
Method Article
Metodi quantitativi per studiare l'attività delle proteine arginina metiltransferasi 1-9 nelle cellule
In questo articolo
Riepilogo
Questi protocolli forniscono la metodologia utilizzata per valutare l'attività enzimatica dei singoli membri della proteina arginina metiltransferasi (PRMT) nelle cellule. Vengono descritte linee guida dettagliate sulla valutazione dell'attività delle PRMT utilizzando biomarcatori endogeni ed esogeni, anticorpi di riconoscimento metil-arginina e composti degli utensili inibitori.
Abstract
Le proteine metiltransferasi (PRMT) catalizzano il trasferimento di un gruppo metile ai residui di arginina delle proteine del substrato. La famiglia PRMT è composta da nove membri che possono monometilato o residui di arginina dimetilato simmetricamente/asimmetricamente. Sono disponibili diversi anticorpi che riconoscono diversi tipi di metilazione dell'arginina di varie proteine; fornendo così strumenti per lo sviluppo di test di biomarcatore di attività PRMT. I test basati su anticorpi PRMT sono impegnativi a causa della sovrapposizione di substrati e specifiche anticorpali basate su motivi. Vengono discussi questi problemi e la configurazione sperimentale per studiare la metilazione dell'arginina fornita dai singoli PRMT. Attraverso l'attenta selezione dei substrati rappresentativi che sono biomarcatori per otto PRMT su nove, è stato progettato un pannello di test di attività PRMT. Qui vengono riportati i protocolli per i saggi cellulari che misurano quantitativamente l'attività enzimatica dei singoli membri della famiglia PRMT nelle cellule. Il vantaggio dei metodi descritti è la loro semplice performance in qualsiasi laboratorio con coltura cellulare e capacità di macchie occidentali fluorescenti. La specificità del substrato e l'affidabilità degli anticorpi scelti sono state completamente convalidate con approcci di knockdown e sovraespressione. Oltre alle linee guida dettagliate dei biomarcatori e degli anticorpi di dosaggio, vengono fornite anche informazioni sull'uso di una raccolta di composti degli utensili inibitori per i PRMT.
Introduzione
La metilazione dell'arginina è un'importante modifica post-traslazionale che regola le interazioni proteina-proteina e proteina-RNA, giocando così un ruolo importante in vari processi cellulari come la sgiunzione pre-mRNA, il danno al DNA, la risposta alla trascrizione e la trasduzione mediata dal fattore di crescita1,2. L'arginina è metilata dalla proteina arginina metiltransferasi (PRMT) con conseguente monometil arginina (Rme1), dimetilarginina asimmetrica (Rme2a) o dimetilginina simmetrica (Rme2s)3. In base al tipo di metilazione, i PRMT sono classificati in tre gruppi: tipo I (PRMT1, 2, 3, 4, 6 e 8), che catalizzano la dimetilazione mono- e asimmetrica; Tipo II (PRMT5 e PRMT9), che catalizzano la dimetilazione mono- e simmetrica; e tipo III (PRMT7), che può solo monometilato arginina3.
A causa di un numero crescente di anticorpi specifici della metilazione dell'arginina disponibili in commercio, l'attività prmt può essere misurata utilizzando l'blotting occidentale. La macchia occidentale a base fluorescente è la tecnica preferita rispetto al rilevamento chemiluminescente grazie a una maggiore gamma dinamica e linearità, una maggiore sensibilità e consentendo il multiplexing4. Per quantificare i livelli di metilazione proteica, è necessaria la normalizzazione del segnale di metilazione ai livelli proteici totali. Scegliendo gli anticorpi per proteine totali e metilate allevati in diverse specie ospiti (ad esempio, topo e coniglio), è possibile utilizzare anticorpi secondari etichettati con fluorofori diversi e il segnale per entrambi gli anticorpi può essere determinato nella stessa banda campione. Gli anticorpi metil-arginina sono stati sviluppati per identificare e caratterizzare proteine monometilate, asimmetricamente o simmetricamente dimetilate in cui la metil-arginina si trova in un contesto specifico. Poiché la maggior parte dei PRMT metilati ricchi di glicina e arginina all'interno dei loro substrati5, sono stati sollevati diversi anticorpi per i peptidi contenenti ripetizioni monometil o asimmetriche, dimetil-arginina-glicina come D5A12, ASYM24 o ASYM25 e SYM11, rispettivamente. Altri anticorpi metil-arginina sono stati generati contro una libreria peptidica contenente arginina asimmetrica, simmetrica dimetil e monometile in un contesto ripetuto che facilita la rilevazione di metil-arginina in questi particolari contesti6. C'è anche un numero crescente di anticorpi che riconoscono un marchio specifico di arginina su una singola proteina che consentono il rilevamento selettivo della metilazione come l'istone H4R3me2a o BAF155-R1064me2a.
Esistono diversi inibitori PRMT disponibili in commercio, che possono essere utilizzati come strumenti per i test cellulari PRMT. Tuttavia, non tutti sono accuratamente caratterizzati per selettività ed effetti fuori bersaglio e alcuni dovrebbero essere usati con cautela. Il Consorzio Genomico Strutturale, in collaborazione con laboratori accademici e partner farmaceutici, ha sviluppato inibitori PRMT potenti, selettivi e permeabili alle cellule ben caratterizzati (sonde chimiche) che possono essere utilizzati senza restrizioni da parte della comunità scientifica. Informazioni su questi inibitori possono essere trovate https://www.thesgc.org/chemical-probes/epigenetics e https://www.chemicalprobes.org/. Le sonde chimiche sono inibitori di piccole molecole con IC50 o Kd < in vitro 100 nM, selettività superiore a 30 volte rispetto alle proteine della stessa famiglia e significativa attività cellulare a 1 μM. Inoltre, ogni sonda chimica ha un analogo chimico stretto che è inattivo rispetto al bersaglioprevisto 7,8,9,10,11,12.
L'obiettivo di questo protocollo è misurare l'attività cellulare dei singoli membri della famiglia PRMT utilizzando il metodo fluorescente western blot. Qui vengono fornite informazioni dettagliate sui biomarcatori di test convalidati, anticorpi e potenti inibitori attivi delle cellule, nonché preziose strategie per un'implementazione efficace del saggio.
Protocollo
1. Coltivazione e placcatura cellulare
NOTA: Cellule di coltura con mezzi raccomandati e test di routine per la contaminazione da micoplasma. Le celle HEK293T, MCF7 e C2C12 sono state scelte come esempi poiché queste linee cellulari sono state utilizzate con successo nei test PRMT.
- La coltura HEK293T, MCF7 e C2C12 in DMEM è integrata con siero bovino fetale al 10% (FBS), penicillina (100 U mL−1)e streptomicina (100 μg mL−1)in piatti trattati con coltura tissutale (TC) da 10 cm.
- Per il test PRMT8, far crescere le cellule HEK293T inducibili PRMT1 in mezzi contenenti doxiciclina (2 μg/mL) per 3 giorni prima dell'inizio del test.
- Per placcare le celle, rimuovere e scartare i supporti dalla piastra.
- Aggiungere 10 mL di PBS (senza ioni Ca+2 e Mg+2) per lavare le cellule e scartare la soluzione.
- Aggiungere 1 mL di Tripsiderina-EDTA (0,25%), incubare per 1 min a temperatura ambiente (RT), quindi scartare la soluzione. Incubare fino a quando le cellule diventano rotonde e staccarsi dalla piastra. Tocca la piastra per aiutare a staccare le celle, se necessario. Per le cellule difficili da tripinare, come C2C12, incubare la piastra per 1-2 min a 37 °C.
NOTA: Evitare l'esposizione delle cellule alla soluzione di tripina per periodi più lunghi (>10 min) in quanto ridurrà la vitalità cellulare. - Aggiungere 1 mL di supporti prebellici alla piastra e pipettare delicatamente le celle su e giù per scomporsi i grumi cellulari. Trasferire le celle in un tubo da 15 ml e aggiungere 3-5 ml di supporti.
- Per misurare il numero di cellulare, mescolare 10 μL di cellule con 10 μL di blu di Trypan e trasferire 10 μL all'emocitometro o utilizzare qualsiasi altro metodo di conteggio cellulare.
- Diluire le cellule alla densità cellulare raccomandata e mettere 500 μL/pozzo in piastre TC a 24 pozzole(tabella 1). Per i saggi endogeni (PRMT1, PRMT4, PRMT5, PRMT7 e PRMT9), passare al passaggio 3.1.
2. Trasfezione cellulare
- Per i test esogeni (PRMT3, PRMT6, PRMT8) trasfetto cellule HEK293T con la quantità raccomandata di DNA (Tabella 2). Le celle HEK293T sono facili da trasfezionare in modo da poter utilizzare qualsiasi reagente di trasfezione, seguendo le istruzioni del produttore.
3. Trattamento composto
NOTA: Non superare lo 0,1% della concentrazione finale di solfossido di dimetile (DMSO) nei mezzi di coltura. Mantenere la stessa concentrazione dmso in ogni pozzo. Gli inibitori selettivi del PRMT (sonde chimiche) e i loro analoghi inattivi strettamente correlati si trovano nella tabella 3.
- Per i test endogeni (PRMT1, PRMT4, PRMT5, PRMT7 e PRMT9), rimuovere i supporti dalle celle e sostituirli con 500 μL di supporti con composto o DMSO da solo (controllo).
NOTA: Di solito ci vogliono 2 giorni per osservare una diminuzione superiore all'80% dei livelli di metilazione R. - Per i saggi esogeni (PRMT3, PRMT6, PRMT8), rimuovere i supporti 4 ore dopo la trasfezione, aggiungere 500 μL di supporti con composto o DMSO da solo (controllo) e incubare per 20-24 h.
4. Preparazione del llysato cellulare
- Rimuovere tutti i supporti dai pozzi, lavare con 100 μL di PBS per rimuovere i supporti residui e aggiungere 60 μL di tampone di lysis (20 mM Tris-HCl pH 8, 150 mM NaCl, 1 mM di acido etilendiamminatetraacetico (EDTA), 10 mM MgCl2, 0,5% Tritone X-100, 12,5 U mL−1 benzonasi, cocktail completo di inibitore della proteasi senza EDTA) ad ogni pozzo.
- Incubare per meno di 1 minuto a RT, dondolando la piastra per distribuire il tampone di lisi sulle cellule. Quindi aggiungere 3 μL di 20% con solfato di dodecil sodico di sodio (SDS), ad una concentrazione finale dell'1 %, e mescolare tremando delicatamente. Trasferire il lysate in tubi di microcentrifugo e tenerlo sul ghiaccio.
NOTA: Aggiungere benzonasi e cocktail inibitore proteico fresco prima dell'uso. L'aggiunta di benzonasi idrolizza rapidamente gli acidi nucleici, riducendo la viscosità del glisato cellulare.
- Incubare per meno di 1 minuto a RT, dondolando la piastra per distribuire il tampone di lisi sulle cellule. Quindi aggiungere 3 μL di 20% con solfato di dodecil sodico di sodio (SDS), ad una concentrazione finale dell'1 %, e mescolare tremando delicatamente. Trasferire il lysate in tubi di microcentrifugo e tenerlo sul ghiaccio.
- Determinare la concentrazione proteica dei campioni utilizzando bca protein assay kit o utilizzare qualsiasi altro metodo che tollera l'1% di SDS in soluzione.
- Aggiungere 2 μL di glisato e standard proteici (0, 1, 2, 4 e 8 μg/mL di BSA nel tampone di lysis) nei pozzi della piastra ben chiara da 96.
- Mescolare il reagente A con reagente B a un rapporto di 50:1 e aggiungere 200 μL per pozzo. Incubare per 20 minuti a 37 °C e leggere l'assorbanza.
- Regolare la concentrazione proteica con tampone di lysis in modo che sia uguale tra i campioni.
- Aggiungere 20 μL di tampone di carico 4x a 60 μL di llysato cellulare e riscaldare a 95 °C per 5 min. Dopo la denaturazione termica, i lire possono essere conservati a -20 °C.
5. Analisi western macchia
- Caricare 5-20 μg di lysato cellulare totale per l'analisi delle proteine istone e 20-100 μg per altre proteine in un gel proteico Bis-Tris 4-12%.
- Eseguire il gel nel tampone di corsa MOPS SDS (MOPS da 50 mM, base Tris da 50 mM, 0,1% SDS con v, 1 mM EDTA, pH 7,7) per circa 2 h a 100 V o fino a quando la parte anteriore del colorante raggiunge il fondo del gel.
- Se si esegue un trasferimento a umido, assemblare il sandwich di trasferimento nel tampone di trasferimento tris-glicina ghiacciato (25 mM Tris, 192 mM glycine, 20% v/v metanolo e 0,05% w/v SDS).
- Posizionare spugne, carta da filtro, membrana PVDF e gel secondo le istruzioni del produttore. Attivare la membrana PVDF immergendosi in metanolo e gel equilibrato nel tampone di trasferimento per 30 s prima del montaggio.
NOTA: Utilizzare la membrana di soffiatura occidentale PVDF consigliata poiché ha bassa autofluorescenza e idoneità per proteine a basso peso molecolare, come gli istoni(Tabella dei materiali).
- Posizionare spugne, carta da filtro, membrana PVDF e gel secondo le istruzioni del produttore. Attivare la membrana PVDF immergendosi in metanolo e gel equilibrato nel tampone di trasferimento per 30 s prima del montaggio.
- Trasferire le proteine dal gel alla membrana PVDF nel tampone di trasferimento tris-glicina a 70 V per 1,5 h sul ghiaccio.
- Membrana a blocchi per 30 minuti in tampone di blocco (5% con latte in salina tamponata con fosfato, PBS). Risciacquare con tampone di lavaggio (PBST: 0,1% v/v Tween-20 in PBS) e incubare con anticorpi primari nella soluzione di albumina sierice bovina (BSA) (5% di BSA in PBST) durante la notte a 4 °C(tabella 3).
NOTA: Per una conservazione più lunga, sterilizzare il filtro soluzione di BSA, aggiungere lo 0,02% con azide di sodio e mantenerlo a 4 °C. - Lavare la membrana 3 x 5 min con PBST. Quindi incubare con anticorpi capra-anti-coniglio (IR800) e anti-topo d'asino (IR680) nel tampone di blocco raccomandato(Tabella dei materiali, Tabella 3)per 30 minuti a RT e lavare 3 x 5 minuti con PBST.
- Leggere il segnale su un'immagine fluorescente western blot a 800 e 700 nm. Utilizzare preferibilmente lo strumento che consente di creare immagini di bande forti e deboli chiaramente in un'unica immagine con alta sensibilità e gamma dinamica, elevato rapporto segnale-rumore, un avviso quando viene raggiunta una saturazione dell'immagine e il multiplexing di due colori fluorescenti nella stessa banda campione.
- Determinare le intensità della banda per l'analisi western blot utilizzando un software appropriato per l'imaging occidentale fluorescente.
Risultati
Di seguito sono presentati esempi di risultati western blot per test cellulari di singoli PRMT. I dettagli dei saggi sono anche riassunti nella tabella 4.
Saggio PRMT1
PRMT1 è il principale contributore alla dimetilazione asimmetrica dell'istonina 4 arginina 3 (H4R3me2a) nellecelle 13. Dopo la perdita dell'attività prmt1, i livelli globali di Rme1 e Rme2 aumentanosignificativamente di 13. Come mostrato nelle figure 1A e 1B, diversi anticorpi possono essere utilizzati per monitorare i cambiamenti globali in Rme1, Rme2a, Rme2s e H4R3me2a. Una significativa diminuzione dei livelli globali di Rme2a e H4R3me2a e gli aumenti di Rme1 e Rme2 possono essere osservati dopo 3 giorni di abbattimento di PRMT1 (Figura 1A, B). Le linee cellulari differiscono nel segnale basale H4R3me2a, quindi, per facilitare il monitoraggio della perdita di attività prmt1, possono essere utilizzate linee cellulari come MCF7 con alti livelli di metilazione basale(Figura 1C). Il tempo ottimale per osservare l'effetto dell'inibizione del PRMT1, ad esempio al momento del trattamento con inibitore prmt di tipo I MS0238, è di 2 giorni(Figura 1D, 1E). Un trattamento più lungo si traduce in una ridotta vitalità e crescita cellulare.
Saggio PRMT3
Per il test cellulare PRMT3, nessuna proteine biomarcatore selettive che cambi di metilazione potrebbe essere rilevata nella macchia occidentale dopo il knockdown o la sovraespressione prmt3. Prmt3 ha dimostrato di dimetilato asimmetricamente H4R3 in vitro14, tuttavia, il marchio è depositato prevalentemente da PRMT1, e quindi è stato progettato un saggio esogeno con PRMT3 sovraespresso. Coerentemente con i risultati in vitro, l'iperespressione del PRMT3 di tipo selvatico ma non del suo mutante catalitico (E338Q) ha portato a un aumento dei livelli di H4R3me2a(figura 2A). Le cellule HEK293T sono state utilizzate poiché hanno una bassa metilazione basale di questo marchio (Figura 1C). Il saggio è stato ulteriormente convalidato con l'inibitore selettivo PRMT3 SGC7077, che ha inibito la metilazione asimmetrica H4R3 dipendente da PRMT3 (Figura 2B).
Saggio PRMT4
PRMT4 dimetilati asimmetricamente BAF155 all'arginina 106415. Poiché l'anticorpo che rileva BAF165-R1064me2a è disponibile in commercio, l'attività di PRMT4 nelle cellule può essere monitorata dalla macchia occidentale rilevando i cambiamenti nei livelli di marchio R1064me2a. La perdita della proteina PRMT4 o l'inibizione dell'attività catalitica con l'inibitore selettivo PRMT4,TP-064 10, si traduce in una diminuzione dei livelli di BAF165-R1064me2a(Figura 3). Un trattamento di 2 giorni è solitamente sufficiente per rimuovere la maggior parte del segnale di metilazione.
Saggio PRMT5
Prmt5 è responsabile della maggior parte della dimetilazione simmetrica dell'arginina proteica. È stato precedentemente riferito che le varie proteine complesse SMN, tra cui SmBB', sono substrati PRMT516. L'attività di PRMT5 può essere monitorata osservando i cambiamenti nei livelli globali di dimetilazione simmetrica dell'arginina o dimetilazione simmetrica delle proteine smbb. La riduzione di PRMT5, ma non di PRMT1, 3, 4, 6 e 7, comporta una diminuzione dei livelli globali di Rme2 (Figura 4A). Nella maggior parte delle linee cellulari, il trattamento delle cellule con inibitori selettivi PRMT5 LLY-28311 e GSK591 per 2-3 giorni ha soppresso la maggior parte del segnale SmBB'Rme2s (Figura 4B). La maggior parte delle cellule sono sensibili all'inibizione della PRMT5, che si traduce in una diminuzione della proliferazione cellulare e della morte cellulare con esposizione prolungata all'inibitore.
Saggio PRMT6
È stato riferito che prmt6 è il principale contributore alla dimetilazione asimmetrica dell'istonina H3 arginina 2 (H3R2me2a) nelle celle17. Nelle cellule HEK293T, il knockdown prmt6 per 3 giorni non è stato sufficiente per osservare una significativa diminuzione dei livelli di H3R2me2a. Tuttavia, l'iperespressione del tipo selvaggio PRMT6 ma non del suo mutante catalitico (V86K/D88A) aumenta i livelli di H3R2me2a, nonché H3R8me2a e H4R3me2a (Figura 5A). Esistono diversi inibitori che inibiscono l'attività di PRMT6 con diversa potenza e selettività: selettivo, inibitore allosterico di PRMT6 SGC687018, inibitore PRMT di tipo I MS0238e inibitore prmt4/6 MS0499. Tutti questi PRMT6 dipendenti da PRMT6 inibiti H3R2 (Figura 5B), così come la dimetilazione asimmetrica H4R3 e H3R8 (dati non mostrati).
Saggio PRMT7
Prmt7 monometilati arginina 469 in forme costitutive e inducibili di HSP70 (HSPA8 e HSPA1/6, rispettivamente)12. Sebbene non ci siano anticorpi disponibili in commercio, che rilevano i livelli di HSP70-R469me1, il marchio può essere rilevato con anticorpi pan monometil. La perdita della proteina PRMT7 o l'inibizione dell'attività catalitica con l'inibitore selettivo PRMT7, SGC302712, si traduce in una diminuzione dei livelli di HSP70-R469me1(Figura 6A, B). SGC3027 è un profarmaco permeabile alle cellule, che nelle cellule viene convertito per reduttasi nell'inibitore selettivo PRMT7 SGC8158, quindi la potenza cellulare può differire tra le linee cellulari. Diverse linee cellulari tumorali esprimono isoforme HSP70 inducibili ad alti livelli e la metilazione può essere difficile da rilevare a causa di una banda non specifica sovrapposta di origine nucleare (Figura 6C). Pertanto, per il saggio cellulare PRMT7, si raccomandano linee cellulari che esprimono principalmente HSPA8 come C2C12, o poiché HSP70 si localizza principalmente nel citoplasma, determinare i livelli di HSP70-R469me1 nella frazione citoplasmatica delle linee cellulari preferite.
Saggio PRMT8
PRMT8 è l'unico PRMT con un modello di espressione limitato dai tessuti - ampiamente espresso nel cervello19. Condivide l'80% di somiglianza di sequenza e ha una preferenza di substrato simile a PRMT119. Differisce dal PRMT1 principalmente all'N-terminale, dove la miristoilazione si traduce nell'associazione di PRMT8 con la membranaplasmatica 20. È stato riferito che PRMT8 insieme alla proteina PRMT1 legante l'RNA EWS21. Poiché l'attività di PRMT8 è bassa nelle linee cellulari non neuronali e L'EWS può anche essere metilata da PRMT1, un saggio in cui il PMRT8 è co-sovraespresso insieme a EWS nelle cellule knockdown PRMT1. Poiché il PRMT1 è un gene essenziale e la sua perdita a lungo termine provoca la morte cellulare, un sistema inducibile in cui prmt1 viene abbattuto per 3 giorni prima dell'utilizzo nel saggio PRMT8 (Figura 7A). La co-espressione del prmt8 di tipo selvaggio, ma non del mutante cataliticamente inattivo (E185Q), insieme a EWS ha comportato un aumento dei livelli di dimetilazione asimmetrica EWS (Figura 7B). Sono stati testati diversi anticorpi asimmetrici della dimetilginina e la metilazione è stata rilevata solo con anticorpi Asym25. Il saggio è stato ulteriormente convalidato con una sonda chimica selettiva PRMT di tipo I, MS0238, che ha ridotto la dimetilazione asimmetrica dipendente da PRMT8 di EWS esogeno (Figura 7B). Sebbene MS023 sia molto potente nell'inibire prmt8 in saggi in vitro, nelle cellule sono necessarie alte concentrazioni di MS023 per vedere l'inibizione della metilazione21.
Saggio PRMT9
È stato dimostrato che PRMT9 dimetilato simmetricamente SAP145 all'arginina 50822. Sfortunatamente, nessun anticorpo disponibile in commercio può riconoscere il marchio. Per il test PRMT9, sono stati utilizzati anticorpi gentilmente dotati dal Dr. Yanzhong Yang (Beckman Research Institute of City of Hope). Quando sovraespresso, il mutante SAP145, di tipo selvaggio ma non R508K, viene metilato da PRMT9 (Figura 8A). Il saggio è stato progettato per monitorare i livelli di SAP145-R508me2 endogeni ed è stato convalidato con Compound X, un prototipo di inibitore prmt9 (work in progress, non ancora pubblicato), che inibisce potentemente PRMT9 in vitro con potenza nanomolare. Il composto X ha diminuito i livelli di SAP145-R508me2s in modo dose-dipendente (Figura 8B).

Figura 1. Saggio cellulare PRMT1. (A) Il knockdown prmt1 si traduce in una diminuzione della dimetilazione asimmetrica globale dell'arginina (Rme2a) e in un aumento dei livelli di dimetilazione simmetrica dell'arginina (Rme2) e della monometilazione (Rme1). L'efficienza di knockdown PRMT1 è presentata nel pannello B. (B) Il knockdown PRMT1 riduce la dimetilazione asimmetrica dell'istono H4R3 (H4R3me2a). (C) I livelli basali di H4R3me2a differiscono tra le diverse linee cellulari. (D) L'inibitore del PRMT di tipo I MS023 diminuisce i livelli di H4R3me2a in modo dose-dipendente. Le cellule MCF7 sono state trattate con MS023 per 2 giorni. (E) Il grafico rappresenta l'adattamento non lineare delle intensità del segnale H4R3me2a normalizzate al totale dell'istono H4. MS023 IC50 = 8,3 nM (n=1). Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2. Saggio cellulare PRMT3. (A) La sovraespressione del mutante catalitico di tipo selvaggio (WT) ma non E338Q di PRMT3 aumenta i livelli di H4R3me2a nelle cellule HEK293T. Le cellule sono state trasfette con PRMT3 con tag FLAG per inibitore selettivo PRMT3 24 h. (B), SGC707, diminuisce la demetilazione asimmetrica H4R3 dipendente da PRMT3 ectopica Fare clic qui per visualizzare una versione più ampia di questa figura.
{kind=link}

Figura 3. Saggio cellulare PRMT4. (A) Il knockdown prmt4 comporta una diminuzione della dimetilazione asimmetrica dell'arginina BAF155-R1064 (cellule HEK293T). (B) L'inibitore selettivo PRMT4, TP-064, diminuisce i livelli di BAF155-R1064Rme2a. Le cellule HEK293T sono state trattate con composto per 2 giorni. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4. Saggio cellulare PRMT5. (A) Il knockdown prmt5 comporta una diminuzione dei livelli globali di dimetilazione simmetrica dell'arginina (cellule MCF7). (B) Gli inibitori selettivi PRMT5 GSK591 e LLY-283 riducono la dimetilazione simmetrica dell'arginina di SmBB (verde), mentre i livelli totali di SmBB(') rimangono invariati (rosso). Le cellule MCF7 sono state trattate con composti per 2 giorni. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 5. Saggio cellulare PRMT6. (A) La sovraespressione di tipo selvaggio (WT) ma non dei livelli di MUTANTE catalitico V86K/D88A PRMT6 aumenta i livelli di H4R3me2a, H3R2me2a e H3R8me2a nelle cellule HEK293T. Le cellule sono state trasfette con PRMT6 con tag FLAG per inibitore selettivo PRMT6 24 h.(B) (SGC6870), inibitore PRMT di tipo I (MS023) e inibitore PRMT4/6 (MS049) diminuiscono i livelli di H3R2me2a dipendenti da PRMT6. Clicca qui per visualizzare una versione più grande di questa figura.
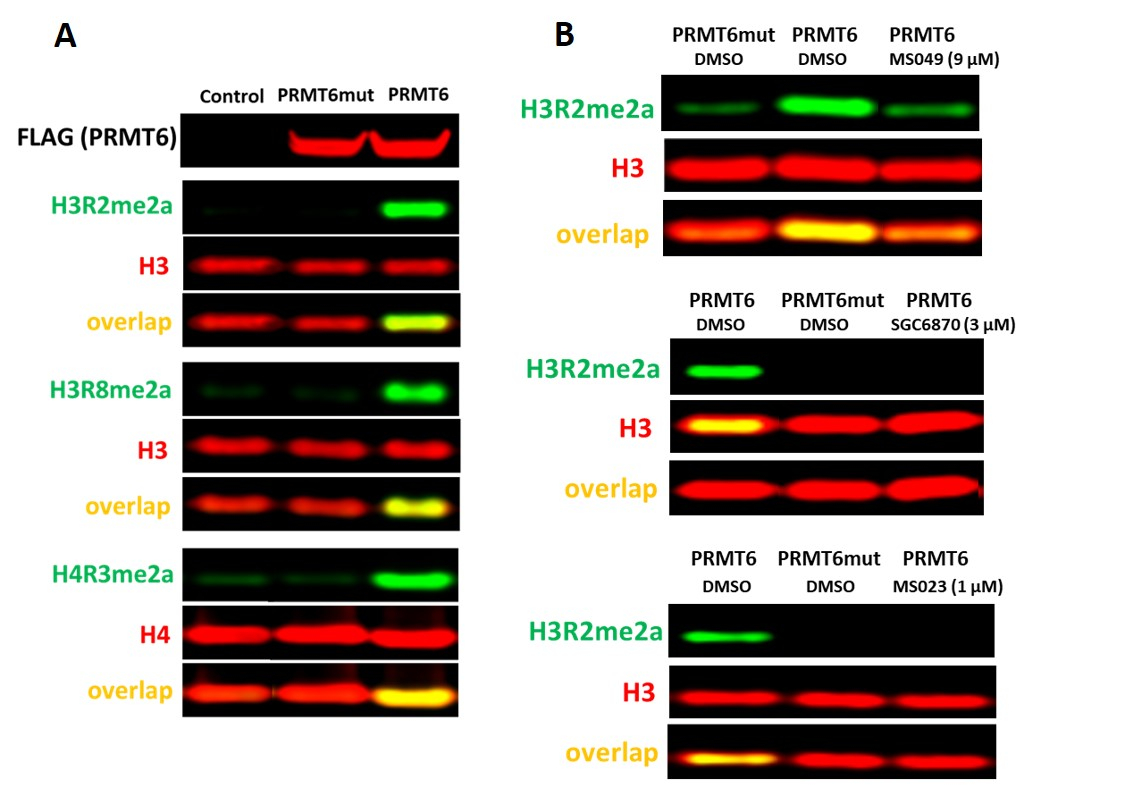{kind=link}

Figura 6. Saggio cellulare PRMT7. (A) Il knockout prmt7 comporta una diminuzione della monometilazione HSP70-R469 (cellule HCT116). (B) Gli inibitori selettivi prmt7, SGC3027, diminuiscono la monometria HSP70-R469 nelle cellule C2C12. Le cellule sono state trattate con composto per 2 giorni. (C) Il rilevamento della metilazione HSP70-R469 dell'HSP70 indutbile (HSPA1/6) con anticorpi pan monometil arginina (Rme1) può essere difficile a causa di una banda non specifica sovrapposta di origine nucleare. Si raccomanda di misurare i livelli di metilazione HSP70 nella frazione citoplasmatica. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 7. Saggio cellulare PRMT8. (A) La metilazione PRMT8 di EWS può essere rilevata quando l'attività di PRMT1 è inibita dal knockdown. Le cellule HEK293T sono state trasdutte con un vettore knockdown PRMT1 inducibile. Dopo 3 giorni di trattamento con doxiciclina, i livelli di PRMT1 sono stati drasticamente ridotti. (B) Quando il PRMT1 viene abbattuto, eWS esogeno viene asimmetricamente dimetilato da PRMT8 di tipo selvatico sovraespresso ma non mutante catalitico (E185Q) di PRMT8. La metilazione è diminuita da un'alta concentrazione di inibitore prmt di tipo I (MS023). Le cellule KD HEK293T PRMT1sono state co-trasfette con tipo selvaggio PRMT8 con tag FLAG o mutante catalitico e EWS con tag GFP e trattate con MS023 per 20 ore. Fare clic qui per visualizzare una versione più ampia di questa figura.
{kind=link}

Figura 8. Saggio cellulare PRMT9. (A) Il tipo selvaggio ma non il mutante R508K SAP145 viene metilato da PRMT9. Le cellule HEK293T sono state trasfette con SAP145 con tag GFP per 1 giorno. (B) Il prototipo di inibitore prmt9 (Composto X) diminuisce la dimetilazione simmetrica R508 dipendente da PRMT9 di SAP145 in modo dipendente dalla dose. Le cellule HEK293T sono state trattate con il composto per 2 giorni. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}
| Prmt | cellule | Densità per ml |
| Prmt1 | McF7 | 1 x 105 |
| Prmt3 | HEK293T | 2 x 105 |
| Prmt4 | HEK293T | 1 x 105 |
| Prmt5 | McF7 | 1 x 105 |
| Prmt6 | HEK293T | 2 x 105 |
| Prmt7 | C2C12 | 1 x 105 |
| Prmt8 | HEK293T (PRMT1 KD)* | 2 x 105 |
| Prmt9 | HEK293T | 2 x 105 |
| *trattare le cellule con doxiciclina (2 μg/mL) 3 giorni prima della placcatura per il test PRMT8 | ||
La tabella 1. Tipi di cellule e densità consigliate per i test PRMT.
| Prmt | μg DNA/24-pozzo | Addgene # | Note aggiuntive |
| Prmt3 | 0.5 FLAG-PRMT3 | 164695 | |
| o 0,5 FLAG-PRMT3 (E338Q) | 164696 | ||
| Prmt6 | 0.5 FLAG-PRMT6 | 164697 | |
| o 0,5 FLAG-PRMT6(V86K/D88A) | 164698 | ||
| Prmt8 | 0.05 EWS-GFP | 164701 | |
| 0,45 PRMT8-FLAG | 164699 | ||
| o 0,45 PRMT8(E185Q)-FLAG | 164700 | ||
| Prmt9 | 0.05 SAP145-GFP | Na | regalo del Dr. Yanzhong Yang, Beckman Research Institute of City of Hope |
| o 0,05 SAP145-R508K-GFP | |||
| 0,45 vettore vuoto |
La tabella 2. La concentrazione di DNA raccomandata per l'esperimento di trasfezione.
| Prmt | anticorpo | Sonda chimica (attività cellulare IC50) | Controllo negativo |
| Prmt1 | H4R3me2a (1:2000) | MS023 -PRMT tipo I | MS094 |
| Rme1 (1:1000) | (PRMT1, PRMT6, PRMT3, PRMT4 IC50 = 9, 56, 1000, 5000 nM, rispettivamente) | ||
| Rme2s (1:2000) | |||
| Rme2a (1:2000) | |||
| Rme2a (ASYM24, 1:3000) | |||
| Rme2a (ASYM25, 1:2000) | |||
| H4 (1:2000) | |||
| B-actin (1:500) | |||
| Prmt3 | H4 (1:2000) | SGC707 (IC50 = 91 nM) | XY-1 |
| H4R3me2a (1:2000) | |||
| FLAG (1:5000) | |||
| Prmt4 | BAF155 (1:200) | TP-064 (IC50 = 43 nM) | TP064N |
| BAF155-R1064me2a (1:3000) | SKI-73 (IC50 = 540 nM)* | SKI-73N* | |
| Prmt5 | anti-SmBB' (1:100) | LLY-283 (IC50 = 30 nM) | LLY-284 |
| Rme2s (#13222, 1:2000) | GSK591 (IC50 = 56 nM) | SGC2096 | |
| Prmt6 | H4R3me2a (1:2000) | SGC6870 (IC50 = 0,9 μM) | SGC6870N |
| H4 (1:2000) | MS023 -PRMT tipo I | MS094 | |
| H3R2me2a ( 1:2000) | (PRMT1, PRMT6, PRMT3, PRMT4 IC50 = 9, 56, 1000, 5000 nM, rispettivamente) | ||
| H3R8me2a (1:2000) | MS049 (PRMT 4, 6 IC50 = 970, 1400 nM, rispettivamente) | MS049N | |
| H3 ( 1:5000) | |||
| FLAG ( 1:5000) | |||
| Prmt7 | Rme1 (1:1000) | SGC3027 (IC50 = 1300 nM) * | SGC3027N* |
| Hsp/Hsc70 (1:2000)* | |||
| Prmt8 | GFP (1:3000) | MS023 (50 μM) | MS094 |
| Rme2a (ASYM25,1:2000) | |||
| FLAG (1:5000) | |||
| Prmt9 | SAP145 (1:1000) | ||
| Regalo sap145-R508me2s del Dr. Yanzhong Yang, Beckman Research Institute of City of Hope (1:1000) (PIMID: 25737013) | |||
| Anticorpi secondari | capra-anti-coniglio IgG-IR800 (1:5000) | ||
| igG-IR680 anti-mouse d'asino (1:5000) | |||
| *- l'anticorpo riconosce HSPA8, HSPA1 e HSPA6 (testato con proteine sovraespresse taggate da GFP), *prodrug - l'IC50 può differire tra varie linee cellulari | |||
La tabella 3. Anticorpi raccomandati e composti della sonda chimica PRMT/utensile di controllo negativo.
| Prmt | Biomarcatore | Lettura del saggio | Convalida del saggio | Linea cellulare consigliata | Ref. | ||||||||
| Prmt1 | H4R3me2a, Rme1, Rme2s, Rme2a | Livelli H4R3me2a normalizzati ai livelli globali H4 H4 Rme1, Rme2a o Rme2 normalizzati a B-actin. | Il knockdown di PRMT1 ha diminuito i livelli basali di H4R3me2a e Rme2a globale e ha aumentato i livelli globali di Rme1 e Rme2 nelle cellule (Fig.1A, B). La sonda chimica PRMT tipo I MS023 ha ridotto i livelli di H4R3me2a in modo dose-dipendente (Fig. 1D). | Le cellule differiscono nei livelli basali di H4R3me2a (Fig. 1C). Le cellule MCF7 hanno alti livelli basali di H4R3me2a, il che lo rende preferibile per i test che monitorano la diminuzione dell'attività di PRMT1. | 8 | ||||||||
| Prmt3 | H4R3me2a | Livelli di metilazione H4R3me2a causati da PRMT3 WT con tag FLAG esogeno o mutante catalitico E338Q (sfondo) normalizzato fino all'istono totale H4 | L'iperespressione di tipo selvaggio PRMT3 ma non del suo mutante catalitico (E338Q) è aumentata di H4R3me2a (Fig. 2A). L'inibitore selettivo PRMT3 SGC707 ha diminuito l'aumento dipendente da PRMT3 dei livelli di H4R3me2a (Fig. 2B) | Le cellule HEK293T hanno bassi livelli basali di H4R3me2a (Fig 1C), che è preferibile per monitorare l'attività esogena di PRMT3 | 7 | ||||||||
| Prmt4 | BAF155-R1064me2a | Livelli di BAF155-R1064me2a normalizzati al totale BAF155 | Il knockdown prmt4 ha diminuito la dimetilazione asimmetrica di BAF155 (Fig. 3A). Il trattamento di 2 giorni con sonda chimica selettiva PRMT4 (TP-064) ha diminuito la dimetilazione asimmetrica di BAF155 (Fig. 3B). | Qualsiasi riga di cella | 10 | ||||||||
| Prmt5 | SmBB'-Rme2 | Livelli di SmBB'-Rme2s rilevati con anticorpi pan Rme2s (CST) normalizzati a SmBB totali' | L'abbattimento di PRMT5 ha comportato una diminuzione dei livelli di dimetilazione simmetrica di SmBB (Fig. 4A). Trattamento di 2 giorni con sonde chimiche selettive PRMT5, GSK591 e LLY285, riduzione dei livelli di SmBB'-Rme2s (Fig. 4B). | Qualsiasi riga di cella | 11 | ||||||||
| Prmt6 | H4R3me2a H3R2me2a H3R8me2a | I livelli di metilazione H4R3me2a, H3R2me2a o H3R8me2a sono aumentati da PRMT6 WT con tag FLAG esogeno ma non da mutante catalitico V86K,D88A (sfondo) normalizzato rispettivamente all'istono totale H4 o H3 | L'iperespressione del tipo selvaggio PRMT6 ma non dei suoi mutanti catalitici (V86K,D88A) ha aumentato i livelli di H3R2me2a, H3R8me2a e H4R3me2a (Fig. 5A). Inibitore allosterico PRMT6 (SGC6870), inibitore PRMT di tipo I MS023 , INIbitore PRMT4/6 MS049 diminuzione dell'aumento dipendente da PRMT6 nei livelli di H3R2me2a (Fig. 5B). | Le cellule HEK293T hanno bassi livelli basali di H4R3me2a, H3R2me2a e H3R8me2a, che è preferibile per monitorare l'attività esogena di PRMT6 | 8,9 | ||||||||
| Prmt7 | HSP70-R469me1 | Livelli di metilazione HSP70-Rme1 normalizzati al totale HSP70 | La monometilazione HSP70 ridotta per ko o knockdown prmt7 (Fig. 6A). Il trattamento di 2 giorni con sonda chimica selettiva PRMT7 SGC3027 ha diminuito la monometilazione HSP70 dipendente da PRMT7 in modo dose-dipendente (Fig. 6B). | C2C12, HT180 Diverse linee cellulari tumorali esprimono una forma induttibile di HSP70 il cui segnale di metilazione si sovrappone a una proteina non specifica di origine nucleare (Fig. 6C). In questo caso, si consiglia di analizzare i livelli di metilazione HSP70 nella frazione citoplasmatica. | 12 | ||||||||
| Prmt8 | EWS-Rme2a | Livelli di metilazione EWS con tag GFP esogeni causati da prmt8 WT o E185Q catalitico (sfondo) con tag FLAG esogeno, normalizzati al segnale GFP totale nelle cellule PRMT1 KO. | Sovraespressione del tipo selvaggio PRMT8 ma non eWS ectopico metilato mutante catalitico E185Q solo nelle cellule PRMT1 KD (Fig. 7A). La sonda chimica PRMT MS023 ha inibito la dimetilazione asimmetrica dell'EWS esogeno da PRMT8 (Fig. 8B). | HEK293T PRMT1 KD (inducibile). L'abbattimento di PRMT1 provoca la morte cellulare, pertanto si consiglia di utilizzare un sistema inducibile. | 8 | ||||||||
| Prmt9 | SAP145-R508me2s | Dimetilazione simmetrica SAP145 dipendente da PRMT9 a R508 normalizzata a SAP145 | La perdita PRMT9 ma non PRMT5 porta a una diminuzione della dimetilazione simmetrica di SAP145. SAP145 WT con tag GFP ma non SAP145mut (R508K) è stato metilato da PRMT9 (Fig. 8A). Trattamento di 2 giorni con Copound X, il prototipo di inibitore prmt9, ridotto i livelli sap145-R508me2s in modo dose-dipendente Fig 8B). | Qualsiasi riga di cella | 21 | ||||||||
La tabella 4. Riepilogo dei saggi PRMT.
Discussione
Qui vengono descritti i protocolli di dosaggio cellulare dettagliati per i membri della famiglia PRMT che utilizzano metodi fluorescenti di soffiatura occidentale. Substrati unici per i quali i cambiamenti nella metilazione dell'arginina possono essere facilmente rilevati in caso di perdita di PRMT individuale o inibizione catalitica e non possono essere compensati da altri membri della famiglia. Alcune proteine sono metilate da più PRMT21,23, suggerendo una sovrapposizione nella specificità del substrato in cui alcuni PRMT contribuiscono solo a una piccola quantità di marchio cellulare in un dato substrato proteico24,25,26,27, ad esempio, sia PRMT8 che PRMT1 contribuiscono alla metilazione di EWS. Pertanto, ogni saggio richiedeva una convalida approfondita di substrati e anticorpi con esperimenti knockdown e/o overexpression e un'ulteriore convalida con inibitori selettivi ben caratterizzati. Sono stati identificati substrati specifici PRMT per i quali è stato possibile rilevare cambiamenti del segno di metilazione entro 2-3 giorni dopo la perdita/inibizione prmt per evitare effetti di composizione di ridotta vitalità e proliferazione cellulare che possono influenzare indirettamente i livelli di marca metil-arginina. Sebbene sia stato possibile trovare substrati unici per PRMT1, 4, 5, 7 e 9; per PRMT3, 6 e 8 è stato necessario utilizzare l'approccio del guadagno di funzione. Diversi anticorpi specifici dell'arginina metile sono stati testati per vari bersagli cellulari, ma nessuno è stato in grado di rilevare cambiamenti significativi entro 3 giorni dal knockdown PRMT3 e PRMT6; pertanto, i test del biomarcatore sono stati sviluppati usando enzimi espressi ectopicamente insieme a mutanti cataliticamente inattivi, che servivano come controllo per la metilazione del substrato di base. PRMT8 è un omologo PRMT1 vicino e condivide preferenze di substrato simili. Poiché non è stato possibile identificare un biomarcatore selettivo PRMT8, è stato sviluppato un saggio nelle cellule di abbattimento PRMT1, in cui PRMT8 è stato co-espresso insieme a EWS. Prmt1 è anche un enzima importante responsabile della metilazione asimmetrica H4R3, quindi, per utilizzare H4R3me2a come biomarcatore per test cellulari PRMT3 e PRMT6, sono state scelte cellule con bassi livelli basali di H4R3me2a e mutanti cataliticamente inattivi sono stati usati come controllo di fondo. Sebbene siano preferiti test endogeni, i saggi esogeni si rivelano inestimabili per testare la potenza cellulare di diversi inibitori selettivi del PRMT7,8,9. Con la crescente conoscenza della biologia PRMT, prevediamo di migliorare i test trovando proteine biomarcatori più specifiche per PRMT3, PRMT6 e PRMT8.
L'uso di anticorpi convalidati e controlli appropriati sono fondamentali per le prestazioni del saggio PRMT. Tutti gli anticorpi qui raccomandati sono stati accuratamente convalidati da esperimenti di knockdown e sovraespressione, tuttavia, le differenze da lotto a lotto, specialmente nel caso di anticorpi policlonali, possono ancora influenzare le loro prestazioni. Pertanto, è fondamentale utilizzare metodi genetici e sonde chimiche insieme ai loro controlli negativi strettamente correlati per confermare l'affidabilità del saggio. Inoltre, per i test PRMT che richiedono un'iperespressione proteica, è fondamentale utilizzare mutanti cataliticamente inattivi insieme a proteine di tipo selvatico per determinare i livelli di metilazione basale.
Questa raccolta di saggi quantitativi per profilare l'attività dei PRMT nelle cellule può essere ampiamente utile per la comunità scientifica poiché può essere implementata rapidamente e facilmente con attrezzature minime e competenze tecniche limitate, coinvolgendo solo la lavorazione cellulare di base e le tecniche fluorescenti di soffiatura occidentale. Gli anticorpi raccomandati e le sonde chimiche per I PRMT possono anche essere utilizzati per test di profilazione proteica basati sull'attività (ABPP) per stabilire l'idoneità di una determinata sonda ABPP, monitorare l'impegno del bersaglio e valutare gli effetti fuori bersaglio utilizzando il formato ABPP competitivo. Gli approcci di sviluppo del saggio qui discussi possono anche essere estrapolati per altre famiglie enzimatiche come la proteina llysina-metiltransferasi e l'acetiltransferasi.
Divulgazioni
Gli autori non hanno interessi finanziari concorrenti o altri interessi contrastanti da dichiarare.
Riconoscimenti
Il Consorzio di Genomica Strutturale è un ente di beneficenza registrato (n. 1097737) che riceve fondi da AbbVie, Bayer AG, Boehringer Ingelheim, Genentech, Genome Canada attraverso l'Ontario Genomics Institute [OGI-196], l'UE e l'EFPIA attraverso l'impresa comune Innovative Medicines Initiative 2 [sovvenzione EUbOPEN 875510], Janssen, Merck KGaA (alias EMD in Canada e negli Stati Uniti), Pfizer, Takeda e wellcome trust [106169/ZZ14/Z].
Materiali
| Name | Company | Catalog Number | Comments |
| 10 cm TC dishes | Greiner bio-one | 664160 | |
| 24-well TC plates | Greiner bio-one | 662160 | |
| 4–12% Bis-Tris Protein Gels | ThermoFisher Scientiffic | NP0323BOX, NP0322BOX,NP0321BOX | |
| Amersham Hybond P PVDF membrane | Millipore-Sigma | 10600021 | |
| anti-Asym 24 | Millipore-Sigma | 07-414 | |
| anti-Asym 25 | Millipore-Sigma | 09-814 | |
| anti-B-actin | Santa Cruz Biotechnologies | sc-47778 | |
| anti-BAF155 | Santa Cruz Biotechnologies | sc-32763 | |
| anti-BAF155-R1064me2a | Millipore-Sigma | ABE1339 | |
| anti-FLAG (#, 1:5000) | Millipore-Sigma | F4799 | |
| anti-GFP | Clontech | 632381 | |
| anti-H3 | Abcam | ab10799 | |
| anti-H3R2me2a | Millipore-Sigma | 04-848 | |
| anti-H3R8me2a | Rockland | 600-401-I67 | |
| anti-H4 | Abcam | ab174628 | |
| anti-H4R3me2a | Active Motif | 39705 | |
| anti-Hsp/Hsc70 | Enzo | ADI-SPA-820 | |
| anti-PRMT1 | Millipore-Sigma | 07-404 | |
| anti-PRMT3 | Abcam | ab191562 | |
| anti-PRMT4 | Bethyl | #A300-421A | |
| anti-PRMT5 | Abcam | ab109451 | |
| anti-PRMT6 | Abcam | ab47244 | |
| anti-PRMT7 | Abcam | ab179822 | |
| anti-Rme1 | CST | 8015 | |
| anti-Rme2a | CST | 13522 | |
| anti-Rme2s | CST | 13222 | |
| anti-Rme2s (ASYM25), Millipore, , 1:2000) | 09-814 | ||
| anti-SAP145 (Abcam, #, 1:1000) | Abcam | ab56800 | |
| anti-SAP145-R508me2s | kind gift from Dr. Yanzhong Yang, Beckman Research Institute of City of Hope | ||
| anti-SmBB’ | Santa Cruz Biotechnologies | sc-130670 | |
| benzonase | PRODUCED IN-HOUSE | ||
| BSA | Millipore-Sigma | A7906 | |
| C2C12 | gift from Dr. Stephane Richard, McGill University | ||
| cOmplete, EDTA-free Protease Inhibitor Cocktail | Millipore-Sigma | 11873580001 | |
| DMEM | Wisent | 319-005-CL | |
| DMSO | Bioshop | DMS666.100 | |
| donkey anti-mouse IgG-IR680 | Licor | 926-68072 | |
| doxycycline | Millipore-Sigma | D9891 | |
| EDTA | Bioshop | EDT111.500 | |
| FBS | Wisent | 80150 | |
| glycine | Bioshop | GLN002.5 | |
| goat-anti-rabbit IgG-IR800 | Licor | 926-32211 | |
| HEK293T | gift from Dr. Sam Benchimol, York University | ||
| Image Studio Software ver 5.2 | Licor | ||
| Loading buffer: NuPAGE LDS Sample Buffer (4x) | ThermoFisher Scientiffic | NP0007 | |
| MCF7 | ATCC® HTB-22™ | ||
| NaCl | Bioshop | SOD001.1 | |
| NuPAGE MOPS SDS Running Buffer | ThermoFisher Scientiffic | NP0001 | |
| Odyssey Blocking Buffer (dilute 4 x with PBST) | Licor | 927-40000 | Intercept (PBS) Blocking Buffer can also be used # 927-70001 |
| Odyssey CLX Imaging System | Licor | model number 9140 | |
| PBS (tissue culture) | Wisent | 311-010-CL | |
| PBS (western blot) | Bioshop | PBS405.4 | |
| penstrep | Wisent | 450-201-EL | |
| Pierce™ BCA Protein Assay Kit | ThermoFisher Scientiffic | 23225 | |
| SDS | Bioshop | SDS001.1 | |
| skim milk powder | Bioshop | SKI400.500 | |
| TC20 automated cell counter | Biorad | 1450102 | |
| Tripsin-EDTA (0.25%) | Wisent | 325-043-EL | |
| Tris | Bioshop | TRS003.5 | |
| Tritton X-100 | Bioshop | TRX506 | |
| trypan blue | GIBCO | 15250-061 | |
| Tween-20 | Bioshop | TWN510.500 |
Riferimenti
- Blanc, R. S., Richard, S. Arginine Methylation: The Coming of Age. Molecular Cell. 65 (1), 8-24 (2017).
- Yang, Y., Bedford, M. T. Protein arginine methyltransferases and cancer. Nature Reviews Cancer. 13 (1), 37-50 (2013).
- Bedford, M. T., Richard, S. Arginine methylation an emerging regulator of protein function. Molecular Cell. 18 (3), 263-272 (2005).
- Eaton, S. L., et al. A guide to modern quantitative fluorescent western blotting with troubleshooting strategies. The Journal of Visualized Experiments. (93), e52099 (2014).
- Thandapani, P., O'Connor, T. R., Bailey, T. L., Richard, S. Defining the RGG/RG motif. Molecular Cell. 50 (5), 613-623 (2013).
- Dhar, S., et al. Loss of the major Type I arginine methyltransferase PRMT1 causes substrate scavenging by other PRMTs. Science Reports. 3, 1311 (2013).
- Kaniskan, H. U., et al. A potent, selective and cell-active allosteric inhibitor of protein arginine methyltransferase 3 (PRMT3). Angewandte Chemie International Edition. 54 (17), 5166-5170 (2015).
- Eram, M. S., et al. A Potent, Selective, and Cell-Active Inhibitor of Human Type I Protein Arginine Methyltransferases. ACS Chemical Biology. 11 (3), 772-781 (2016).
- Shen, Y., et al. Discovery of a Potent, Selective, and Cell-Active Dual Inhibitor of Protein Arginine Methyltransferase 4 and Protein Arginine Methyltransferase 6. Journal of Medicinal Chemistry. 59 (19), 9124-9139 (2016).
- Nakayama, K., et al. TP-064, a potent and selective small molecule inhibitor of PRMT4 for multiple myeloma. Oncotarget. 9 (26), 18480-18493 (2018).
- Bonday, Z. Q., et al. LLY-283, a Potent and Selective Inhibitor of Arginine Methyltransferase 5, PRMT5, with Antitumor Activity. ACS Medicinal Chemistry Letters. 9 (7), 612-617 (2018).
- Szewczyk, M. M., et al. Pharmacological inhibition of PRMT7 links arginine monomethylation to the cellular stress response. Nature Communications. 11 (1), 2396 (2020).
- Goulet, I., Gauvin, G., Boisvenue, S., Cote, J. Alternative splicing yields protein arginine methyltransferase 1 isoforms with distinct activity, substrate specificity, and subcellular localization. Journal of Biological Chemistry. 282 (45), 33009-33021 (2007).
- Siarheyeva, A., et al. An allosteric inhibitor of protein arginine methyltransferase 3. Structure. 20 (8), 1425-1435 (2012).
- Stefansson, O. A., Esteller, M. CARM1 and BAF155: an example of how chromatin remodeling factors can be relocalized and contribute to cancer. Breast Cancer Research. 16 (3), 307 (2014).
- Pesiridis, G. S., Diamond, E., Van Duyne, G. D. Role of pICLn in methylation of Sm proteins by PRMT5. Journal of Biological Chemistry. 284 (32), 21347-21359 (2009).
- Guccione, E., et al. Methylation of histone H3R2 by PRMT6 and H3K4 by an mLL complex are mutually exclusive. Nature. 449 (7164), 933-937 (2007).
- Yudao Shen, ., L, F., et al. A First-in-class, Highly Selective and Cell-active Allosteric Inhibitor of Protein Arginine Methyltransferase 6 (PRMT6). BioRxiv. , 1-21 (2020).
- Pahlich, S., Zakaryan, R. P., Gehring, H. Identification of proteins interacting with protein arginine methyltransferase 8: the Ewing sarcoma (EWS) protein binds independent of its methylation state. Proteins. 72 (4), 1125-1137 (2008).
- Lee, J., Sayegh, J., Daniel, J., Clarke, S., Bedford, M. T. PRMT8, a new membrane-bound tissue-specific member of the protein arginine methyltransferase family. Journal of Biological Chemistry. 280 (38), 32890-32896 (2005).
- Kim, J. D., Kako, K., Kakiuchi, M., Park, G. G., Fukamizu, A. EWS is a substrate of type I protein arginine methyltransferase, PRMT8. International Journal of Molecular Medicine. 22 (3), 309-315 (2008).
- Yang, Y., et al. PRMT9 is a type II methyltransferase that methylates the splicing factor SAP145. Nature Communications. 6, 6428 (2015).
- Rakow, S., Pullamsetti, S. S., Bauer, U. M., Bouchard, C. Assaying epigenome functions of PRMTs and their substrates. Methods. 1175, 53-65 (2020).
- Musiani, D., et al. Proteomics profiling of arginine methylation defines PRMT5 substrate specificity. Science Signaling. 12 (575), (2019).
- Musiani, D., Massignani, E., Cuomo, A., Yadav, A., Bonaldi, T. Biochemical and Computational Approaches for the Large-Scale Analysis of Protein Arginine Methylation by Mass Spectrometry. Current Protein and Peptide Science. 21 (7), 725-739 (2020).
- Shishkova, E., et al. Global mapping of CARM1 substrates defines enzyme specificity and substrate recognition. Nature Communications. 8, 15571 (2017).
- Pawlak, M. R., Banik-Maiti, S., Pietenpol, J. A., Ruley, H. E. Protein arginine methyltransferase I: substrate specificity and role in hnRNP assembly. Journal of Cellular Biochemistry. 87 (4), 394-407 (2002).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati