ، ملونة حسب نوع الخلية. (ز) مخططات دائرية تقارن ترددات أنواع الخلايا المحددة في كلتا العينتين. (ح) المقاييس الخاصة بالعينات المتتابعة: عدد النوى، ومتوسط عدد الجينات، والنسخ لكل نواة. (ط) مخططات الكمان التي توضح توزيع عدد الجينات والنصوص المكتشفة لكل نوع من أنواع الخلايا في كلتا العينتين. أستر. = الخلايا النجمية ، أوليج. = oligodendrocytes ، Vasc. = خلايا الأوعية الدموية ، CRCs = خلايا Cajal-Retzius ، Neur. = الخلايا العصبية ، Imm. = الخلايا المناعية ، OPCs = خلايا السلائف oligodendrocytes. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}
4. تحضير معلق النواة المفردة لإجراء تسلسل الحمض النووي الريبي أحادي النوى (التوقيت: 30 دقيقة)
- بعد الفرز ، أضف 1 مل من PBS يحتوي على 1٪ BSA إلى أنبوب التجميع من أجل جمع القطرات على جدار الأنبوب وتدور بسرعة 500 × جم لمدة 5 دقائق عند 4 درجات مئوية. تجاهل الطافي ، وترك 50 ميكرولتر.
ملاحظة: تعامل مع نوى الطرد المركزي بعناية حيث قد يكون من الصعب ملاحظة أي حبيبات في قاع الأنبوب. سيساعد استخدام جهاز طرد مركزي دلو متأرجح على التخلص من المادة الطافية دون تعطيل الحبيبات. - ماصة بلطف لإعادة تعليق نوى الطرد المركزي. أضف 5 ميكرولتر من معلق النوى إلى 5 ميكرولتر من تريبان الأزرق في أنبوب دقيق سعة 0.5 مل.
تنبيه: تعامل مع تريبان الأزرق بعناية لأنه يشكل خطرا على الصحة ، ويمكن أن يسبب السرطان ، ويشتبه في أنه يضر بالخصوبة أو الطفل الذي لم يولد بعد. ارتد قفازات واقية وملابس وحماية للعين والوجه. لا تتعامل حتى تتم قراءة جميع احتياطات السلامة وفهمها. - قياس التركيز وتقييم صلاحية تعليق الخلية الواحدة باستخدام مقياس الدم أو عداد الخلايا الآلي (انظر جدول المواد). قم بإعداد المكتبة وتسلسل النوى كما هو مفصل في الخطوة 5.
ملاحظة: أظهرت العينات التي اعتبرت ذات نوعية جيدة للتسلسل شكل نوى مستديرة ومنتظمة تحت المجهر بدون حطام خلوي (الشكل 2 د). إن وجود هالة حول الغشاء النووي أو تجميع نوى متعددة معا هي علامات على وجود نوى تالفة ولا ينبغي النظر في مثل هذه المعلقات الخلوية ل snRNA-seq (الشكل 2D). كان التركيز المقاس للنوى في حدود 300-700 نواة / ميكرولتر.
5. إعداد المكتبة وتسلسلها
ملاحظة: يعتمد وصف الخطوات التالية على منصة التسلسل الداخلية المستخدمة في هذه الدراسة (انظر جدول المواد). لذلك ، قد تختلف بعض الإعدادات عند استخدام نظام أساسي مختلف. هنا ، يتم وصف الخطوات الرئيسية فقط ويجب تحديد كل معلمة باتباع الإرشادات والبروتوكولات من الشركة المصنعة المختارة وإن كان ذلك مع التحسين قبل الاستخدام الأول. من الأهمية بمكان التأكد من أن إعداد المكتبات يتم في أسرع وقت ممكن بعد تركيز معلقات النوى المصنفة لتجنب تدهور الحمض النووي الريبي وضمان الجودة المثلى للتسلسل.
- قم بتحميل ما بين 7000 و 10000 نواة في شريحة أحادية الخلية للموائع الدقيقة.
- تقسيم النوى المحملة في قطرات مقياس نانولتر باستخدام وحدة التحكم المقدمة والكواشف من المورد المختار. Lyse nucles داخل كل قطرة ونسخ عكسي للحمض النووي الريبي.
ملاحظة: داخل قطرة ، تشترك جميع cDNA الناتجة في نفس الرمز الشريطي للخلية. - إعداد المكتبات ل snRNA-seq باتباع إرشادات المورد المختار وضمان التوافق مع منصة التسلسل. تحقق من جودة وتركيز المكتبات النهائية باستخدام الرحلان الكهربائي أو القياس الفلوري أو الطرق القائمة على qPCR ، وإذا أمكن ، قم بتجميعها بشكل متساو قبل التسلسل.
- قامت Denature بتجميع مكتبات التعبير الجيني 3ʹ وتخفيفها وفقا لتوصية الشركة المصنعة.
- قم بإجراء تسلسل فهرسة مزدوج أو أحادي أو مزدوج على منصة تسلسل من الجيل التالي بعمق تسلسل يبلغ 50000 زوج قراءة لكل خلية.
النتائج
يصف البروتوكول المقدم هنا طريقة لإعداد تعليق لنوى مفردة غير عصبية معزولة من DG لأداء snRNA-seq. مع أو بدون FANS ، كشف التجميع المعلوماتي الحيوي عن مجموعات منفصلة جيدا من النوى تتوافق مع أنواع الخلايا المعروفة داخل DG (الشكل 2E ، F). ضمن العينة غير المصنفة بواسطة FACS ، تألفت غالبية النوى عالية الجودة التي تم تسلسلها من ثلاث مجموعات من الخلايا العصبية (84.9٪ من إجمالي النوى لهذه العينة ، الشكل 2E ، G ، H). من المتوقع حدوث مثل هذه النتائج ، مع الأخذ في الاعتبار أن مجموعات الخلايا الأكثر تمثيلا في DG هي الخلايا العصبية الحبيبية ، والخلايا العصبية المثيرة الأخرى (الخلايا العصبية الاستثارية المسماة) ، والخلايا العصبية المثبطة10. كانت المجموعات غير العصبية التي تم تحديدها تتكون في الغالب من أنواع الخلايا الدبقية (11.1٪) ، بما في ذلك الخلايا النجمية والخلايا قليلة التغصن وخلايا السلائف قليلة التغصن (OPCs) والخلايا المناعية (3.3٪) وخلايا Cajal-Retzius (0.6٪). عند إجراء FANS لاستبعاد السكان الإيجابيين ل NeuN (عينة مصنفة من NeuN سلبية FACS ؛ الشكل 2F ، G ، H) ، أصبحت مجموعات الخلايا الدبقية سائدة (81.3٪). يسمح عزل عدد أكبر من النوى الدبقية بتقسيم أفضل للمجموعات المختلفة التي من شأنها أن تتجمع معا بدون FANS. في الواقع ، عند إعادة تجميع وتحليل جينات معينة إما معبرا عنها في NSCs أو في الخلايا النجمية ، انفصلت أربع مجموعات فرعية (الشكل التكميلي 2 أ ، ب). بالنظر إلى علامات خلوية أكثر تحديدا وتقييم مستويات التعبير الجيني عبر أنواع الخلايا ، تم اكتشاف مجموعة صغيرة من NSCs منفصلة بشكل منفصل عن مجموعات الخلايا النجمية الرئيسية مع تعبير أعلى عن Hopx و Notch2 وتقريبا لا يوجد تعبير عن Aldh1a1 أو Aqp4 (الشكل التكميلي 2C). ومع ذلك ، بسبب التداخل في التعبير الجيني بين الخلايا النجمية و NSCs ، ستكون هناك حاجة إلى مزيد من التحليل لتحديد وتحديد أنواع فرعية مختلفة من الخلايا على وجه التحديد. علاوة على ذلك ، كانت عينة FANS سلبية NeuN تحتوي على مجموعات إضافية مصنفة على أنها خلايا وعائية (2.3٪) تشمل الخلايا البطانية ، والخلايا المحيطة ، والخلايا الوعائية عند الرجوع إليها للتعبير عن علامات خاصة بالخلية (البيانات غير معروضة).
باتباع الإرشادات الخاصة بالبروتوكول المختار لإنشاء مكتبات للتسلسل ، تم الحصول على ملفات تعريف تعبير عالية الجودة مع أو بدون FANS. بالنسبة للعينات المتسلسلة عند 50000 قراءة / نواة ، تم اكتشاف 2510 جينا في المتوسط لكل نواة للعينة غير المصنفة بواسطة FACS (5578 نسخة ، الشكل 2H) و 1665.5 جينا (3508 نسخة) لعينة FANS سلبية NeuN ، بعد تصفية نوى منخفضة الجودة (الشكل 2H ، I). تؤكد هذه المقاييس أن هذا البروتوكول يولد تنميطا نسخيا عالي الجودة لنوى مفردة يمكن مقارنته بالدراسات التي تستخدم مناهج مختلفة22,23 وأن عملية فرز FACS لا تلحق الضرر بالنوى ل snRNA-seq اللاحقة. والجدير بالذكر أن الاختلاف في عدد الجينات والنسخ لكل نواة بين العينتين لا يرجع إلى انخفاض جودة البيانات ولكن إلى ارتفاع نسبة الخلايا العصبية في العينة غير المصنفة من قبل FACS (84.9٪ مقارنة ب 1.7٪ في عينة FANS سلبية NeuN) ، والتي لها نشاط نسخ أعلى (2,660 جينا / نواة و 6,170 نسخة / نواة في عينة غير مصنفة من قبل FACS) من متوسط نشاط النسخ لجميع أنواع الخلايا غير العصبية (1,090 جينا / نواة و 1,785 النصوص / النواة ، الشكل 2I).
تظهر هذه النتائج التمثيلية معا أن اختيار نوى NeuN السلبية باستخدام FANS هو أداة قوية لعزل أنواع الخلايا منخفضة الوفرة من أنسجة المخ التي تم تشريحها حديثا وإجراء تنميط نسخي أحادي النوى عالي الجودة لمجموعات الخلايا المتميزة هذه عبر طرق snRNA-seq.
الشكل التكميلي 1: التحقق من صحة التلوين المناعي ل FANS. تم تحضين معلق النوى (A) بدون الجسم المضاد NeuN-AF 488 كعنصر تحكم سلبي أو (B) مع الجسم المضاد وتشغيله من خلال فارز FACS للتحقق من صحة ظروف التلوين المناعي. الرجاء الضغط هنا لتنزيل هذا الملف.
الشكل التكميلي 2: تحليل التعبير الجيني وإعادة تجميع مجموعة الخلايا النجمية. (أ) مخطط تقريب وإسقاط مشعب موحد لتقليل الأبعاد (UMAP) يوضح تجميع 4968 نواة بناء على ملفات تعريف التعبير على مستوى الجينوم من الشكل 2F. تم إجراء مكالمات من نوع الخلية بناء على علامات نوع الخلية. (ب) تتكون مجموعة الخلايا النجمية من 2579 نواة تم اختيارها من (أ) لمزيد من الإعداد الفرعي لدراسة الأنواع الفرعية الخلوية المحتملة. تم الكشف عن أربعة أنواع فرعية بواسطة تجميع سورات (0-3) ، موضحة بألوان مختلفة. ج: مستويات التعبير الجيني لواسمات خلوية محددة عبر أنواع الخلايا الأربعة. تم الحصول على جميع قطع الأراضي باستخدام حزمة Seurat R24. باختصار ، تم تسوية أعداد RNA-seq لكل خلية من خلال التعبير الكلي وضربها في عامل المقياس (10000). ثم تم تحويل هذه النتيجة. تم قياس القيم المحولة (تم قياس التباين إلى واحد) وتوسيطها (متوسط الضبط على الصفر) داخل كل خلية قبل تطبيق UMAP لحساب التضمينات ، والتي تم استخدامها كقيم على محوري x و y. تمثل الرسوم البيانية ناتج تقنية تقليل الأبعاد على مخطط مبعثر 2D حيث تمثل كل نقطة خلية ذات إحداثيات x و y بناء على تضمين الخلية التي تحددها تقنية الاختزال. يتم وضع الخلايا ذات التوقيعات الجينية المتشابهة بالقرب من بعضها البعض بواسطة عمليات التضمين. الرجاء الضغط هنا لتنزيل هذا الملف.
الشكل التكميلي 3: تحليل التعبير الجيني ل NeuN في السلالة العصبية. (أ) مخطط UMAP يوضح تجميع النسب العصبي من مجموعة البياناتالمتاحة للجمهور 15. تم إنشاء UMAPs كما في الشكل التكميلي 2. (ب) مستويات التعبير الجيني لعلامات خلوية محددة عبر السلالة العصبية تظهر الخلايا النجمية (Aquaporin 4 = Aqp4) ، NSCs (بروتين المجال المثلي فقط = Hopx) ، NeuN / Rbfox3 (NSCs والخلايا السلفية الوسيطة [IPCs]) ، وخلايا الدراجات (كيناز المعتمد على السيكلين 6 = Cdk6). الرجاء الضغط هنا لتنزيل هذا الملف.
الجدول التكميلي 1: تركيبات الوسائط والمخازن المؤقتة المستخدمة في الدراسة. الرجاء الضغط هنا لتنزيل هذا الملف.
Discussion
لتنفيذ هذا البروتوكول بنجاح ، يعد تشريح DG الخطوة الحاسمة الأولى ، والتي تتطلب بعض الممارسة لإبقائه غير تالف والحد من التلوث من الأنسجة المحيطة. من التجربة ، يمكن الحصول على فصل DG عن الحصين بسرعة كبيرة من قبل باحث ماهر يمكنه بعد ذلك العمل على تحسين تقنيته لزيادة سرعة التشريح وبالتالي تحسين نضارة الأنسجة لتوليد بيانات عالية الجودة. وعلى نفس المنوال ، يتطلب تحضير وإعادة تعليق النوى المفردة الاتساق عبر الظروف المختلفة المستخدمة في تجربة واحدة ، ولكن أيضا تجنب السحب المفرط الذي يمكن أن يعطل الغشاء النووي الذي يطلق الحمض النووي الريبي المحيط الذي سيؤدي إلى تحيز نتائج التسلسل. بالإضافة إلى التوصيات المذكورة سابقا لإعداد نوى عالية الجودة ، يجب أيضا النظر في تركيز معلق النواة المفردة قبل الشروع في التسلسل. في الواقع ، وفقا لإرشادات الشركة المصنعة ، يجب تخفيف المستحضر بتركيز أعلى من 1200 nuc / μL ، لأن هذا المستوى من تركيز النوى سيكون أكثر عرضة لتشكيل مضاعفات تؤثر على تحليلات المعلوماتية الحيوية النهائية. وتجدر الإشارة إلى أن تسلسل العينات بتركيزات نوى أقل من 500 نيومتر مكعب / ميكرولتر قد لا يكون مجديا بسبب التكلفة المتضمنة. يوصى أيضا باتباع نصيحة مستخدم متقدم لنظام مراقبة الأصول الميدانية لإعداد جميع البوابات والبقاء متسقا مع الإعدادات عبر العينات والتكرارات البيولوجية. وبالمثل ، فإن إعداد المكتبات لتسلسل الحمض النووي الريبي يستلزم بعض التدريب لتحقيق نتائج عالية الجودة ومعظم البائعين لديهم دعم ممتاز لتحقيق ذلك بكفاءة. تم اختبار هذه الطريقة فقط مع الأنسجة الطازجة في هذه الدراسة. ومع ذلك ، تم إجراء FANS أيضا مع الأنسجة المجمدة25. لذلك من المعقول أن نفترض أن هذا البروتوكول يمكن إجراؤه باستخدام الأنسجة المجمدة وإن كان ذلك مع تحسين طفيف.
تم تطوير هذا البروتوكول مع وضع تطبيق معين في المصب في الاعتبار ، وهو التحقيق في مجموعات الخلايا بخلاف الخلايا العصبية داخل مكانة الحصين العصبية. في الواقع ، تشير خطوط الأدلة المتزايدة إلى أن ضعف AHN في الشيخوخة يمكن أن يعزى إلى الخلايا المحيطة داخل الكوة1،2،3،9. على وجه الخصوص ، تظهر الخلايا النجمية والخلايا قليلة التغصن كمنظم رئيسي ل AHN. ومع ذلك ، فإن عزلهم عن DG إلى جانب تسلسل الحمض النووي الريبي قد أدى إلى نتائج مختلطة ، مما يجعل هذه الفرضية صعبة التقييم باستخدام هذه التقنية 1,17. سمح هذا النهج لفرز نوى NeuN السلبية FACS بعزل المزيد من الخلايا النجمية والخلايا قليلة التغصن مقارنة بالعينات التي لم يتم فرزها بواسطة FACS ، مما يتيح تحليلا معلوماتيا حيويا أفضل. هذا البروتوكول قابل للتطبيق في جميع الأعمار عبر العمر ، وتوفر البيانات التمثيلية المقدمة هنا مع أنسجة من الحيوانات القديمة دليلا على مفهوم أن هذه الطريقة قوية للتحقيق في مكانة الحصين العصبية الهرمة. لتوسيع استخدام هذه الطريقة وتكييفها مع الأسئلة البيولوجية المختلفة ، من المهم مراعاة أنه يمكن اختبار مستضدات الغشاء النووي العصبي الأخرى مع معايرة شاملة لأفضل الأجسام المضادة التي تم التحقق من صحتها لهذه العلامات. على سبيل المثال ، عند دراسة عملية التمايز العصبي من NSCs في DG ، تبدأ بعض أنواع الخلايا مثل خلايا النوع 2 أو الخلايا العصبية في التعبير عن NeuN (الشكل التكميلي 3). ومن ثم، ستكون هناك حاجة إلى مولد ضد آخر لدراسة هذه الأنواع من الخلايا على وجه التحديد. على العكس من ذلك ، لا يزال يتم تحديد بعض الخلايا العصبية في هذه الدراسة بعد فرز FACS السلبي ل NeuN ربما بسبب انخفاض أو عدم وجود تعبير عن NeuN في هذه المجموعات السكانية (على سبيل المثال ، الخلايا العصبية القشرية Cajal-Retzius19). بالإضافة إلى ذلك ، تم الإبلاغ عن NeuN في مجموعات فرعية من oligodendrocytes26 ، والتي يمكن أن تعطي نتائج متحيزة إذا كانت هذه المجموعات الفرعية ذات أهمية. وبالتالي ، ينبغي النظر بعناية في اختيار المستضد عند البدء في استخدام FANS لتجنب إدراج أو استبعاد مجموعات الخلايا التي من شأنها أن تحول دون إجابة دقيقة على سؤال بيولوجي محدد. بالاتفاق مع هذا ، يوصى أيضا بالتحقق من صحة كل نتيجة تسلسل من خلال المقايسات المتعامدة (على سبيل المثال ، الكيمياء المناعية أو نطاق الحمض النووي الريبي) قبل التحقق من صحة أو دحض الفرضية المختبرة باستخدام هذا البروتوكول. أخيرا ، يمكن تطوير الخطوة التي تتضمن FANS لتشمل أكثر من جسم مضاد واحد مع استراتيجية فرز أكثر تفصيلا لاستبعاد و / أو تضمين مجموعات الخلايا المرغوبة.
في نهاية المطاف ، يمكن أن يكون للتكنولوجيات الموصوفة في هذا البروتوكول بعض القيود عند استخدامها مع الأنواع الأخرى. على سبيل المثال ، يتم تعريف المكانة بشكل جيد للغاية في القوارض مع وجود NSCs التكاثري والهادئ أو الخلايا العصبية المولودة حديثا المقيدة داخل مناطق فرعية محددة من DG ، ولكن لا يزال من غير الواضح كيف يجب تحديد مكانة الحصين العصبية في الأنواع الأخرى. في الواقع ، لا يتم محاذاة الخلايا التكاثرية داخل منطقة متصلة من DG في الرئيسيات غير البشرية والبشر ولكنها منتشرة حولها وقد تكون موجودة أيضا في اللوزة7. لذلك ، فإن تشريح وعزل مناطق أوسع من DG في الأنواع الأخرى من المحتمل أن يؤثر على استخدام هذا البروتوكول. على وجه الخصوص ، يجب تحسين خطوات التفكك والتثليج لإعداد الأنسجة أثناء العمل مع قطع أكبر من الأنسجة27,28. فيما يتعلق بالتحليل المعلوماتي الحيوي ، في حين أن القوارض التي تحتوي على سلالات متجانسة للغاية ومشروحة جيدا ، فإن التباين الجيني للجينوم البشري جنبا إلى جنب مع الأعداد غير الكافية من العلامات الخلوية للتمييز بوضوح بين مجموعات الخلايا المختلفة (على سبيل المثال ، NSCs والخلايا النجمية) يتطلب الكثير من التطبيع للتحليل الذي يمكن أن يؤدي إلى استنتاجات مختلفة عند تحديد مجموعة صغيرة من الخلايا7 ، 11. في مثل هذه الحالات ، قد يظل إثراء الخلايا خيارا مفضلا أو يجب استخدامه جنبا إلى جنب مع استراتيجيات أخرى لزيادة القوة التحليلية.
ومع ذلك ، يمكن للنهج الحالي أن يتيح التحقيق في دور مجموعات الخلايا غير المدروسة وإن كانت مهمة في تنظيم AHN. يمكن أن يكون هذا هو الحال بشكل خاص بالنسبة لمجموعات الخلايا النجمية ، التي تلعب دورا مركزيا في ظهور وتطور الأمراض التنكسية العصبية29,30. أظهرت هذه الدراسة أنه يمكن تحديد الخلايا النجمية وغيرها من مجموعات الخلايا النادرة وتوصيفها ببساطة عن طريق استبعاد الغالبية العظمى من الخلايا العصبية الموجودة داخل DG. لم تتمكن الدراسات الأخرى التي تستخدم مناهج مختلفة من تحقيق استرداد مماثل للنوى من نفس النطاق من مجموعات الخلايا5،11،17. علاوة على ذلك ، تظهر نتائج هذه الدراسة أنه من الممكن استخدام هذا النهج لعزل مجموعة NSC دون إثراء محدد لمجتمع الخليةهذا 15.
في الختام ، سيكون اتباع هذه الطريقة وتحسينها خطوة إلى الأمام لمعالجة الأسئلة المعلقة المتعلقة بالدور السياقي للمكانة العصبية الحصينية لتعديل AHN. على وجه الخصوص ، يمكن أن يجلب رؤى جديدة حول مستويات التعبير الجيني في الأدمغة المسنة والمريضة في مجموعات الخلايا المرتبطة بتنظيم AHN9 ، أو يدعم تحديد عدم التجانس المحتمل ل NSCs1 أو يعالج دور الأوعية الدموية في AHN. في النهاية ، يمكن تكييف هذه الطريقة مع منافذ الخلايا الجذعية البالغة الأخرى ذات الأسئلة والقضايا المماثلة.
Disclosures
SG و TL و SK هم موظفون في Merck Sharp & Dohme LLC ، وهي شركة تابعة لشركة Merck & Co.، Inc. ، Rahway ، NJ ، الولايات المتحدة الأمريكية المعروفة باسم MSD خارج الولايات المتحدة وكندا. SG هي أحد المساهمين في شركة Merck & Co.، Inc. ، راهواي ، نيوجيرسي ، الولايات المتحدة الأمريكية.
Acknowledgements
يود المؤلفون أن يشكروا لاكلان هاريس وبييرو ريغو على الدعم الفني وجيسون إم أوسلانر وديت لوفات على تقديم ملاحظاتهم على المخطوطة. تم دعم هذا العمل من خلال دعم المنح من MRC والتعاون البحثي قبل المنافسة مع MSD ، ومعهد فرانسيس كريك ، الذي يتلقى تمويله من أبحاث السرطان في المملكة المتحدة (FC0010089) ، ومجلس البحوث الطبية في المملكة المتحدة (FC0010089) ، و Wellcome Trust (FC0010089) وجائزة Wellcome Trust Investigator Award إلى FG (106187 / Z / 14 / Z). نعتذر للعديد من المؤلفين الذين لم نتمكن من مناقشة أعمالهم والاستشهاد بها بسبب ضيق المساحة.
Materials
| Name | Company | Catalog Number | Comments |
| 0.5ml microtube | Eppendorf | 30124537 | |
| 10.00µm Flouresbrite YG Carboxylate Microspheres | Polysciences | 15700-10 | |
| 15 mL polypropylene centrifuge tubes | Corning | 430052 | |
| 2 pairs of sterile Dumont #5 forceps | Fine Science Tools | 11252-30 | |
| 4′,6-diamidino-2-phenylindole (DAPI) | Sigma Aldrich | D9564-10MG | |
| 4150 TapeStation System | Agilent | N/A | |
| 5 mL round bottom high clarity polypropylene test tube with snap cap | Falcon | 352063 | |
| 5 mL round bottom polystyrene test tube with cell strainer snap cap | Falcon | 352235 | |
| 50 mL polypropylene centrifuge tubes | Corning | 430829 | |
| 70 µm cell strainer | Falcon | 352350 | |
| 8 peak SPHERO Rainbow Calibration Particles | BD Biosciences | RCP-30-5A | |
| Accudrop Beads | BD Biosciences | N/A | |
| Allegra X-30R Centrifuge | Beckman Coulter | N/A | |
| Anti-NeuN antibody, clone A60, Alexa Fluor 488 conjugated | Millipore | MAB377X | |
| BD FACSAria Fusion Flow Cytometer | BD Biosciences | N/A | |
| Beckman Coulter MoFlo XDP | Beckman Coulter | N/A | |
| Chromium Controller | 10x Genomics | N/A | |
| Chromium Next GEM Single Cell 3' Reagent Kits v3.1 | 10x Genomics | PN-1000121; PN-1000120; PN-1000213 | |
| BSA 7.5% | Gibco | 15260037 | |
| Dithiothreitol (DTT) | Thermo Scientific | R0861 | |
| Dounce tissue grinder set: mortar, loose pestle (A) and tight pestle (B) | KIMBLE | D8938-1SET | |
| Eppendorf Tubes Protein LoBind 1.5ml | Eppendorf | 30108116 | |
| Halt, 100x Protease inhibitor | ThermoFisher | 78429 | |
| HiSeq 4000 Sequencing System | Illumina | N/A | Sequencing configuration: 28-8-0-91 |
| KCl | Any chemical supplier | Laboratory made | |
| LUNA-FX7 Automated Cell counter | Logos Biosystems | N/A | |
| MgCl2 | Any chemical supplier | Laboratory made | |
| N°10 guarded sterile disposable scalpels | Swann-Morton | 6601 | |
| Nuclease-free water | Sigma Aldrich | W4502-1L | |
| Pair of sterile student surgical scissors | Fine Science Tools | 91401-12 | |
| PBS | Any chemical supplier | Laboratory made | |
| RNase Inhibitor 40 U µl-1 | Ambion | AM2684 | |
| RNasin 40 U µl-1 | Promega | N211A | |
| Sterile Petri dish | Corning | 430167 | |
| Sucrose | Sigma Aldrich | 59378-500G | |
| Tris buffer, pH 8.0 | Any chemical supplier | Laboratory made | |
| Triton X-100 10% (v/v) | Sigma Aldrich | T8787-250ML | |
| Trypan blue | Invitrogen | T10282 |
References
- Gillotin, S. Targeting impaired adult hippocampal neurogenesis in ageing leveraging intrinsic mechanisms regulating neural stem cell activity. Ageing Research Reviews. 71, 101447 (2021).
- Urban, N., Blomfield, I. M., Guillemot, F. Quiescence of adult mammalian neural stem cells: A highly regulated rest. Neuron. 104 (5), 834-848 (2019).
- Hanspal, M. A., Gillotin, S. A new age in understanding adult hippocampal neurogenesis in Alzheimer's disease. Neural Regeneration Research. 17 (12), 2615-2618 (2022).
- Zhang, H., et al. Single-nucleus transcriptomic landscape of primate hippocampal aging. Protein & Cell. 12 (9), 695-716 (2021).
- Franjic, D., et al. Transcriptomic taxonomy and neurogenic trajectories of adult human, macaque, and pig hippocampal and entorhinal cells. Neuron. 110 (3), 452-469 (2022).
- Moreno-Jimenez, E. P., Terreros-Roncal, J., Flor-Garcia, M., Rabano, A., Llorens-Martin, M. Evidences for adult hippocampal neurogenesis in humans. Journal of Neuroscience. 41 (12), 2541-2553 (2021).
- Sorrells, S. F., et al. Positive controls in adults and children support that very few, if any, new neurons are born in the adult human hippocampus. Journal of Neuroscience. 41 (12), 2554-2565 (2021).
- Zhou, Y., et al. Molecular landscapes of human hippocampal immature neurons across lifespan. Nature. 607, 527-533 (2022).
- Bonafina, A., Paratcha, G., Ledda, F. Deciphering new players in the neurogenic adult hippocampal niche. Frontiers in Cell and Developmental Biology. 8, 548 (2020).
- Amaral, D. G., Scharfman, H. E., Lavenex, P. The dentate gyrus: fundamental neuroanatomical organization (dentate gyrus for dummies). Progress in Brain Research. 163, 3-22 (2007).
- Habib, N., et al. Massively parallel single-nucleus RNA-seq with DroNc-seq. Nature Methods. 14 (10), 955-958 (2017).
- Flor-Garcia, M., et al. Unraveling human adult hippocampal neurogenesis. Nature Protocols. 15 (2), 668-693 (2020).
- Moreno-Jimenez, E. P., et al. Adult hippocampal neurogenesis is abundant in neurologically healthy subjects and drops sharply in patients with Alzheimer's disease. Nature Medicine. 25 (4), 554-560 (2019).
- Kalinina, A., Lagace, D. Single-cell and single-nucleus RNAseq analysis of adult neurogenesis. Cells. 11 (10), 1633 (2022).
- Harris, L., et al. Coordinated changes in cellular behavior ensure the lifelong maintenance of the hippocampal stem cell population. Cell Stem Cell. 28 (5), 863-876 (2021).
- Shin, J., et al. Single-cell RNA-seq with Waterfall reveals molecular cascades underlying adult neurogenesis. Cell Stem Cell. 17 (3), 360-372 (2015).
- Artegiani, B., et al. A single-cell RNA sequencing study reveals cellular and molecular dynamics of the hippocampal neurogenic niche. Cell Reports. 21 (11), 3271-3284 (2017).
- Nott, A., Schlachetzki, J. C. M., Fixsen, B. R., Glass, C. K. Nuclei isolation of multiple brain cell types for omics interrogation. Nature Protocols. 16 (3), 1629-1646 (2021).
- Sarnat, H. B., Nochlin, D., Born, D. E. Neuronal nuclear antigen (NeuN): a marker of neuronal maturation in early human fetal nervous system. Brain Development. 20 (2), 88-94 (1998).
- . Guidance on the Operation of ASPA Available from: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_dat/file/662364/Guidance_on_the_Operation_of_ASPA.pdf (2022)
- Hagihara, H., Toyama, K., Yamasaki, N., Miyakawa, T. Dissection of hippocampal dentate gyrus from adult mouse. Journal of Visualized Experiments. (33), e1543 (2009).
- Habib, N., et al. Disease-associated astrocytes in Alzheimer's disease and aging. Nature Neuroscience. 23 (6), 701-706 (2020).
- Ding, J., et al. Systematic comparison of single-cell and single-nucleus RNA-sequencing methods. Nature Biotechnology. 38 (6), 737-746 (2020).
- Hao, Y., et al. Integrated analysis of multimodal single-cell data. Cell. 184 (13), 3573-3587 (2021).
- Mussa, Z., Tome-Garcia, J., Jiang, Y., Akbarian, S., Tsankova, N. M. Isolation of adult human astrocyte populations from fresh-frozen cortex using fluorescence-activated nuclei sorting. Journal of Visualized Experiments. (170), e62405 (2021).
- Zhang, Y., et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. Journal of Neuroscience. 34 (36), 11929-11947 (2014).
- Marti-Mengual, U., Varea, E., Crespo, C., Blasco-Ibanez, J. M., Nacher, J. Cells expressing markers of immature neurons in the amygdala of adult humans. European Journal of Neuroscience. 37 (1), 10-22 (2013).
- Zhang, X. M., et al. Doublecortin-expressing cells persist in the associative cerebral cortex and amygdala in aged nonhuman primates. Frontiers in Neuroanatomy. 3, 17 (2009).
- Ding, Z. B., et al. Astrocytes: a double-edged sword in neurodegenerative diseases. Neural Regeneration Research. 16 (9), 1702-1710 (2021).
- Phatnani, H., Maniatis, T. Astrocytes in neurodegenerative disease. Cold Spring Harbor Perspectives in Biology. 7 (6), 020628 (2015).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved