
Method Article
研究蛋白质精氨酸甲基转移酶1-9细胞活性的方法
摘要
这些协议提供了用于评估细胞中蛋白质精氨酸甲基转移酶(PRMT)家族中个体成员的酶活性的方法。详细介绍了使用内源性和外源性生物标志物、甲基-精氨酸识别抗体和抑制剂工具化合物评估PRMT活性的指南。
摘要
蛋白质甲基转移酶 (PRMTs) 催化甲基组向基质蛋白精氨酸残留物的转移。PRMT 家族由九名成员组成,可单体或对称/不对称地使用二甲基酸精氨酸残留物。有几个抗体识别不同类型的各种蛋白质的精氨酸甲基化:从而为PRMT活性生物标志物检测的发展提供了工具。PRMT 基于抗体的检测由于基材重叠和基于图案的抗体特异性而具有挑战性。讨论了这些问题以及研究单个PRMT所促成的精氨酸甲基化的实验设置。通过仔细选择具有代表性的基材,为九分之八的PRMT提供生物标志物,设计了一个PRMT活动检测小组。在这里,报告了以定量测量PRMT家族中单个成员在细胞中的酶活性而进行细胞检测的协议。上述方法的优点是它们在任何具有细胞培养和荧光西斑功能的实验室中都具有直接的性能。基板特异性和所选抗体可靠性通过击倒和过度表达方法得到充分验证。除了检测生物标志物和抗体的详细指南外,还提供了关于使用抑制剂工具为PRMT收集化合物的信息。
引言
精氨酸甲基化是调节蛋白质-蛋白质和蛋白质-RNA相互作用的重要转化后修饰,因此在MRNA前拼接、DNA损伤、转录反应和生长因子介导1、2等各种细胞过程中发挥着重要作用。精氨酸是由蛋白质精氨酸甲基转移酶 (PRMTs) 导致单甲基精氨酸 (Rme1), 不对称二甲基金宁 (Rme2a), 或对称二甲基金素 (Rme2s)3.根据甲基化类型,PRMT分为三组:I型(PRMT1、2、3、4、6和8),它们催化单二元化和不对称二元化:II型(PRMT5和PRMT9),促进单二元化和对称二元化:和类型III(PRMT7),它只能单甲基酸酯精氨酸3。
由于越来越多的市售精氨酸甲基化特异性抗体,PRMT活性可以用西方印迹来测量。荧光为基础的西方污点是首选的技术比化疗检测,由于更大的动态范围和线性,更高的灵敏度,并允许多路复用4。要量化蛋白质甲基化水平,需要将甲基化信号正常化到总蛋白质水平。通过选择在不同宿主物种(如小鼠和兔子)中培养的总和甲基化蛋白的抗体,可以使用标有不同氟磷的二次抗体,并在同一样本带中确定两种抗体的信号。开发甲基-精氨酸抗体是为了识别和描述甲基化、不对称或对称的二甲基化蛋白质,其中甲基-精氨酸是在特定环境中发现的。由于大多数PRMT的基底5内含有甲基酸甘油和精氨酸丰富的图案,因此分别为含有单甲基或不对称、对称二甲基-精氨酸-甘氨酸重复的肽(如D5A12、ASYM24或ASYM25和SYM11)提出了若干抗体。其他甲基-精氨酸抗体产生于肽库中,该库含有不对称、对称的二甲基和单甲基精氨酸,在重复的语境中促进在这些特定环境中检测甲基-精氨酸6。也有越来越多的抗体,识别单一蛋白质上的特定精氨酸标记,使甲基化选择性检测,如高石H4R3me2a或BAF155-R1064me2a。
有几个市售的PRMT抑制剂,可以用作PRMT细胞测定工具。然而,并非所有这些都具有完全的选择性和偏离目标的效果,有些应谨慎使用。结构基因组联盟与学术实验室和制药合作伙伴合作,开发出具有良好特征的强效、选择性和细胞渗透性PRMT抑制剂(化学探针),科学界可以毫无限制地使用这种抑制剂。这些抑制剂的信息可以在 https://www.thesgc.org/chemical-probes/epigenetics 和 https://www.chemicalprobes.org/ 上找到。化学探针是小分子抑制剂,体外IC50或K d<100纳米,选择性超过同一家族蛋白质的30倍,细胞活性显著,为1μM。 此外,每个化学探针都有一个紧密的化学模拟,对预定目标7、8、9、10、11、12不活跃。
该协议的目标是使用荧光西斑法测量单个PRMT家庭成员的细胞活性。这里提供了有关验证的检测生物标志物、抗体和强效细胞活性抑制剂的详细信息,以及成功实施检测的宝贵策略。
研究方案
1. 细胞培养和电镀
注:培养细胞与推荐的介质和测试定期支原体污染。HEK293T、MCF7 和 C2C12 细胞被选为示例,因为这些细胞系在 PRMT 检测中成功使用。
- 文化 HEK293T, MCF7 和 C2C12 在 DMEM 补充 10% 胎儿牛血清 (FBS), 青霉素 (100 U mL−1),和链霉素 (100μg mL−1)在 10 厘米组织培养处理 (TC) 菜肴.
- 对于PRMT8检测,在检测开始前3天内,在含有多氧环素(2μg/mL)的介质中生长PRMT1不可推卸的HK293T细胞。
- 要将细胞镀上,从板中取出并丢弃介质。
- 添加 10 mL 的 PBS(无 Ca+2 和 Mg+2 离子)以清洗细胞并丢弃溶液。
- 加入 1 mL 的 Trypsin-EDTA (0.25%),在室温 (RT) 下孵育 1 分钟,然后丢弃溶液。孵化,直到细胞变得圆,并从盘子分离。如果需要,点击板帮助分离细胞。对于难以尝试的细胞,如 C2C12,在 37 °C 下孵育板 1-2 分钟。
注意:避免细胞长时间接触尝试普辛溶液(>10分钟),因为它会降低细胞的生存能力。 - 在板中加入 1 mL 的预热介质,然后轻轻地上下移动器单元,以分解细胞团。将细胞转移到 15 mL 管中,并添加 3-5 mL 的介质。
- 要测量细胞数量,将 10 μL 的细胞与 10 μL 的 Trypan 蓝色混合,并将 10 μL 传输到血细胞仪或使用任何其他细胞计数方法。
- 稀释细胞以推荐细胞密度,并将 500 μL/井放入 24 井 TC 板(表 1)中。对于内源性测定(PRMT1、PRMT4、PRMT5、PRMT7 和 PRMT9),移动到步骤 3.1。
2. 细胞传输
- 对于外源性测定(PRMT3、PRMT6、PRMT8)使用推荐的DNA量(表2)对 HEK293T 细胞进行脱血。HEK293T 细胞易于转染,因此任何转染试剂都可以按照制造商的指示使用。
3. 复合处理
注:文化媒体中最终二甲基硫化物(DMSO)浓度不超过0.1%。在每口井中保持相同的 DMSO 浓度。选择性PRMT抑制剂(化学探针)及其密切相关的非活性模拟可以在 表3中找到。
- 对于 内源性检测 (PRMT1、PRMT4、PRMT5、PRMT7 和 PRMT9),从细胞中去除介质,并单独用化合物或 DMSO(控制)替换 500μL 介质。
注意:观察R甲基化水平下降超过80%通常需要2天。 - 对于 外源性检测 (PRMT3、PRMT6、PRMT8),在传输后4小时移除介质,单独添加500μL复合物或DMSO(控制)介质,孵育20-24小时。
4. 细胞利萨特制备
- 从井中取出所有介质,用 100 μL 的 PBS 清洗以去除残留介质,并添加 60 μL 的溶解缓冲(20 mM Tris-HCl pH 8, 150m NaCl, 1 mM 乙酰胺乙酰酸 (EDTA), 10 mM MgCl2, 0.5% 特里顿 X-100, 12.5 U mL−1 苯酶, 完整的 EDTA 无蛋白酶抑制剂鸡尾酒) 到每口井.
- 在 RT 孵化不到 1 分钟,摇动板在细胞上分配裂解缓冲。然后加入3 μL的20%w/v硫酸钠(SDS),到最终1%的浓度,并通过轻轻摇晃混合。将利萨酸盐转移到微中燃料管中,并将其保持在冰上。
注意:使用前加入苯甲酶和蛋白质抑制剂鸡尾酒。加入苯酶快速水解核酸,降低细胞的乳酸粘度。
- 在 RT 孵化不到 1 分钟,摇动板在细胞上分配裂解缓冲。然后加入3 μL的20%w/v硫酸钠(SDS),到最终1%的浓度,并通过轻轻摇晃混合。将利萨酸盐转移到微中燃料管中,并将其保持在冰上。
- 使用 BCA 蛋白质测定套件确定样品的蛋白质浓度,或使用任何其他在溶液中耐受 1% SDS 的方法。
- 在裂解缓冲区中加入 2μL 的裂解水和蛋白质标准(0、1、2、4 和 8 μg/mL BSA)添加到 96 井透明板的井中。
- 将试剂 A 与试剂 B 以 50:1 的比例混合,每口井添加 200 μL。在37°C孵化20分钟,并阅读吸收。
- 用裂解缓冲器调整蛋白质浓度,使样品中的蛋白质浓度相等。
- 将 20 μL 的 4 倍加载缓冲区添加到 60μL 的电池裂解油中,并在 95 °C 下加热 5 分钟。热变性后,裂解液可储存在-20°C。
5. 西方污点分析
- 将总细胞裂解水的 5-20 μg 装载到 4-12% 的 Bis-Tris 蛋白质凝胶中,用于分析高石蛋白,将其他蛋白质的 20-100 μg 放入 4-12% 的 Bis-Tris 蛋白凝胶中。
- 在 MOPS SDS 中运行缓冲区(50 mM MOPS、50 mM 三元基础、0.1% SDS w/v、1 mM EDTA、pH 7.7)中运行凝胶,在 100 V 处运行约 2 小时,或直到染料前部到达凝胶底部。
- 如果执行湿转移,在冰冷的三合院-甘油转移缓冲区(25 mM 三重奏、192 mM 胶氨酸、20% v/v 甲醇和 0.05% w/v SDS)中组装转移三明治。
- 根据制造商的说明放置海绵、滤纸、PVDF 膜和凝胶。通过在组装前将甲醇和等效凝胶浸泡在转移缓冲液中 30s 来激活 PVDF 膜。
注意:使用推荐的PVDF西印迹膜,因为它具有低自流和适合低分子量蛋白质,如石酮(材料表)。
- 根据制造商的说明放置海绵、滤纸、PVDF 膜和凝胶。通过在组装前将甲醇和等效凝胶浸泡在转移缓冲液中 30s 来激活 PVDF 膜。
- 在3S-Glycine中将蛋白质从凝胶转移到PVDF膜,在70 V时在冰上以1.5小时的速度转移缓冲区。
- 在阻塞缓冲区块膜 30 分钟(磷酸盐缓冲盐水中 5% w/v 牛奶,PBS)。用洗涤缓冲器冲洗(PBST:PBS中0.1%v/v Tween-20),并在牛血清白蛋白(BSA)溶液(PBST中5%BSA)中用主要抗体在4°C(表3)过夜孵化。
注意:对于更长的存储空间,过滤消毒 BSA 溶液,加入 0.02% w/v 钠 azide,并保持在 4 °C。 - 使用PBST清洗膜3 x 5分钟。然后在推荐的阻塞缓冲器(材料表,表3)中用山羊防兔(IR800)和驴抗鼠标(IR680)抗体在RT孵育30分钟,用PBST清洗3×5分钟。
- 在 800 和 700 nm 的荧光西污点成像仪上阅读信号。最好使用该仪器,使图像强和微弱波段清晰在一个图像具有高灵敏度和动态范围,高信号噪声比,当图像饱和达到警告,以及在同一样本带的两种荧光颜色的复用。
- 使用适当的软件进行荧光西成像,确定西斑分析的带状强度。
结果
下面介绍了单个PRMT细胞测定西面污点结果示例。分析细节也总结在表4。
PRMT1 检测
PRMT1是13号细胞中希斯通4精氨酸3不对称二甲基化(H4R3me2a)的主要贡献者。在PRMT1活动中断后,全球Rme1和Rme2s水平显著增加13。如图1A和1B所示,几个抗体可用于监测Rme1、Rme2a、Rme2s以及H4R3me2a的全球变化。全球 Rme2a 和 H4R3me2a 水平显著下降,Rme1 和 Rme2s 在PRMT1击倒 3 天后(图 1A,B)可观察到其增加。细胞系在基底H4R3me2a信号中不同,因此,为了便于监测PRMT1活性的损失,可以使用基底甲基化水平高的MCF7等细胞系(图1C)。观察PRMT1抑制作用的最佳时间是2天(图1D,1E)。更长的治疗导致细胞存活率和生长率降低。
PRMT3检测
对于PRMT3细胞测定,在PRMT3击倒或过度表达后,在西方污点中无法检测到甲基化变化的选择性生物标志物蛋白。PRMT3在体外14中被证明为非对称的二甲基甲酸酯H4R3,但是,该标记主要由PRMT1沉积,因此设计了具有过度表达PRMT3的外源性检测。与体外发现一致,过度表达野生型PRMT3,但未表达其催化突变体(E338Q)导致H4R3me2a水平(图2A)增加。HEK293T细胞被使用,因为它们有低基底甲基化这个标记(图1C)。该检测结果通过PRMT3选择性抑制剂SGC7077进一步验证,该抑制了PRMT3依赖型H4R3不对称甲基化(图2B)。
PRMT4检测
PRMT4 非对称二甲基酸盐 BAF155 在精氨酸 106415.由于检测 BAF165-R1064me2a 的抗体已市售,通过检测 R1064me2a 标记水平的变化,可以通过西方污点监测细胞中的 PRMT4 活性。PRMT4蛋白的丧失或与PRMT4选择性抑制剂TP-06410的催化活性抑制,导致BAF165-R1064me2a水平下降(图3)。2天的治疗通常足以消除大部分甲基化信号。
PRMT5检测
PRMT5 负责大多数蛋白质精氨酸对称二甲基化。此前有报道称,包括SmBB'在内的各种SMN复合蛋白是PRMT5基板16。PRMT5活性可以通过观察全球对称精氨酸二甲基化或SmBB蛋白质对称二甲基化水平的变化来监测。 PRMT5的击倒,但不是 PRMT1、3、4、6 和 7 的击倒导致全球 Rme2s 水平下降(图 4A)。在大多数细胞系中,用PRMT5选择性抑制剂LLY-28311 和GSK591治疗细胞2-3天抑制了大多数SmBB'Rme2s信号(图4B)。大多数细胞对PRMT5抑制敏感,这会导致细胞增殖和细胞死亡与长期抑制剂暴露的减少。
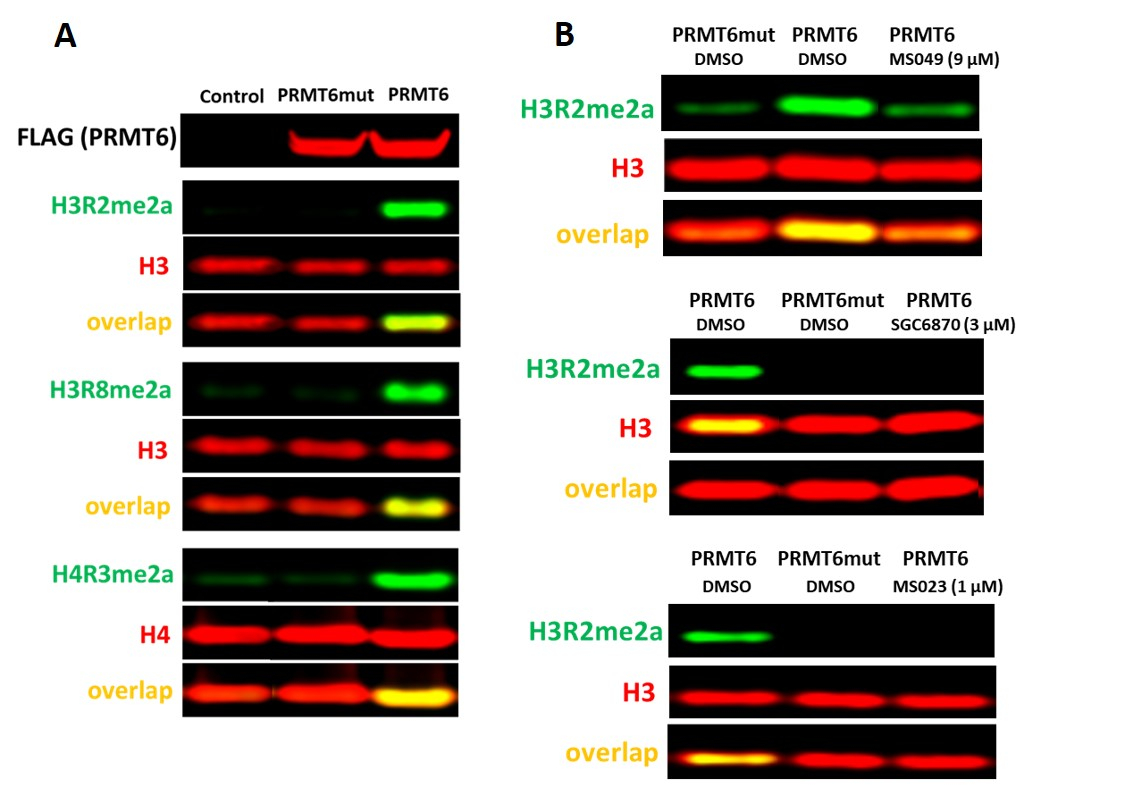
PRMT6检测
据报道,PRMT6是17号细胞中高石H3精氨酸2不对称二甲基化(H3R2me2a)的主要贡献者。在 HEK293T 细胞中 ,PRMT6 击倒 3 天不足以观察 H3R2me2a 水平的显著下降。然而,过度表达野生类型的PRMT6,但不是其催化突变体(V86K/D88A)增加H3R2me2a的水平,以及H3R8me2a和H4R3me2a(图5A)。有几个抑制剂,抑制PRMT6活动具有不同的效力和选择性:选择性,异体PRMT6抑制剂SGC687018,PRMTI型抑制剂MS0238,和PRMT4/6抑制剂MS0499。所有这些抑制PRMT6依赖H3R2(图5B),以及H4R3和H3R8不对称二元化(数据未显示)。
PRMT7检测
PRMT7 单甲基酸盐精氨酸 469 在组成和不可推导形式的 HSP70 (HSPA8 和 HSPA1/6, 分别)12.虽然没有市售抗体,可检测HSP70-R469me1水平,但可以通过泛单甲基抗体检测出该标记。PRMT7蛋白的丧失或与PRMT7选择性抑制剂SGC302712的催化活性抑制,导致HSP70-R469me1(图6A,B)水平下降。SGC3027是一种可渗透细胞的药物,在细胞中通过还原酶转化为PRMT7选择性抑制剂SGC8158,因此细胞电效可能因细胞线而异。几个癌细胞系在高水平上表达难以受精的HSP70等形,由于核原产地的不特定带重叠(图6C),甲基化可能难以检测。因此,对于PRMT7细胞测定,建议主要表达HSPA8(如C2C12)的细胞系,或者由于HSP70主要在细胞质中定位,因此确定首选细胞系细胞质部分的HSP70-R469me1水平。
PRMT8检测
PRMT8是唯一具有组织限制表达模式的PRMT-主要表现在大脑19。它共享80%的序列相似性,并具有类似的基板偏好PRMT119。它不同于PRMT1主要在N终点站,其中肌基化导致PRMT8与等离子膜20的关联。据报道,PRMT8与PRMT1甲基酸酯RNA结合蛋白EWS21。由于PRMT8活性在非神经细胞系中较低,EWS也可以通过PRMT1甲基化,在PRMT1击倒细胞中,PMRT8与EWS共同过度表达的检测方法已经开发出来。由于PRMT1是一种必需的基因,其长期损失会导致细胞死亡,因此PRMT1在使用PRMT8检测之前被击倒3天(图7A)。野生型PRMT8,但不是催化不活跃突变体(E185Q)与EWS共同表达,导致EWS非对称二元化水平增加(图7B)。测试了几种不对称的二甲基苯丙胺抗体,仅用Asym25抗体检测到甲基化。该检测结果通过PRMT I型选择性化学探针MS0238进一步验证,该探针减少了PRMT8依赖外源EWS(图7B)的非对称二甲基化。虽然MS023在体外测定中抑制PRMT8非常有效,但在MS023高浓度的细胞中,需要看到甲基化抑制21。
PRMT9检测
PRMT9被证明对称二甲基酸酯SAP145在精氨酸50822。不幸的是,没有市售抗体可以识别标记。在PRMT9检测中,使用了杨彦忠博士(希望之城贝克曼研究所)赠送的抗体。过度表达时,野生类型但未 R508K 突变 SAP145 由 PRMT9 (图 8A)甲基化。该检测旨在监测内源性SAP145-R508me2s的水平,并验证了化合物X,一个原型PRMT9抑制剂(工作正在进行中,尚未发布),它有力地抑制PRMT9 体外 与纳米分子效力。化合物 X 以剂量依赖的方式降低了 SAP145-R508me2s 水平 (图 8B)。

图1。PRMT1细胞检测。 (A) PRMT1 击倒导致全球不对称精氨酸二甲基化 (Rme2a) 减少,对称精氨酸二甲基化 (Rme2s) 和单甲基化 (Rme1) 水平增加。PRMT1 击倒效率呈现在面板 B.(B) PRMT1 击倒中,可降低高石 H4R3 (H4R3me2a) 的不对称二元化。(C) 基底 H4R3me2a 水平因细胞系而异。(D) I型 PRMT 抑制剂 MS023 以剂量依赖的方式降低 H4R3me2a 水平。MCF7细胞接受MS023治疗2天。(E) 该图表示 H4R3me2a 信号电能的非线性拟合,该信号电能正常化为总高石 H4。MS023 IC50 = 8.3 nM (n=1)。 请单击此处查看此图的较大版本。
{kind=link}

图2。PRMT3细胞测定。 (A) 野生型 (WT) 的过度表达,而不是 PRMT3 的 E338Q 催化突变,可增加 HEK293T 细胞中的 H4R3me2a 水平。细胞被转染为 FLAG 标记的 PRMT3 24 h.(B) PRMT3 选择性抑制剂 SGC707,减少异位 PRMT3 依赖 H4R3 不对称脱甲基化 请单击此处查看此图的更大版本。
{kind=link}

图3。PRMT4细胞测定。 (A) PRMT4 击倒导致 BAF155-R1064 非对称精氨酸二甲基化 (HEK293T 细胞) 的减少。(B) PRMT4选择性抑制剂TP-064可降低BAF155-R1064Rme2a水平。HEK293T细胞用化合物治疗2天。 请单击此处查看此图的较大版本。
{kind=link}

图4。PRMT5细胞检测。 (A) PRMT5 击倒导致全球对称精氨酸二甲基化水平 (MCF7 细胞) 的降低。(B) PRMT5选择性抑制剂GSK591和LLY-283,减少SmBB的对称精氨酸二甲基化(绿色),而SMBB的总水平保持不变(红色)。MCF7细胞用化合物治疗2天。 请单击此处查看此图的较大版本。
{kind=link}

图5。PRMT6细胞检测。 (A) 野生类型 (WT) 的过度表达,但不是 V86K/D88A 催化突变 PRMT6 增加 H4R3me2a、H3R2me2a 和 H3R8me2a 在 HEK293T 细胞中的水平。细胞被转染为 FLAG 标记的 PRMT6 24 h.(B) PRMT6 选择性抑制剂 (SGC6870)、PRMT I 型抑制剂 (MS023) 和 PRMT4/6 抑制剂 (MS049) 降低 PRMT6 依赖 H3R2me2a 水平。 请单击此处查看此图的较大版本。
{kind=link}

图6。PRMT7细胞检测。 (A) PRMT7 淘汰赛导致 HSP70-R469 单甲基化 (HCT116 细胞) 减少。(B) PRMT7选择性抑制剂SGC3027可减少C2C12细胞中的HSP70-R469单体化。细胞用化合物治疗2天。(C) 检测HSP70-R469甲基化的可致病性HSP70(HSPA1/6)与泛单甲基精氨酸抗体(Rme1)可能很难,由于核原产地的重叠不特定波段。建议测量细胞质分数中的 HSP70 甲基化水平。 请单击此处查看此图的较大版本。
{kind=link}

图7。PRMT8细胞检测。 (A) 当PRMT1活动被击倒时,可以检测到 EWS的PRMT8 甲基化。HEK293T细胞被用一个不可推断的 PRMT1 击倒向量进行转化。经过3天的多氧环素治疗,PRMT1水平大幅下降。(B) 当 PRMT1 被击倒时,外源性EWS被过度表达的野生PRMT8型二甲基化,而不是PRMT8的催化突变体(E185Q)。甲基化由高浓度的PRMT I型抑制剂(MS023)降低。HEK293T PRMT1KD 细胞与 FLAG 标记的 PRMT8 野生类型或催化突变体和 GFP 标记的 EWS 共同传输,并与 MS023 一起处理 20 小时。
{kind=link}

图8。PRMT9细胞检测。 (A) 野生类型,但不是R508K突变SAP145是由PRMT9甲基化。HEK293T 细胞与 GFP 标记的 SAP145 一起转染 1 天。(B) 原型 PRMT9 抑制剂 (复合 X) 以剂量依赖的方式减少 PRMT9 依赖 R508 SAP145 的对称二甲基化。HEK293T细胞与化合物一起治疗2天。 请单击此处查看此图的较大版本。
{kind=link}
| 普拉姆特 | 细胞 | 每毫升密度 |
| PRMT1 | 麦克夫 7 | 1 x 105 |
| PRMT3 | 赫克293T | 2 x 105 |
| PRMT4 | 赫克293T | 1 x 105 |
| PRMT5 | 麦克夫 7 | 1 x 105 |
| PRMT6 | 赫克293T | 2 x 105 |
| PRMT7 | C2C12 | 1 x 105 |
| PRMT8 | 赫克293T(PRMT1千瓦)* | 2 x 105 |
| PRMT9 | 赫克293T | 2 x 105 |
| *在PRMT8检测电镀前3天用多氧环素(2μg/mL)治疗细胞 | ||
表1。建议用于 PRMT 检测的细胞类型和密度。
| 普拉姆特 | 微克DNA/24井 | 阿金# | 其他说明 |
| PRMT3 | 0.5 旗- PRMT3 | 164695 | |
| 或 0.5 升 - PRMT3 (E338Q) | 164696 | ||
| PRMT6 | 0.5 旗-PRMT6 | 164697 | |
| 或 0.5 旗-PRMT6 (V86K/D88A) | 164698 | ||
| PRMT8 | 0.05 EWS-GFP | 164701 | |
| 0.45 PRMT8 旗 | 164699 | ||
| 或 0.45 PRMT8 (E185Q) - 标志 | 164700 | ||
| PRMT9 | 0.05 SAP145-GFP | 那 | 希望之城贝克曼研究所杨彦忠博士的礼物 |
| 或0.05 SAP145-R508K-GFP | |||
| 0.45 空矢量 |
表2。推荐用于转染实验的DNA浓度。
| 普拉姆特 | 抗体 | 化学探针(细胞活性IC50) | 负控制 |
| PRMT1 | H4R3me2a (1:2000) | MS023-PRMT类型I | MS094 |
| Rme1 (1:1000) | (PRMT1、PRMT6、PRMT3、PRMT4 IC50 =9、56、1000、5000 nM) | ||
| Rme2s (1:2000) | |||
| Rme2a (1:2000) | |||
| Rme2a (ASYM24, 1:3000) | |||
| Rme2a (ASYM25, 1:2000) | |||
| H4 (1:2000) | |||
| B-行动 (1:500 ) | |||
| PRMT3 | H4 (1:2000) | SGC707 (IC50 = 91 纳米) | XY-1 |
| H4R3me2a (1:2000) | |||
| 旗 (1:5000) | |||
| PRMT4 | BAF155 (1:200) | TP-064 (IC50 = 43 纳米) | TP064N |
| BAF155-R1064me2a (1:3000) | SKI-73 (IC50 = 540 纳米)* | 滑雪-73N* | |
| PRMT5 | 反烟雾(1:100) | 283 (IC50 = 30 nM) | LLY-284 |
| Rme2s (#13222, 1:2000) | 葛兰素史克591 (IC50 = 56 nM) | SGC2096 | |
| PRMT6 | H4R3me2a (1:2000) | SGC6870 (IC50 = 0.9μM) | SGC6870N |
| H4 (1:2000) | MS023-PRMT类型I | MS094 | |
| H3R2me2a (1:2000) | (PRMT1、PRMT6、PRMT3、PRMT4 IC50 =9、56、1000、5000 nM) | ||
| H3R8me2a (1:2000) | MS049(PRMT 4,6 IC50 =970,1400纳米,分别为) | MS049N | |
| H3 (1:5000) | |||
| 旗 (1:5000) | |||
| PRMT7 | Rme1 (1:1000) | SGC3027(IC50=1300纳米)* | SGC3027N* |
| 赫斯普/赫克70 (1:2000)* | |||
| PRMT8 | GFP (1:3000) | MS023 (50 微米) | MS094 |
| Rme2a (ASYM25,1:2000) | |||
| 旗 (1:5000) | |||
| PRMT9 | SAP145 (1:1000) | ||
| SAP145-R508me2s - 希望之城贝克曼研究所杨彦忠博士赠送的实物礼物(1:1000)(PIMID:25737013) | |||
| 次生抗体 | 山羊抗兔IgG-IR800 (1:5000) | ||
| 驴反鼠标 IgG-IR680 (1:5000) | |||
| *-抗体识别HSPA8,HSPA1和HSPA6(测试过度表达的GFP标记蛋白质),*prodrug-IC50可能因不同细胞系而异 | |||
表3。推荐抗体和PRMT化学探针/负控制工具化合物。
| 普拉姆特 | 生物标志物 | 分析读出 | 分析验证 | 推荐的细胞系 | 裁判。 | ||||||||
| PRMT1 | H4R3me2a,雷姆1,雷姆2s,雷梅2a | H4R3me2a 级别标准化为总 H4 全球 Rme1、Rme2a 或 Rme2s 级别,并标准化为 B-actin。 | PRMT1 的击倒降低了基底 H4R3me2a 和全球 Rme2a 水平,并增加了细胞中的全球 Rme1 和 Rme2s 水平(图.1A,B)。PRMT I型化学探针MS023以剂量依赖的方式降低了H4R3me2a的水平(图1D)。 | 细胞在基底H4R3me2a水平(图1C)不同。MCF7 细胞具有较高的基底 H4R3me2a 水平,因此最好用于检测 PRMT1 活性的减少。 | 8 | ||||||||
| PRMT3 | H4R3梅2a | H4R3me2a 甲基化水平由外源标记的 PRMT3 WT 或催化 E338Q 突变体(背景)标准化为总高石 H4 | 野生类型的PRMT3过度表达,但不是其催化突变体(E338Q)增加了H4R3me2a(图2A)。PRMT3选择性抑制剂SGC707减少了PRMT3依附增加的H4R3me2a水平(图2B) | HEK293T细胞具有低基底H4R3me2a水平(图1C),这是更好的监测外源PRMT3活性 | 7 | ||||||||
| PRMT4 | BAF155-R1064me2a | BAF155-R1064me2a 级别正常化为 BAF155 | PRMT4 击倒减少了 BAF155(图 3A) 的非对称二元化。使用PRMT4选择性化学探针(TP-064)进行2天治疗,减少BAF155(图3B)的非对称二元化。 | 任何细胞系 | 10 | ||||||||
| PRMT5 | 斯姆布 - 雷姆 2s | 用平底锅 Rme2s 抗体 (CST) 检测到的 Smbb '- Rme2s 水平标准化为总 Smbb' | PRMT5 的击倒导致 SmBB 对称二甲基化水平降低(图 4A)。使用PRMT5选择性化学探针GSK591和LLY285进行2天治疗,降低了SmBB'-Rme2s水平(图4B)。 | 任何细胞系 | 11 | ||||||||
| PRMT6 | H4R3梅2a H3R2米2a H3R8梅2a | H4R3me2a、H3R2me2a 或 H3R8me2a 甲基化水平通过外源标记的 PRMT6 WT 而增加,但不催化 V86K、D88A 突变体(背景)分别正常化为总高石 H4 或 H3 | 野生型PRMT6的过度表达,但不是其催化突变体(V86K,D88A)增加了H3R2me2a,H3R8me2a和H4R3me2a水平(图5A)。异体PRMT6抑制剂(SGC6870),PRMT I型抑制剂MS023,PRMT4/6抑制剂MS049减少PRMT6依赖性增加H3R2me2a水平(图5B)。 | HEK293T细胞具有低基底H4R3me2a、H3R2me2a和H3R8me2a水平,更适合监测外源PRMT6活性 | 8,9 | ||||||||
| PRMT7 | HSP70-R469me1 | HSP70-Rme1 甲基化水平标准化为 HSP70 总数 | PRMT7 淘汰赛或击倒减少了 HSP70 单体化(图 6A)。使用PRMT7选择性化学探针SGC3027进行2天治疗,以剂量依赖的方式减少PRMT7依赖性HSP70单体化(图6B)。 | C2C12, HT180 几个癌细胞系表示一种可诱导的HSP70形式,其甲基化信号与核原产地的非特异性蛋白质重叠(图6C)。在这种情况下,我们建议分析细胞质分数中的 HSP70 甲基化水平。 | 12 | ||||||||
| PRMT8 | 埃姆斯- 雷梅 2a | 外源性GFP标记的EWS甲基化水平由外源标记PRMT8 WT或E185Q催化突变体(背景)引起,在PRMT1 KO细胞中正常化为总GFP信号。 | 过度表达野生类型PRMT8,但不是催化E185Q突变甲基化异位EWS只在PRMT1 KD细胞(图7A)。PRMT I型化学探针MS023通过PRMT8抑制了外源EWS的非对称二甲基化(图8B)。 | HEK293T PRMT1 KD(不可推断)。 PRMT1击倒导致细胞死亡,因此我们建议使用不可推断的系统。 | 8 | ||||||||
| PRMT9 | SAP145-R508me2s | PRMT9 依赖 SAP145 对称二元化,R508 标准化为 SAP145 | 损失 PRMT9 但不是 PRMT5 会导致 SAP145 的对称二元化减少。GFP 标记的 SAP145 WT,但未标记为 SAP145mut (R508K) 是由 PRMT9(图 8A) 甲基化。使用原型PRMT9抑制剂Copound X进行2天治疗,以剂量依赖的方式降低SAP145-R508me2s水平,图8B)。 | 任何细胞系 | 21 | ||||||||
表4。PRMT 分析摘要。
讨论
在这里,详细的细胞检测协议为PRMT家族的成员描述,使用荧光西斑点方法。选择独特的基材,在单独的PRMT损失或催化抑制时很容易检测到精氨酸甲基化的变化,并且无法由其他家庭成员进行补偿。有些蛋白质是由多个PRMT21,23甲基化,这表明基质特异性重叠,其中一些PRMT只贡献少量细胞标记在给定的蛋白质基板24,25,26,27,例如,PRMT8和PRMT1有助于EWS的甲基化。因此,每次检测都需要通过击倒和/或过度表达实验对基板和抗体进行彻底验证,并进一步验证具有良好特征的选择性抑制剂。已确定PRMT特定基板,可在PRMT丢失/抑制后的2-3天内检测到甲基化标记变化,以避免可能间接影响甲基-精氨酸标记水平的细胞生存能力降低和增殖的复合效应。虽然有可能找到PRMT1、4、5、7和9的独特基材:对于PRMT3、6和8,必须采用功能增益方法。对几个精氨酸甲基特异性抗体进行了各种细胞靶点测试,但没有一种能够在PRMT3和PRMT6击倒后3天内检测出显著变化:因此,生物标志物检测是利用异位表达的酶与催化不活跃突变体一起开发的,作为基线基板甲基化的控制。PRMT8 是一个密切的 PRMT1 同源,并共享类似的基板偏好。由于无法识别PRMT8选择性生物标志物,因此开发了PRMT1击倒细胞中的检测方法,其中PRMT8与EWS共同表达。PRMT1 也是负责 H4R3 不对称甲基化的主要酶,因此,使用 H4R3me2a 作为 PRMT3 和 PRMT6 细胞测定的生物标志物,选择基底 H4R3me2a 水平低的细胞以及催化不活跃突变体作为背景控制。虽然内源性检测是首选,外源性检测证明对测试几个选择性PRMT抑制剂7,8,9的细胞效力是无价的。随着PRMT生物学知识的不断增长,我们期望通过为PRMT3、PRMT6和PRMT8找到更具体的生物标志物蛋白来改进检测。
使用经验证的抗体和适当的控制对PRMT检测性能至关重要。这里推荐的所有抗体都经过击倒和过度表达实验的彻底验证,但是,批次差异,特别是在多克隆抗体的情况下,仍然可能影响其性能。因此,使用遗传方法和化学探针及其密切相关的负控件来确认检测可靠性至关重要。此外,对于需要蛋白质过度表达的PRMT检测,使用催化不活跃突变体以及野生型蛋白质来确定基底甲基化水平至关重要。
这套用于分析细胞中PRMT活性的定量检测方法对科学界大有裨益,因为它可以以最少的设备和有限的技术专长快速、轻松地实施,仅涉及基本的细胞培养和荧光西方印迹技术。建议的PRMT抗体和化学探针也可用于基于活动的蛋白质分析(ABPP)检测,以确定给定ABPP探头的适用性,使用具有竞争力的ABPP格式监测目标参与度并评估目标外效应。这里讨论的检测开发方法也可以推断出其他酶家族,如蛋白莱辛-甲基转移酶和乙酰转移酶。
披露声明
作者没有任何相互竞争的经济利益或其他相互冲突的利益可以申报。
致谢
结构基因组学联合会是一个注册慈善机构(不:1097737),从拜耳股份公司的艾伯维获得资金, 博林格·英格尔海姆、吉恩泰克、加拿大基因组通过安大略基因组研究所[OGI-196]、欧盟和EFPIA通过创新药物倡议2联合承诺[EUbOPEN赠款875510]、詹森、默克KGaA(加拿大和美国的EMD)、辉瑞、竹田和威康信托[106169/ZZ14/Z]。
材料
| Name | Company | Catalog Number | Comments |
| 10 cm TC dishes | Greiner bio-one | 664160 | |
| 24-well TC plates | Greiner bio-one | 662160 | |
| 4–12% Bis-Tris Protein Gels | ThermoFisher Scientiffic | NP0323BOX, NP0322BOX,NP0321BOX | |
| Amersham Hybond P PVDF membrane | Millipore-Sigma | 10600021 | |
| anti-Asym 24 | Millipore-Sigma | 07-414 | |
| anti-Asym 25 | Millipore-Sigma | 09-814 | |
| anti-B-actin | Santa Cruz Biotechnologies | sc-47778 | |
| anti-BAF155 | Santa Cruz Biotechnologies | sc-32763 | |
| anti-BAF155-R1064me2a | Millipore-Sigma | ABE1339 | |
| anti-FLAG (#, 1:5000) | Millipore-Sigma | F4799 | |
| anti-GFP | Clontech | 632381 | |
| anti-H3 | Abcam | ab10799 | |
| anti-H3R2me2a | Millipore-Sigma | 04-848 | |
| anti-H3R8me2a | Rockland | 600-401-I67 | |
| anti-H4 | Abcam | ab174628 | |
| anti-H4R3me2a | Active Motif | 39705 | |
| anti-Hsp/Hsc70 | Enzo | ADI-SPA-820 | |
| anti-PRMT1 | Millipore-Sigma | 07-404 | |
| anti-PRMT3 | Abcam | ab191562 | |
| anti-PRMT4 | Bethyl | #A300-421A | |
| anti-PRMT5 | Abcam | ab109451 | |
| anti-PRMT6 | Abcam | ab47244 | |
| anti-PRMT7 | Abcam | ab179822 | |
| anti-Rme1 | CST | 8015 | |
| anti-Rme2a | CST | 13522 | |
| anti-Rme2s | CST | 13222 | |
| anti-Rme2s (ASYM25), Millipore, , 1:2000) | 09-814 | ||
| anti-SAP145 (Abcam, #, 1:1000) | Abcam | ab56800 | |
| anti-SAP145-R508me2s | kind gift from Dr. Yanzhong Yang, Beckman Research Institute of City of Hope | ||
| anti-SmBB’ | Santa Cruz Biotechnologies | sc-130670 | |
| benzonase | PRODUCED IN-HOUSE | ||
| BSA | Millipore-Sigma | A7906 | |
| C2C12 | gift from Dr. Stephane Richard, McGill University | ||
| cOmplete, EDTA-free Protease Inhibitor Cocktail | Millipore-Sigma | 11873580001 | |
| DMEM | Wisent | 319-005-CL | |
| DMSO | Bioshop | DMS666.100 | |
| donkey anti-mouse IgG-IR680 | Licor | 926-68072 | |
| doxycycline | Millipore-Sigma | D9891 | |
| EDTA | Bioshop | EDT111.500 | |
| FBS | Wisent | 80150 | |
| glycine | Bioshop | GLN002.5 | |
| goat-anti-rabbit IgG-IR800 | Licor | 926-32211 | |
| HEK293T | gift from Dr. Sam Benchimol, York University | ||
| Image Studio Software ver 5.2 | Licor | ||
| Loading buffer: NuPAGE LDS Sample Buffer (4x) | ThermoFisher Scientiffic | NP0007 | |
| MCF7 | ATCC® HTB-22™ | ||
| NaCl | Bioshop | SOD001.1 | |
| NuPAGE MOPS SDS Running Buffer | ThermoFisher Scientiffic | NP0001 | |
| Odyssey Blocking Buffer (dilute 4 x with PBST) | Licor | 927-40000 | Intercept (PBS) Blocking Buffer can also be used # 927-70001 |
| Odyssey CLX Imaging System | Licor | model number 9140 | |
| PBS (tissue culture) | Wisent | 311-010-CL | |
| PBS (western blot) | Bioshop | PBS405.4 | |
| penstrep | Wisent | 450-201-EL | |
| Pierce™ BCA Protein Assay Kit | ThermoFisher Scientiffic | 23225 | |
| SDS | Bioshop | SDS001.1 | |
| skim milk powder | Bioshop | SKI400.500 | |
| TC20 automated cell counter | Biorad | 1450102 | |
| Tripsin-EDTA (0.25%) | Wisent | 325-043-EL | |
| Tris | Bioshop | TRS003.5 | |
| Tritton X-100 | Bioshop | TRX506 | |
| trypan blue | GIBCO | 15250-061 | |
| Tween-20 | Bioshop | TWN510.500 |
参考文献
- Blanc, R. S., Richard, S. Arginine Methylation: The Coming of Age. Molecular Cell. 65 (1), 8-24 (2017).
- Yang, Y., Bedford, M. T. Protein arginine methyltransferases and cancer. Nature Reviews Cancer. 13 (1), 37-50 (2013).
- Bedford, M. T., Richard, S. Arginine methylation an emerging regulator of protein function. Molecular Cell. 18 (3), 263-272 (2005).
- Eaton, S. L., et al. A guide to modern quantitative fluorescent western blotting with troubleshooting strategies. The Journal of Visualized Experiments. (93), e52099 (2014).
- Thandapani, P., O'Connor, T. R., Bailey, T. L., Richard, S. Defining the RGG/RG motif. Molecular Cell. 50 (5), 613-623 (2013).
- Dhar, S., et al. Loss of the major Type I arginine methyltransferase PRMT1 causes substrate scavenging by other PRMTs. Science Reports. 3, 1311 (2013).
- Kaniskan, H. U., et al. A potent, selective and cell-active allosteric inhibitor of protein arginine methyltransferase 3 (PRMT3). Angewandte Chemie International Edition. 54 (17), 5166-5170 (2015).
- Eram, M. S., et al. A Potent, Selective, and Cell-Active Inhibitor of Human Type I Protein Arginine Methyltransferases. ACS Chemical Biology. 11 (3), 772-781 (2016).
- Shen, Y., et al. Discovery of a Potent, Selective, and Cell-Active Dual Inhibitor of Protein Arginine Methyltransferase 4 and Protein Arginine Methyltransferase 6. Journal of Medicinal Chemistry. 59 (19), 9124-9139 (2016).
- Nakayama, K., et al. TP-064, a potent and selective small molecule inhibitor of PRMT4 for multiple myeloma. Oncotarget. 9 (26), 18480-18493 (2018).
- Bonday, Z. Q., et al. LLY-283, a Potent and Selective Inhibitor of Arginine Methyltransferase 5, PRMT5, with Antitumor Activity. ACS Medicinal Chemistry Letters. 9 (7), 612-617 (2018).
- Szewczyk, M. M., et al. Pharmacological inhibition of PRMT7 links arginine monomethylation to the cellular stress response. Nature Communications. 11 (1), 2396 (2020).
- Goulet, I., Gauvin, G., Boisvenue, S., Cote, J. Alternative splicing yields protein arginine methyltransferase 1 isoforms with distinct activity, substrate specificity, and subcellular localization. Journal of Biological Chemistry. 282 (45), 33009-33021 (2007).
- Siarheyeva, A., et al. An allosteric inhibitor of protein arginine methyltransferase 3. Structure. 20 (8), 1425-1435 (2012).
- Stefansson, O. A., Esteller, M. CARM1 and BAF155: an example of how chromatin remodeling factors can be relocalized and contribute to cancer. Breast Cancer Research. 16 (3), 307 (2014).
- Pesiridis, G. S., Diamond, E., Van Duyne, G. D. Role of pICLn in methylation of Sm proteins by PRMT5. Journal of Biological Chemistry. 284 (32), 21347-21359 (2009).
- Guccione, E., et al. Methylation of histone H3R2 by PRMT6 and H3K4 by an mLL complex are mutually exclusive. Nature. 449 (7164), 933-937 (2007).
- Yudao Shen, ., L, F., et al. A First-in-class, Highly Selective and Cell-active Allosteric Inhibitor of Protein Arginine Methyltransferase 6 (PRMT6). BioRxiv. , 1-21 (2020).
- Pahlich, S., Zakaryan, R. P., Gehring, H. Identification of proteins interacting with protein arginine methyltransferase 8: the Ewing sarcoma (EWS) protein binds independent of its methylation state. Proteins. 72 (4), 1125-1137 (2008).
- Lee, J., Sayegh, J., Daniel, J., Clarke, S., Bedford, M. T. PRMT8, a new membrane-bound tissue-specific member of the protein arginine methyltransferase family. Journal of Biological Chemistry. 280 (38), 32890-32896 (2005).
- Kim, J. D., Kako, K., Kakiuchi, M., Park, G. G., Fukamizu, A. EWS is a substrate of type I protein arginine methyltransferase, PRMT8. International Journal of Molecular Medicine. 22 (3), 309-315 (2008).
- Yang, Y., et al. PRMT9 is a type II methyltransferase that methylates the splicing factor SAP145. Nature Communications. 6, 6428 (2015).
- Rakow, S., Pullamsetti, S. S., Bauer, U. M., Bouchard, C. Assaying epigenome functions of PRMTs and their substrates. Methods. 1175, 53-65 (2020).
- Musiani, D., et al. Proteomics profiling of arginine methylation defines PRMT5 substrate specificity. Science Signaling. 12 (575), (2019).
- Musiani, D., Massignani, E., Cuomo, A., Yadav, A., Bonaldi, T. Biochemical and Computational Approaches for the Large-Scale Analysis of Protein Arginine Methylation by Mass Spectrometry. Current Protein and Peptide Science. 21 (7), 725-739 (2020).
- Shishkova, E., et al. Global mapping of CARM1 substrates defines enzyme specificity and substrate recognition. Nature Communications. 8, 15571 (2017).
- Pawlak, M. R., Banik-Maiti, S., Pietenpol, J. A., Ruley, H. E. Protein arginine methyltransferase I: substrate specificity and role in hnRNP assembly. Journal of Cellular Biochemistry. 87 (4), 394-407 (2002).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。