
Method Article
Gen-Editierung von primären Rhesusaffen-B-Zellen
In diesem Artikel
Zusammenfassung
Wir stellen eine Methode zur Kultivierung und Gen-Editierung von primären Rhesusaffen-B-Zellen unter Verwendung von CRISPR/Cas9 und rekombinantem Adeno-assoziiertem Virus-Serotyp 6 für die Untersuchung von B-Zell-Therapien vor.
Zusammenfassung
B-Zellen und ihre Nachkommen sind die Quellen hochexprimierter Antikörper. Ihre hohen Proteinexpressionsfähigkeiten zusammen mit ihrer Häufigkeit, der leichten Zugänglichkeit über peripheres Blut und der Zugänglichkeit für einfache adoptive Transfers haben sie zu einem attraktiven Ziel für Gen-Editing-Ansätze gemacht, um rekombinante Antikörper oder andere therapeutische Proteine zu exprimieren. Die Gen-Editierung von primären B-Zellen der Maus und des Menschen ist effizient, und Mausmodelle für In-vivo-Studien haben sich als vielversprechend erwiesen, aber die Machbarkeit und Skalierbarkeit für größere Tiermodelle wurde bisher nicht nachgewiesen. Wir haben daher ein Protokoll entwickelt, um primäre B-Zellen von Rhesusaffen in vitro zu editieren, um solche Studien zu ermöglichen. Wir berichten über Bedingungen für die In-vitro-Kultur und Gen-Editierung von primären Rhesusaffen-B-Zellen aus mononukleären Zellen oder Splenozyten des peripheren Blutes mit CRISPR/Cas9. Um die gezielte Integration von großen (<4,5 kb) Kassetten zu erreichen, wurde ein schnelles und effizientes Protokoll zur Herstellung des rekombinanten Adeno-assoziierten Virus-Serotyps 6 als homologiegesteuerte Reparaturvorlage unter Verwendung eines Tetracyclin-fähigen, selbststummschaltenden adenoviralen Helfervektors aufgenommen. Diese Protokolle ermöglichen die Untersuchung prospektiver B-Zell-Therapeutika bei Rhesusaffen.
Einleitung
B-Zellen sind die Grundlage der humoralen Immunität. Bei Aktivierung durch verwandte Antigen- und Sekundärsignale entstehen aus naiven B-Zellen des Keimzentrums B-Zellen, Gedächtnis-B-Zellen und Plasmazellen1. Letzteres ist die Quelle der sezernierten Antikörper, die die Schutzfunktionen der meisten derzeit verfügbaren Impfstoffe vermitteln2. Plasmazellen wurden als Antikörperfabriken beschrieben, da sie große Mengen an Antikörpern in das Serum sezernieren - etwa 2 ng / Tag / Zelle3, was 7-16 g / l Serum entspricht, was Antikörper zu einem der drei am häufigsten vorkommenden Proteine im Serum4 macht. B-Zellen sind reichlich im Blut vorhanden und können daher leicht gewonnen und in ein Individuum zurückgegossen werden.
Diese Merkmale haben B-Zellen zu einem Ziel von Zelltherapiebemühungen gemacht, um den B-Zell-Rezeptor (BCR) zu geneditieren und breit neutralisierende Antikörper (bNAbs) gegen das humane Immundefizienzvirus (HIV)5,6,7,8,9,10,11,12,13,14,15 und andere Proteine zu exprimieren 16, 17,18,19,20,21. Solche Ansätze haben in zahlreichen Mausstudien in vivoPotenzial gezeigt 7,8,10,11,16,22. Für die klinische Translation 9,15,23 müssen jedoch noch einige Hürden überwunden werden, darunter Sicherheit, Dauer und Ausmaß der therapeutischen Wirksamkeit sowie die Skalierung auf größere Tiere wie nichtmenschliche Primaten (NHPs). In der Tat sind NHPs und insbesondere Rhesusaffen, die eine lange Geschichte in der Antikörper- und HIV-Forschunghaben 24,25, das am besten geeignete Modell, um diese Parameter zu testen.
Hier haben wir Protokolle entwickelt, die es ermöglichen, diese Probleme anzugehen. Bisher haben nur wenige Studien versucht, Rhesusaffen-B-Zellen ex vivo zu kultivieren, und nur eine positive Selektion unter Verwendung von CD20 wurde für die Reinigung von Rhesusaffen-B-Zellenberichtet 26,27,28. Wir haben ein Protokoll für die Isolierung von unberührten Rhesusaffen-B-Zellen durch die negative Depletion anderer Zelltypen erstellt. Darüber hinaus werden Kultivierungsbedingungen für die gezielte Genom-Editierung von Rhesusaffen-B-Zellen definiert. Dieses Protokoll beschreibt die Verwendung von CRISPR/Cas9-Ribonukleoproteinen (RNPs) und rekombinantem Adeno-assoziiertem Virus-Serotyp 6 (rAAV6) als homologiegesteuerte Reparaturvorlage (HDRT) zur Gen-Editierung kultivierter Rhesusaffen-B-Zellen. Mit diesem Protokoll wurden Bearbeitungseffizienzen von bis zu 40 % bei großen (~1,5 kb) Einfügungen erreicht. Wir stellen auch eine schnelle und kostengünstige Methode zur Herstellung von rAAV6 unter Verwendung eines Tetracyclin-fähigen, selbststummschaltenden adenoviralen Helfers29 vor, um die schnelle Testung von HDRTs in diesem Format zu ermöglichen. Zusammengenommen beschreiben diese Protokolle einen effizienten Arbeitsablauf für die Genom-Editierung von Rhesusaffen-B-Zellen (Abbildung 1), der die Bewertung von B-Zell-Therapien in einem NHP-Modell ermöglicht.
Um die Experimente zu starten, kann Spendermaterial aus kommerziellen Quellen bestellt oder durch Phlebotomie oder Splenektomie gewonnen werden. In dieser Studie wurden die Phlebotomien und Blutentnahmen wie zuvor beschrieben30 mit dem Gerinnungshemmer EDTA durchgeführt. Um Milz-, primäre Rhesusaffen-B-Zellen zu erhalten, wurden partielle (25%-50%) oder totale Splenektomien unter Verwendung von Techniken durchgeführt, über die zuvor berichtet wurde31. Die Tiere wurden vor der Operation über Nacht gefastet. Kurz gesagt, während der Operation wurde der Bauch abgeschnitten und dreimal abwechselnd mit Chlorhexidin und 70% Isopropylalkohol präpariert. Ein Schnitt (5-10 cm) wurde im Bauch gemacht, um die Milz zu identifizieren und zu isolieren. Das Gefäßsystem der Milz wurde entweder mit Nähten oder Gefäßklammern ligiert. Der Schnitt wurde in zwei Schichten mit 4-0 PDS-Polydioxanon-Nähten verschlossen. Die Splenektomie wurde einmalig für ein einzelnes Tier durchgeführt. Einzellige Suspensionen wurden aus Makakenmilz durch Mazeration durch Zellsiebe hergestellt. Mononukleäre Zellen aus Blut und Milzzellsuspensionen wurden mittels Dichtegradientenzentrifugation hergestellt und in flüssigem Stickstoff gelagert.
Protokoll
Alle tierexperimentellen Verfahren und Experimente wurden gemäß Protokollen durchgeführt, die vom Institutional Animal Care and Use Committee des National Institute of Allergy and Infectious Diseases, National Institutes of Health, genehmigt wurden. Eine Zusammenfassung der folgenden Protokolle ist in Abbildung 1 dargestellt. Männliche und weibliche Rhesusaffen (Macaca mulatta) indischer Genetik im Alter von 2 bis 8 Jahren wurden gemäß den Richtlinien des Ausschusses für die Pflege und Verwendung von Versuchstieren in einer Einrichtung der Biosicherheitsstufe 2 untergebracht und gepflegt.
VORSICHT: Alle Experimente wurden in Übereinstimmung mit den universellen Vorsichtsmaßnahmen für durch Blut übertragbare Krankheitserreger, mit sterilen/aseptischen Techniken und geeigneter Ausrüstung der Biosicherheitsstufe 2 in Laminar-Flow-Hauben durchgeführt.
1. rAAV6-Produktion
- Bereiten Sie die Reagenzien für die rAAV6-Produktion vor.
- Entwerfen und klonen Sie die homologiegesteuerte Reparaturschablone zwischen den invertierten Terminalwiederholungen (ITRs) von AAV2 im Vektor pAAV unter Verwendung von Standardtechniken. Stellen Sie sicher, dass die Homologiearme auf beiden Seiten mindestens ~ 250 bp betragen, aber nur 60 bp ausreichen können, obwohl längere Homologiearme bevorzugt werden, wenn das Konstruktionsdesign dies zulässt. Wenn Zielsequenzen einer der verwendeten sgRNAs im HDRT vorhanden sind, entfernen Sie sie mit stillen Mutationen, die am effektivsten in dem an den Protospacer angrenzenden Motiv oder der Seed-Region der Zielstelle sind.
HINWEIS: Es kann eine Gensynthese in Kombination mit einer Gibson-Assemblierung für eine effiziente Klonierung32 durchgeführt werden. Bereiten Sie einen Maxiprep eines korrekten Klons für die Transfektion vor. Für das sgRNA-Design wird CHOPCHOP33 empfohlen, und eine Liste weiterer Tools finden Sie unter https://zlab.bio/guide-design-resources. Die maximale Verpackungskapazität für AAV einschließlich ITRs beträgt ~4,7 kb. AAV6 ist der am häufigsten verwendete Serotyp zur Bearbeitung hämatopoetischer Zellen, insbesondere B-Zellen9. Andere Serotypen von AAV für die Gen-Editierung von Rhesusaffen-B-Zellen wurden nicht getestet, aber AAV28 und AAV-DJ10,11 wurden in Mausstudien verwendet. - Bereiten Sie 293AAV-Nährmedium und Produktionsmedium gemäß Tabelle 1 und Tabelle 2 vor. Sterilfilter durch eine 0,2 μm Polyethersulfon (PES) Membranfiltereinheit. Bei 4 °C lagern.
- Bereiten Sie 1x Polyethylenimin (PEI) -Lösung (1 mg/ml, 100 ml) vor.
- In einem 250-ml-Glasbecher ~ 70 ml H2O in einer Mikrowelle für ~ 30 s erhitzen und dann 100 mg PEI hinzufügen. Fügen Sie einen Magnetrührer hinzu und rühren Sie, bis sich das PEI weitgehend aufgelöst hat.
- Stellen Sie den pH-Wert mit 1 M HCl auf 7 ein, füllen Sie dann mit H2O auf 100 ml auf, warten Sie 10 Minuten, überprüfen Sie den pH-Wert erneut und passen Sie ihn gegebenenfalls an.
- Sterilfiltrieren Sie die PEI-Lösung durch eine 0,2 μm PES-Membranfiltereinheit, aliquot, und lagern Sie sie bei −20 °C. Nach dem Auftauen kann die Lösung bei 4 °C bis zu 2 Monate gelagert werden.
- Bereiten Sie 5x Polyethylenglykol (PEG)/NaCl-Lösung vor.
- Wiegen Sie 400 g PEG 8.000 und 24 g NaCl ab.
- Geben Sie einen Magnetrührer in ein 2-Liter-Glasbecherglas, fügen Sie das abgewogene PEG 8.000 und NaCl hinzu und spülen Sie es mit ~ 550 ml deionisiertem Wasser ab.
- Unter Hitze umrühren und auf 80-90 °C zum Kochen bringen, bis es sich vollständig aufgelöst hat.
- Stellen Sie den pH-Wert mit 1 M NaOH auf ~7,4 ein, stellen Sie dann das Volumen mit einem Messzylinder auf 1 l ein und füllen Sie es mit dem Magnetrührer in eine 2-l-Glasflasche.
- Autoklavieren Sie die Flasche, den Magnetrührer und die Lösung in einem Wasserbad für 30 Minuten bei 121 °C.
- Kühlen Sie die Lösung nach dem Autoklavieren in einem kalten Raum unter Rühren mit dem Magnetrührer ab, um eine Trennung in verschiedene Phasen zu verhindern. Bei Bedarf aliquotieren und bei 4 °C lagern.
- Bereiten Sie den Formulierungspuffer vor.
- Mischen Sie 500 ml DPBS mit 50 μl 10% Pluronic F-68. Sterilfilter durch eine 0,2 μm PES-Membranfiltereinheit und Lagerung bei Raumtemperatur (RT).
- Entwerfen und klonen Sie die homologiegesteuerte Reparaturschablone zwischen den invertierten Terminalwiederholungen (ITRs) von AAV2 im Vektor pAAV unter Verwendung von Standardtechniken. Stellen Sie sicher, dass die Homologiearme auf beiden Seiten mindestens ~ 250 bp betragen, aber nur 60 bp ausreichen können, obwohl längere Homologiearme bevorzugt werden, wenn das Konstruktionsdesign dies zulässt. Wenn Zielsequenzen einer der verwendeten sgRNAs im HDRT vorhanden sind, entfernen Sie sie mit stillen Mutationen, die am effektivsten in dem an den Protospacer angrenzenden Motiv oder der Seed-Region der Zielstelle sind.
- Zellkultur, Transfektion und Transduktion für die rAAV6-Produktion
- Tauen, kultivieren und frieren Sie 293AAV-Zellen ein, wie vom Hersteller beschrieben, unter Verwendung des oben genannten 293AAV-Kulturmediums und Trypsin-EDTA zur Spaltung. Es wird empfohlen, einige frühe Passagen einzufrieren und die Zellen für die AAV-Produktion zu verwenden, bevor sie Passage 40 erreichen.
- Für die rAAV6-Produktion werden vier 15-cm-Zellkulturschalen mit 5 x 106 Zellen in jeweils 30 ml ausgesät. Die Zellen sind in der Regel 1-2 Tage nach der Aussaat bereit für die Transfektion, wenn sie 80%-90% Konfluenz erreichen.
- Tauen Sie ein Maxiprep des pAAV-Plasmids auf, das das HDRT enthält, um es in AAV6 zu verpacken. Resuspendieren Sie 85,6 μg des pAAV-Plasmids in 3 ml reinem DMEM-Medium.
- 342 μl 1 mg/ml PEI-Lösung werden in 3 ml reinem DMEM-Medium gelöst. Inkubieren Sie beide Lösungen 10 Minuten lang bei RT.
- Mischen Sie beide 3-ml-Röhrchen in ein Röhrchen mit ~ 6,4 ml Transfektionsmischung und inkubieren Sie sie 20 Minuten lang bei RT.
- Tauen Sie in der Zwischenzeit den Tetracyclin-fähigen, selbstschalldämpfenden Hilfsvektor RepCap6 aus dem -80 °C-Gefrierschrank in einem 37 °C-Wasserbad auf. Um die 293AAV-Zellen zu transduzieren, wird der Helfervektor bei einer Multiplizität der Infektion (MOI) von 25 unter Verwendung der mittleren infektiösen Gewebekulturdosis (TCID 50) und unter der Annahme von 1,15 x10 7 Zellen/Schale hinzugefügt; In der Regel werden 2-10 μl pro 15-cm-Schale verwendet. Schaukeln und schwenken Sie das Geschirr vorsichtig, um es zu verteilen.
- Nach der Inkubation der Transfektionsmischung 1,6 ml davon tropfenweise auf jede der vier 15 cm großen Schalen geben. Über Nacht bei 37 °C und 5 % CO2 inkubieren.
HINWEIS: Wenn die interessierenden rAAV6-Vektoren bereits verfügbar sind, können diese Vektoren alternativ verwendet werden, um das zu verpackende virale Genom bereitzustellen, was die Notwendigkeit von Plasmiden mit diesem System negiert und vergleichbare rAAV6-Titer liefert. Für diesen Ansatz werden die 293AAV-Zellen mit dem gewünschten rAAV6 bei einem MOI von 50 (basierend auf rAAV6-Genomkopien [GC]/ml) zusammen mit dem Helfervektor co-transduziert. - Am nächsten Tag das Nährmedium vorsichtig absaugen, entsorgen und durch 30 ml vorgewärmtes Produktionsmedium ersetzen. Vor der Ernte weitere 96 Stunden inkubieren. Es wird kein weiterer Medienwechsel empfohlen, um die Erträge zu maximieren.
- Gewinnung und Reinigung des rekombinanten AAV6 aus dem Medium
- Sammeln Sie den gesamten Zellüberstand in einer Filtereinheit mit einer 0,2 μm PES-Membran, die mindestens 50 % größer ist als das Volumen des zu filternden Mediums, ohne die Zellen aus der Schale zu entfernen. Filtern Sie dann den Überstand.
HINWEIS: Wenn höhere Ausbeuten an rAAV6 gewünscht werden, können die Zellen geerntet und das rAAV aus dem Zellpellet unter Verwendung kommerzieller Kits oder etablierter Protokolle extrahiert werden34,35. Da AAV6 meist in das Medium36 sezerniert wird, wurde nur Überstand verwendet, wodurch Arbeit, Kosten und Zeit reduziert wurden. - 5x PEG/NaCl-Lösung wird mit 25% des gesammelten Volumens in den filtrierten Überstand gegeben; Dies sind typischerweise 30 ml, wenn vier 15-cm-Schalen mit 30 ml verwendet werden.
- Durch Umdrehen gut mischen und dann über Nacht bei 4 °C inkubieren, um die Viruspartikel auszufällen.
HINWEIS: AAV-Partikel sind in dieser Lösung bis zu 2 Tage stabil. - Kühlen Sie eine Schaufelzentrifuge mit 250 mL Röhrcheneinsätzen auf 4 °C vor. Bereiten Sie eine 4-ml-Zentrifugalfiltereinheit mit einem Cutoff-Wert von 100 kDa und einen hydrophilen PES-Spritzenfilter von 0,22 μm vor, indem Sie jede Membran mit 2 ml 10% Pluronic F-68 für mindestens 1 h bei RT vorbehandeln.
- Das AAV-PEG/NaCl-Gemisch wird in ein 250-ml-Röhrchen überführt, bei 2.500 x g 1 h bei 4 °C zentrifugiert und dann der gesamte Überstand vorsichtig durch Absaugen entfernt.
- Resuspendieren Sie das beige bis weiße Viruspellet durch Vortexen in 4 ml 1 M HEPES, bis es vollständig resuspendiert ist. Bei Bedarf 5 Minuten stehen lassen und erneut wirbeln. Resuspendieren Sie mit einer serologischen 5-ml-Pipette und überführen Sie das Gesamtvolumen in ein 15-ml-Röhrchen.
- Fügen Sie in einem Abzug der Virussuspension ein gleiches Volumen Chloroform hinzu - typischerweise 4 ml.
- 2 min kräftig wirbeln und dann bei 1.000 x g 5 min bei RT zentrifugieren.
- Sammeln Sie die oberste Schicht (AAV-haltiger Überstand) in einem neuen 50-ml-Röhrchen und entsorgen Sie die untere Schicht (Chloroform).
ACHTUNG: Chloroformhaltige Lösungen sind gefährliche Abfälle. Befolgen Sie die institutionellen Richtlinien für die Entsorgung. - Legen Sie den AAV-haltigen Überstand unter einen Abzug und lassen Sie das restliche Chloroform 30 Minuten lang verdampfen.
- Waschen Sie in der Zwischenzeit die vorbehandelte Zentrifugalfiltereinheit und den Spritzenfilter.
- Geben Sie 1,5 ml Formulierungspuffer in die vorbehandelte Zentrifugalfiltereinheit. Zentrifugieren bei 3.500 x g für 10 min bei 15 °C in einem schwingenden Schaufelrotor. Wiederholen Sie diesen Schritt mit 4 ml Formulierungspuffer, um die Membran zu waschen.
- Spülen Sie den Spritzenfilter zweimal mit 5 ml Formulierungspuffer mit einer 5-ml-Spritze aus.
- Füllen Sie den ~4 ml AAV-haltigen Überstand aus der Chloroformextraktion in eine 5-ml-Spritze, setzen Sie den gewaschenen Spritzenfilter ein und filtern Sie direkt in die Zentrifugalfiltereinheit.
- Zentrifugieren Sie bei 3.500 x g für 25 min bei 15 °C und vergewissern Sie sich dann, dass die AAV-Lösung im Filter etwa zwischen 50 und 100 μl liegt. Wenn das Volumen der Lösung >100 μl beträgt, zentrifugieren Sie weiter.
- Nach dem Entfernen des Filtrats werden 4 ml Formulierungspuffer in den Becher der Zentrifugalfiltereinheit gegeben und die Lösung durch Pipettieren gleichmäßig gemischt. Zentrifugieren Sie 3.500 x g für 25 min bei 15 °C und bestätigen Sie dann, dass die AAV-Lösung im Filter zwischen 50 und 100 μl liegt. Wenn das Volumen der Lösung >100 μl beträgt, zentrifugieren Sie weiter. Wiederholen Sie diesen Schritt für eine weitere Wäsche.
- Bestätigen Sie nach der abschließenden Zentrifugation, dass das Volumen der Lösung 50-70 μl beträgt. Wenn nicht, zentrifugieren Sie weiter. Übertragen Sie das Präparat in ein 1,5-ml-Röhrchen. Falls gewünscht, aliquotieren und bei −80 °C lagern.
- Sammeln Sie den gesamten Zellüberstand in einer Filtereinheit mit einer 0,2 μm PES-Membran, die mindestens 50 % größer ist als das Volumen des zu filternden Mediums, ohne die Zellen aus der Schale zu entfernen. Filtern Sie dann den Überstand.
- Rekombinante AAV6-Titerbestimmung mittels qPCR
HINWEIS: Die qPCR-Primer glühen im ITR-Bereich und sollten daher für alle in pAAV klonierten Konstrukte geeignet sein.- Tauen Sie ein Aliquot des zu titerierenden rAAV6 und ein Aliquot des AAV6-Referenzmaterials auf. Das AAV6-Referenzmaterial sollte nahe bei 4 x 1011 GC/ml liegen; Andernfalls passen Sie die Verdünnungen entsprechend an.
- Führen Sie einen DNase-I-Aufschluss durch, um die verbleibende freie Plasmid-DNA in der rAAV6-Präparation zu entfernen, indem 2,0 μl der Probe oder des AAV6-Referenzmaterials mit 15,6 μl nukleasefreiem H 2 O,2,0μl 10x DNase I-Puffer und 0,4 μl DNase I kombiniert werden.
- Vorsichtig mischen und 30 Minuten bei 37 °C inkubieren und dann auf Eis geben. Dies ist Verdünnung 1 (siehe Tabelle 3).
- Bereiten Sie fünffache serielle Verdünnungen aller Proben und des AAV6-Referenzmaterials wie in Tabelle 3 unten mit Wasser vor.
- Bereiten Sie einen SYBR Green qPCR-Mastermix vor. Mischen Sie pro Vertiefung 4,7 μl nukleasefreies Wasser mit 10 μl SYBR Green-Mastermix, 0,15 μl ITR-Primer vorwärts bei 100 μM und 0,15 μl ITR-Primer umgekehrt bei 100 μM.
HINWEIS: Jede Probe wird doppelt gemessen, mit 16 Vertiefungen für den Referenzstandard, 8 Vertiefungen pro Probe und 2 Vertiefungen für eine No-Template-Kontrolle. Bereiten Sie 10 % mehr Mastermix vor, um Pipettierfehler zu berücksichtigen. - Laden Sie in einer optischen 96-Well- oder 384-Well-Reaktionsplatte 15 μl/Well des SYBR Green qPCR-Mastermixes.
- Als nächstes laden Sie 5 μl Proben und AAV6-Referenzmaterial oder nukleasefreies Wasser für die No-Template-Kontrolle. Für den AAV6-Referenzstandard Lastverdünnung 2 bis Verdünnung 9. Für Proben wird die Verdünnung 5 bis 8 belastet. Messen Sie jede Verdünnung in zweifacher Ausfertigung. Vermeiden Sie Blasen.
- Versiegeln Sie die beladene Platte mit einer optisch transparenten Folie, zentrifugieren Sie sie bei 800 x g für 1 Minute bei RT und laden Sie die Platte mit dem entsprechenden 96-Well- oder 384-Well-Setup in das qPCR-Gerät.
- Einrichten und Ausführen des qPCR-Geräts mit SYBR-Detektion unter den folgenden Zyklusbedingungen: 98 °C für 3 min, dann 40 Zyklen von 98 °C für 15 s und 58 °C für 30 s, gefolgt von einer Schmelzkurve.
- Analysieren Sie die Daten mit der Software des Instruments unter Verwendung der Konzentration des AAV6-Referenzmaterials in Genomkopien pro Millimeter (GC/ml) als Standardkurve (siehe Tabelle 3). Berechnen Sie die Endkonzentration der Probe, indem Sie sie mit dem Verdünnungsfaktor multiplizieren.
- Stellen Sie sicher, dass die Standardkurve R2 nahe bei 1,0 liegt, die PCR-Effizienz 90 % bis 110 % beträgt, die Basislinie entfernt wurde, die Schmelzkurve einen einzelnen Peak zeigt, sichdie Ct-Werte entsprechend den Verdünnungen ändern und die Duplikate innerhalb von 0,5 Ct liegen; Andernfalls schließen Sie Ausreißer aus. Erwarten Sie Renditen wie in Abbildung 2.
2. Vorbereitung von B-Zell-Medien und -Stimuli
- Auftaumedium vorbereiten: Kombinieren Sie RPMI-1640 mit 20% FCS. Sterilfilter durch eine 0,2 μm PES-Membranfiltereinheit. Bei 4 °C lagern.
- B-Zellkulturmedium vorbereiten: Kombinieren Sie die Reagenzien in Tabelle 4 und filtern Sie sie dann durch eine 0,2-μm-PES-Membranfiltereinheit. Bei 4 °C lagern.
- Resuspendieren Sie jedes der B-Zell-Stimulanzien in Tabelle 5 in Stammkonzentrationen in B-Zellkulturmedium, mit Ausnahme von CpG ODN, das in nukleasefreiem Wasser resuspendiert werden sollte. Bei −80 °C lagern.
- Wenn Sie eine Nicht-B-Zell-negative Depletion durchführen (optionaler Schritt 4), bereiten Sie DPBS (kein Kalzium, kein Magnesium) mit 2% FCS (DPBS 2% FCS) vor. Sterilfilter durch eine 0,2 μm PES-Membranfiltereinheit. Bei 4 °C lagern.
3. Präparation und Kultivierung von Rhesusaffen-B-Zellen
ANMERKUNG: Kryokonservierte Rhesusaffen-PBMCs oder Splenozyten werden verwendet, um die Zellkultur30,31 aufzubauen.
- Auftaumedium und B-Zellkulturmedien in einem 37 °C heißen Wasserbad vorwärmen. Tauen Sie die B-Zell-Stimulanzien aus Tabelle 5 auf Eis auf.
- Bereiten Sie ein Röhrchen mit vorgewärmtem Auftaumedium in geeigneter Größe vor. Diese sollte idealerweise mehr als das 10-fache des Volumens der aufgetauten Zellen betragen.
- In einem Wasserbad bei 37 °C werden jeweils ein bis zwei Kryozellen mit PBMCs oder Splenozyten aufgetaut und in das vorbereitete Röhrchen mit vorgewärmtem Medium umgefüllt. Spülen Sie die Kryoröhren, um alle Zellen zu sammeln.
- Zentrifugieren Sie die Zellen bei 200 x g für 10 min bei RT.
HINWEIS: Diese Zentrifugationseinstellungen reduzieren die Thrombozytenkontamination bei gleichzeitiger Erhaltung der PBMC-Ausbeute. Es können höhere Geschwindigkeiten wie 350 x g für 5 min verwendet werden. - Resuspendieren Sie die Zellen zum Waschen in 10 ml Auftaumedium.
- Wiederholen Sie Schritt 3.4 und Schritt 3.5 für insgesamt drei Zentrifugationen, um das Gefriermedium zu entfernen. Nach der letzten Zentrifugation resuspendieren Sie die Zellen mit geschätzten ~5 x 106 Zellen/ml in B-Zellkulturmedium.
HINWEIS: Das obige Protokoll kultiviert ganze PBMC- oder Splenozytenpräparate mit Kontamination durch andere Zellen. Wenn reinere B-Zellkulturen erforderlich sind, wenn auch mit deutlich reduzierter Gesamt-B-Zellausbeute, fahren Sie mit Schritt 4 fort. Es wurden keine Unterschiede in der Bearbeitungseffizienz zwischen den beiden Methoden beobachtet. - Zur Zählung wird ein Aliquot von 10 μl Zellen nach Bedarf mit B-Zellkulturmedium verdünnt. Zählen Sie mit einem Hämozytometer und einer Trypanblau-Färbung, wobei gleiche Volumina resuspendierter Zellen und Trypanblau 0,4% ige Lösung kombiniert werden.
- Stellen Sie die Zellkonzentration auf 3 x 106 Zellen/ml mit B-Zellkulturmedium entsprechend der Zellzahl ein. Fügen Sie dann die B-Zell-Stimulanzien zu ihren endgültigen Konzentrationen gemäß Tabelle 5 hinzu und mischen Sie.
- Übertragen Sie die Zellen in eine geeignete Zellkulturschale. Insgesamt werden 0,6 x 10 6-0,7 x 106 Zellen/cm2 empfohlen. Inkubation der Zellen bei 37 °C mit 5% CO 2 für 48 h ±2 h.
4. Optionale negative Depletion von Nicht-B-Zellen
HINWEIS: Ausbeute und Reinheit hängen vom Input-Prozentsatz der B-Zellen unter den PBMCs ab, der sich zwischen den einzelnen Rhesusaffen drastisch unterscheiden kann27. Erwarten Sie eine Reinheit von 80 % bis 95 %, einen Wirkungsgrad von 60 % und 1 x 10 6-1,5 x 106 Zellen von 1 x 107 PBMCs.
- Nach dem letzten Waschen (Schritt 3.6) resuspendieren die Zellen bei 1 x 108 Zellen/ml in DPBS 2% FCS und humanem Fc-Block verdünnt 1:200. Die Zellzahlen basieren auf der Anzahl der aufgetauten Zellen.
- Inkubieren Sie 15 Minuten lang auf Eis, um die Fc-Rezeptoren zu blockieren, und fügen Sie dann die biotinylierten Antikörper in Tabelle 6 hinzu. Weitere 20 Minuten auf Eis inkubieren.
- Das Röhrchen mit DPBS 2% FCS auffüllen und bei 200 x g 10 min bei 4 °C schleudern.
- Resuspendieren Sie die Zellen in DPBS 2% FCS bei 80% des Volumens aus Schritt 4.1 (d. h. 80 μL pro 1 x 107 Zellen).
- Fügen Sie der Zellsuspension magnetische Streptavidinkügelchen mit 20 % des Volumens aus Schritt 4.1 hinzu (d. h. 20 μl Kügelchen pro 1 x 107 Zellen).
- Inkubieren Sie die Zellen 15 Minuten lang auf Eis und rühren Sie gelegentlich um.
- Bereiten Sie in der Zwischenzeit pro 1 x 108 Zellen einen Magnetabscheider mit einer großen magnetischen Verarmungssäule und einem Vorabscheidefilter vor. Spülen Sie den Vorabscheidefilter und die Säule mit 2 ml DPBS 2% FCS durch Schwerkraftströmung und entsorgen Sie den Durchfluss. Installieren Sie ein 15-ml-Auffangröhrchen.
HINWEIS: Die Verwendung anderer Säulen wie positiver Selektionssäulen oder anderer magnetischer Perlenreinigungssysteme kann die Reinheit drastisch reduzieren. - Füllen Sie die Zellen nach der Inkubation auf 0,5 ml mit DPBS 2% FCS auf, wenn das Volumen <0,5 ml beträgt. Wenn das Volumen ≥0,5 ml beträgt, fahren Sie einfach fort.
- Laden Sie die Zellsuspension in den Vorabscheidefilter auf der vorbereiteten Säule und sammeln Sie den Durchfluss in das 15-ml-Röhrchen.
- Die ungebundenen angereicherten B-Zellen werden zweimal durch Zugabe von 1 ml DPBS 2% FCS in den Vorabscheidefilter eluiert. Sammeln Sie die ungebundenen Zellen durch Schwerkraft in derselben Röhre.
HINWEIS: Eine zusätzliche Elution kann die Ausbeute geringfügig erhöhen. Die Reinheit und Effizienz kann durch die Durchflusszytometrie der Eingangszellen, angereicherten Zellen und Zellen, die auf der Säule zurückgehalten werden, bewertet werden. Um die auf der Säule verbliebenen Zellen zu erhalten, entfernen Sie die Säule vom Magneten und spülen Sie sie mit dem mitgelieferten Kolben mit 3 ml DPBS 2% FCS ab. Falls gewünscht, bewerten Sie die Reinheit durch Durchflusszytometrie wie in Abbildung 3 unter Verwendung der Reagenzien in Tabelle 7. - Zentrifugieren Sie die angereicherten B-Zellen bei 200 x g für 10 min bei 4 °C.
- Resuspendieren Sie die Zellen bei geschätzten ~ 5 x 106 Zellen/ml in B-Zellkulturmedium und fahren Sie mit Schritt 3.7 fort.
5. Primäre Rhesusaffen-B-Zell-Gen-Editierung
- Nachdem Sie die Rhesusaffen-B-Zellen für 48 h ± 2 h aktiviert haben, bereiten Sie die Reagenzien für die Elektroporation und Transduktion vor.
- Vorwärm-DMSO, nukleasefreier Duplex-Puffer, Puffer T und Puffer E (10-μl-Elektroporationskit) oder E2 (100-μl-Elektroporationskit) aus dem Elektroporationskit auf RT.
- Tauen Sie die rAAV6-HDRT- und B-Zell-Stimulanzien aus Tabelle 5 auf Eis auf.
- Resuspendieren Sie die CRISPR-Cas9-sgRNAs bei 100 μM im Duplexpuffer. 10 Minuten bei RT rekonstituieren und durch Vortexen und Schnippen mischen. Bewahren Sie die rekonstituierten sgRNAs bis zur Verwendung auf Eis auf. Bei −80 °C lagern.
HINWEIS: CRISPR-Cas9 sgRNAs können mit verschiedenen Online-Tools (siehe 1.1.1) entworfen werden und können in ihrer Schneideffizienz drastisch variieren. Eine empirische Prüfung der Schneideffizienz wird mit Assays wie TIDE37 oder ICE38 empfohlen. - Bereiten Sie pro 10 μl Elektroporation 550 μl B-Zellkulturmedium mit allen Stimulanzien aus Tabelle 5 vor und fügen Sie 1% DMSO hinzu. Skalieren Sie die Volumina um das 10-fache für 100-μl-Elektroporationen. Optional können 10% dieses Mediums ohne Antibiotika-Antimykotikum hergestellt werden, was die Zelllebensfähigkeit nach der Transfektion leicht erhöht.
- Bereiten Sie pro 10-μl-Elektroporation eine Vertiefung einer 48-Well-Zellkulturplatte mit 50 μl des B-Zellkulturmediums mit Stimulanzien und ohne Antibiotika-Antimykotikum vor, falls verwendet. Für 100-μl-Elektroporationen pipettieren Sie 500 μl in die Vertiefungen einer 6-Well-Platte.
- Fügen Sie dem Medium in den Vertiefungen rAAV6 HDRT hinzu, bis zu 20 % des Volumens im Vertiefungs. Ziel sind MOIs im Bereich von 1 x 10 5-1 x 10 6 basierend auf der Anzahl der Zellen pro Transfektion (10 μL Elektroporation: 5 x 10 5 Zellen; 100 μL Elektroporation:5 x 106 Zellen) und der GC in der rAAV6-Präparation anzustreben. Hohe rAAV6-Stammkonzentrationen von 5 x 1013 GC/ml bis 5 x 1014 GC/ml werden empfohlen, um hohe MOIs mit geringen Volumina zu erreichen.
HINWEIS: Niedrigere MOIs können zu einer verringerten Bearbeitungseffizienz führen, und MOIs von 5 x 105 liegen im Allgemeinen nahe an der maximalen Bearbeitungseffizienz, die wir gesehen haben. Ein Einfluss unterschiedlicher MOIs auf die Lebensfähigkeit von B-Zellen wurde nicht beobachtet. Es wird empfohlen, Kontrollen ohne rAAV6 HDRT, ohne RNP-Transfektion und ohne beides einzuschließen. - Das zubereitete Geschirr und das restliche Medium vorwärmen, indem man es in einen Inkubator bei 37 °C mit 5% CO2 überführt.
- Bereiten Sie pro 10 μl Elektroporation 1,15 μl Ribonukleoprotein (RNP) vor: Mischen Sie 0,4 μl 61 μM Cas9 mit 0,75 μl 100 μM sgRNA in Duplexpuffer. Bereiten Sie sich aufgrund eines Pipettierfehlers und zur Vermeidung von Blasen beim Laden der Elektroporationsspitzen extra vor (30% mehr für eine einzelne Elektroporation wird empfohlen). 10-fach skalieren für 100 μL Spitzen.
- Inkubieren Sie das RNP mindestens 15 Minuten lang bei RT, bevor Sie es mit den Zellen mischen. Nach der Inkubation können mehrere RNPs kombiniert werden, wenn mehr als ein Locus gleichzeitig anvisiert werden soll. Es wurden keine signifikanten Unterschiede in der Effizienz bei bis zu drei Loci gleichzeitig beobachtet.
- Bereiten Sie in der Zwischenzeit die Zellen für die Elektroporation vor. Halten Sie die Zellen immer auf RT, um Temperaturschocks zu vermeiden. Ernten Sie die Zellen nach 48 h ± 2 h Kultur in ein geeignetes Gefäß. Spülen Sie das Geschirr mit DPBS aus, um die maximale Anzahl von Zellen zu sammeln.
- Zentrifugieren Sie die Zellen bei 200 x g für 10 min bei RT. Verwerfen Sie den Überstand und resuspendieren Sie die Zellen in DPBS bei ~2 x 106 Zellen/ml.
- Kombinieren Sie 10 μl Trypanblau 0,4% ige Lösung mit 10 μl der Zellsuspension und zählen Sie mit einem Hämozytometer.
HINWEIS: Erwarten Sie zu diesem Zeitpunkt aufgrund von Verlusten während der Ernte und des Waschens etwa 60% der Zellen, die 48 Stunden ± 2 Stunden früher in Kultur gebracht wurden. - In der Zwischenzeit zentrifugieren Sie die Zellen bei 200 x g für 10 min bei RT. Entsorgen Sie den Überstand und achten Sie darauf, dass die verbleibenden DPBS minimiert werden. Resuspendieren Sie die Zellen in vorgewärmtem (RT) Puffer T bei 5,55 x 107 Zellen/ml basierend auf der obigen Zellzahl.
- Richten Sie das Transfektionssystem ein, indem Sie das Gerät einschalten und auf 1.350 V, 15 ms und 1 Impuls einstellen. Platzieren Sie die Pipettenstation in der Laminar-Flow-Haube
- Bereiten Sie für jeden Satz von 10 Elektroporationen ein Transfektionsröhrchen mit 3 ml Puffer E (für 10 μl-Transfektionen) oder E2 (für 100 μl-Transfektionen) vor. Führen Sie das Röhrchen in die Pipettenstation ein.
- Kombinieren Sie pro 10 μl Elektroporation 1,15 μl RNP mit 9 μl Zellen. Stellen Sie sicher, dass ein ausreichendes Volumen (+ 30%) vorhanden ist, um zu vermeiden, dass Luft in die Elektroporationsspitze gesaugt wird. Vor der Elektroporation 1-2 Minuten bei RT inkubieren.
- Saugen Sie 10 μl oder 100 μl RNP und Zellgemisch in die entsprechend große Elektroporationsspitze einer Elektroporationspipette ein, führen Sie die beladene Pipette in die Pipettenstation ein und starten Sie die Elektroporation. Stellen Sie sicher, dass die Spitzen vollständig frei von Luftblasen sind, um Lichtbögen zu vermeiden. Beobachten Sie während der Elektroporation, ob kein Lichtbogen auftritt.
- Die elektroporierten Zellen werden sofort in das vorbereitete, vorgewärmte, kleine Volumen des Mediums mit oder ohne rAAV6 in der 48-Well- (10-μl-Transfektionen) oder 6-Well-Platte (100-μl-Transfektionen) ausgeworfen. Wiederholen Sie die Schritte 5.15 bis 5.17 mit den verbleibenden Beispielen. Kontrollproben ohne Transfektion in die Kulturvertiefungen geben.
- Inkubieren Sie die Zellen bei 37 °C mit 5 % CO 2 für 4 h ±2 h und fügen Sie dann das vorbereitete, vorgewärmte B-Zellkulturmedium hinzu, das Stimulanzien, DMSO und Antibiotika/Antimykotikum enthält: 450 μl für 10 μl Transfektionen oder 4,5 ml für 100 μl Transfektionen.
- Die Inkubation bei 37 °C mit 5% CO2 für 12-24 h fortsetzen. Wechseln Sie dann das Medium zu einem B-Zellkulturmedium, das Stimulanzien und Antibiotika/Antimykotikum ohne DMSO enthält, wenn eine längere Kultivierung gewünscht wird. Die Analyse der genomischen DNA kann nach 24 h durchgeführt werden. Digitale Tröpfchen-PCR unter Verwendung eines Primers außerhalb des Homologiearms und eines Primers innerhalb des Inserts kann verwendet werden, um die Bearbeitungseffizienz39 zu quantifizieren. Führen Sie PCRs durch, um die Insertionsstelle zu amplifizieren, und führen Sie eine Sanger-Sequenzierung durch, um die korrekte Bearbeitung zu überprüfen.
- Für die Analyse des Proteingehalts werden die Zellen nach der Elektroporation 40-48 h lang kultiviert, um Änderungen der Proteinexpression zu ermöglichen, und eine Analyse mittels Durchflusszytometrie unter Verwendung der Reagenzien in Tabelle 7 durchgeführt.
Ergebnisse
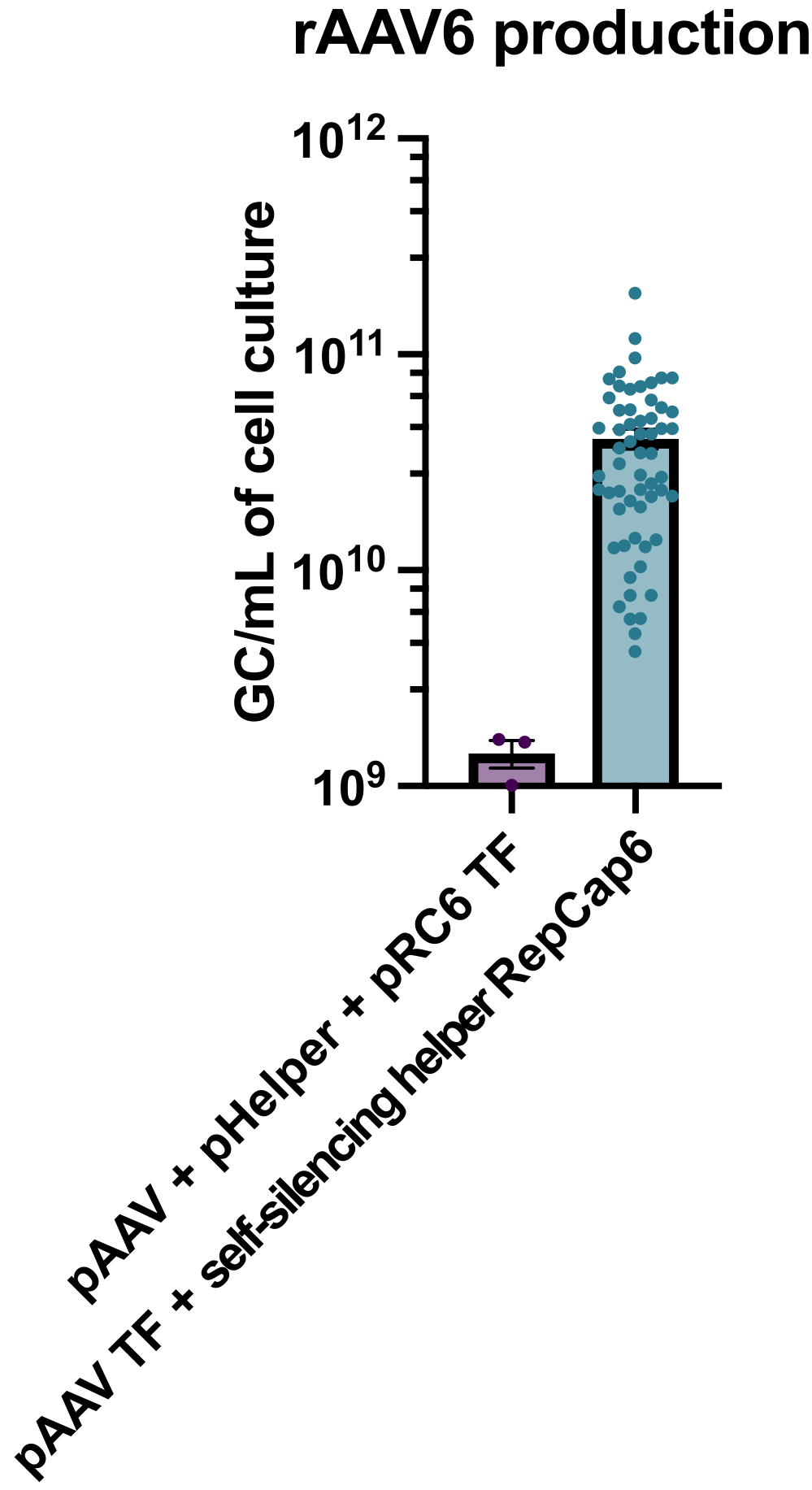
Die Produktion von rAAV6 unter Verwendung des Tetracyclin-fähigen, selbstsilencierenden adenoviralen Helfers führte zu einer Produktion von durchschnittlich 4 x10 10 GC/ml Zellkulturmedium und übertraf damit die Produktion mit einer standardmäßigen, helferfreien Dreifachtransfektion um das 30- bis 40-fache (Abbildung 2).
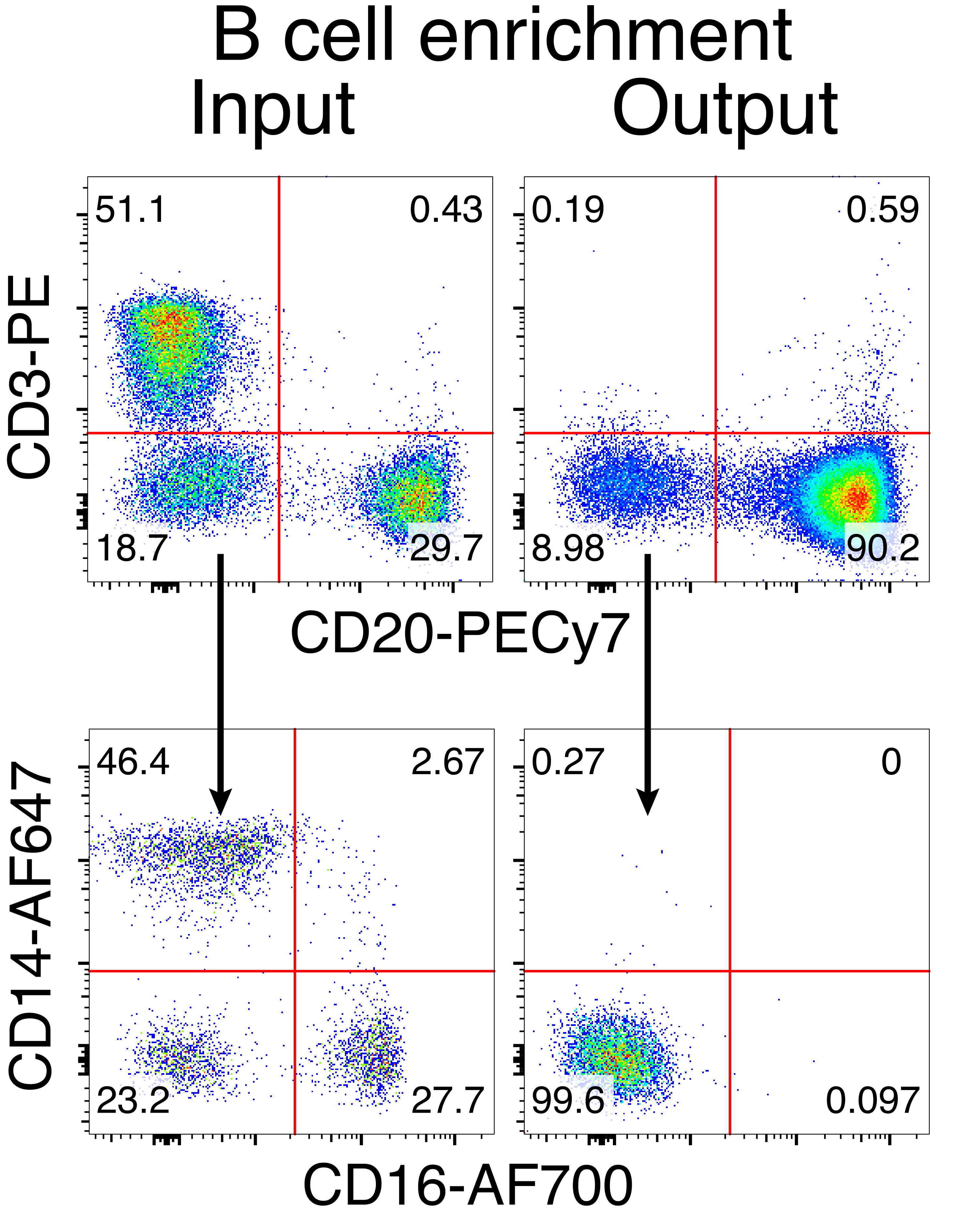
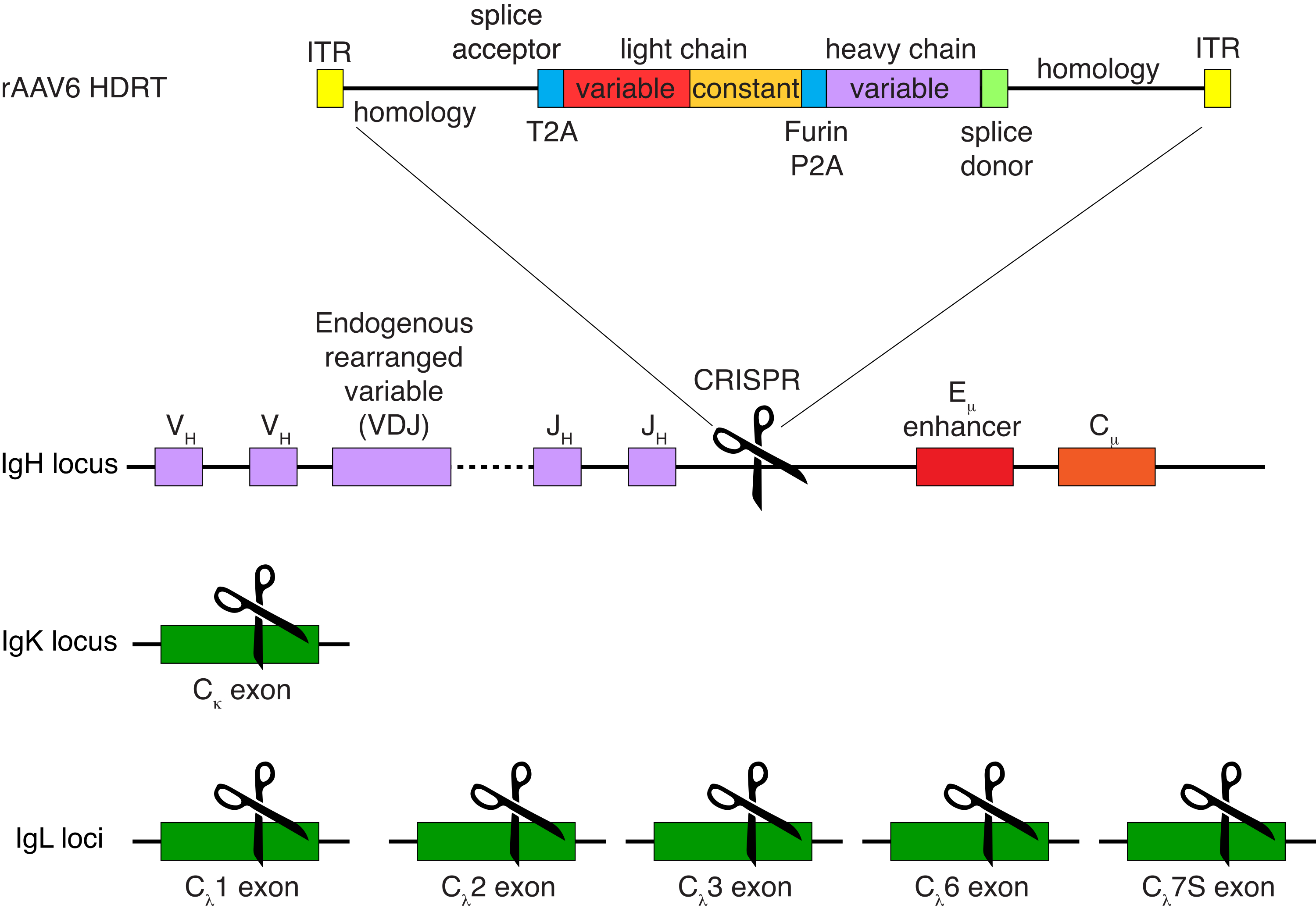
Die optionale Aufreinigung von Rhesusaffen-B-Zellen führte zur Eliminierung der überwiegenden Mehrheit der CD3+ T-Zellen und CD14+ und/oder CD16+ myeloischen Zellen, wobei routinemäßig Reinheiten von 80%-95% CD20+ B-Zellen erhalten wurden (Abbildung 3). Basierend auf unseren früheren Designs in murinen B-Zellen7 haben wir eine Methode entwickelt, um die B-Zell-Rezeptor-Spezifität von Rhesusaffen-B-Zellen zu editieren und gleichzeitig den Allelausschluss in der überwiegenden Mehrheit der B-Zellen aufrechtzuerhalten, indem endogene Antikörper-Leichtketten durch die Störung ihrer konstanten Region gelöscht werden. Wir konstruierten ein promotorloses HDRT, das in den IGH-Locus zwischen dem letzten IGHJ-Gen und dem Eμ-Enhancer von Rhesusaffen-B-Zellen eingefügt werden sollte (Abbildung 4). Dieses Konstrukt nutzt den endogenen VH-Promotor der natürlich umgelagerten vorgelagerten VDJ-Region in reifen B-Zellen und wird daher nicht von episomalen AAV-Genomen exprimiert. Darüber hinaus erfordert dieses Konstrukt das Spleißen in nachgeschaltete Antikörper-Schwerketten-Konstantenregionen, die auf der Zelloberfläche exprimiert werden sollen. Daher zeigt die spezifische Antigenbindung auf der Zelloberfläche, die durch Durchflusszytometrie gezeigt wird, eine korrekte Ziellocus-Integration an und dass die eingefügte Sequenz funktionsfähig ist.
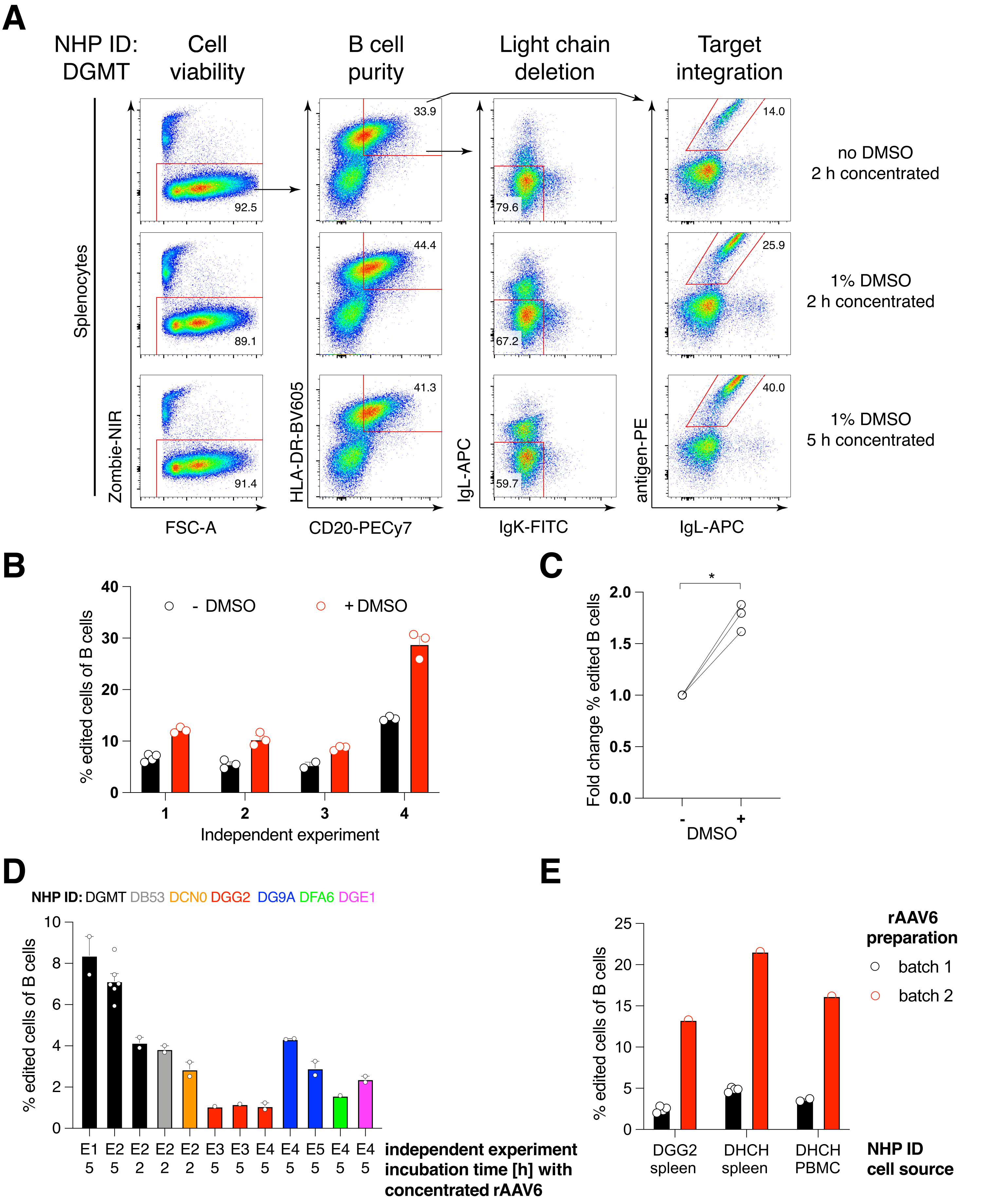
Wir haben ein solches Konstrukt, das für den Antikörper Ab1485, ein von Rhesusaffen abgeleitetes Anti-HIV-bNAb40, kodiert, in rAAV6 verpackt und es verwendet, um aktivierte primäre Rhesusaffen-Splenozyten- oder PBMC-Kulturen zu editieren, wie oben beschrieben (Abbildung 5A). Das Protokoll behielt eine hohe Zellviabilität (~90%) bei, während gleichzeitig die Leichtkettenexpression in ~80% der B-Zellen gelöscht wurde. Die Mehrzahl der B-Zellen exprimierte noch den Isotyp IgM (Abbildung 5B). Die Zugabe des rAAV6, das für das Ab1485 HDRT kodiert, führte in 16%-21% der B-Zellen zu Gen-Editierung und Ab1485-Oberflächenexpression (Abbildung 5A), wenn auch mit einer geringeren Fluoreszenzintensität für Antikörperketten als für unbearbeitete B-Zellen (Abbildung 5A rechtes Bild, Abbildung 5C). Dies kann das Ergebnis einer Epitopkonkurrenz zwischen der Antigenfärbung und den Monoklonalen sein, die zum Nachweis der Oberflächen-BCR in der Durchflusszytometrie verwendet werden, sowie einer tatsächlich reduzierten Proteinexpression aufgrund der polycistronischen Natur des HDRT und des weniger effizienten Spleißens. Die Zugabe von 1% DMSO und ausgedehnte, konzentrierte Inkubationen mit dem rAAV6 HDRT erhöhten im Allgemeinen die Bearbeitungseffizienz (Abbildung 6A-C). Mit dieser speziellen Methode, die typischerweise 5 % bis 20 % und bis zu 40 % beträgt, wird eine Bearbeitungseffizienz in Abhängigkeit vom einzelnen Rhesusaffen (Abbildung 5A, Abbildung 6A-E) und der Qualität des rAAV6-HDRT-Batches (Abbildung 6E) erreicht. Insgesamt stellen wir Protokolle für eine effiziente rAAV6-Produktion sowie die Kultivierung, Aufreinigung und Genomierung von Rhesusaffen-B-Zellen vor.
| Reagenzien | Volumen | Lager | Endkonzentration |
| DMEM, hohe Glukose | 500 ml | 1 x | ~ 88.5% |
| FCS, hitzeinaktiviert | 50 ml | 1 x | ~ 8.85% |
| Antibiotikum/Antimykotikum | 5 mL | 100 x | 1 x |
| Glutamin | 5 mL | 200 mM | 2 mM |
| Natriumpyruvat | 5 mL | 100 mM | 1 mM |
Tabelle 1: Das 293AAV-Zellkulturmedium.
| Reagenzien | Volumen | Lager | Endkonzentration |
| DMEM, hohe Glukose | 500 ml | 1 x | ~ 95.2% |
| FCS, hitzeinaktiviert | 10 ml | 1 x | ~ 1,9% |
| Antibiotikum/Antimykotikum | 5 mL | 100 x | 1 x |
| Glutamin | 5 mL | 200 mM | 2 mM |
| Natriumpyruvat | 5 mL | 100 mM | 1 mM |
Tabelle 2: Das 293AAV-Zellproduktionsmedium.
| Verdünnungs-Serie | Volumen der Probe (μl) | Verdünnungsmittel und Volumen | Verdünnungsfaktor | Gesamtverdünnung | Referenz AAV6 |
| GC/ml | |||||
| Verdünnung 1 | 2 μl Probe oder AAV-Referenzstandard bei 4,1 x 1011 GC/ml | 18 μL DNAseI-Puffer und -Enzym | 10 x | 10 x | 4.1 x 1010 |
| Verdünnung 2 | 15 μL Dil. 1 | 60 μL H2O | 5 x | 50 x | 8.2 x 109 |
| Verdünnung 3 | 20 μL Dil. 2 | 80 μL H2O | 5 x | 250 x | 1.6 x 109 |
| Verdünnung 4 | 20 μL Dil. 3 | 80 μL H2O | 5 x | 1250 x | 3.3 x 108 |
| Verdünnung 5 | 20 μL Dil. 4 | 80 μL H2O | 5 x | 6250x | 6.6 x 107 |
| Verdünnung 6 | 20 μL Dil. 5 | 80 μL H2O | 5 x | 31250 x | 1.3 x 107 |
| Verdünnung 7 | 20 μL Dil. 6 | 80 μL H2O | 5 x | 156250 x | 2.6 x 106 |
| Verdünnung 8 | 20 μL Dil. 6 | 80 μL H2O | 5 x | 781250 x | 5.24 x 105 |
| Verdünnung 9 | 20 μL Dil. 7 | 80 μL H2O | 5 x | 3906250 x | 1.05 x 105 |
Tabelle 3: qPCR-Verdünnungstabelle.
| Reagenz | Volumen | Lager | Endkonzentration |
| RPMI-1640 | 420 mL | 1 x | 84% |
| FCS, hitzeinaktiviert | 50 ml | 1 x | 10% |
| Antibiotikum/Antimykotikum | 5 mL | 100 x | 1 x |
| Glutamin | 5 mL | 200 mM | 2 mM |
| Natriumpyruvat | 5 mL | 100 mM | 1 mM |
| HEPES | 5 mL | ca. 1 m | 10 mM |
| 2-B-Mercapto-Ethanol | 550 μL | 55 mM | 55 μM |
| Nicht-essentielle Aminosäuren | 5 mL | 100 x | 1 x |
| Insulin-Transferin-Selen | 5 mL | 100 x | 1 x |
Tabelle 4: B-Zellkulturmedium.
| Reagenz | Verdünnung | Lager | Endkonzentration |
| MegaCD40L | 1:1000 | 100 μg/ml | 100 ng/ml |
| CpG ODN | 1:300 | 1 mg/ml | 3,33 μg/ml |
| Menschliches BAFF | 1:1000 | 40 μg/ml | 40 ng/ml |
| Menschliches IL-2 | 1:1000 | 50 μg/ml | 50 ng/ml |
| Menschliches IL-10 | 1:1000 | 50 μg/ml | 50 ng/ml |
Tabelle 5: B-Zell-Stimulanzien.
| Antikörper | Klonen | Verdünnung | Abschließende Konz. |
| Anti-Human-CD3 | FN-18 | 1:40 | 2,5 μg/ml |
| Anti-Human-CD8a | RPA-T8 | 1:200 | 2,5 μg/ml |
| Anti-Human-CD14 | M5E2 | 1:200 | 2,5 μg/ml |
| Anti-Human-CD16 | 3G8 | 1:200 | 2,5 μg/ml |
| Anti-Mensch-CD33 | AC104.3E3 | 1:50 | 1 Test |
| Anti-Mensch-CD64 | 10.1 | 1:800 | 0,625 μg/ml |
| Anti-Mensch-CD66 | TET2 | 1:11 | 1 Test |
| Anti-Mensch-CD89 | Autobahn A59 | 1:800 | 0,625 μg/ml |
Tabelle 6: Antikörper für den optionalen Abbau von Nicht-B-Zellen.
| Reagenz | Typ/Klon | Arbeitsverdünnung/-konzentration |
| Anti-Mensch CD14 AlexaFluor647 | M5E2 | 1:50 |
| Anti-Mensch CD16 AlexaFluor700 | 3G8 | 1:50 |
| Anti-Human-CD20 PECy7 | 2H7 | 1:50 |
| Anti-Human-CD3-PE | SP34-2 | 1:50 |
| Zombie-NIR | - | 1:500 |
| Anti-Human-HLA-DR BV605 | L243 | 1:200 |
| Anti-Mensch Ig-Leichtkette Lambda APC | MHL-38 | 1:50 |
| Anti-Mensch Kappa Lichterkette FITC | polyklonal | 1:500 |
| Anti-Human-IgM BV421 | MHM-88 | 1:50 |
| RC1-Antigen, zufällig biotinyliert | - | 5 μg/ml |
| Streptavidin-PE | - | 1:500 |
Tabelle 7: Durchflusszytometrische Reagenzien für die Analyse.

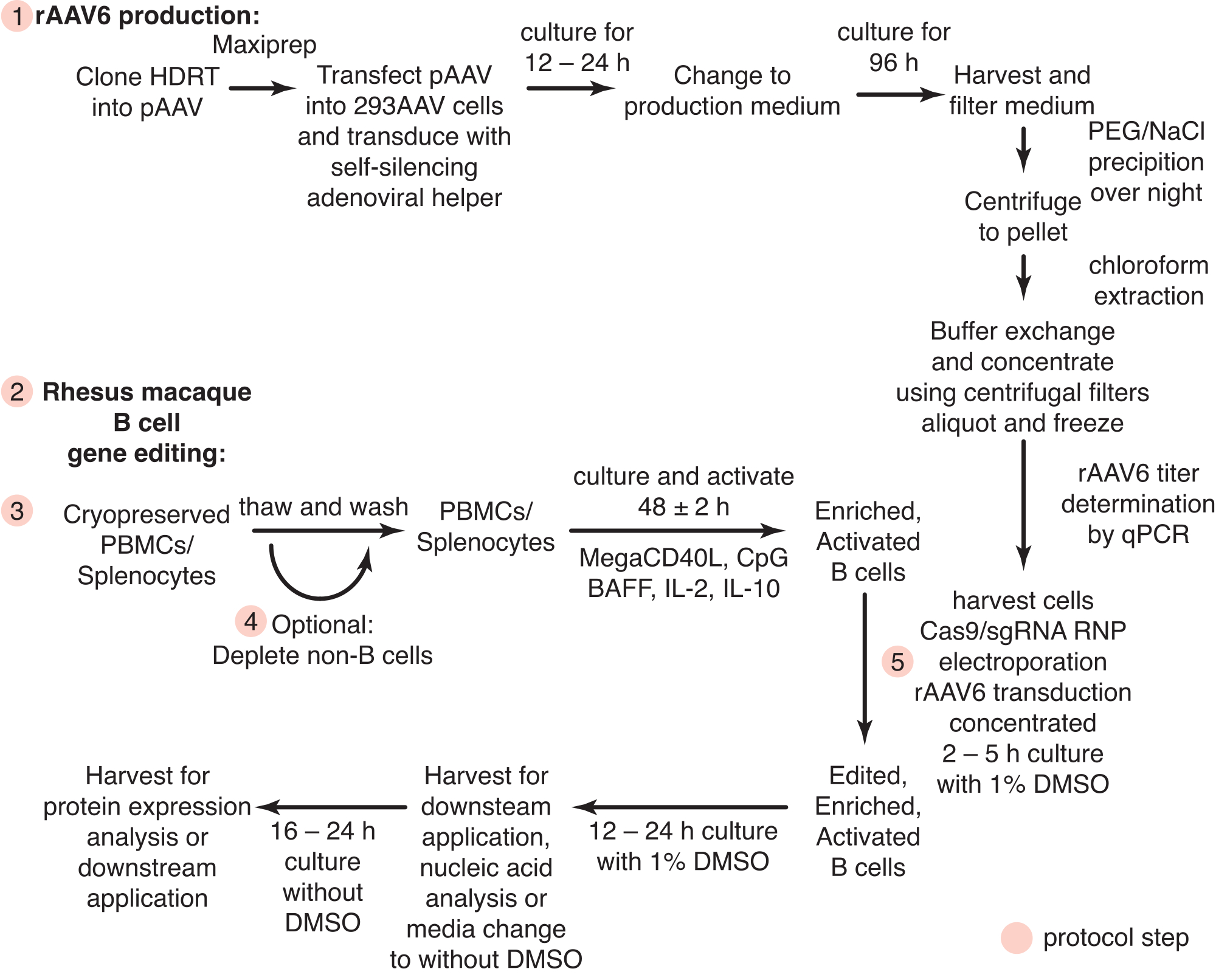
Abbildung 1: Schematische Darstellung der rAAV6-Produktion und der Gen-Editierung von primären Rhesusaffen-B-Zellen. Die Protokolle gliedern sich in die rAAV6-Produktion (Schritt 1) und die Gen-Editierung von Rhesusaffen-B-Zellen (Schritte 2-5), einschließlich eines optionalen Schritts zur Depletion von Nicht-B-Zellen (Schritt 4). Schritte in den Protokollen sind mit roten Kreisen gekennzeichnet. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 2: Hohe rAAV6-Ausbeuten unter Verwendung eines selbststummschaltenden adenoviralen Helfers. rAAV6 wurde mit den hier beschriebenen Methoden (pAAV-Transfektion [TF] + Selbst-Silencing-Helfer RepCap6, Selbst-Silencing-Adenoviraler Helfer) oder typischer Helfer-freier Dreifach-Transfektion von pAAV, pHelper und pRepCap6 (pRC6) hergestellt. rAAV6 wurde nur aus dem Zellüberstand gereinigt. Die Methoden, die die selbststummschaltenden adenoviralen Helfervektoren verwendeten, produzierten 30-40-fach mehr rAAV, titeriert durch qPCR, wie oben beschrieben. Jeder Punkt repräsentiert eine individuelle rAAV-Produktion unter Verwendung verschiedener pAAV-Konstrukte aus 2 bis 20 unabhängigen Experimenten. Der Mittelwert ± SEM wird aufgetragen. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 3: B-Zell-Anreicherung durch die negative Depletion von Nicht-B-Zellen. Rhesusaffen-B-Zellen wurden unter Verwendung des beschriebenen Protokolls aus PBMCs angereichert und auf eine Reinheit von 90% angereichert. Der Input und Output vor der Anreicherung nach der Anreicherung werden angezeigt. Eingezäunt auf lebenden Singulett-PBMCs. Repräsentativ für fünf unabhängige Experimente. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 4: Targeting-Strategie zur Editierung der B-Zell-Rezeptor-Spezifität von Rhesusaffen-B-Zellen. rAAV6 wurde mit dem abgebildeten HDRT hergestellt. Das HDRT besteht aus einem 266 bp 5'-Homologiearm, gefolgt von 111 bp des Rhesusaffen-IGHM-Exon-1-Spleißakzeptors, dann einem GSG-Linker mit einer selbstspaltenden 2A-Peptidsequenz (T2A) des Thosea-Asigna-Virus , gefolgt von einer Leader-Sequenz und der vollständigen leichten Kette des Rhesusaffen-Antikörpers Ab1485 als Rhesusaffe IGLC1. Darauf folgen eine Furin-Spaltstelle, ein GSG-Linker und eine porcine teschovirus-selbstspaltende 2A-Peptidsequenz (Furin-P2A), gefolgt von einer weiteren Leader-Sequenz und der Schwerkettenvariablen Ab1485, gefolgt von 52 bp der Rhesusaffen-IGHJ4-Spleißspendersequenz, um das Spleißen in nachgeschaltete Antikörper-Schwerketten-Konstantenregionen zu ermöglichen, und einem 514-bp-Homologiearm. Dieses Konstrukt wurde unter Verwendung der sgRNA-Zielsequenz GAGATGCCAGAGCAAACCAG in den IGH-Locus zwischen dem letzten IGHJ-Gen und dem Eμ-Enhancer gezielt. Beide Homologiearme wurden so konzipiert, dass sie an der Schnittstelle dieser sgRNA enden, wodurch die Zielsequenz entfernt wird und eine optimale Integrationseffizienz ermöglicht wird. Um den allelischen Ausschluss und die Expression eines einzelnen B-Zell-Rezeptors aufrechtzuerhalten, haben wir gleichzeitig endogene Leichtketten mit sgRNAs deletiert, die auf den Rhesusaffen IGKC mit der Zielsequenz GGCGGGAAGATGAAGACAGA und IGLC1, IGLC2, IGLC3, IGLC6 und IGLC7S unter Verwendung der Zielsequenz CTGATCAGTGACTTCTACCC abzielen. Die HDRT enthielt stille Mutationen, die die Spaltung der IGLC1-Sequenz durch diese sgRNA verhinderten. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 5: Gen-Editierung von primären Rhesusaffen-B-Zellen . (A) Primäre Splenozyten (oberes Bild) oder PBMCs (unteres Bild) desselben Rhesusaffen wurden ohne Abbau von Nicht-B-Zellen kultiviert und wie oben beschrieben editiert. Die Targeting-Strategie war wie in Abbildung 4 dargestellt. Zwei Tage nach der Elektroporation wurden die Zellen geerntet und für die durchflusszytometrische Analyse oberflächengefärbt. Die linke Spalte wurde auf Singulett-Zellen eingezäunt, und die anderen Spalten wurden dann eingezäunt, wie in der oberen Zeile angegeben. Die Viabilität der Zellen, die Reinheit der B-Zellen, die Deletionseffizienz der Leichtketten und die Knock-in-Effizienz von Ab1485 durch Färbung mit dem spezifischen Antigen RC141 sind in unbehandelten, RNP-transfizierten oder RNP-transfizierten + rAAV6-transduzierten Proben (MOI = 5 x 105) angegeben. Stellvertretend für sechs unabhängige Experimente mit Zellen verschiedener Rhesusaffen. (B) IgM-Expression auf kultivierten Rhesusaffen-B-Zell-Kontrollen oder nach Editierung und (C) geometrische mittlere Fluoreszenzintensität (gMFI) von IgM auf B-Zellen, die die Ig-Expression aufgrund von IgLC- und IgKC-Targeting nicht verloren haben (uneditiert) oder B-Zellen, die das erwartete Antigen binden (editiert). Der rote Punkt zeigt den gMFI von kultivierten, nicht transfizierten Kontroll-B-Zellen an. gibt p < 0,0001 in einem gepaarten t-Test an. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 6: Auswirkungen von DMSO, verlängerter konzentrierter Inkubation mit rAAV6 HDRT, rAAV-Batch-Qualität und Reproduzierbarkeit zwischen verschiedenen Spender-NHPs auf die Gen-Editing-Effizienz in primären Rhesusaffen-B-Zellen. (A) Splenozyten wurden wie beschrieben kultiviert und editiert. Nach der Elektroporation wurden 5 x 10 5 Zellen in Medium mit oder ohne 1% DMSO kultiviert und in 50 μL Medium, das rAAV6 HDRT enthielt, bei einem MOI von 5 x 10 5 für entweder 2 h oder5 h inkubiert, bevor weitere 450 μl Medium zugegeben wurden. Die Zellen wurden 2 Tage nach der Elektroporation mittels Durchflusszytometrie analysiert, wie in Abbildung 5 dargestellt. Stellvertretend für vier unabhängige Experimente. (B) Quantifizierung von (A) über vier unabhängige Experimente. Die Punkte kennzeichnen technische Replikate mit Transfektionseinstellungen von 1.350 V, 10-20 ms und 1 Puls-Elektroporationsdauer und DMSO-Konzentrationen im Bereich von 0,75 % bis 1,25 %. (C) Durchschnittliche Falzänderung der Bearbeitungseffizienz von (B). * p > 0,05 im Mann-Whitney-U-Test. (D) Bearbeitungseffizienz gegenüber unabhängigen Experimenten mit verschiedenen Makaken unter Verwendung einer kommerziellen rAAV6-Charge mit geringerer Effizienz. (E) Editiereffizienz unter Verwendung von zwei verschiedenen kommerziellen Chargen von rAAV6, in die das gleiche Konstrukt in den B-Zellen von zwei verschiedenen NHPs im selben Experiment verpackt wurde. Die Punkte kennzeichnen technische Replikate mit Transfektionseinstellungen von 1.350 V, 10-20 ms und 1 Impulselektroporation. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}
Diskussion
Die hier vorgestellten Protokolle bieten eine schnelle und effiziente Methode zur Erzeugung hoher Ausbeuten und Titer von rAAV6s als HDRTs und neuartige Methoden zur effizienten Gen-Editierung primärer Rhesusaffen-B-Zellen in vitro.
Das rAAV6-Produktionsprotokoll ist vergleichsweise einfach und schnell und ermöglicht die gleichzeitige Herstellung und Prüfung vieler verschiedener Konstrukte ohne übermäßigen Arbeitsaufwand. Falls gewünscht, kann rAAV6 unter Verwendung etablierter Protokolle wie der Iodixanol-Gradienten-Ultrazentrifugation34 oder der wässrigen Zweiphasenpartitionierung35 vor dem Pufferaustausch und der Konzentration weiter gereinigt werden.
Obwohl es die Gesamtausbeute verringerte, entschieden wir uns dafür, nur serumreduziertes Zellkulturmedium für die rAAV6-Aufreinigung anstelle der Aufreinigung aus dem Zellpellet zu verwenden, da der Großteil von rAAV6 in das Medium36 freigesetzt wird und die Reinigung aus dem Zellpellet mehr Kosten und Arbeit verursacht. Die Verwendung des selbstinaktivierenden adenoviralen Helfers erhöhte die Ausbeute im Durchschnitt um das 30- bis 40-fache und ermöglichte das Testen von in AAV6 verpackten Konstrukten in einer einzigen 15-cm-Schale. Obwohl unsere Aufreinigungsmethode einfach ist, erhalten wir mit dieser Methode eine relativ geringe Variation der Gen-Editing-Effizienz oder der Zelllebensfähigkeit nach der Transduktion unter Verwendung verschiedener Zelllinien oder anderer Primärzellen (Daten nicht gezeigt).
Wir haben ein Rhesusaffen-B-Zell-Reinigungsprotokoll entwickelt, um unberührte primäre B-Zellen unter Verwendung der negativen Depletion unerwünschter Populationen zu erhalten. Obwohl es für die Genom-Editierung dieser Zellen nicht notwendig ist, bietet es eine Möglichkeit, eine relativ reine Population von primären Rhesusaffen-B-Zellen für diese oder andere Anwendungen zu erhalten, falls andere Zelltypen die experimentellen Ziele stören. Die Reinheit geht jedoch auf Kosten einer verringerten Gesamtausbeute an B-Zellen. Bemerkenswert ist, dass sowohl für die angereicherten als auch für die nicht angereicherten B-Zellkulturen der Anteil der B-Zellen in den ursprünglichen PBMC- oder Splenozytenpräparaten entscheidend ist. Insbesondere für PBMCs empfehlen wir, verschiedene Makaken auf Individuen mit einem hohen Anteil an B-Zellen im peripheren Blut zu untersuchen, um eine hohe Anzahl von B-Zellen für Experimente zu erhalten, da dieser Wert zwischen Individuen dramatisch variieren kann27. PBMCs können durch regelmäßige Blutungen oder Leukapherese erhalten werden42.
Das Gen-Editing-Protokoll führt zu einer effizienten Gen-Editierung, typischerweise zwischen 60 % und 80 % der Knock-in-B-Zellen und 5 % -20 % der Knock-in-B-Zellen, obwohl wir bis zu 90 % BCR-Knock-out und 40 % BCR-Knock-in-B-Zellen erreicht haben (Abbildung 5 und Abbildung 6).
Die wichtigsten Parameter für die effiziente Bearbeitung von Rhesusaffen-B-Zellen sind die Schneideeffizienz der sgRNA, die Elektroporationsparameter, der MOI und die Qualität der rAAV6-Präparation. Die Schneideeffizienz von Kandidaten-sgRNAs sollte empirisch bestimmt werden, um eine optimale Bearbeitung und Gestaltung des HDRT zu ermöglichen. Die hier vorgestellten Elektroporationsparameter balancieren Effizienz und Viabilität aus, um die maximale Gesamtzahl der editierten B-Zellen und nicht den höchsten Prozentsatz der editierten B-Zellen zu erhalten. Wenn ein höherer Prozentsatz an bearbeiteten Zellen erforderlich ist, werden erhöhte Spannungen (bis zu 1.750 V) oder veränderte Pulslängen (10-30 ms) empfohlen, obwohl mehr Zelltod beobachtet werden kann. Wir stellten auch eine etwas höhere Bearbeitungseffizienz in Milz-B-Zellen im Vergleich zu B-Zellen aus PBMCs desselben Individuums fest (Abbildung 5); Der zugrunde liegende Grund dafür ist jedoch derzeit nicht bekannt.
Wir fanden heraus, dass die Zugabe von 1% DMSO nach der Elektroporation die Gen-Editing-Effizienz in Rhesusaffen-B-Zellen signifikant um ~40% erhöhte, ohne die Zelllebensfähigkeit zu beeinträchtigen (Abbildung 6A-C), in Übereinstimmung mit Berichten in anderen Zellen43. Eine verlängerte Kultur in 1% DMSO sollte jedoch vermieden werden und kann die Lebensfähigkeit der Zellen beeinträchtigen. DMSO kann auf Wunsch komplett weggelassen werden.
Die Kultivierung der Zellen in einem kleinen Volumen nach mehrstündiger Elektroporation zusammen mit dem rAAV6 führt zu höheren Editiereffizienzen, wahrscheinlich aufgrund der besseren Transduktion von HDRT durch das rAAV6 und damit der höheren intrazellulären Konzentration von HDRT zum relevanten Zeitpunkt, wenn Cas9 aktiv ist. Wir fanden heraus, dass die Kultivierung der Zellen auf diese Weise für bis zu 8 Stunden die Lebensfähigkeit der Zellen nicht beeinträchtigte, aber die Bearbeitungseffizienz stieg nicht dramatisch über 5 Stunden hinaus (Abbildung 6). Wenn nur Knock-out statt Knock-In erforderlich ist, kann dieser Schritt entfallen.
Abschließend stellen wir umfassende Protokolle für die Gen-Editierung von Rhesusaffen-B-Zellen in vitro und die Produktion von rAAV6 HDRT vor, die für das effiziente Einklopfen gewünschter Konstrukte erforderlich sind. Diese Protokolle ermöglichen die schnelle und kostengünstige Prüfung vieler Konstrukte, die als rAAV6 verpackt sind, und ermöglichen die präklinische Prüfung der Machbarkeit und Skalierbarkeit von B-Zelltherapien in einem relevanteren nicht-menschlichen Primatenmodell.
Offenlegungen
Interessenkonflikte werden nicht deklariert.
Danksagungen
Wir danken Harry B. Gristick und Pamela Bjorkman für die Bereitstellung des RC1-Antigens und den gesamten Laboratorien von Nussenzweig und Martin für die kritische Diskussion. Diese Arbeit wurde durch das Stipendium der Bill and Melinda Gates Foundation INV-002777 (an M.C.N.) und das intramurale Forschungsprogramm des National Institute of Allergy and Infectious Diseases, National Institutes of Health, unterstützt. (R.G. und M.A.M). M.C.N. ist ein HHMI-Ermittler.
Materialien
| Name | Company | Catalog Number | Comments |
| 1.5 mL tube sterile, Dnase, Rnase and purogen free | Stellar Scientific | T17-125 | or similar |
| 10 mL serological pipette, polystyrene, sterile, nonpyrogenic, DNase-/RNase-free, and Human DNA-free | Corning | 4488 | or similar |
| 15 cm tissue culture dish | Falcon | 353025 | or similar |
| 15 mL polypropylene conical tybe | Falcon | 352097 | or similar |
| 25 mL serological pipette, polystyrene, sterile, nonpyrogenic, DNase-/RNase-free, and Human DNA-free | Corning | 4489 | or similar |
| 250 mL polypropylene conical tybe | Corning | 430776 | or similar |
| 293AAV cell line | Cell Biolabs | AAV-100 | |
| 2-B-mercapto-ethanol, 55mM (1000x) | Gibco | 21985-023 | |
| 48-well tissue culture plate | Corning | 3548 | or similar |
| 5 mL serological pipette, polystyrene, sterile, nonpyrogenic, DNase-/RNase-free, and Human DNA-free | Corning | 4487 | or similar |
| 5 mL syringes with Luer-Lok Tip | BD | 309646 | or similar |
| 50 mL polypropylene conical tybe | Falcon | 352070 | or similar |
| 50 mL serological pipette, polystyrene, sterile, nonpyrogenic, DNase-/RNase-free, and Human DNA-free | Corning | 4490 | or similar |
| 6-well tissue culture plate | Falcon | 353046 | or similar |
| AAV-6 Packaging System (plasmids) | Cell Biolabs | VPK-406 | |
| AAV6 Reference Materials (full capsids) | Charles River | RS-AAV6-FL | |
| Accu-jet S Pipette Controller | Brand | 26350 | or similar pipette controller |
| Antibiotic/Antimycotic 100x | Gibco | 15260-062 | |
| anti-human CD14 AlexaFluor647 | Biolegend | 301812 | |
| anti-human CD14 biotin | BioLegend | 301826 | |
| anti-human CD16 AlexaFluor700 | BD Biosciences | 557920 | |
| anti-human CD16 biotin | BioLegend | 302004 | |
| anti-human CD20 PECy7 | Biolegend | 302312 | |
| anti-human CD3 biotin | Thermo Fisher | APS0309 | |
| anti-human CD3 PE | BD Biosciences | 552127 | |
| anti-human CD33 biotin | Miltenyi | 130-113-347 | |
| anti-human CD64 biotin | BioLegend | 305004 | |
| anti-human CD66 biotin | Miltenyi | 130-100-143 | |
| anti-human CD89 biotin | BioLegend | 354112 | |
| anti-human CD8a biotin | BioLegend | 301004 | |
| anti-human HLA-DR BV605 | Biolegend | 307640 | |
| anti-human Ig light chain lambda APC | Biolegend | 316610 | |
| anti-human IgM BV421 | Biolegend | 314516 | |
| anti-Human Kappa Light Chain FITC | Fisher Scientific | A18854 | |
| Autoclave | Steris | Amsco Lab 250 | or similar |
| Cell culture CO2 incubator | Fisher Scientific | 51026331 | or similar |
| Centrifugal Filter Unit (Amicon Ultra - 4, 100 kDa) | Millipore | UFC810024 | |
| Centrifuge 5920 R | Eppendorf | EP022628188 | or any other, coolable swinging bucket centrifuge with inserts for 96-well plates, 15, 50 and 250 mL size tubes |
| Chloroform | Fisher Scientific | C298SK-4 | |
| Cpg ODN | Invivogen | tlrl-2395 | |
| Dimethyl sulfoxide (DMSO) | Sigma-Aldrich | 34869-500ML | |
| DMEM, High Glucose | Gibco | 11965092 | |
| DNaseI (RNase-free) | New England Biolabs | M0303L | |
| DPBS, no calcium, no magnesium | Gibco | 14190144 | |
| Electroporation kit (Neon Transfection System 10 µL) | Fisher Scientific | MPK1096 | or other sizes or 100 uL transfection kit MPK 10096 |
| Electroporation system (Neon Transfection System) | Fisher Scientific | MPK5000 | |
| FCS | Hyclone | SH30910.03* | |
| Ficoll-PM400 (Ficoll-Paque PLUS) | Cytiva | 17144002 | or similar |
| Fume Hood | Fisher Scientific | FH3943810244 | or similar |
| Glutamine 200 mM | Gibco | 25030-081 | |
| Graduated Cylinder 1L | Corning | 3022-1L | or similar |
| Hemocytometer | Sigma-Aldrich | Z375357-1EA | or similar |
| HEPES 1M | Gibco | 15630-080 | |
| HEPES 1M | Gibco | 15630-080 | |
| Hot Plate Magnetic Stirrer | Fisher Scientific | SP88857200 | or similar |
| Human BAFF | Peprotech | 310-13 | |
| Human BD Fc Block | BD | 564220 | |
| Human IL-10 | Peprotech | 200-10 | |
| Human IL-2 | Peprotech | 200-02 | |
| Hydrochloric acid | Fisher Scientific | A144S-500 | |
| Hydrophilic Polyethersulfone Syringe Filters, (Supor membrane), Sterile - 0.2 µm, 25 mm | Pall | 4612 | |
| Insulin-Transferin-Selenium, 100x | Gibco | 41400-045 | |
| ITR primer forward: GGAACCCCTAGTGATGGAGTT | Integrated DNA Technologies | custom | |
| ITR primer reverse: CGGCCTCAGTGAGCGA | Integrated DNA Technologies | custom | |
| Laminar flow biosafety cabinet | The Baker Company | SG403A | or similar |
| Large magnetic depletion (LD) Column | Miltenyi Biotec | 130-042-901 | |
| Magentic seperator (MidiMACS separator and multistand) | Miltenyi Biotec | 130-090-329 | |
| Magnetic stir bar | Fisher Scientific | 14-512-127 | or similar |
| Magnetic streptavidin beads (Streptavidin MicroBeads) | Miltenyi Biotec | 130-048-101 | |
| Maxiprep kit | Machery-Nagel | 740414.5 | or similar |
| Media Bottles 2L with cap | Cole-Parmer | UX-34514-26 | or similar |
| MegaCD40L | Enzo | ALX-522-110-C010 | |
| MicroAmp Optical 384-well Reaction Plate | Fisher Scientific | 4309849 | |
| MicroAmp Optical Adhesive Film | Fisher Scientific | 4311971 | |
| Microcentrifuge 5424 R | Eppendorf | 5404000014 | or any other table top centrifuge for 1.5 mL tubes |
| Microwave oven | Panasonic | NN-SD987SA | or similar |
| Nikon TMS Inverted Phase Contrast Microscope | Nikon | TMS | or any other Inverted phase-contrast microscope for cell culture |
| Non-essential amino acids, 100x | Gibco | 11140-050 | |
| Nuclease-free Duplex buffer | Integrated DNA Technologies | 11-01-03-01 | |
| Nuclease-free Water | Qiagen | 129115 | |
| pH meter | Mettler Toledo | 30019028 | or similar |
| Pipetman Classic Starter Kit, 4 Pipette Kit, P2, P20, P200, P1000 and tips | Gilson | F167380 | or similar set of pipettes and tips |
| Pluronic F-68 10 % | Gibco | 24040-032 | |
| Polyethylene Glycol 8000 | Fisher Scientific | BP233-1 | |
| Polyethylenimine, Linear, MW 25000, Transfection Grade (PEI 25K | Polysciences | 23966-100 | |
| Precision Balance | Mettler Toledo | ME4001TE | or similar |
| Pre-Separation Filters (30 µm) | Miltenyi Biotec | 130-041-407 | |
| Pyrex glass beaker 2 L | Cole-Parmer | UX-34502-13 | or similar |
| Pyrex glass beaker 250 mL | Millipore Sigma | CLS1000250 | or similar |
| qPCR Instrument | Fisher Scientific | 4485691 | or similar |
| RC1 antigen randomly biotinylated | Bjorkman lab, CalTech | in house | |
| RPMI-1640 | Gibco | 11875-093 | |
| S.p. Cas9 Nuclease | Integrated DNA Technologies | 1081059 | |
| Scientific 1203 Water Bath | VWR | 24118 | or any water bath set to 37 °C |
| Sodium chloride | Sigma-Aldrich | S7653-5KG | |
| Sodium hydroxide | Sigma-Aldrich | S8045-500G | |
| Sodium Pyruvate 100 mM | Gibco | 11360-070 | |
| Sterile Disposable Filter Units with PES Membranes | Thermo Scientific Nalgene | 567-0020 | |
| Streptavidin-PE | BD Biosciences | 554061 | |
| SYBR Green Master Mix | Fisher Scientific | A25742 | |
| Tetracycline-enabled, self-silencing adenoviral vector RepCap6 | Oxgene | TESSA-RepCap6 | |
| Trypan Blue Solution, 0.4% | Gibco | 15250061 | |
| Trypsin-EDTA (0.05%), phenol red | Gibco | 25300054 | |
| Water Purification System | Millipore Sigma | ZEQ7000TR | or similar |
| Zombie-NIR | Biolegend | 423106 |
Referenzen
- Victora, G. D., Nussenzweig, M. C. Germinal centers. Annual Review of Immunology. 40, 413-442 (2022).
- Plotkin, S. A. Correlates of protection induced by vaccination. Clinical and Vaccine Immunology. 17 (7), 1055-1065 (2010).
- Brinkmann, V., Heusser, C. H. T cell-dependent differentiation of human B cells into IgM, IgG, IgA, or IgE plasma cells: High rate of antibody production by IgE plasma cells, but limited clonal expansion of IgE precursors. Cellular Immunology. 152 (2), 323-332 (1993).
- Chernecky, C. C., Berger, B. J. Protein Electrophoresis - Serum., 6th edition. , Elsevier Saunders. St Louis, MO. 917-920 (2013).
- Balazs, A. B., et al. Antibody-based protection against HIV infection by vectored immunoprophylaxis. Nature. 481 (7379), 81-84 (2011).
- Greiner, V., et al. CRISPR-mediated editing of the B cell receptor in primary human B cells. iScience. 12, 369-378 (2019).
- Hartweger, H., et al. HIV-specific humoral immune responses by CRISPR/Cas9-edited B cells. Journal of Experimental Medicine. 216 (6), 1301-1310 (2019).
- Huang, D., et al. Vaccine elicitation of HIV broadly neutralizing antibodies from engineered B cells. Nature Communications. 11, 5850(2020).
- Jeske, A. M., Boucher, P., Curiel, D. T., Voss, J. E. Vector strategies to actualize B cell-based gene therapies. Journal of Immunology. 207 (3), 755-764 (2021).
- Nahmad, A. D., et al. In vivo engineered B cells secrete high titers of broadly neutralizing anti-HIV antibodies in mice. Nature Biotechnology. 40 (8), 1241-1249 (2022).
- Nahmad, A. D., et al. Engineered B cells expressing an anti-HIV antibody enable memory retention, isotype switching and clonal expansion. Nature Communications. 11, 5851(2020).
- Voss, J. E., et al. Reprogramming the antigen specificity of B cells using genome-editing technologies. eLife. 8, 42995(2019).
- Pesch, T., et al. Molecular design, optimization, and genomic integration of chimeric B cell receptors in murine B cells. Frontiers in Immunology. 10, 2630(2019).
- Cheong, T. C., Compagno, M., Chiarle, R. Editing of mouse and human immunoglobulin genes by CRISPR-Cas9 system. Nature Communications. 7, 10934(2016).
- Rogers, G. L., Cannon, P. M. Genome edited B cells: A new frontier in immune cell therapies. Molecular Therapy. 29 (11), 3192-3204 (2021).
- Hung, K. L., et al. Engineering protein-secreting plasma cells by homology-directed repair in primary human B cells. Molecular Therapy. 26 (2), 456-467 (2018).
- Johnson, M. J., Laoharawee, K., Lahr, W. S., Webber, B. R., Moriarity, B. S. Engineering of primary human B cells with CRISPR/Cas9 targeted nuclease. Scientific Reports. 8, 12144(2018).
- Wu, C. M., et al. Genetic engineering in primary human B cells with CRISPR-Cas9 ribonucleoproteins. Journal of Immunological Methods. 457, 33-40 (2018).
- Luo, B., et al. Engineering of alpha-PD-1 antibody-expressing long-lived plasma cells by CRISPR/Cas9-mediated targeted gene integration. Cell Death and Disease. 11 (11), 973(2020).
- Laoharawee, K., et al. Genome engineering of primary human B cells using CRISPR/Cas9. Journal of Visualized Experiments. (165), e61855(2020).
- Laoharawee, K., Johnson, M. J., Moriarity, B. S. CRISPR/Cas9-mediated genome engineering of primary human B cells. Methods in Molecular Biology. 2115, 435-444 (2020).
- Moffett, H. F., et al. B cells engineered to express pathogen-specific antibodies protect against infection. Science Immunology. 4 (35), (2019).
- Hartweger, H., Nussenzweig, M. C. CRISPR comes a-knock-in to reprogram antibodies in vivo. Nature Biotechnology. 40 (8), 1183-1184 (2022).
- Nishimura, Y., Martin, M. A. Of mice, macaques, and men: Broadly neutralizing antibody immunotherapy for HIV-1. Cell Host & Microbe. 22 (2), 207-216 (2017).
- Shedlock, D. J., Silvestri, G., Weiner, D. B. Monkeying around with HIV vaccines: Using rhesus macaques to define 'gatekeepers' for clinical trials. Nature Reviews Immunology. 9 (10), 717-728 (2009).
- Kreuser, S., et al. Efficient methods for generation and expansion of, and gene delivery to rhesus macaque plasma B cells. bioRxiv. , (2021).
- Gujer, C., Sundling, C., Seder, R. A., Karlsson Hedestam, G. B., Lore, K. Human and rhesus plasmacytoid dendritic cell and B-cell responses to Toll-like receptor stimulation. Immunology. 134 (3), 257-269 (2011).
- Kim, J. S., et al. Cell enrichment-free massive ex-vivo expansion of peripheral CD20(+) B cells via CD40-CD40L signals in non-human primates. Biochemical and Biophysical Research Communications. 473 (1), 92-98 (2016).
- Su, W., et al. Self-attenuating adenovirus enables production of recombinant adeno-associated virus for high manufacturing yield without contamination. Nature Communications. 13, 1182(2022).
- Endo, Y., et al. Short- and long-term clinical outcomes in rhesus monkeys inoculated with a highly pathogenic chimeric simian/human immunodeficiency virus. Journal of Virology. 74 (15), 6935-6945 (2000).
- Balaphas, A., Buchs, N. C., Meyer, J., Hagen, M. E., Morel, P. Partial splenectomy in the era of minimally invasive surgery: The current laparoscopic and robotic experiences. Surgical Endoscopy. 29 (12), 3618-3627 (2015).
- Gibson, D. G., et al. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nature Methods. 6 (5), 343-345 (2009).
- Labun, K., et al. CHOPCHOP v3: Expanding the CRISPR web toolbox beyond genome editing. Nucleic Acids Research. 47, 171-174 (2019).
- Strobel, B., Miller, F. D., Rist, W., Lamla, T. Comparative analysis of cesium chloride- and iodixanol-based purification of recombinant adeno-associated viral vectors for preclinical applications. Human Gene Therapy Methods. 26 (4), 147-157 (2015).
- Guo, P., et al. Rapid and simplified purification of recombinant adeno-associated virus. Journal of Virological Methods. 183 (2), 139-146 (2012).
- Vandenberghe, L. H., et al. Efficient serotype-dependent release of functional vector into the culture medium during adeno-associated virus manufacturing. Human Gene Therapy. 21 (10), 1251-1257 (2010).
- Brinkman, E. K., Chen, T., Amendola, M., van Steensel, B. Easy quantitative assessment of genome editing by sequence trace decomposition. Nucleic Acids Research. 42 (22), 168(2014).
- Conant, D., et al. Inference of CRISPR edits from Sanger trace data. The CRISPR Journal. 5 (1), 123-130 (2022).
- Wilkinson, A. C., et al. Cas9-AAV6 gene correction of beta-globin in autologous HSCs improves sickle cell disease erythropoiesis in mice. Nature Communications. 12, 686(2021).
- Wang, Z., et al. A broadly neutralizing macaque monoclonal antibody against the HIV-1 V3-Glycan patch. eLife. 9, 61991(2020).
- Escolano, A., et al. Immunization expands B cells specific to HIV-1 V3 glycan in mice and macaques. Nature. 570 (7762), 468-473 (2019).
- Pathiraja, V., Matar, A. J., Gusha, A., Huang, C. A., Duran-Struuck, R. Leukapheresis protocol for nonhuman primates weighing less than 10 kg. Journal of the American Association for Laboratory Animal Science. 52 (1), 70-77 (2013).
- Stratigopoulos, G., De Rosa, M. C., LeDuc, C. A., Leibel, R. L., Doege, C. A. DMSO increases efficiency of genome editing at two non-coding loci. PLoS One. 13 (6), 0198637(2018).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten