
Method Article
מיון שלילי של גרעינים המופעלים על ידי פלואורסצנציה של נוירונים בשילוב עם ריצוף RNA של גרעין יחיד כדי לחקור את הנישה הנוירוגנית של ההיפוקמפוס
In This Article
Summary
מוצגת כאן שיטה לרצף גרעינים בודדים שבודדו מהפיתול המשונן של העכבר, שאינה כוללת את רוב תאי העצב באמצעות מיון גרעינים המופעלים על ידי פלואורסצנציה (FAN). גישה זו מייצרת פרופילי ביטוי איכותיים ומקלה על המחקר של רוב סוגי התאים האחרים המיוצגים בנישה, כולל אוכלוסיות נדירות כגון תאי גזע עצביים.
Abstract
נוירוגנזה היפוקמפלית בוגרת (AHN), המורכבת מתחזוקה לכל החיים של תאי גזע עצביים מתרבים ושקטים (NSCs) בתוך האזור התת-גרגירי (SGZ) של הפיתול המשונן (DG) והתמיינותם מנוירונים שזה עתה נולדו לתאי גרגיר בשכבת תאי הגרגיר, מאומתת היטב במחקרים רבים. שימוש בבעלי חיים מהונדסים גנטית, במיוחד מכרסמים, הוא כלי רב ערך לחקור מסלולי איתות המווסתים AHN ולחקור את התפקיד של כל סוג תא המרכיבים את הנישה הנוירוגנית בהיפוקמפוס. כדי לענות על כך, לשיטות המשלבות בידוד גרעינים בודדים עם ריצוף מהדור הבא הייתה השפעה משמעותית בתחום ה-AHN לזיהוי חתימות גנים עבור כל אוכלוסיית תאים. עם זאת, יש צורך בעידון נוסף של טכניקות אלה כדי ליצור פרופיל פנוטיפי של אוכלוסיות תאים נדירות יותר בתוך ה-DG. כאן אנו מציגים שיטה המשתמשת במיון גרעינים מופעלים פלואורסצנטיים (FANS) כדי להוציא את רוב אוכלוסיות הנוירונים מתרחיף גרעינים יחיד שבודד מ-DG שזה עתה נותח, על ידי בחירת גרעינים לא מוכתמים עבור האנטיגן NeuN, על מנת לבצע ריצוף RNA של גרעינים בודדים (snRNA-seq). שיטה זו היא אבן דרך פוטנציאלית לחקור עוד יותר את הוויסות הבין-תאי של ה-AHN ולחשוף סמנים ומנגנונים תאיים חדשים בין מינים.
Introduction
הדור המתמשך של נוירונים בהיפוקמפוס בבגרות, הידוע גם בשם נוירוגנזה בהיפוקמפוס למבוגרים (AHN), קשור לתפקודים קוגניטיביים כגון למידה, רכישה/פינוי זיכרון והפרדת דפוסים, ונראה כי הוא מנגנון חשוב של עמידות בהזדקנות ומחלות ניווניות של מערכת העצבים למניעת ליקויים קוגניטיביים 1,2,3 . מכרסמים היו המודל המועדף לחקור AHN באמצעות מספר שיטות, כולל אימונוציטוכימיה ושיטות ריצוף מהדור הבא (NGS). התרגום של תוצאות אלה למינים אחרים נותר שנוי במחלוקת. ואכן, AHN נצפה ברוב המינים, אך המידה שבה הוא נמשך לאורך כל החיים, במיוחד אצל בני אדם 4,5,6,7,8, נתונה לוויכוח קבוע.
עד כה, מסלולי איתות פנימיים וחיצוניים שונים אושרו לווסת AHN1. עם זאת, ההשפעה של תקשורת בין-תאית על AHN רק מתעוררת9. ניתן לייחס זאת תחילה לספציפיות לא מספקת של סמני התאים הידועים כיום כדי לבצע ניתוח in vivo עם בעלי חיים מהונדסים גנטית. ואכן, מחקרים רבים הסתמכו על סמנים כגון דאבלקורטין או חלבון חומצי פיברילארי גליאלי (GFAP) המתבטאים במספר סוגי תאים1. שנית, המורכבות והרמה הגבוהה של מגוון התאים בנישה ההיפוקמפלית הבוגרת10 מביאה אתגרים טכניים לפרופיל כל סוג תא. זה המקרה במיוחד עבור ניתוח ביואינפורמטי עם סמנים תאיים חופפים המשמשים בצינורות אנליטיים עבור אוכלוסיות שונות, כגון תאי גזע עצביים או תאי גליה, וכתוצאה מכך מסקנות שנויות במחלוקת בעת הערכת AHN 7,11. שלישית, המספר העצום של תאי עצב מערער את החקירה של אוכלוסיות תאים פחות בשפע, כגון אסטרוציטים, אוליגודנדרוציטים או תאים אפנדימליים, למרות שתפקידם בוויסות הכוונון העדין של AHN הופך להיות בולט9. יחד, מגבלות אלה משפיעות על היכולת לתרגם תוצאות של מכרסמים למינים אחרים. הדבר מועצם במיוחד על ידי הקושי לשחזר במבחנה רקמה מורכבת, כגון הנישה הנוירוגנית בהיפוקמפוס, ועל ידי המכשולים הרבים לגישה לרקמות באיכות גבוהה יחד עם היעדר פרוטוקולים סטנדרטיים לעיבוד רקמות במחקרים המערבים רקמות אנושיות12,13. לכן חיוני לפתח גישות חדשות לפרופיל אוכלוסיות תאים ולזיהוי סמנים תאיים חדשים בתוך הפיתול המשונן (DG) שבסופו של דבר יובילו להבנה טובה יותר של התרומות השונות של כל סוג תא לוויסות AHN.
כדי להשיג זאת, בידוד של תאים בודדים (sc) וגרעינים בודדים (sn) בשילוב עם ריצוף RNA הפך לחיוני לחקר רקמות מורכבות כגון DG14. לפיכך, אסטרטגיות של העשרת תאים לבידוד תאים בודדים מנישה היפוקמפלית בוגרת של עכברים בוצעו בעיקר כדי לבחון תאי גזע עצביים15,16. אסטרטגיה מעניינת להעשרת תאים שאינם נוירונים מה-DG יושמה על ידי ריצוף GluR1/Cd24 תאים בודדים שליליים כפולים שהביאו לריצוף של 1,408 תאים ללא אשכולות מובחנים בין אסטרוציטים לתאי גזע עצביים לאחר ניתוח ביואינפורמטי17. ייתכן שהסיבה לכך היא העיכול האנזימטי הקשה הנדרש להכנת תא בודד שפוגע בשלמות התא וב-RNA. כדי לעקוף בעיה טכנית זו, פותחו מספר שיטות המשתמשות בבידוד גרעינים בודדים במקום זאת והן מתאימות במיוחד לרקמות מורכבות11,18. עם זאת, הדומיננטיות של תאי עצב בתוך ה-DG או באופן רחב יותר בתוך מערכת ההיפוקמפוס-אנטורינל יוצרת הטיית דגימה כדי לחקור את מכלול אוכלוסיות התאים הקיימות באזורי מוח אלה. בנוסף, המספר המצומצם של תאים לטעינה לצורך הכנת ספריות תאים בודדים מדגיש את נוכחותה של אוכלוסיית התאים העיקרית בצינורות אנליטיים של גרעינים בודדים מרוצפים. ואכן, אשכולות נוירונים גדולים עוברים לעתים קרובות ביאורים וניתוחים, בעוד שאוכלוסיות תאים אחרות סובלות מתת-ייצוג או מפספסות 5,11.
בניסיון להתגבר על הטיות אלה ולהיות מסוגלים ליצור פרופיל של סוגי תאים שאינם תאי עצב הנמצאים ב-DG של העכבר, הומצאה שיטה במחקר זה תוך שימוש בעקרון של מיון גרעינים מופעלים פלואורסצנטיים (FANS)18 שאינה כוללת את רוב האוכלוסיות העצביות על ידי בחירה שלילית של גרעינים בודדים מוכתמים עם אנטיגן גרעיני עצבי (NeuN, ידוע גם בשם Rbfox3). בחירה זו באנטיגן הונחתה על ידי הספרות המתארת את NeuN כסמן עצבי אמין19 ועל ידי הצורך להשתמש בחלבון גרעיני לגישה זו. לאחר מכן הוכנו תאים ממוינים FACS שליליים ל-NeuN לריצוף RNA על פלטפורמת 10x Genomics. התוצאות מראות כי הדרה של תאים המבטאים NeuN מאפשרת פרופיל תעתיקי ספציפי ואיכותי מסוג תא של אוכלוסיות תאי גליה ותאים נדירים.
Protocol
טיפול בבעלי חיים והליכים ניסיוניים בוצעו בהתאם להנחיות מכון פרנסיס קריק, כמו גם להנחיות ולחוקים של משרד הפנים הבריטי.

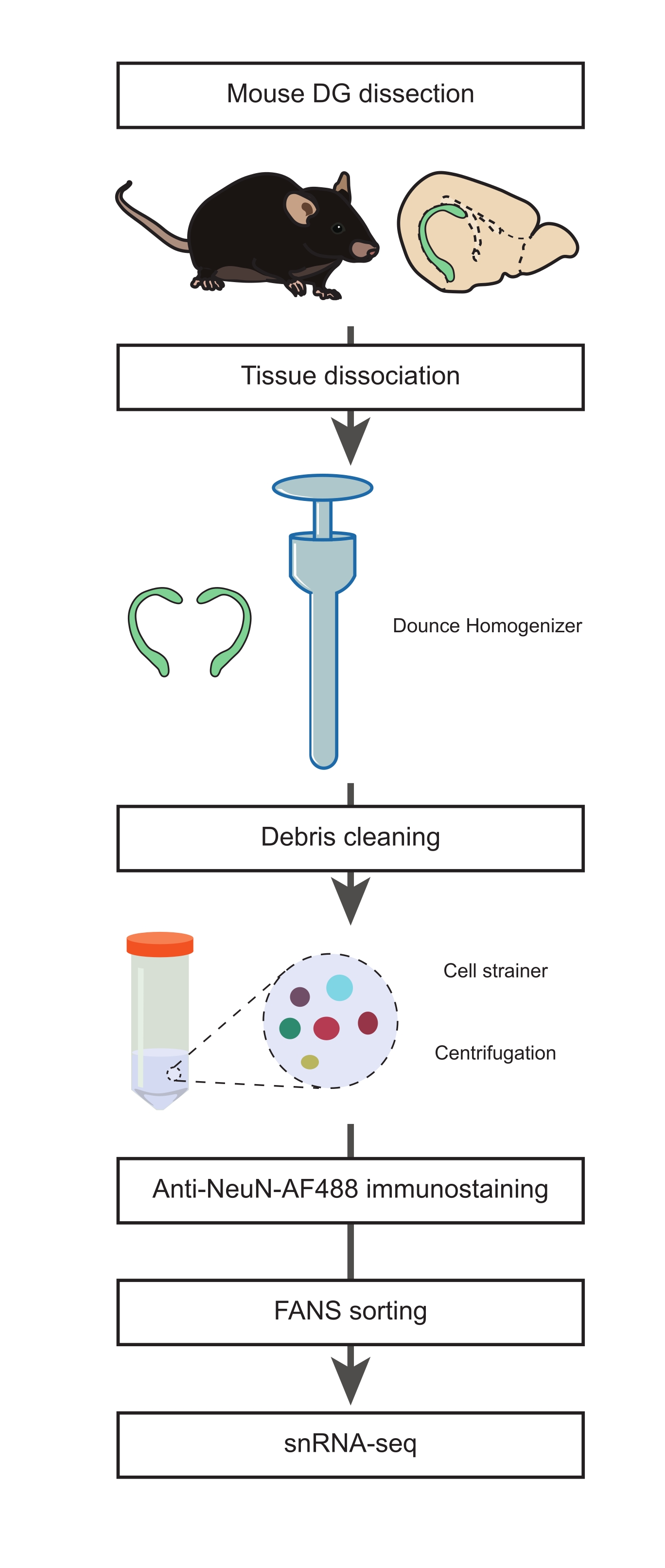
איור 1: הכנת תרחיף גרעינים יחיד מה-DG המנותח של עכברים בוגרים עבור snRNA-seq של אוכלוסיות שאינן נוירונים. דיאגרמת זרימה המתארת את השלבים העיקריים של הפרוטוקול הכוללים דיסקציה של DG עכבר, הכנת השעיה של גרעינים בודדים, חיסון NeuN ומיון שלילי של NeuN-FANS לפני שתמשיך עם snRNA-seq. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}
1. נתיחת DG (תזמון: 15 דקות)
- הכן מדיית בידוד גרעינים 1 ו-2 (NIM1 ו-NIM2), מאגר הומוגניזציה (HB) ומדיית שטיפה (WM) (טבלה משלימה 1). הניחו את כל המאגרים, חומרי ההדפסה, הריאגנטים והכלים על הקרח עד הצורך. שים את ההומוגנייזר של Dounce (ראו טבלת חומרים) על קרח במהלך ההכנה (לפחות שעה אחת לפני שלב ההומוגניזציה).
הערה: NIM1 ניתן להכנה ולאחסון בטמפרטורה של 4°C למשך עד 6 חודשים. NIM2, HB ו-WM צריכים להיות מוכנים טריים.
אזהרה: טפל ב-DTT, מעכב הפרוטאז וב-Triton X-100 בזהירות. תרכובות אלה מגרות את העור והעיניים, רעילות מאוד ומסוכנות לסביבה הימית. בזמן השימוש בכימיקלים אלה יש ללבוש כפפות מגן, בגדים, להגן על העיניים והפנים, לשטוף ידיים ביסודיות לאחר הטיפול, ולהימנע משחרור לסביבה. - הרדמת עכבר C57Bl/6J זכר בן 22 חודשים על ידי נקע צוואר הרחם בעקבות הליך לוח זמנים 1 של משרד הפנים20.
הערה: ראה דיון ברציונל לגבי השימוש בעכבר בן 22 חודשים במחקר זה. עם זאת, פרוטוקול זה יכול להתבצע בכל גיל לאורך תוחלת החיים. - נתחו את המוח מעכבר שעבר המתת חסד והעבירו אותו לצלחת פטרי בגודל 10 ס"מ מלאה ב-PBS קר כקרח (איור 1). מניחים את צלחת הפטרי על קרח. הסר את המוח הקטן באמצעות אזמל לחתוך את המוח לשניים בין שתי ההמיספרות (לאורך ציר sagittal).
- ממלאים צלחת פטרי חדשה בקוטר 10 ס"מ ב-PBS קר כקרח ומניחים אותה על קרח. העבירו חצי מהמוח לצלחת הפטרי החדשה. באמצעות משקפת, נתחו את ה-DG וחזרו על שלב זה כדי לקבל את ה-DG השני מהמחצית השנייה של המוח.
הערה: שלבים אלה (שלבים 1.2-1.4) הותאמו מתוך הליךשתואר קודם לכן 21. חשוב להתקדם מהר ככל האפשר בשלב זה כדי לשמור על שלמות התאים. - מעבירים את שני ה-DGs להומוגנייזר של Dounce שעבר קירור מראש ומוסיפים 1 מ"ל של HB קר.
2. דיסוציאציה של רקמות, בידוד גרעינים בודדים וחיזוק חיסוני אנטי-NeuN (תזמון: שעתיים)
- הומוגניזציה של הרקמה עם 10 משיכות של המזיק "A" רופף, ואחריו 15 משיכות של מזיק "B" הדוק.
הערה: יש לבצע הומוגניזציה של Dounce עם חומר המליטה על הקרח עם משיכות עדינות כדי להפחית את החום הנגרם על ידי חיכוך וקצף. כל המאגרים והציוד צריך להיות precooled ולשמור על קרח במהלך ההליך. - להעביר את homogenate לתוך צינור prechilled 15 מ"ל; לשטוף את Dounce homogenizer עם 1 מ"ל של HB קר ולשלב לאותו צינור. הוסף 3 מ"ל של HB לצינור 15 מ"ל ודגרה 5 דקות על קרח. מערבבים 2x על ידי היפוך הצינור בעדינות.
- יש להרטיב מראש מכסה מסננת בקוטר 70 מיקרומטר עם 0.5 מ"ל HB על פני מבחנה של 50 מ"ל. מסננים את תרחיף הגרעינים משלב 2.2 על ידי הטיה עדינה של צינור 15 מ"ל לתוך מסננת התא. לשטוף את מסננת התא עם 0.5 מ"ל של HB.
- הסר את מסננת התאים וצנטריפוגה את המבחנה ב 500 x g במשך 5 דקות ב 4 מעלות צלזיוס, באמצעות צנטריפוגה דלי נדנדה. השליכו את הסופר-נטנט.
הערה: מאמץ ההומוגנט יעזור להפחית את הפסולת, שהיא קריטית לציטומטריה של הזרימה ולמדרגות snRNA-seq במורד הזרם. - יש להשעות את הכדור בעדינות ב-4 מ"ל של HB באמצעות פיפטה P1000. דגירה על קרח במשך 5 דקות. סובב ב 500 x גרם במשך 10 דקות ב 4 מעלות צלזיוס. השליכו את הסופר-נטנט והחזירו את הכדור ב-3 מ"ל של זיכרון עבודה.
- יש להרטיב מראש מכסה מסננת בקוטר 35 מיקרומטר מעל מבחנה של 15 מ"ל עם 0.5 מ"ל של זיכרון עבודה (WM. מסננים את מתלה הגרעינים משלב 2.5 דרך מסננת התא, ומזרימים בעדינות 0.5 מ"ל בכל פעם באמצעות פיפטה P1000.
- לשטוף את מכסה המסננת עם 0.5 מ"ל של WM ומניחים את הצינור על קרח. מעבירים את התסנין לצינור חדש של 15 מ"ל ולצנטריפוגה למשך 5 דקות ו-4 מעלות צלזיוס ב-500 x גרם. השליכו את הסופר-נטנט והחזירו את הכדור ב-3 מ"ל של זיכרון עבודה.
- סובב ב 500 x גרם במשך 5 דקות ב 4 מעלות צלזיוס. יש להשליך את ה-supernatant ולהשהות את הכדור ב-1 מ"ל של זיכרון עבודה עם הנוגדן המצומד של העכבר anti-NeuN, Alexa Fluor 488-מצומד (anti-NeuN-AF488, 1:32,000) ו-1 מיקרוגרם/מ"ל DAPI. דגירה במשך 45 דקות על קרח בחושך.
הערה: כדי לייעל את החיסון של גרעינים מבודדים, מומלץ לבצע טיטרציה של הנוגדן כדי לקבוע את הדילול האופטימלי לניתוח ומיון ציטומטריה של זרימה. לאחר מכן, הפעילו בקרות מתאימות כדי לוודא שתנאי הצביעה הם אופטימליים. לדוגמה, עם הנוגדן המצומד anti-NeuN-AF488, הופעלה בקרה שלילית (כלומר, ללא תוספת של הנוגדן, איור משלים 1A) ובקרה חיובית (כלומר, צביעה עם הנוגדן, איור משלים 1B) כדי להעריך הפרדה של אוכלוסיות לא מוכתמות ומוכתמות. כאשר מתחילים לעבוד עם נוגדן מצומד AF488, מומלץ להפעיל בקרת איזוטיפ מצומד AF488 כדי להעריך את הספציפיות. אם נעשה שימוש בנוגדן שאינו מצומד, ייתכן שיהיה צורך בבקרה נוספת כגון הוספת נוגדן משני רק להכנת גרעינים כדי להעריך קשירה לא ספציפית של הנוגדן המשני.
3. מיון גרעינים מופעל פלואורסצנטי (FANS) כדי לא לכלול אוכלוסיות נוירונים (תזמון: 45 דקות)
- העבירו את תרחיף הגרעינים המוכתמים באימונו למבחנה של 5 מ"ל ושמרו אותו על הקרח עד לתחילת הליך הציטומטריה של הזרימה.
הערה: אם עובדים עם פיסות רקמה גדולות יותר משני DG של עכברים, ייתכן שיידרש דילול נוסף עם מאגר WM כדי למנוע סתימת FACS אם צפיפות הגרעינים בתמיסה הופכת גבוהה. - מערבבים את הדגימות במשך 3 שניות במהירות נמוכה לפני הכנסת הצינורות למכשיר FACS (ראו טבלת חומרים).
הערה: (הגדרת FACS) יש ליישר את מכונות המיון בתחילת ההליך עם חלקיקי כיול בהתאם להמלצות היצרן. עיכוב הטיפה כויל באמצעות חרוזים או מיקרוספרות (ראו טבלת חומרים) על פי מודל FACS. הדגימות מוינו בטמפרטורה של 4 מעלות צלזיוס במצב טוהר. כדי להקטין את נפח האיסוף, הגרעינים מוינו דרך פיית 70 מיקרומטר בלחץ המומלץ עבור ציטומטר הזרימה. גרעינים מוינו לצינורות קשירה נמוכים של 1.5 מ"ל (ראו טבלת חומרים) המכילים 50 מיקרון של זיכרון עבודה. כל צינורות האיסוף היו מצופים ב- PBS + 5% BSA ב 4 מעלות צלזיוס במהלך הלילה כדי להפחית את הסיכון של גרעינים להידבק לדפנות הצינור. - כדי לקבל את הנתונים מדגימה של תרחיף הגרעינים המוכתמים, הגדר את השערים בגובה DAPI ובאזור DAPI כדי לא לכלול את פסולת התאים ואת הגרעינים המצטברים (איור 2A). יתר על כן, הפרד גרעינים בודדים מכל אגרגטים או פסולת תאים שהוכתמו על-ידי DAPI על-ידי הגדרת השערים באזור פיזור הצד (SSC) של היומן ובאזור פיזור הלוג קדימה (FCS) (איור 2B).
- הגדר את השערים עבור האנטי-NeuN-AF488 ואזור ה-FSC, כדי לבודד את האוכלוסייה השלילית של NeuN-AF488, כפי שמוצג באיור 2C.
- לאחר הניתוח, תוך שימוש באסטרטגיית הגידור שתוארה לעיל, מיין את האוכלוסייה השלילית של NeuN-AF488 בצינור איסוף של 1.5 מ"ל מלא ב- 50 μL של זיכרון עבודה.
הערה: בעקבות אסטרטגיית החיתוך שתוארה לעיל והליך הנתיחה לבידוד ה-DG ממוחו של עכבר בוגר, האוכלוסייה השלילית של NeuN-AF488 צפויה לייצג ~14% מהגרעינים הבודדים.

איור 2: בידוד ופרופיל תעתיק של אוכלוסיות תאים שאינם נוירונים מה-DG. (A-C) אסטרטגיית Gating לבידוד גרעינים בודדים שליליים של NeuN-AF488 ולמניעת פסולת תאים. (A) FANS מנקדים חלקת מדגם מייצג של גרעינים מבודדים, המתארת את הגדרת השער לבחירת גרעיני DAPI+ והדרה של פסולת תאים ואגרגטים. (B) בחירה נוספת של גרעינים בודדים רלוונטיים באמצעות אזור FSC ואזור SSC. (C) השערים עבור NeuN-AF488 כדי לא לכלול את האוכלוסייה החיובית ולמיין עבור הגרעינים הבודדים השליליים. (D) מיקרוגרף של מתלה גרעינים יחיד טוב עם כמות מינימלית של פסולת ושיעור גבוה יותר של גרעינים באיכות טובה (צורה עגולה, חץ שחור) בהשוואה לגרעינים באיכות גרועה (חץ לבן). סרגלי קנה מידה = 50 μm, 10 μm (כניסה). (ה,ו) ניתוח נתוני snRNA-seq ופרופיל של אוכלוסיות התאים הנפרדות שבודדו מה-DG של עכברים זכרים בני 22 חודשים C57BL/6J. קירוב סעפת אחיד והקרנה עבור חלקות הפחתת ממדים (UMAP) של פרופילי גרעינים בודדים מהתאים (E) שאינם ממוינים על-ידי FACS ו-(F) תאים ממוינים FACS שליליים של NeuN, צבועים לפי סוג התא. (G) תרשימי עוגה המשווים את התדרים של סוגי תאים מזוהים בשתי הדגימות. (H) מדדים מתאימים לדגימות המרוצפות: מספר גרעינים, מספר חציוני של גנים ותעתיקים לכל גרעין. (I) עלילות כינור המציגות את התפלגות מספר הגנים והתעתיקים שזוהו עבור כל סוג תא בשתי הדגימות. אסטר. = אסטרוציטים, אוליג. = אוליגודנדרוציטים, Vasc. = תאים וסקולריים, CRCs = תאי Cajal-Retzius, Neur. = נוירונים, Imm. = תאים חיסוניים, OPCs = תאים מבשרי אוליגודנדרוציטים. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}
4. הכנת מתלה הגרעינים הבודדים לביצוע ריצוף RNA של גרעין יחיד (תזמון: 30 דקות)
- לאחר המיון, הוסף 1 מ"ל של PBS המכיל 1% BSA לצינור האיסוף על מנת לאסוף טיפות על דופן הצינור ולהסתובב ב 500 x g במשך 5 דקות ב 4 מעלות צלזיוס. יש להשליך את הסופר-נטנט ולהשאיר 50 מיקרו-ליטר.
הערה: טפל בגרעינים הצנטריפוגים בזהירות מכיוון שייתכן שיהיה קשה לצפות בכל כדור בתחתית הצינור. שימוש בצנטריפוגה עם דלי נדנדה יעזור להשליך את הסופר-נאטנט מבלי להפריע לכדור. - פיפט בעדינות כדי להשעות גרעינים צנטריפוגטיים. הוסף 5 μL של תרחיף גרעינים ל 5 μL של כחול טריפאן במיקרו-צינורית של 0.5 מ"ל.
התראה: טפל בכחול טריפאן בזהירות מכיוון שהוא מסוכן לבריאות, עלול לגרום לסרטן וחשוד בפגיעה בפוריות או בילד שטרם נולד. יש ללבוש כפפות מגן, בגדים והגנה על העיניים והפנים. אין לטפל עד שכל אמצעי הזהירות נקראו והובנו. - מדוד את הריכוז והערך את הכדאיות של תרחיף התא הבודד באמצעות המוציטומטר או מונה תאים אוטומטי (ראה טבלת חומרים). בצע הכנה וריצוף של הגרעינים כמפורט בשלב 5.
הערה: דגימות שנחשבו לאיכותיות לריצוף הראו צורת גרעין עגולה וקבועה מתחת למיקרוסקופ ללא פסולת תאים (איור 2D). נוכחות של הילה סביב קרום הגרעין או צבירה של גרעינים מרובים יחד הם סימנים לגרעינים פגומים, ואין להתייחס לתרחיפים כאלה של תאים עבור snRNA-seq (איור 2D). הריכוז הנמדד של גרעינים היה בטווח של 300-700 גרעינים/μL.
5. הכנת ספרייה וריצוף
הערה: תיאור השלבים הבאים מבוסס על פלטפורמת הריצוף הפנימית ששימשה במחקר זה (ראה טבלת חומרים). לכן, הגדרות מסוימות עשויות להיות שונות בעת שימוש בפלטפורמה אחרת. כאן, רק את השלבים העיקריים מתוארים וכל פרמטר צריך להיקבע על פי הנחיות ופרוטוקולים של היצרן הנבחר אם כי עם אופטימיזציה לפני השימוש הראשון. חשוב לוודא שהכנת הספריות מתבצעת מהר ככל האפשר לאחר ריכוז תרחיף גרעינים ממוינים כדי למנוע פגיעה ב-RNA ולהבטיח איכות אופטימלית של הרצף.
- טען בין 7,000 ל-10,000 גרעינים לתוך שבב חד-תאי מיקרופלואידי.
- גרעינים טעונים במחיצה בטיפות בקנה מידה ננוליטר באמצעות הבקר שסופק והריאגנטים מהספק הנבחר. גרעיני Lyse בתוך כל טיפה ומתמללים RNA הפוך.
הערה: בתוך טיפה כל ה- cDNA שנוצר שיתף את אותו ברקוד תא. - הכן ספריות עבור snRNA-seq בהתאם להנחיות מהספק הנבחר והבטח תאימות לפלטפורמת הריצוף. בדוק את האיכות והריכוז של ספריות סופיות באמצעות אלקטרופורזה, פלואורומטריה או שיטות מבוססות qPCR, ואם רלוונטי, אחסן אותן באופן שווה לפני הרצף.
- דנטורציה איגדה 3ʹ ספריות ביטוי גנים ודילול בהתאם להמלצת היצרן.
- בצע רצף אינדקס זוגי, יחיד או כפול בפלטפורמת ריצוף מהדור הבא עם עומק רצף של 50,000 זוגות קריאה לכל תא.
תוצאות
הפרוטוקול המוצג כאן מתאר שיטה להכנת השעיה של גרעינים בודדים שאינם נוירונים שבודדו מה-DG כדי לבצע snRNA-seq. עם או בלי מאווררים, אשכולות ביואינפורמטיים חשפו קבוצות מופרדות היטב של גרעינים המתאימים לסוגי תאים ידועים בתוך ה-DG (איור 2E,F). בתוך הדגימה שאינה ממוינת על ידי FACS, רוב הגרעינים האיכותיים שרצפו כללו שלוש קבוצות של תאי עצב (84.9% מסך הגרעינים עבור דגימה זו, איור 2E,G,H). תוצאות כאלה צפויות, בהתחשב בכך שאוכלוסיות התאים המיוצגות ביותר ב- DG הן נוירונים גרגיריים, נוירונים מעוררים אחרים (המסומנים נוירונים מעוררים), ונוירונים מעכבים10. האשכולות הלא-עצביים שזוהו היו מורכבים ברובם מסוגי תאי גליה (11.1%), כולל אסטרוציטים, אוליגודנדרוציטים ותאים מבשרי אוליגודנדרוציטים (OPCs), תאי מערכת החיסון (3.3%) ותאי קחאל-רציוס (0.6%). בעת ביצוע FANS כדי לא לכלול אוכלוסיות חיוביות של NeuN (מדגם ממוין FACS שלילי של NeuN; איור 2F,G,H), אשכולות של תאי גליה הפכו לדומיננטיים (81.3%). הבידוד של מספר גדול יותר של גרעיני גליה מאפשר פילוח טוב יותר של אוכלוסיות שונות שיתקבצו יחד ללא מאווררים. ואכן, בעת קיבוץ מחדש וניתוח גנים ספציפיים המתבטאים בתאי גזע עצביים או באסטרוציטים, ארבעה תתי-צבירים מופרדים (איור משלים 2A,B). בהסתכלות על סמנים תאיים ספציפיים יותר והערכת רמות ביטוי הגנים בכל סוגי התאים, זוהה אשכול קטן של תאי גזע עצביים הנפרדים בנפרד מהאוכלוסיות האסטרוציטיות העיקריות עם ביטוי גבוה יותר של Hopx ו-Notch2 וכמעט ללא ביטוי של Aldh1a1 או Aqp4 (איור משלים 2C). עם זאת, בגלל החפיפה בביטוי גנים בין אסטרוציטים לתאי גזע עצביים, יידרש ניתוח נוסף כדי ליצור פרופיל ספציפי ולזהות תתי-סוגים שונים של תאים. יתר על כן, בדגימת ה-FANS השלילית של NeuN היו אשכולות נוספים שסומנו כתאים וסקולריים (2.3%) הכוללים תאי אנדותל, פריציטים ותאי לפטומנינגיאליים של כלי הדם כאשר הם מוצלבים לביטוי של סמנים ספציפיים לתאים (הנתונים אינם מוצגים).
בעקבות ההנחיות לפרוטוקול שנבחר ליצירת ספריות לריצוף, התקבלו פרופילי ביטוי באיכות גבוהה עם או בלי מאווררים. עבור דגימות שרוצפו ב-50,000 קריאות/גרעינים, זוהו בממוצע 2,510 גנים לכל גרעין עבור הדגימה שאינה ממוינת על ידי FACS (5,578 תעתיקים, איור 2H) ו-1,665.5 גנים (3,508 תעתיקים) עבור דגימת FANS שלילית של NeuN, לאחר סינון גרעינים באיכות נמוכה (איור 2H,I). מדדים אלה מאשרים כי פרוטוקול זה מייצר פרופיל תעתיק באיכות גבוהה של גרעינים בודדים בדומה למחקרים המשתמשים בגישות שונות22,23 וכי תהליך מיון FACS אינו פוגע בגרעינים עבור snRNA-seq הבאים. יש לציין כי ההבדל במספר הגנים והתעתיקים לכל גרעין בין שתי הדגימות אינו נובע מאיכות נתונים נמוכה יותר, אלא מהשיעור הגבוה של תאי עצב בדגימה שאינה ממוינת על ידי FACS (84.9% לעומת 1.7% בדגימת FANS שלילית של NeuN), שיש להם פעילות שעתוק גבוהה יותר (2,660 גנים/גרעין ו-6,170 תעתיקים/גרעין בדגימה שאינה ממוינת FACS) מאשר פעילות השעתוק הממוצעת של כל סוגי התאים שאינם נוירונים (1,090 גנים/גרעין ו-1,785 תמלילים/גרעין, איור 2I).
יחד, תוצאות מייצגות אלה מראות כי הבחירה של גרעינים שליליים של NeuN באמצעות FANS היא כלי רב עוצמה לבידוד סוגי תאים בשפע נמוך מרקמת מוח שזה עתה נותחה ולבצע פרופיל תעתיק גרעיני יחיד באיכות גבוהה של אוכלוסיות תאים מובחנות אלה באמצעות שיטות snRNA-seq.
איור משלים 1: אימות של החיסון עבור FANS. תרחיף גרעינים דוגר (A) ללא נוגדן anti-NeuN-AF 488 כבקרה שלילית או (B) עם הנוגדן ורץ דרך סדרן FACS כדי לאמת את תנאי החיסון. אנא לחץ כאן כדי להוריד קובץ זה.
איור משלים 2: ניתוח ביטוי גנים והתקבצות מחדש של צביר האסטרוציטים. (A) תרשים קירוב והקרנה אחידים של סעפת עבור הפחתת ממדים (UMAP) המציג את ההתקבצות של 4968 גרעינים בהתבסס על פרופילי ביטוי כלל-גנומיים מאיור 2F. שיחות מסוג תא נעשו על סמך סמנים מסוג תא. (B) צביר אסטרוציטים המורכב מ-2579 גרעינים שנבחרו מתוך (A) לתת-הגדרה נוספת כדי לחקור תת-סוגים פוטנציאליים של תאים. ארבעה תתי-סוגים זוהו על ידי אשכולות Seurat (0-3), המוצגים על ידי צבעים שונים. (C) רמות ביטוי גנים של סמנים תאיים ספציפיים בארבעת סוגי התאים. כל המגרשים הושגו באמצעות חבילת Seurat R24. בקצרה, ספירות RNA-seq נורמלו עבור כל תא על ידי הביטוי הכולל והוכפלו בגורם קנה המידה (10,000). תוצאה זו עברה לאחר מכן שינוי ביומן. הערכים שעברו שינוי צורה שונה היו בקנה מידה (שונות שגודלה השתנה לאחד) וממורכזים (ממוצע מוגדר לאפס) בתוך כל תא לפני החלת UMAP לחישוב ההטבעות, ששימשו כערכים על צירי x ו- y. גרפים מייצגים את הפלט של טכניקת הפחתה ממדית בחלקת פיזור דו-ממדית שבה כל נקודה מייצגת תא עם קואורדינטות x ו- y בהתאמה בהתבסס על ההטבעות של התאים שנקבעו על ידי טכניקת ההפחתה. תאים עם חתימות גנים דומות ממוקמים קרוב זה לזה על ידי ההטבעות. אנא לחץ כאן כדי להוריד קובץ זה.
איור משלים 3: ניתוח ביטוי גנים של NeuN בשושלת הנוירוגנית. (A) תרשים UMAP המציג את ההתקבצות של שושלת נוירוגנית מתוך מערך נתונים זמין לציבור15. UMAPs נוצרו כמו באיור משלים 2. (B) רמות ביטוי גנים של סמנים תאיים ספציפיים ברחבי השושלת הנוירוגנית המראים אסטרוציטים (Aquaporin 4 = Aqp4), תאי גזע עצביים (חלבון הומיאודומין בלבד = הופקס), NeuN/Rbfox3 (תאי גזע עצביים ותאי אב ביניים [IPCs]), ותאי מחזור (קינאז תלוי ציקלין 6 =Cdk6). אנא לחץ כאן כדי להוריד קובץ זה.
טבלה משלימה 1: קומפוזיציות של מדיה ומאגרים ששימשו במחקר. אנא לחץ כאן כדי להוריד קובץ זה.
Discussion
כדי לבצע בהצלחה פרוטוקול זה, כריתת ה- DG היא הצעד הקריטי הראשון, הדורש תרגול מסוים כדי לשמור עליו לא פגום ולהגביל את הזיהום מהרקמות הסובבות. מניסיון, הפרדת ה-DG מההיפוקמפוס יכולה להירכש מהר מאוד על ידי חוקר מיומן שיכול לעבוד על זיקוק הטכניקה שלהם כדי להגביר את מהירות הדיסקציה ולכן לשפר את טריות הרקמה כדי לייצר נתונים באיכות גבוהה. ברוח דומה, הכנה וחידוש של גרעינים בודדים דורשת עקביות בין התנאים השונים המשמשים בניסוי יחיד, אך גם הימנעות מפיפטינג מוגזם שעלול לשבש את שחרור הרנ"א הסביבתי שיטה את תוצאות הריצוף. בנוסף להמלצות שהוזכרו קודם לכן להכנת גרעינים באיכות גבוהה, יש לשקול גם את ריכוז ההשעיה של הגרעין הבודד לפני שממשיכים בריצוף. ואכן, על פי הנחיות היצרן, יש לדלל תכשיר עם ריכוז גבוה מ-1,200 nuc/μL, שכן רמה זו של ריכוז גרעינים תהיה בעלת סיכון גבוה יותר ליצירת הכפלות המשפיעות על ניתוחים ביואינפורמטיים במורד הזרם. יש לציין כי ריצוף דגימות עם ריכוזי גרעינים מתחת ל-500 nuc/μL עשוי שלא להיות כדאי בשל העלות הכרוכה בכך. מומלץ גם לעקוב אחר עצתו של משתמש FACS מתקדם כדי להגדיר את כל ה- gating ולהישאר עקביים עם ההגדרות על פני דגימות ושכפולים ביולוגיים. כמו כן, הכנת ספריות לריצוף RNA כרוכה בהכשרה מסוימת כדי להשיג תוצאות באיכות גבוהה ולרוב הספקים יש תמיכה מצוינת להשיג זאת ביעילות. שיטה זו נבדקה רק עם רקמה טרייה במחקר זה; עם זאת, FANS בוצע גם עם רקמה קפואה25. לכן סביר להניח כי פרוטוקול זה יכול להתבצע עם רקמה קפואה אם כי עם אופטימיזציה קלה.
פרוטוקול זה פותח מתוך מחשבה על יישום מסוים במורד הזרם, שהוא לחקור אוכלוסיות תאים שאינן נוירונים בתוך הנישה הנוירוגנית בהיפוקמפוס. ואכן, קווי ראיות הולכים וגדלים מצביעים על כך שניתן לייחס פגיעה ב-AHN בהזדקנות לתאים הסובבים בתוך הנישה 1,2,3,9. בפרט, אסטרוציטים ואוליגודנדרוציטים מתגלים כרגולטורים מרכזיים של AHN; עם זאת, הבידוד שלהם מה-DG בשילוב עם ריצוף RNA הניב תוצאות מעורבות, מה שהופך את ההשערה הזו למאתגרת להערכה בטכניקה זו 1,17. גישה זו של מיון FACS גרעינים שליליים של NeuN אפשרה בידוד של יותר אסטרוציטים ואוליגודנדרוציטים בהשוואה לדגימות שלא מוינו על ידי FACS, מה שמאפשר ניתוח ביואינפורמטי טוב יותר. פרוטוקול זה ישים בכל הגילאים לאורך תוחלת החיים והנתונים המייצגים המוצגים כאן עם רקמות מבעלי חיים ישנים מספקים הוכחת היתכנות לכך ששיטה זו חזקה לחקר הנישה הנוירוגנית המזדקנת בהיפוקמפוס. כדי להרחיב את השימוש בשיטה זו ולהתאים אותה לשאלות ביולוגיות שונות, חשוב לשקול כי אנטיגנים אחרים של ממברנה גרעינית עצבית יכולים להיבדק יחד עם טיטרציה יסודית של הנוגדנים המאומתים הטובים ביותר עבור סמנים אלה. לדוגמה, כאשר חוקרים את תהליך ההתמיינות העצבית מתאי גזע עצביים ב-DG, סוגי תאים מסוימים, כגון תאים מסוג 2 או נוירובלסטים, מתחילים לבטא את NeuN (איור משלים 3). לכן, יהיה צורך באנטיגן אחר כדי לחקור באופן ספציפי את סוגי התאים האלה. לעומת זאת, חלק מתאי העצב עדיין זוהו במחקר זה לאחר מיון FACS שלילי ל-NeuN, אולי בגלל ביטוי נמוך או ללא ביטוי של NeuN באוכלוסיות אלה (למשל, נוירונים קחאל-רציוס קליפת המוח19). בנוסף, דווח כי NeuN באה לידי ביטוי בתת-אוכלוסיות של אוליגודנדרוציטים26, מה שיכול לתת תוצאות מוטות אם תת-אוכלוסיות אלה היו מעניינות. לכן, הבחירה של אנטיגן כאשר מתחילים להשתמש FANS צריך להיחשב בזהירות כדי למנוע הכללה או הדרה של אוכלוסיות תאים שימנעו תשובה מדויקת לשאלה ביולוגית ספציפית. בהסכמה עם זה, מומלץ גם שכל תוצאת ריצוף תאושר עוד יותר על ידי בדיקות אורתוגונליות (למשל, אימונוהיסטוכימיה או היקף RNA) לפני אימות או הפרכה של ההשערה הנבדקת בפרוטוקול זה. לבסוף, ניתן לפתח עוד יותר את השלב הכרוך ב-FANS כך שיכלול יותר מנוגדן אחד עם אסטרטגיית מיון משוכללת יותר כדי לא לכלול ו/או לכלול אוכלוסיות תאים רצויות.
בסופו של דבר, לטכנולוגיות המתוארות בפרוטוקול זה עשויות להיות מגבלות מסוימות כאשר משתמשים בהן עם מינים אחרים. לדוגמה, הנישה מוגדרת היטב במכרסמים עם נוכחות של תאי גזע עצביים מתרבים ושקטים או נוירונים שזה עתה נולדו המוגבלים בתוך תת-אזורים ספציפיים של DG, אך עדיין לא ברור כיצד יש להגדיר את הנישה הנוירוגנית בהיפוקמפוס במינים אחרים. ואכן, תאים מתרבים אינם מיושרים בתוך אזור רציף של ה-DG בפרימטים ובבני אדם שאינם בני אדם, אלא מפוזרים סביבו ועשויים להימצא גם באמיגדלה7. לכן, ניתוח ובידוד אזורים רחבים יותר מאשר DG במינים אחרים עשוי להשפיע על השימוש בפרוטוקול זה. במיוחד, שלבי הדיסוציאציה והטריטורציה להכנת רקמה יצטרכו להיות אופטימליים תוך כדי עבודה עם חתיכות גדולות יותר של רקמה27,28. באשר לאנליזה ביואינפורמטית, בעוד שלמכרסמים גזעיים יש גנום הומוגני מאוד ומבואר היטב, השונות הגנטית של הגנום האנושי בשילוב עם מספר לא מספיק של סמנים תאיים כדי להבחין בבירור בין אוכלוסיות תאים שונות (למשל, תאי גזע עצביים ואסטרוציטים) דורשת הרבה נורמליזציה לניתוח שיכול להוביל למסקנות שונות כאשר צביר קטן של תאים מזוהה7, 11. במצבים כאלה, העשרת תאים עשויה עדיין להיות אפשרות מועדפת או שיש להשתמש בה לצד אסטרטגיות אחרות להגברת הכוח האנליטי.
עם זאת, הגישה הנוכחית יכולה לאפשר לחקור את תפקידן של אוכלוסיות תאים מוחלשות, אם כי בעלות פוטנציאל חשוב, בוויסות של AHN. זה יכול להיות המקרה במיוחד עבור אוכלוסיות של אסטרוציטים, אשר ממלאים תפקיד מרכזי בהתפרצות והתקדמות של מחלות נוירודגנרטיביות29,30. מחקר זה הראה כי ניתן לזהות אסטרוציטים ואוכלוסיות תאים נדירים אחרים ולתאר אותם פשוט על ידי אי הכללת הרוב המכריע של תאי העצב הנמצאים בתוך ה-DG. מחקרים אחרים שהשתמשו בגישות שונות לא הצליחו להשיג התאוששות דומה של גרעינים מאותו טווח של אוכלוסיות תאים 5,11,17. יתר על כן, תוצאות מחקר זה מראות כי ניתן להשתמש בגישה זו כדי לבודד אשכול NSC ללא העשרה ספציפית של אוכלוסיית תאים זו15.
לסיכום, מעקב ושיפור שיטה זו יהיה צעד קדימה כדי להתמודד עם שאלות יוצאות דופן הקשורות לתפקיד ההקשר של נישה נוירוגנית בהיפוקמפוס עבור אפנון של AHN. בפרט, הוא יכול להביא תובנות חדשות על רמות ביטוי גנים במוחות זקנים וחולים באוכלוסיות תאים הקשורות לוויסות של AHN9, לתמוך בזיהוי הטרוגניות פוטנציאלית של תאי גזע עצביים1 או לטפל בתפקיד כלי הדם ב-AHN. בסופו של דבר, שיטה זו יכולה להיות מותאמת לנישות אחרות של תאי גזע בוגרים עם שאלות וסוגיות דומות.
Disclosures
SG, TL ו- SK הם עובדים של Merck Sharp & Dohme LLC, חברת בת של Merck & Co., Inc., Rahway, ניו ג'רזי, ארה"ב הידועה בשם MSD מחוץ לארה"ב וקנדה. SG היא בעלת מניות של Merck & Co., Inc., Rahway, ניו ג'רזי, ארה"ב.
Acknowledgements
המחברים רוצים להודות ללכלן האריס ופיירו ריגו על התמיכה הטכנית ולג'ייסון מ. אוסלנר ודיטה לובאט על מתן משוב על כתב היד. עבודה זו נתמכה על ידי תמיכת מענקים מ- MRC ושיתוף פעולה מחקרי טרום תחרותי עם MSD, מכון פרנסיס קריק, המקבל את המימון שלו מ- Cancer Research UK (FC0010089), המועצה למחקר רפואי בבריטניה (FC0010089), קרן Wellcome (FC0010089) ועל ידי פרס חוקר אמון Wellcome ל- FG (106187 / Z / 14 / Z). אנו מתנצלים בפני הסופרים הרבים שאת עבודתם לא יכולנו לדון ולצטט בגלל חוסר מקום.
Materials
| Name | Company | Catalog Number | Comments |
| 0.5ml microtube | Eppendorf | 30124537 | |
| 10.00µm Flouresbrite YG Carboxylate Microspheres | Polysciences | 15700-10 | |
| 15 mL polypropylene centrifuge tubes | Corning | 430052 | |
| 2 pairs of sterile Dumont #5 forceps | Fine Science Tools | 11252-30 | |
| 4′,6-diamidino-2-phenylindole (DAPI) | Sigma Aldrich | D9564-10MG | |
| 4150 TapeStation System | Agilent | N/A | |
| 5 mL round bottom high clarity polypropylene test tube with snap cap | Falcon | 352063 | |
| 5 mL round bottom polystyrene test tube with cell strainer snap cap | Falcon | 352235 | |
| 50 mL polypropylene centrifuge tubes | Corning | 430829 | |
| 70 µm cell strainer | Falcon | 352350 | |
| 8 peak SPHERO Rainbow Calibration Particles | BD Biosciences | RCP-30-5A | |
| Accudrop Beads | BD Biosciences | N/A | |
| Allegra X-30R Centrifuge | Beckman Coulter | N/A | |
| Anti-NeuN antibody, clone A60, Alexa Fluor 488 conjugated | Millipore | MAB377X | |
| BD FACSAria Fusion Flow Cytometer | BD Biosciences | N/A | |
| Beckman Coulter MoFlo XDP | Beckman Coulter | N/A | |
| Chromium Controller | 10x Genomics | N/A | |
| Chromium Next GEM Single Cell 3' Reagent Kits v3.1 | 10x Genomics | PN-1000121; PN-1000120; PN-1000213 | |
| BSA 7.5% | Gibco | 15260037 | |
| Dithiothreitol (DTT) | Thermo Scientific | R0861 | |
| Dounce tissue grinder set: mortar, loose pestle (A) and tight pestle (B) | KIMBLE | D8938-1SET | |
| Eppendorf Tubes Protein LoBind 1.5ml | Eppendorf | 30108116 | |
| Halt, 100x Protease inhibitor | ThermoFisher | 78429 | |
| HiSeq 4000 Sequencing System | Illumina | N/A | Sequencing configuration: 28-8-0-91 |
| KCl | Any chemical supplier | Laboratory made | |
| LUNA-FX7 Automated Cell counter | Logos Biosystems | N/A | |
| MgCl2 | Any chemical supplier | Laboratory made | |
| N°10 guarded sterile disposable scalpels | Swann-Morton | 6601 | |
| Nuclease-free water | Sigma Aldrich | W4502-1L | |
| Pair of sterile student surgical scissors | Fine Science Tools | 91401-12 | |
| PBS | Any chemical supplier | Laboratory made | |
| RNase Inhibitor 40 U µl-1 | Ambion | AM2684 | |
| RNasin 40 U µl-1 | Promega | N211A | |
| Sterile Petri dish | Corning | 430167 | |
| Sucrose | Sigma Aldrich | 59378-500G | |
| Tris buffer, pH 8.0 | Any chemical supplier | Laboratory made | |
| Triton X-100 10% (v/v) | Sigma Aldrich | T8787-250ML | |
| Trypan blue | Invitrogen | T10282 |
References
- Gillotin, S. Targeting impaired adult hippocampal neurogenesis in ageing leveraging intrinsic mechanisms regulating neural stem cell activity. Ageing Research Reviews. 71, 101447 (2021).
- Urban, N., Blomfield, I. M., Guillemot, F. Quiescence of adult mammalian neural stem cells: A highly regulated rest. Neuron. 104 (5), 834-848 (2019).
- Hanspal, M. A., Gillotin, S. A new age in understanding adult hippocampal neurogenesis in Alzheimer's disease. Neural Regeneration Research. 17 (12), 2615-2618 (2022).
- Zhang, H., et al. Single-nucleus transcriptomic landscape of primate hippocampal aging. Protein & Cell. 12 (9), 695-716 (2021).
- Franjic, D., et al. Transcriptomic taxonomy and neurogenic trajectories of adult human, macaque, and pig hippocampal and entorhinal cells. Neuron. 110 (3), 452-469 (2022).
- Moreno-Jimenez, E. P., Terreros-Roncal, J., Flor-Garcia, M., Rabano, A., Llorens-Martin, M. Evidences for adult hippocampal neurogenesis in humans. Journal of Neuroscience. 41 (12), 2541-2553 (2021).
- Sorrells, S. F., et al. Positive controls in adults and children support that very few, if any, new neurons are born in the adult human hippocampus. Journal of Neuroscience. 41 (12), 2554-2565 (2021).
- Zhou, Y., et al. Molecular landscapes of human hippocampal immature neurons across lifespan. Nature. 607, 527-533 (2022).
- Bonafina, A., Paratcha, G., Ledda, F. Deciphering new players in the neurogenic adult hippocampal niche. Frontiers in Cell and Developmental Biology. 8, 548 (2020).
- Amaral, D. G., Scharfman, H. E., Lavenex, P. The dentate gyrus: fundamental neuroanatomical organization (dentate gyrus for dummies). Progress in Brain Research. 163, 3-22 (2007).
- Habib, N., et al. Massively parallel single-nucleus RNA-seq with DroNc-seq. Nature Methods. 14 (10), 955-958 (2017).
- Flor-Garcia, M., et al. Unraveling human adult hippocampal neurogenesis. Nature Protocols. 15 (2), 668-693 (2020).
- Moreno-Jimenez, E. P., et al. Adult hippocampal neurogenesis is abundant in neurologically healthy subjects and drops sharply in patients with Alzheimer's disease. Nature Medicine. 25 (4), 554-560 (2019).
- Kalinina, A., Lagace, D. Single-cell and single-nucleus RNAseq analysis of adult neurogenesis. Cells. 11 (10), 1633 (2022).
- Harris, L., et al. Coordinated changes in cellular behavior ensure the lifelong maintenance of the hippocampal stem cell population. Cell Stem Cell. 28 (5), 863-876 (2021).
- Shin, J., et al. Single-cell RNA-seq with Waterfall reveals molecular cascades underlying adult neurogenesis. Cell Stem Cell. 17 (3), 360-372 (2015).
- Artegiani, B., et al. A single-cell RNA sequencing study reveals cellular and molecular dynamics of the hippocampal neurogenic niche. Cell Reports. 21 (11), 3271-3284 (2017).
- Nott, A., Schlachetzki, J. C. M., Fixsen, B. R., Glass, C. K. Nuclei isolation of multiple brain cell types for omics interrogation. Nature Protocols. 16 (3), 1629-1646 (2021).
- Sarnat, H. B., Nochlin, D., Born, D. E. Neuronal nuclear antigen (NeuN): a marker of neuronal maturation in early human fetal nervous system. Brain Development. 20 (2), 88-94 (1998).
- . Guidance on the Operation of ASPA Available from: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_dat/file/662364/Guidance_on_the_Operation_of_ASPA.pdf (2022)
- Hagihara, H., Toyama, K., Yamasaki, N., Miyakawa, T. Dissection of hippocampal dentate gyrus from adult mouse. Journal of Visualized Experiments. (33), e1543 (2009).
- Habib, N., et al. Disease-associated astrocytes in Alzheimer's disease and aging. Nature Neuroscience. 23 (6), 701-706 (2020).
- Ding, J., et al. Systematic comparison of single-cell and single-nucleus RNA-sequencing methods. Nature Biotechnology. 38 (6), 737-746 (2020).
- Hao, Y., et al. Integrated analysis of multimodal single-cell data. Cell. 184 (13), 3573-3587 (2021).
- Mussa, Z., Tome-Garcia, J., Jiang, Y., Akbarian, S., Tsankova, N. M. Isolation of adult human astrocyte populations from fresh-frozen cortex using fluorescence-activated nuclei sorting. Journal of Visualized Experiments. (170), e62405 (2021).
- Zhang, Y., et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. Journal of Neuroscience. 34 (36), 11929-11947 (2014).
- Marti-Mengual, U., Varea, E., Crespo, C., Blasco-Ibanez, J. M., Nacher, J. Cells expressing markers of immature neurons in the amygdala of adult humans. European Journal of Neuroscience. 37 (1), 10-22 (2013).
- Zhang, X. M., et al. Doublecortin-expressing cells persist in the associative cerebral cortex and amygdala in aged nonhuman primates. Frontiers in Neuroanatomy. 3, 17 (2009).
- Ding, Z. B., et al. Astrocytes: a double-edged sword in neurodegenerative diseases. Neural Regeneration Research. 16 (9), 1702-1710 (2021).
- Phatnani, H., Maniatis, T. Astrocytes in neurodegenerative disease. Cold Spring Harbor Perspectives in Biology. 7 (6), 020628 (2015).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved