
Method Article
对成年 冈比亚按蚊 的有效口服RNA干扰(RNAi)给药
摘要
口服由细菌产生的dsRNA,一种常规用于 秀丽隐杆线虫 的RNA干扰(RNAi)递送方法,在这里成功地应用于成年蚊子。我们的方法允许在不使用注射的情况下进行稳健的反向遗传学研究和传播阻断载体研究。
摘要
二十年来,RNA干扰一直是逆向遗传分析中广泛使用的工具。在成年蚊子中,双链RNA(dsRNA)给药主要 通过 注射完成,这需要很长时间,不适合野外应用。为了克服这些局限性,我们在这里提出了一种更有效的方法,通过口服递送dsRNA到成年 冈比亚按蚊来稳健地活化RNAi。在 大肠杆菌 菌株HT115(DE3)中产生长dsRNA,并在10%蔗糖中将热杀灭的含dsRNA细菌的浓缩悬浮液 随意地提供给 成年蚊子。在治疗期间每2天更换一次棉球。使用这种方法靶向 双性 (参与性别分化的基因)或 叉头 (编码唾液腺转录因子)分别导致靶基因表达和/或蛋白质免疫荧光信号降低,分别通过定量实时荧光显微镜(qRT-PCR)或荧光共聚焦显微镜测量。还观察到唾液腺形态缺陷。这种高度灵活,用户友好,低成本,省时的dsRNA递送方法可以广泛适用于对昆虫载体生理学及其他方面重要的靶基因。
引言
许多疾病都是由蚊子传播的,使蚊子生理学和遗传学研究成为一项重要任务。在过去的20年中,RNAi在这些生物体中的使用一直很突出,并允许许多蚊子基因1,2,3,4,5的功能表征。dsRNA递送最常用的技术是显微注射,其缺点是可以伤害蚊子,并且需要大量的时间和精力。RNAi的口服给药方法已经过测试,但主要在蚊子的幼虫阶段6,7,8,9。成年蚊子中口服dsRNA尚未得到充分探索,可以成为研究病媒生物学和病媒控制的有用工具。
疟疾由 按蚊 传播,当受感染的雌性蚊子从未受感染的宿主身上取血粉并注射含有疟疾寄生虫的唾液时10。为了最终在蚊子的唾液中传播,寄生虫必须克服许多障碍,包括逃避蚊子的免疫系统,穿越中肠屏障以及侵入唾液腺11。蚊子唾液腺(SG)结构是寄生虫入侵的关键,并且该结构由关键的唾液腺表达转录因子以及性二态性的决定因素控制。几种高度保守的转录因子对于唾液腺的细胞规格和稳态维持以及用于血液喂养的唾液蛋白的产生和分泌是必需的12,13,14。叉头(Fkh)是一种有翅螺旋转录因子,作为昆虫SG结构和功能的主要调节因子(基于果蝇和蚕蛾的研究)15,16,17,18,19,20。在 果蝇 SGs中,Fkh与Sage(一种SG特异性的基本螺旋 - 环螺旋(bHLH)转录因子)一起发挥作用,以促进SG生存和唾液产生19。 果蝇 中唾液产生的重要阳性共调节因子是CrebA,这是一种经过充分研究的亮氨酸拉链转录因子,可上调分泌途径基因21,22,23的表达。在女性唾液腺中也存在很强的形态分化,这可能起着关键作用,不仅在血液喂养中,而且在寄生虫侵入这种组织的能力方面也是如此24。
许多参与确定唾液腺存活、结构、生理学和性二态性的基因具有复杂的时空表达谱25,26,27,并且dsRNA诱导RNAi的传统递送方法并不总是有效地靶向该组织或其他组织中的这些类型的基因。然而,在幼虫期埃及伊蚊和冈比亚蚊的口服递送dsRNA已成功用于沉默dsx基因9,28的雌性特异性形式。以前在蚊子唾液腺中使用dsRNA的研究发现,尽管需要大量的dsRNA,但沉默效果相对持久(至少13天)29。在这里,测试了热杀灭的大肠杆菌菌株HT115(DE3)表达dsx,fkh或CrebA的序列特异性dsRNA在成年雌性蚊子中诱导这些基因的RNAi沉默的能力。口服dsRNA诱导的冈比亚猪笼草基因敲低,mRNA水平明显降低,表型与这些基因的功能丧失一致。因此,这种方法可能会敲低各种唾液腺基因的功能。
研究方案
1. 将dsRNA克隆到 大肠杆菌 表达载体中
- 选择要插入到适当的载体中以表达dsRNA的靶基因序列。使用以下方法从 Vectorbase.org 检索表达式值。
- 在主页搜索框中搜索目的基因(例如 ,表1)。
- 在生成的基因页面中,导航到 8.转录组学 部分。
- 寻找列出的相关RNA-seq和微阵列基因表达实验。
- 将感兴趣的值转录到电子表格软件中并创建数据表。
- 选择至少使用一个T7启动子的市售质粒。如果所选质粒只有一个T7启动子(与大多数商业质粒一样),请在反向引物中加入第二个T7启动子,用于扩增目的基因的dsDNA。
注意:靶基因的dsRNA序列可以使用Web应用程序E-RNAi选择用于RNAi试剂30的设计。长dsRNA(约400 bp)或短发夹dsRNA(shRNA)都可以根据特定的基因序列进行设计。在克隆之前,应对这些序列进行扩增和测序以进行身份确认。本研究中使用的选定基因区域,质粒和启动子列在 补充文件1中。 - 根据前面描述的简单一步过程9,31执行克隆。为此,纯化PCR产物并连接到线性化的质粒DNA。使用连接产物进行感受态 大肠杆菌 细胞32的热休克转化。通过蓝/白筛选选择转化的细胞。使用 T7 引物 PCR 确认插入片段的方向,并使用 M13 引物确认序列。
注意:当选择用于转化的质粒携带编码β半乳糖苷酶的lacZ基因时,可以使用白色/蓝色筛选。白色菌落应在lacZ内包含所需的插入片段,并且可以选择以进一步确认靶序列33的存在和取向。 - 从第一次转化中纯化质粒,并用它来转化前述 的大肠杆菌 HT115(DE3)。在确认带有插入物的质粒存在于主管 的大肠杆菌 HT115(DE3)中后,使细菌的甘油储备一次性使用。
注意:应获取或准备在每次实验中使用的适当的非相关对照dsRNA。在这种情况下,使用拟南芥中不相关的基因aintegumenta(蚂蚁)的序列。
2. 制备表达dsRNA的热杀灭细菌
- 从含有100μg/ mL氨苄西林和12.5μg/ mL四环素的50mLLuria Broth(LB)中培养含有表达dsRNA的DsRNA的大 肠 杆菌菌株HT115(DE3)的培养物,在37°C下的平台振荡器(180 rpm)上生长12小时。
- 将细菌培养物(1:1000)稀释到含有100μg/ mL氨苄西林和12.5μg/ mL四环素的2x酵母胰蛋白(2x YT)培养基中。
- 通过添加40μM(终浓度)异丙基β-D-1-硫代四乙酸吡喃糖苷(IPTG)来诱导dsRNA的产生。
- 当细胞达到O.D.600 = 0.4时,大约在37°C诱导2小时后,以180rpm搅拌,制备热杀灭细菌的浓缩悬浮液,如Taracena等人9所述。通过离心(4000×g,4°C,10分钟)沉淀细胞,并在一体积的磷酸钠缓冲液(PBS)中洗涤细胞。
- 在相同条件下再次旋转,在PBS中重新悬浮至初始体积的1/100,并在70°C下放置1小时。
- 制作400μL等分试样热杀灭细菌,并将这些等分试样储存在-20°C直至进一步使用(不要储存超过一周)。这种热杀灭细菌的悬浮液含有用于RNAi实验的特定dsRNA。对靶基因dsRNA细菌和每次实验中使用的不相关的dsRNA对照进行该程序。
3. 用表达dsRNA的热杀菌喂养蚊子
- 解冻一等分试样的dsRNA(HT115(DE3)细菌悬浮液),并与1.6mL含有0.2%对羟基苯甲酸甲酯的12%糖溶液混合。
- 将一个小棉球浸泡在该溶液中,并将浸泡的棉球放入装有5天大蚊子的笼子内。确保蚊子以这种溶液为食,同时拾取糖和含有dsRNA的细菌。
- 每隔一天更换浸泡在dsRNA糖溶液中的棉球,连续8天。
- 将蚊子笼保持在恒定条件下,即27°C和80%相对湿度,光周期为12小时:12小时:光照:黑暗光周期,由30分钟的黎明和30分钟的黄昏期隔开。
4. 测定靶基因表达水平
- 通过将容器放在冰上一分钟或直到蚊子停止移动来冷麻醉蚊子。一旦蚊子被麻醉,将它们放在寒冷的表面上,以隔离雌性进行解剖。
- 将70%乙醇喷洒到蚊子上,并将它们放在装有PBS的玻璃表面上。用一对镊子,稳定地固定蚊子头,非常缓慢地拉动胸部,使唾液腺释放到PBS中。
- 将唾液腺保存在冰冷的PBS中,直到10个人被解剖。使用硫氰酸胍-苯酚-氯仿法将10个SG用于RNA提取。将RNA沉淀悬浮在30μL无RNase水中。
- 使用1μL等分试样在上一步中从SG中提取的RNA,以读取260和280nm处的吸光度,并通过与稀释因子相乘来计算每个样品的RNA浓度。260/280比值~2.0表示RNA质量好。
- 使用1μg纯化的RNA使用商业逆转录试剂盒合成互补DNA(cDNA)。
- 根据制造商的建议,对cDNA进行1:10稀释以制备RT-PCR反应。对于每个样品,为靶基因准备一个反应,并同时与管家(HK)基因建立反应。将每个基因反应设置在一式三份技术中,以消除方法随机变异的影响。
注意:在这里, An. gambiae 核糖体 S7 基因(GeneBank:L20837.1)和 肌动蛋白 (VectorBase:AGAP000651)已被用作HK基因。 - 按照SYBR-green制造商的适应症,使用最终浓度为300 nM的所有引物。用标准PCR条件扩增:95°C 10分钟,然后在95°C下循环40次,每次15秒,在60°C下循环60秒。
注意:为了量化基因表达,使用δ-δ-Ct方法(ΔΔCt)。Delta Ct(ΔCt)是靶基因的Ct和管家基因的Ct之间的差异。ΔΔCt是实验组ΔCt与对照组ΔCt之间的差异35。
5. 表型评估:成功采血
- 为了评估血液进食的能力,将15只雌性蚊子的组用目标处理并控制dsRNA放在小笼子(直径12厘米)上,并饿死它们4小时。
- 使用设定为37°C的循环水浴,玻璃蚊子喂食器(直径24毫米)和parafilm膜,为蚊子提供除锡的羊血。
注意:血液可以从商业供应商处获得,该供应商无菌地从美国原产的健康供体动物中提取血液,并在没有抗凝剂或添加剂的情况下手动脱洗。 - 通过直接观察,计数并记录从前五名女性中成功获得血粉的探测尝试次数,以便在每组中完全充血。
注意:为了避免蚊子的显着代谢变化,这可能会干扰影响寻血行为的能量资源,饥饿保持在最低限度(4小时)。因此,并非每只蚊子都会热切地寻找血粉,我们将充血雌性的数量限制在五只(每组总数的三分之一),以减少时间变量的影响,例如暴露于人类气味,腔室和觅食表面之间的温度变化等。
6. 表型评价:唾液腺形态及相关蛋白的下调
- 按照步骤4.2所述,在1x磷酸盐缓冲盐水(PBS)中分离新鲜组织,并在冰冷的丙酮中固定90秒。除去丙酮后,在1x PBS中冲洗几次。与一抗抗体在4°C下孵育过夜,抗血清(参见 材料表)稀释成1x PBS。
注:参见用于鉴定唾液蛋白一抗的材料 表 (按蚊 抗血小板蛋白,AAPP;粘蛋白2,MUC2),SG转录因子(叉头,fkh;鼠尾草,鼠尾草;环AMP响应元件结合蛋白A,CrebA)和分泌囊泡的标志物(Rab11)。这些抗体被用作SG形式和功能的读数。然而,任何适合免疫荧光的抗体都应该适合于该方案。 - 在1x PBS中洗涤几次。加入在1x PBS中稀释的二抗(荧光),并在室温下在黑暗中孵育2小时。加入任何复染[如4',6-二脒-2-苯基吲哚(DAPI;DNA),小麦胚芽凝集素(WGA;用于甲壳素),鬼球蛋白(用于F-肌动蛋白)和/或尼罗河红(用于脂质)]在2小时孵育结束前30分钟。
- 在1x PBS中洗涤三次。然后,将组织安装在具有1mm厚盖玻片的标准显微镜载玻片上的100%甘油中,并储存在-20°C,直到使用荧光共聚焦显微镜成像。
注意:要获得定量数据,成像设置必须保持恒定。在这里,仅包括通过整个3D体积组织的最大强度投影图像,并且所有图像定量在治疗之间(在实验中)基于非SG组织残留物(脂肪体,角质层或头部)中的DAPI信号进行归一化。
结果
首先,来自VectorBase的微阵列表达数据用于扫描发育阶段36,37 的潜在靶标,以确定与当前研究相关的所有基因的表达状态(表1)。正如预期的那样,我们选择的所有靶基因在成年SG中均表现出表达, aapp 和 鼠尾草 的水平特别高(表1)。同样值得注意的是, f-Agdsx 在成年女性SG9中的表达水平很高。
使用网络应用程序E-RNAi评估来自每个基因的特定片段以用作dsRNA ,以设计RNAi试剂30。 然后克隆含有每个靶基因特有序列的约400 bp区域(图1A),转化为适当的细菌菌株,并用于制备热杀灭细菌的悬浮液,诱导其产生dsRNA。成年蚊子在蔗糖浸泡的棉球上喂食8天,该棉球含有 用于f-Agdsx,fkh或 蚂蚁 (不相关的阴性对照)的dsRNA细菌悬浮液。
对于雌性蚊子的RNAi喂养的分析,首先确定f-Agdsx或fkh dsRNA喂养是否诱导基因沉默。在用fkh-dsRNA喂养的组中观察到fkh转录本水平降低98.8%(±2.1)(图1B),表明dsRNA非常有效地降低了SGs中fkh转录本的丰度。令人惊讶的是,在用dsRNA处理的f-Agdsx的蚊子中,fkh mRNA水平降低了82.0%(±18.9),其中f-Agdsx的减少率为89.86%(±4.48), 这表明fkh可能是唾液腺中F-Dsx的目标。随着fkh表达水平的显着降低,fkh敲低蚊子表现出血液喂养所需的探测尝试次数显着增加。平均而言,这些蚊子表现出比对照组或f-Agdsx dsRNA喂养的蚊子完全充血的五倍(图1C)。这导致询问fkh敲低RNAi治疗是否导致关键转录调节因子(SG TFs Sage和CrebA)(图2),分泌蛋白(AAPP和粘蛋白)和分泌机制[Nile Red(脂质)和Rab11(分泌囊泡)]的定位和/或分布发生变化(图4)。重要的是,在不同的叶区域,叶和单个SG中观察到染色强度的显着差异。
正如预测的那样,与蚂蚁对照RNAi相比,fkh RNAi之后的所有SG裂片的鼠尾草和CrebA染色水平显着降低(图2A)。线扫描图谱中最高最大强度值(红色虚线和数字标签)和最低最大强度值(蓝色虚线和数字标签)的减少表明组织内高信号和低信号区域的减少(图2A,B)。这些数据表明,An. gambiae fkh RNAi是有效的,并且fkh调节冈比亚猪笼草和CrebA中SG TFs Sage和CrebA的产生和/或稳定性,类似于它们在果蝇SGs19,38,39中的遗传关系。
当考虑高度丰富的唾液成分蛋白时,与对照RNAi治疗相比,fkh RNAi后所有三个SG叶中按蚊抗血小板蛋白(AAPP)40,41的水平降低(图3A,B;绿色)。另一方面,没有观察到粘蛋白水平的变化(图3A,B;紫色)。这些数据表明,Fkh对不同唾液蛋白基因的表达有不同的贡献。
最后,观察到两个分泌标志物(图4A,B):Rab11(与顶端再生内体相关的囊泡)42和尼罗河红(脂质)。在fkh RNAi处理后的远端侧叶(DL)叶中观察到Rab11荧光降低(图4A v vs. 4B v;绿色)。然而,内侧(M)和近端侧叶(PL)中的Rab11信号增加(图4A vii,ix与4B vii,ix;绿色)也发生。与对照RNAi治疗相比,fkh RNAi后在尼罗河红信号(图4A,B;紫色)中未观察到明显的差异。这些数据表明,fkh减少可能以复杂的方式改变一些分泌机制的作用,这在SG叶之间有所不同。
| 数据: | 戈尔采夫 | 内拉·奥维耶多 | 内拉·奥维耶多 | 面包师傅 | 面包师傅 | 面包师傅 | 面包师傅 | ||
| 基因符号 | 功能 | 阿加普编号 | 胚胎(25小时) | L3幼虫 | L3 升 | 成年女性 身体(3天) | 成年男性 身体(3天) | 成年女性 SG (3 天) | 成年男性 SG (3 天) |
| 断续器 | 唾液蛋白 | AGAP009974 | 3.92 | 4.38 | 4.33 | 3.81 | 2.46 | 11.92 | 2.69 |
| 克雷巴 | txn 因子 | AGAP001464 | 6.28 | 5.22 | 5.92 | 2.99 | 2.96 | 3.27 | 3.13 |
| " | txn 因子 | AGAP011038 | 4.50 | 4.46 | 5.23 | 2.96 | 2.86 | 3.05 | 2.88 |
| 断续器 | txn 因子 | AGAP004050 | 4.91 | 5.39 | 5.55 | 3.72 | 4.00 | 4.57 | 4.01 |
| 断续器 | txn 因子 | AGAP001671 | 5.18 | 4.67 | 5.25 | 2.99 | 3.09 | 3.21 | 3.05 |
| 断续器 | 唾液蛋白 | AGAP012020 | 4.59 | 5.53 | 5.63 | 2.96 | 3.07 | 3.08 | 3.26 |
| 拉布11 | 水疱贩运 | AGAP004559 | 10.21 | 7.47 | 8.60 | 4.90 | 3.79 | 3.38 | 2.96 |
| 圣人 | txn 因子 | AGAP013335 | 5.32 | 5.96 | 8.89 | 3.40 | 3.33 | 7.37 | 7.23 |
表 1:An 的平均 log2 微阵列表达谱 。 甘比亚 感兴趣的基因。 图中显示了从Vectorbase收集的基因名称、功能类别、Vectorbase(AGAP)标识符和平均log2微阵列表达数据。这些数据表明,与整个个体相比,我们感兴趣的基因(参与唾液腺(SG)细胞生物学和分泌)在幼虫3期(L3)和成体SG中表达和富集。

图1:成年冈比亚猪笼草的f-Agdsx和fkh敲低降低了SG中的fkh mRNA水平并影响女性的血液进食能力。 (A)该方法中用于dsRNA生产的质粒设计的代表性图像。通过将第二个T7启动子序列包含在用于扩增待克隆到pGEMT质粒中的插入片段的3'引物中,将其添加到质粒中。然后将质粒转化为大肠杆菌HT115(DE3)细菌,并在10%糖水中由诱导热杀灭细菌的悬浮液制成进料溶液。(B)用dsRNA喂养溶液喂养f-Agdsx或fkh的动物,fkh转录本水平显着降低(具有多重比较的单因子方差分析;n = 15)。然而,只有用fkh dsRNA(C)喂养的组在获取血粉所需的咬人尝试次数上显示出显着差异。该组中的蚊子平均需要五倍于对照组或dsx-dsRNA喂养组所需的探测尝试次数的五倍(具有多重比较的单因子方差分析;n = 15)。误差线表示均值的标准误差 (SEM)。每个实验在三个单独的生物重复中进行。请点击此处查看此图的大图。
{kind=link}

图2:成年冈比亚菌唾液腺中的fkh敲低降低了SG转录因子水平。 显示的是第13天成年雌性An. gambiae SGs在口服8天(第5-13天)后暴露于(A)非相关dsRNA对照(蚂蚁)或(B)靶向SG TF叉头(fkh,AGAP001671)的dsRNA在10%蔗糖中染色的染料DAPI(DNA;红色),标记小麦胚芽凝集素(WGA,甲壳素/ O-GlcNAcylation;蓝色), 反对SG TFs Sage(绿色)和CrebA(紫色)的反血清。显示的比例尺长度为微米。SG (i) 用白色破折号勾勒轮廓。放大的波瓣图像中的黄线(由黄色框包围的区域,并标记为"插图")表示进行信号强度线扫描的位置。在图像下方的图形中绘制对应于每个缩放波瓣的线扫描的绿色和紫色通道强度(在SG中始终从左到右);X 轴 = 距离(以像素为单位),Y 轴 = 灰度单位(像素强度)。像素强度的动态范围由红色(最大值)和蓝色(最小值)虚线分隔,相应的值显示在每个图形上。MIP = 通过整个 SG 深度的最大强度 3D 投影。DL:远端外侧叶;M:内叶;PL:近端外叶;SD:唾液管。请点击此处查看此图的大图。
{kind=link}

图3:成虫冈比亚菌唾液腺中的fkh敲低降低了SG分泌的蛋白质水平。 显示的是第13天成年雌性An. gambiae SGs在口服8天(第5-13天)后暴露于(A)非相关dsRNA对照(蚂蚁),或(B)靶向SG TF叉头(fkh,AGAP001671)的dsRNA在10%蔗糖中用染料DAPI染色(DNA;红色),标记小麦胚芽凝集素(WGA,甲壳素/ O-GlcNAcylation;蓝色), 和唾液蛋白AAPP(绿色)和粘蛋白(MUC2,紫色)。显示的比例尺长度为微米。SG (i) 用白色破折号勾勒轮廓。放大的波瓣图像中的黄线(由黄色框包围的区域)表示信号强度的线扫描的位置。对应于每个波瓣的线扫描的绿色和紫色通道强度在图像下方的图形中绘制(在SG中始终从左到右);X 轴 = 距离(以像素为单位),Y 轴 = 灰度单位(像素强度)。像素强度的动态范围由红色(最大值)和蓝色(最小值)虚线分隔,相应的值显示在每个图形上。MIP = 通过整个 SG 深度的最大强度 3D 投影。DL:远端外侧叶;M:内叶;PL:近端外叶;SD:唾液管。斜体"DL"标签 (Bi) 表示同一 DL 波瓣的两个可见区域。请点击此处查看此图的大图。
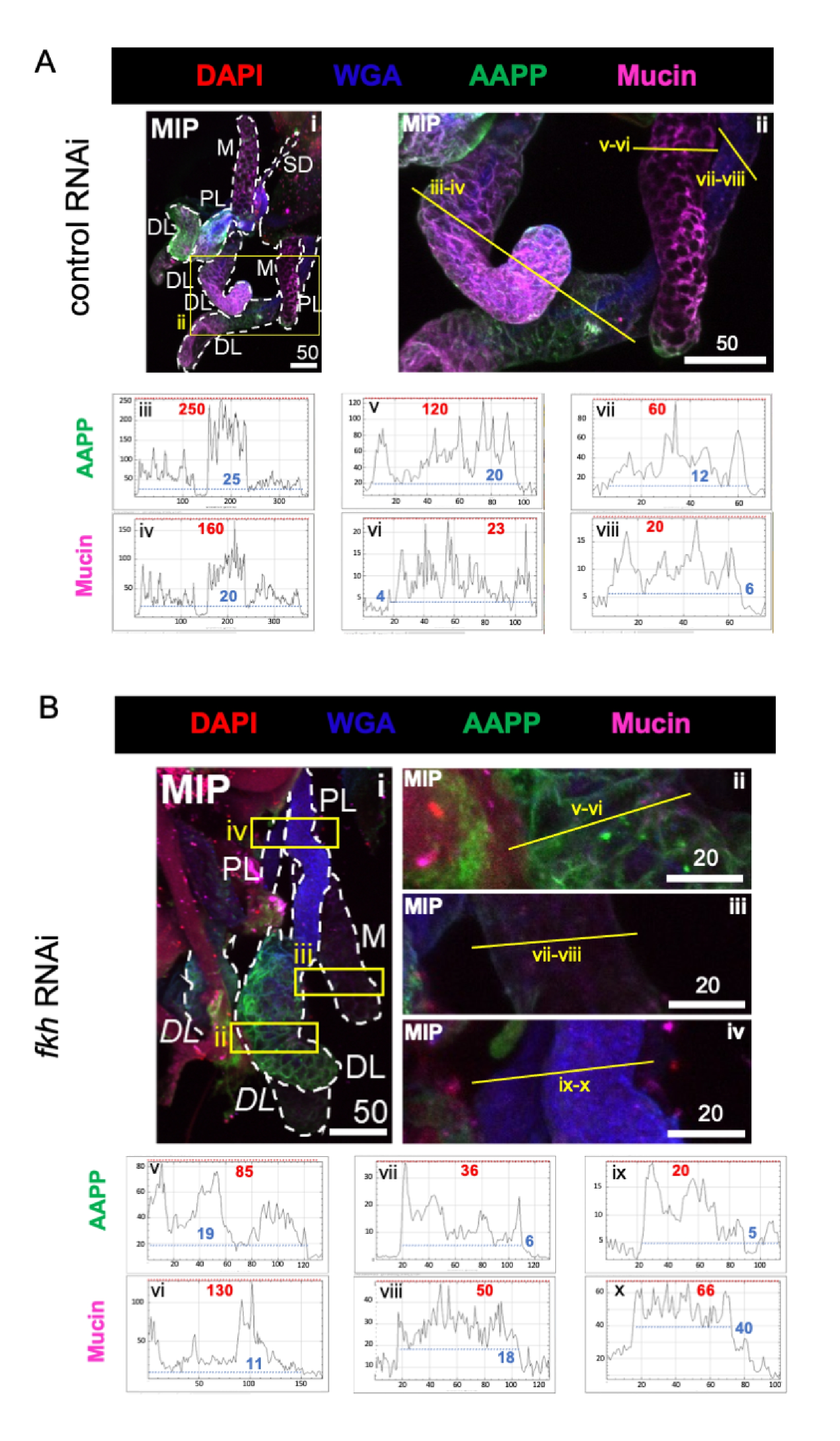{kind=link}

图4:成年冈比亚菌唾液腺中的fkh敲低可减少SG分泌标志物。 显示的是第13天成年雌性An. gambiae SGs在口服8天(第5-13天)后暴露于(A)非相关dsRNA对照(蚂蚁),或(B)靶向SG TF叉头(fkh,AGAP001671)的dsRNA在10%蔗糖中用染料DAPI染色(DNA;红色),标记小麦胚芽凝集素(WGA,甲壳素/ O-GlcNAcylation;蓝色), 尼罗河红(脂质;紫色)和抗血清对抗回收的内体囊泡标记Rab11(绿色)。显示的比例尺长度为微米。SG (i) 用白色破折号勾勒轮廓。放大的波瓣图像中的黄线(由黄色框包围的区域)表示信号强度的线扫描的位置。在图像下方的图形中绘制对应于每个波瓣的线扫描的绿色和紫色通道强度(在SG中始终从左到右);X 轴 = 距离(以像素为单位),Y 轴 = 灰度单位(像素强度)。像素强度的动态范围由红色(最大值)和蓝色(最小值)虚线分隔,相应的值显示在每个图形上。MIP = 通过整个 SG 深度的最大强度 3D 投影。DL:远端外侧叶;M:内叶;PL:近端外叶;SD:唾液管。请点击此处查看此图的大图。
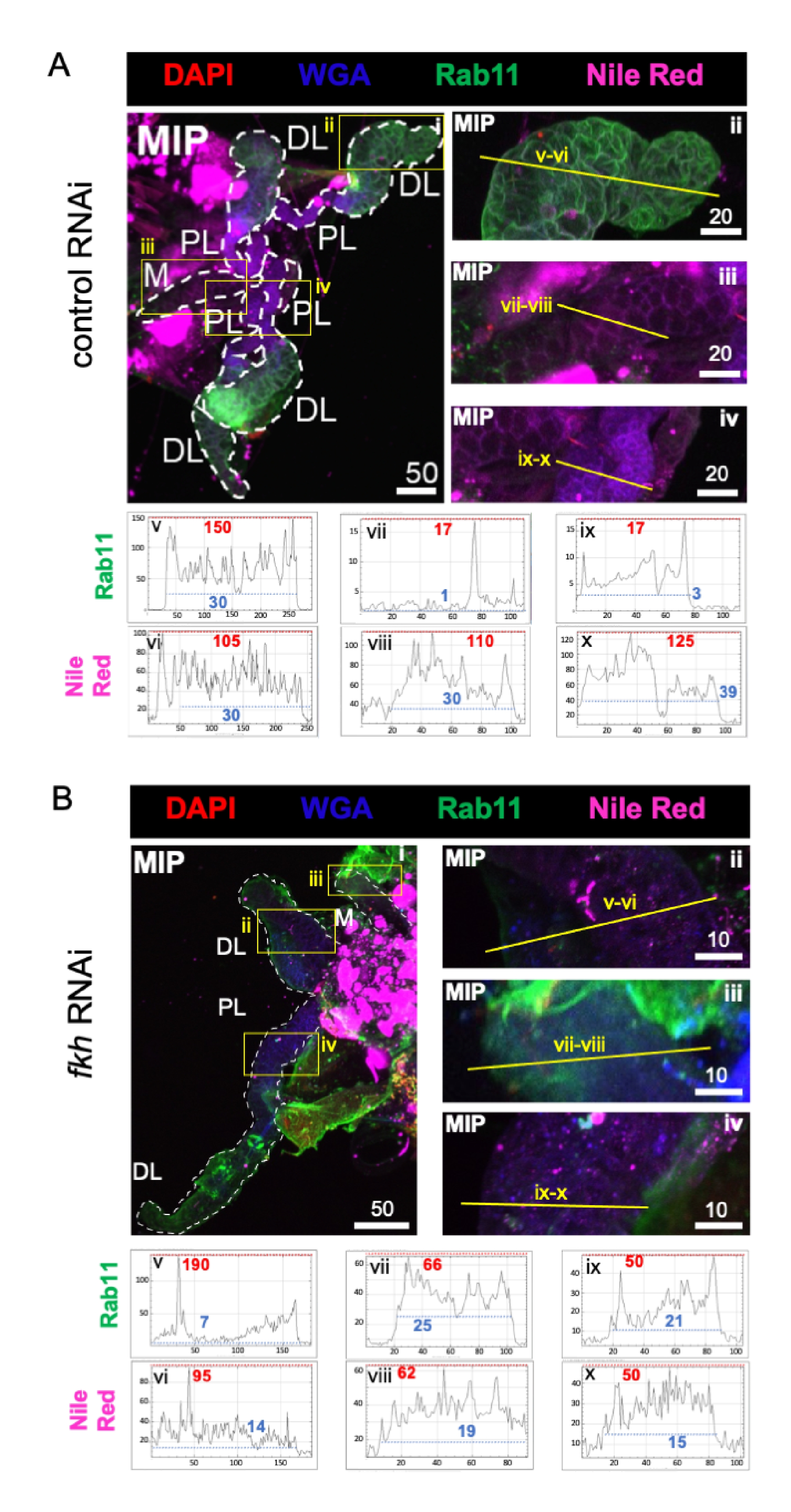{kind=link}
补充文件 1.请点击此处下载此文件。
讨论
通过经口喂养将dsRNA有效地传递给 冈比亚 蚊的能力对实验室和现场的载体生物学研究具有广泛的意义。显微注射长期以来一直被接受为蚊子中化学物质,抗体,RNAi和遗传修饰策略的首选递送方式43,44。通过使用口服给药可以避免大量物理操作,细胞损伤和压力的后果,这也可能适用于大规模或现场应用。先前的研究表明,RNAi在个体成年蚊子29中无处不在,允许在所有组织中起作用,包括唾液腺。通过给蚊子喂食大量表达dsRNA 的大肠杆菌,这些大肠杆菌 在很长一段时间内异步消化,人们可以潜在地在笼子中的所有个体中实现对RNAi的一致和均匀的暴露。这种方法允许喂养大量的蚊子,并根据靶基因分析所得表型的潜在变异性。然而,一个重要的考虑因素是细菌在棉纤维中异质分布的可能性,因此dsRNA。每天用于蚊子糖喂养的400μL细菌将含有约≤4.6μg的dsRNA,如前所述和计算了9 ,但每只蚊子摄入的dsRNA量没有单独确定。如果构建dsRNA构建体成为常规,这种简单的治疗方案允许任何蚊子研究人员快速同化该技术。 先验地,与学习和应用显微注射到相似样本大小所需的时间相比,治疗期间的时间消耗(每天30分钟)是微不足道的。
喂养dsRNA通常用于模型生物秀丽隐杆线虫45的反向遗传学研究。这种大量使用强调了口服给药方法的价值。在转化的大肠杆菌中构建An. gambiae全基因组文库,类似于秀丽隐杆线虫46,47中存在的文库,将允许在蚊子中以更大的规模进行快速反向遗传筛查。然而,重要的是要注意,该方法的效率在很大程度上取决于转录本的内源性水平,并且如果表达不限于靶组织,而是更广泛地表达4,8,44。此外,有证据表明,一些杀虫剂可以诱发蚊子的行为回避48,并且用可能诱发对蚊子产生不利影响的细菌喂养可能会引发类似的避免模式。在实验室的受控环境中,蚊子没有其他食物来源,他们别无选择,无法避免含有大肠杆菌的糖水,对营养来源的需求可能会压倒避免细菌的本能。但是,如果策略旨在用于控制较少的设置,则应考虑这一点。
有可能同时靶向多个基因(使用一个构建体,多个构建体或转化的细菌分离物的混合物),但需要进一步的研究来评估有效性。这一点的另一个重要考虑因素是评估使用单个或多个目标时可能的脱靶或协同效应。建立适当的对照基因和基团是实验设计的重要组成部分。此外,人们很容易推测这种方法可用于靶向其它病原体或病毒49。以前在蚊子中诱导RNAi的工作是在直接注射试剂的条件下进行的,因此不存在 大肠杆菌 。大 肠杆菌 可以提供一个保护室,允许随着时间的推移更慢地释放dsRNA,确保暴露在更长的时间内或多或少是连续的29。
最后,这些结果表明,通过调整暴露的时间范围(长度和开始日)和大 肠杆菌 的使用量,可以调节该技术的效果。这一特征使我们能够通过反复试验确定最佳敲低条件,从而研究必需基因(dsx 和 fkh)的功能。这大大增加了使用这种技术研究目标基因的可能性。
总之,研究发现,口服RNAi给成年蚊子可以简单,通用,并且是研究蚊子基因功能和创建用于病媒控制的新型和可塑性工具的有力方法。
披露声明
作者报告说,他们没有利益冲突需要披露。
致谢
作者希望感谢疾病预防控制中心昆虫学处和寄生虫病和疟疾司的工作人员和科学家,以及Brian Trigg和Michelle Chiu在JHU的细菌制备方面的帮助和/或对这项工作的有益讨论。我们感谢JHMRI Insectary和经理Chris Kizito获得并饲养 了冈比亚 蚊子。我们感谢黄伟(JHSPH)协助获得质粒PJet GFP和pPB47 GFP用于本研究。这项工作的资金由以下机构提供:NIH R21AI153588(DJA),约翰霍普金斯疟疾研究所博士后奖学金(MW);由Good Ventures基金会和开放慈善项目向CDC基金会提供的赠款,题为"支持蚊子中女性发育的冷冻保存和抑制,以协助疟疾研究,开放慈善项目",2017年。我们非常感谢JHU显微镜设施工作人员的协助以及适用的NIH对所用显微镜的资助支持(NIH拨款编号:S10OD016374)。本手稿中的发现和结论是作者的,并不一定代表CDC的观点。商号仅用于识别,并不意味着得到疾病控制和预防中心、公共卫生服务局或美国卫生与公众服务部的认可。
材料
| Name | Company | Catalog Number | Comments |
| 1 Kb Plus DNA Ladder | Thermo Fisher Scientific | 10787018 | |
| 2x Yeast Extract Tryptone (2xYT) Medium | BD Difco | DF0440-17 | |
| AAPP | n/a | n/a | Antisera. 1:50 dilution (rabbit); gift from Fabrizio Lombardo |
| AccuStart II PCR Supermix | Quantabio | 95137-100 | |
| Agarose | Millipore Sigma | A9539 | |
| Ampicillin | Millipore Sigma | A5354 | |
| Anopheles gambiae G3 | BioDefense and Emerging Infections (BEI) Malaria Research and Reference Reagent Resource Center (MR4) | MRA-112 | |
| BugDorm | BioQuip | 1452 | |
| Centrifuge 5810R | Eppendorf | P022628181 | |
| CrebA | DSHB | CrebA Rbt-PC | Antisera. 1:50 dilution (rabbit); generated by the Andrew Lab |
| Damiens diet | BioServ | ||
| DAPI | Life Technologies | n/a | 4′,6-diamidino-2-phenylindole; 1:200 dilution. |
| Defibrinated sheep blood | HemoStat | DSB050 | |
| Escherichia coli HT115 (DE3) | |||
| Ethidium bromide | Millipore Sigma | E7637 | |
| High-Capacity cDNA Reverse Transcription Kit | Thermo Fisher Scientific | 4368814 | |
| Isopropyl β-D-1-thiogalactopyranoside | Millipore Sigma | I5502 | |
| JM109 Competent cells | Promega | L2005 | |
| Luria Broth Media | Thermo Fisher Scientific | 10855001 | |
| Mucin 2 | Proteintech | Muc2; 27 675-1-AP | Antisera. 1:100 dilution (mouse). |
| Nanodrop 2000 | Thermo Fisher Scientific | ||
| Nile Red | Sigma | n/a | Lipid dye; 1:50 dilution. |
| Owl EasyCast B2 Mini Gel Horizontal Electrophoresis | Thermo Fisher Scientific | Model B2 | |
| pGEMT easy | Promega | A3600 | |
| Power SYBR-green PCR master MIX | Applied Biosystems | 4367659 | |
| PureLink PCR purification kit | Thermo Fisher Scientific | K31001 | |
| QuantaStudio 6 | Applied Biosystems | ||
| QuantStudio6 Real Time PCR System | Applied Biosystems | ||
| Rab11 | n/a | n/a | Antisera. 1:100 dilution (rabbit); generated by the Andrew Lab |
| Rh-WGA | Vector Labs | n/a | Rhodamine-conjugated wheat germ agglutinin (chitin, O-GlcNAcylation dye); 1:40 dilution |
| Sage | n/a | n/a | Antisera. 1:50 dilution (rat); generated by the Andrew Lab |
| T4 DNA ligase | Promega | M1801 | |
| Tetracycline | Millipore Sigma | 87128 | |
| Trizol | Thermo Fisher Scientific | 15596018 | |
| Zeiss LSM700 fluorescence confocal microscope | Zeiss | ||
| ANTIBODIES | |||
| Chicken anti-Rat IgG (H+L), Alexa Fluor 647 | Thermo Fisher Scientific | A21472 | |
| Goat anti-Mouse IgG (H+L), Alexa Fluor 647 | Thermo Fisher Scientific | A28181 | |
| IgG (H+L) Goat anti-Rabbit, Alexa Fluor 488 | Thermo Fisher Scientific | A27034 | |
| Rabbit anti-Goat IgG (H+L), Alexa Fluor 488 | Thermo Fisher Scientific | A27012 | |
| PRIMERS | |||
| ACT-2f: TACAACTCGATCATGAAGTGCGA | CDC Biotechnology Core Facility Branch | n/a | qRT-PCR primer |
| ACT-3r: CCCGGGTACATGGTGGTACCGC CGGA | CDC Biotechnology Core Facility Branch | n/a | qRT-PCR primer |
| FKH_RNAi_F: GCCGACTTATGCTTAGCCCA | CDC Biotechnology Core Facility Branch | n/a | qRT-PCR primer |
| FKH_RNAi_R: TAGCCGTCAATTCCTCCTGC | CDC Biotechnology Core Facility Branch | n/a | qRT-PCR primer |
| newDSX-f: AGAGGGCGGGGAAATTCTAGT | CDC Biotechnology Core Facility Branch | n/a | qRT-PCR primer |
| newDSX-r: GGGCTTGTGGCAGTACGAATA | CDC Biotechnology Core Facility Branch | n/a | qRT-PCR primer |
| S7qf1: AGAACCAGCAGACCACCATC | CDC Biotechnology Core Facility Branch | n/a | qRT-PCR primer |
| S7qr1: GCTGCAAACTTCGGCTATTC | CDC Biotechnology Core Facility Branch | n/a | qRT-PCR primer |
参考文献
- Hoa, N. T., Keene, K. M., Olson, K. E., Zheng, L. Characterization of RNA interference in an Anopheles gambiae cell line. Insect Biochemistry and Molecular Biology. 33, 949-957 (2003).
- Caplen, N., Zheng, Z., Falgout, B., Morgan, R. Inhibition of viral gene expression and replication in mosquito cells by dsRNA-triggered RNA interference | Elsevier enhanced reader. Molecular Therapy. 6, 243-251 (2002).
- Brown, A. E., Catteruccia, F. Toward silencing the burden of malaria: progress and prospects for RNAi-based approaches. BioTechniques. , 38-44 (2006).
- Airs, P. M., Bartholomay, L. C. RNA interference for mosquito and mosquito-borne disease control. Insects. 8, (2017).
- Blandin, S., et al. Reverse genetics in the mosquito Anopheles gambiae: targeted disruption of the Defensin gene. EMBO Reports. 3 (9), 852-856 (2002).
- Garver, L., Dimopoulos, G. Protocol for RNAi assays in adult mosquitoes (A. gambiae). Journal of Visualized Experiments: JoVE. (5), e230 (2007).
- Whyard, S., et al. Silencing the buzz: a new approach to population suppression of mosquitoes by feeding larvae double-stranded RNAs. Parasites & Vectors. 8, 96 (2015).
- Wiltshire, R. M., Duman-Scheel, M. Advances in oral RNAi for disease vector mosquito research and control. Current Opinion in Insect Science. 40, 18-23 (2020).
- Taracena, M. L., Hunt, C. M., Benedict, M. Q., Pennington, P. M., Dotson, E. M. Downregulation of female doublesex expression by oral-mediated RNA interference reduces number and fitness of Anopheles gambiae adult females. Parasites & Vectors. 12, 170 (2019).
- Grassi, B. Studi di uno zoologo sulla malaria. Real Accademia dei Lincei. 3, 229 (1901).
- Smith, R. C., Jacobs-lorena, M. Plasmodium - Mosquito interactions: A tale of roadblocks and detours. Advances in Insect Physiology. 39, (2010).
- Das, S., et al. Transcriptomic and functional analysis of the Anopheles gambiae salivary gland in relation to blood feeding. BMC Genomics. 11, 1-14 (2010).
- Francischetti, I. M. B., Valenzuela, J. G., Pham, V. M., Garfield, M. K., Ribeiro, J. M. C. Toward a catalog for the transcripts and proteins (sialome) from the salivary gland of the malaria vector Anopheles gambiae. Journal of Experimental Biology. 205, 2429-2451 (2002).
- Henderson, K. D., Isaac, D. D., Andrew, D. J. Cell fate specification in thedrosophila salivary gland: The integration of homeotic gene function with the DPP signaling cascade. Developmental Biology. 205, 10-21 (1999).
- Mach, V., Ohno, K., Kokubo, H., Suzuki, Y. The Drosophila fork head factor directly controls larval salivary gland-specific expression of the glue protein gene Sgs3. Nucleic Acids Research. 24 (12), 2387-2394 (1996).
- Weiserova, M., et al. Mini-Mu transposition of bacterial genes on the transmissible plasmid. Folia Microbiologica. 32 (5), 368-375 (1987).
- Abrams, E. W., Mihoulides, W. K., Andrew, D. J. Fork head and Sage maintain a uniform and patent salivary gland lumen through regulation of two downstream target genes, PH4αSG1 and PH4αSG2. Development. 133, 3517-3527 (2006).
- Myat, M. M., Isaac, P. P., Andrew, D. J. Early genes required for salivary gland fate determination and morphogenesis in Drosophila melanogaster. Advances in Dental Research. 14, 89-98 (2000).
- Fox, R. M., Vaishnavi, A., Maruyama, R., Andrew, D. J. Organ-specific gene expression: the bHLH protein Sage provides tissue specificity to Drosophila FoxA. Development of Cell Biology. 140, 2160-2171 (2013).
- Maruyama, R., Grevengoed, E., Stempniewicz, P., Andrew, D. J. Genome-wide analysis reveals a major role in cell fate maintenance and an unexpected role in endoreduplication for the Drosophila FoxA gene fork head. PLOS ONE. 6, 20901 (2011).
- Johnson, D. M., et al. CrebA increases secretory capacity through direct transcriptional regulation of the secretory machinery, a subset of secretory cargo, and other key regulators. Traffic. 21, 560-577 (2020).
- Fox, R. M., Hanlon, C. D., Andrew, D. J. The CrebA/Creb3-like transcription factors are major and direct regulators of secretory capacity. Journal of Cell Biology. 191, 479-492 (2010).
- Abrams, E. W., Andrew, D. J. CrebA regulates secretory activity in the Drosophila salivary gland and epidermis. Development. 132, 2743-2758 (2005).
- Wells, M. B., Andrew, D. J. Anopheles salivary gland architecture shapes plasmodium sporozoite availability for transmission. mBio. 10 (4), 01238 (2019).
- Pei-Wen, L., Xiao-Cong, L., Jin-Bao, G., Yan, L., Xiao-Guang, C. Molecular cloning, characterization and expression analysis of sex determiantion gene doublesex from Anopheles gambiae (Diptera: Culicidae). Acta Entomologica Sinica. 58 (2), 122-131 (2015).
- Scali, C., Catteruccia, F., Li, Q., Crisanti, A. Identification of sex-specific transcripts of the Anopheles gambiae doublesex gene. The Journal of Experimental Biology. 208, 3701-3709 (2005).
- Price, D. C., Egizi, A., Fonseca, D. M. Characterization of the doublesex gene within the Culex pipiens complex suggests regulatory plasticity at the base of the mosquito sex determination cascade. BMC Evolutionary Biology. 15, 1-13 (2015).
- Mysore, K., et al. siRNA-mediated silencing of doublesex during female development of the dengue vector mosquito Aedes aegypti. PLoS Neglected Tropical Diseases. 9, 1-21 (2015).
- Boisson, B., et al. Gene silencing in mosquito salivary glands by RNAi. FEBS Letters. 580, 1988-1992 (2006).
- Horn, T., Boutros, M. E-RNAi: a web application for the multi-species design of RNAi reagents-2010 update. Nucleic Acids Research. 38, 332-339 (2010).
- Taracena, M. L., et al. Genetically modifying the insect gut microbiota to control chagas disease vectors through systemic RNAi. PLoS Neglected Tropical Diseases. 9, (2015).
- Sambrook, J., Fritsch, E. F., Maniatis, T. . Molecular Cloning: A Laboratory Manual. , (1989).
- Ullmann, A., Jacob, F., Monod, J. Characterization by in vitro complementation of a peptide corresponding to an operator-proximal segment of the β-galactosidase structural gene of Escherichia coli. Journal of Molecular Biology. 24, 339-343 (1967).
- Timmons, L. Bacteria-mediated RNAi-General outline. Carnegie Institution of Washington. , (2000).
- Pfaffl, M. W. Relative quantification. Real-time PCR. , 63-82 (2004).
- Neira-Oviedo, M., et al. The RNA-Seq approach to studying the expression of mosquito mitochondrial genes. Insect Molecular Biology. 20, 141-152 (2011).
- Baker, D. A., et al. A comprehensive gene expression atlas of sex- and tissue-specificity in the malaria vector, Anopheles gambiae. BMC Genomics. 12, (2011).
- Loganathan, R., Hoon, J., Wells, M. B., Andrew, D. J. Secrets of secretion - How studies of the Drosophila salivary gland have informed our understanding of the cellular networks underlying secretory organ form and function. Cellular Networks in Development. , 143 (2021).
- Chung, S., Hanlon, C. D., Andrew, D. J. Building and specializing epithelial tubular organs: The Drosophila salivary gland as a model system for revealing how epithelial organs are specified, form and specialize. Wiley Interdisciplinary Reviews: Developmental Biology. 3, 281-300 (2014).
- Yoshida, S., et al. Inhibition of collagen-induced platelet aggregation by anopheline antiplatelet protein, a saliva protein from a malaria vector mosquito. Blood. 111, 2007-2014 (2008).
- Wells, M. B., Villamor, J., Andrew, D. J. Salivary gland maturation and duct formation in the African malaria mosquito Anopheles gambiae. Scientific Reports. 7 (1), 601 (2017).
- Takahashi, S., et al. Rab11 regulates exocytosis of recycling vesicles at the plasma membrane. Journal of Cell Science. 125, 4049-4057 (2012).
- Catteruccia, F., Levashina, E. A. RNAi in the malaria vector, Anopheles gambiae. Methods in Molecular Biology. 555, 63-75 (2009).
- Balakrishna Pillai, A., et al. RNA interference in mosquito: understanding immune responses, double-stranded RNA delivery systems and potential applications in vector control. Insect Molecular Biology. 26, 127-139 (2017).
- Fire, A., et al. Potent and specific genetic interference by double-stranded RNA in caenorhabditis elegans. Nature. 391, 806-811 (1998).
- Kamath, R. S., Ahringer, J. Genome-wide RNAi screening in Caenorhabditis elegans. Methods. 30, 313-321 (2003).
- Kamath, R. S., et al. Systematic functional analysis of the Caenorhabditis elegans genome using RNAi. Nature. 421, 231-237 (2003).
- Carrasco, D., et al. Behavioural adaptations of mosquito vectors to insecticide control. Current Opinion in Insect Science. 34, 48-54 (2019).
- Magalhaes, T., et al. Induction of RNA interference to block Zika virus replication and transmission in the mosquito Aedes aegypti. Insect Biochemistry and Molecular Biology. , 111 (2019).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。