
Method Article
Afinación de la degradación para lograr un agotamiento específico y eficiente de las proteínas
En este artículo
Resumen
Aquí, presentamos un protocolo para agotar de forma efectiva y específica una proteína de interés en la levadura Saccharomyces cerevisiae utilizando el sistema AID.
Resumen
El receptor de unión a la auxina vegetal, TIR1, reconoce las proteínas que contienen un motivo específico de degron inducible en auxina (AID) en presencia de auxina, apuntando a su degradación. Este sistema se explota en muchos eucariotas no vegetales, de modo que una proteína diana, etiquetada con el motivo AID, se degrada a la adición de auxina. El nivel de expresión TIR1 es crítico; la expresión excesiva conduce a la degradación de la proteína etiquetada con AID incluso en ausencia de auxina, mientras que la baja expresión conduce a un agotamiento lento. Se creó un sistema de AYUDA inducible por estradiol, con la expresión de TIR1 bajo el control de un promotor inducible de estradiol. El nivel de TIR1 es ajustable cambiando el tiempo de incubación con el estradiol antes de la adición de auxina. Este protocolo describe cómo agotar rápidamente una proteína diana utilizando el sistema AID. El tiempo adecuado de incubación de estradiol depende de la abundancia de la proteína diana. Por lo tanto, el agotamiento eficiente depende de una sincronización óptima que también minimice el agotamiento independiente de la auxina.
Introducción
Las mutaciones condicionales, como los mutantes sensibles a la temperatura, son una poderosa herramienta para el estudio de proteínas esenciales, permitiendo el crecimiento celular bajo la condición permisiva pero causando pérdida de función en condiciones no permisivas. Sin embargo, metabolismo celular puede ser seriamente perturbado por el cambio en las condiciones de crecimiento necesario para inducir el defecto y también puede crear efectos fuera del objetivo. Se han desarrollado varios métodos, en los que la proteína de interés se secuestra condicionalmente1 o su expresión se controla 2,3 mediante la adición de una molécula pequeña. Este protocolo utiliza la auxina y el sistema de degron inducible de auxina (AID) para agotar eficientemente una proteína diana.
El sistema AID tiene su origen en las plantas, donde se utiliza una auxina (en este protocolo indole-3-ácido acético (IAA), estimula la interacción dela proteína Aux/IAA con TIR1, miembro del complejo de ligasas ubiquitinas SCF U3 4. La interacción compleja de SCF causa poliubiquitinación de proteínas de la familia Aux/IAA, lo que resulta en su degradación por el proteosoma5,6.
Este sistema fue previamente adaptado para su uso en la levadura Saccharomyces cerevisiae7,8 expresando la proteína TIR1 de Oriza sativa (osTIR) en células de levadura, donde es capaz de interactuar con la levadura endógena Complejo SCF. La proteína de interés fue etiquetada con un motivo de la proteína Aux/IAA IAA17 para apuntar a su degradación. Los truncamientos funcionales de IAA17 se desarrollaron más tarde, como AID*8,9,10, que contiene el motivo 43 aminoácido sin ayuda sensible de Arabidopsis thaliana IAA17, junto con una etiqueta de epítopo para permitir Detección.
El sistema inicialmente adaptado para su uso en levadura en ciernes7,8 expresó la proteína osTIR1 de un promotor de GAL de levadura. La expresión requiere cambiar a medio de crecimiento con galactosa como única fuente de carbono, lo que, por desgracia, resulta en un cambio diónico con cambios de amplio alcance en el metabolismo celular11. Por otro lado, se ha informado de que la expresión constitutiva de TIR1 puede conducir a la degradación de la proteína diana en ausencia de auxina/IAA12 si el nivel de expresión es alto, mientras que la expresión TIR1 baja causa un agotamiento ineficiente. Se desarrolló un sistema de AID mejorado denominado AID en el que el osTIR está bajo el control de un promotor inducible que es ajustable para adaptarse a la proteína diana, con un efecto mínimo sobre el metabolismo celular. Para lograrlo, se construyó un factor de transcripción artificial (ATF) en el que el activador de transcripción viral VP16 se fusiona con un receptor de estrógenos y un dominio de unión al ADN (DBD) de cuatro dedos Zn. Cuando está presente el estradiol (un estrógeno), el ATF puede entrar en el núcleo e inducir la transcripción del osTIR uniéndose a su promotor (Z4EVpr)13,12.
la expresión osTIR suele ser detectable unos 20 minutos después de la adición de é-estradiol12. Sin embargo, la duración óptima de la expresión osTIR para lograr un agotamiento eficiente de la proteína etiquetada con auxina, evitando el agotamiento antes de la adición de auxina, debe determinarse empíricamente para cada proteína diana. Un tiempo aproximado para esta pre-incubación se puede estimar a partir de los valores de abundancia en la base de datos del genoma de Saccharomyces (SGD https://www.yeastgenome.org/). Como se puede observar en la Figura 1, la proteína abundante, Dcp1 (2880 a 4189 moléculas/célula), requiere 40 minutos de pre-incubación con el estradiol, sin agotamiento independiente de la auxina observado. La proteína mucho menos abundante, Prp2 (172 a 211 moléculas/célula), se agota fuertemente después de sólo 20 minutos de pre-incubación. Es aconsejable probar dos tiempos de pre-incubación adicionales, 10 a 20 minutos antes o después de este tiempo estimado inicial (20 min es el tiempo mínimo que se recomienda). El tiempo óptimo de pre-incubación es el momento en el que la proteína objetivo no se ha agotado antes de añadir auxina y una vez que se añade auxina el agotamiento es aceptable o los niveles de proteína se acercan al mínimo posible. Por lo tanto, de la Figura 1b, para Prp22 con 30 min de pre-incubación, los niveles no han disminuido mucho 10 minutos después de la adición de auxin. Comparando esto con 40 min de pre-incubación y 15 min con IAA, donde hay poco agotamiento adicional, no hay ningún beneficio en incubar con auxina de más de 10 minutos o pre-incubación por más de 30 min, particularmente porque hay evidencia de no auxina agotamiento dependiente a 40 min. Para Dcp1 con 40 min de pre-incubación (el último punto en el que el nivel de proteína es aproximadamente 100% antes de la adición de auxina), 15 a 20 minutos de agotamiento con auxina es aceptable. Se recomienda mantener el tiempo de agotamiento lo más corto posible para reducir los efectos secundarios sobre el metabolismo celular14.
En este artículo se demuestra cómo utilizar el sistema DE Ayuda mediante la optimización del momento de la incubación de é-estradiol para la expresión de osTIR para lograr un rápido agotamiento de la proteína diana en la adición de IAA sin agotamiento antes de agregar auxina.
Protocolo
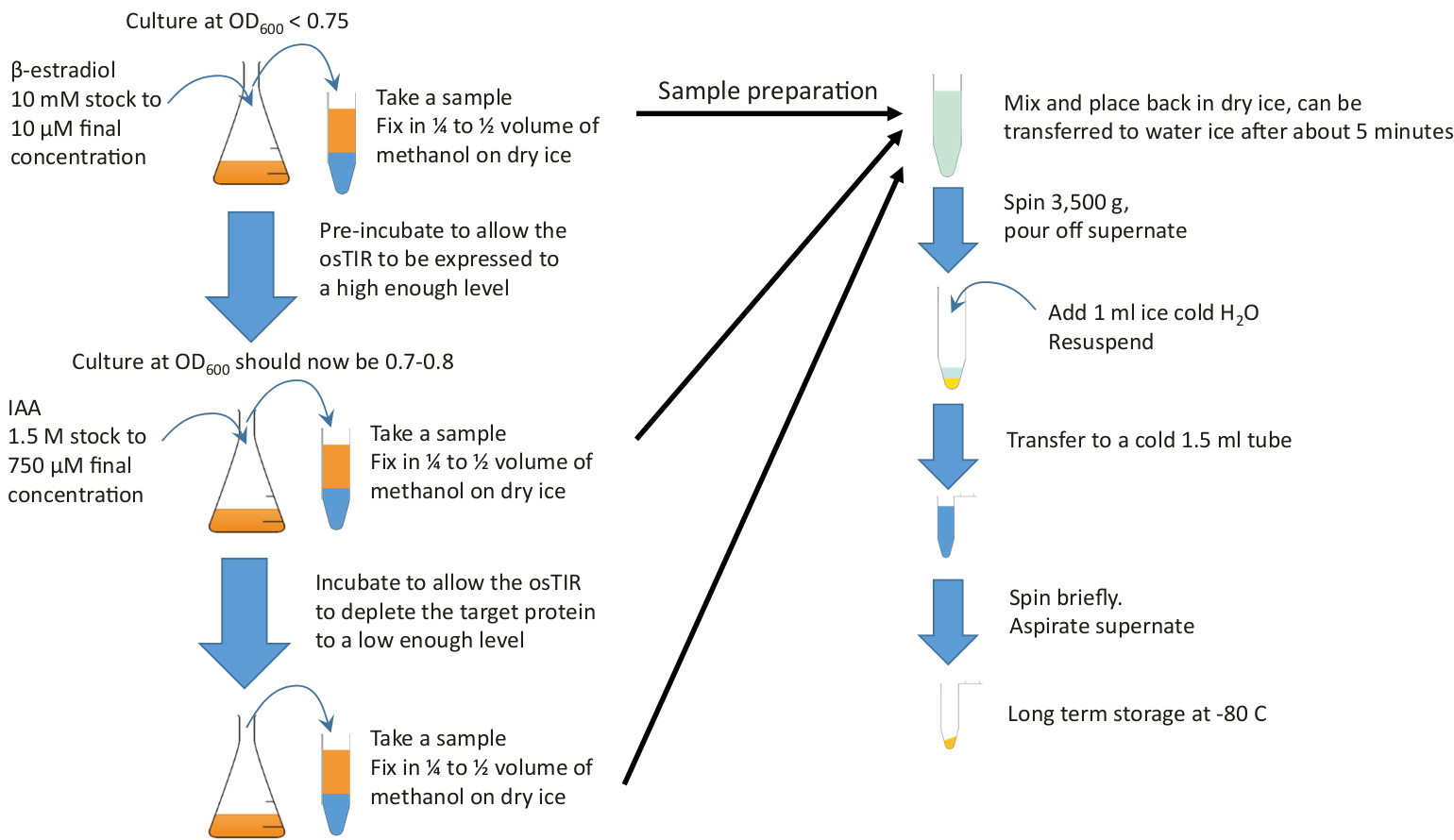
NOTA: Consulte la Figura 2 para obtener un resumen gráfico.
1. Preparación de la tensión
-
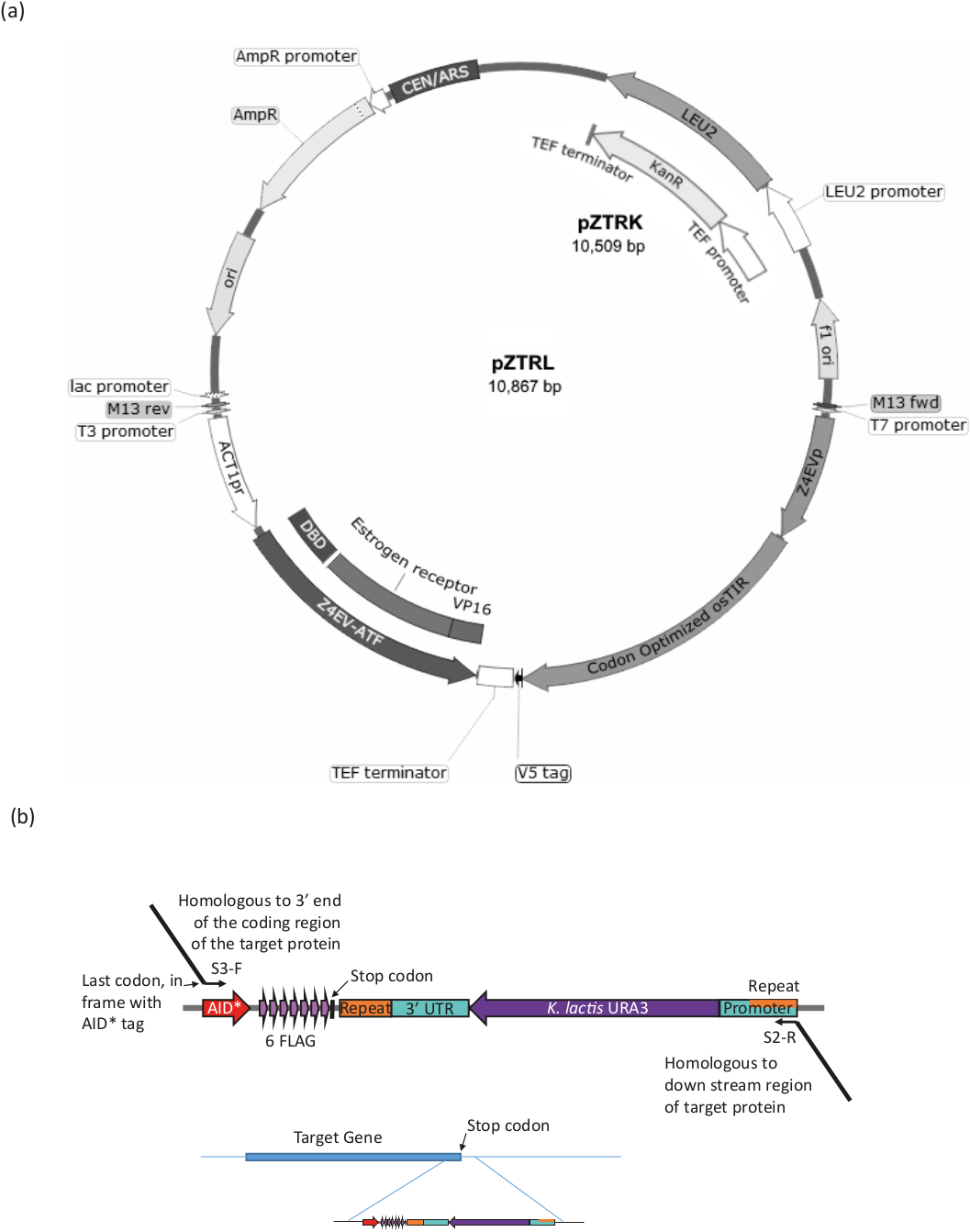
Usando una ura3-strain, introduce el sistema AID (es decir, genes que codifican el factor de transcripción sensible al estradiol (ATF) y el osTIR) y etiqueta AID* la proteína diana (ver Figura 3 y Tabla 1 para un resumen del procedimiento).
- Transformar15 pZTRK (marcador de resistencia G418) o pZTRL (marcadorLEU2) plásmido (disponible en el Centro de Recursos Genéticos de Levadura) en la cepa de levadura ura3- o utilizar el plásmido como plantilla para producir el producto PCR para Integración.
- PCR amplificar el ATF (marcado Z4EVATF en el mapa plásmido) y osTIR utilizando una polimerasa de alta fidelidad de cualquiera de los plásmidos pZTRK o pZTRL. Utilice imprimadores con extensiones de 50 a 60 bases de 3' con homología a la región genómica, para dirigir la integración mediante la recombinación homóloga16. Para la integración genómica de los dos componentes por separado o juntos, véase la Tabla 1 para los cebadores y las condiciones.
NOTA: La cepa pZ4EV-NTR1 tiene los componentes ya integrados en el genoma (disponibles en el Centro de Recursos Genéticos de Levadura, Japón). - Asegúrese de que la proteína diana esté etiquetada con AID* mediante el procedimiento Longtine17 (consulte la figura 3b y la tabla 1).
- Realizar un análisis de crecimiento en la cepa sin é-estradiol e IAA presentes para determinar si la etiqueta AID* afecta al crecimiento y predecir la tasa de crecimiento para su uso en el paso 1.5.
2. Procedimiento General para el Agotamiento
- Calcular la cantidad de cultivo necesario para todas las muestras que se van a recopilar; por ejemplo, 10 ml de cultivo en OD600 de 0,8 es suficiente para la extracción de proteínas, ARN y ADN para una sola muestra, por lo que para 6 muestras, se necesitan al menos 60 ml de cultivo.
- A partir de un cultivo nocturno, establezca suficiente nueva cultura en OD600 0.1 a 0.2 y deje crecer a 30 oC. Se recomienda un medio rico como YPDA, aunque se pueden utilizar otras condiciones de crecimiento:
Extracto de levadura 10 g Peptona 20 g Glucosa 20 g Sulfato de Adenina 40 mg H2O a 1 L
NOTA: Autoclave o filtro esterilizar; la esterilización por filtro se prefiere como complejos de péptidos/azúcar producidos por el precipitado autoclave en el metanol utilizado en la recolección de muestras. - Prepárese para recibir las muestras.
- Poner 30 a 50% del volumen de muestra de metanol previsto en un tubo. Por ejemplo, si se va a tomar una muestra de 10 ml, coloque 5 ml de metanol en un tubo de halcón de 15 ml y cierre el tubo firmemente. Una vez cerrado, etiquete el tubo y colóquelo en hielo seco o a -80 oC para enfriar.
PRECAUCION: Dispensar el metanol en una campana de humo. - Etiquetar tubos de 1,5 ml para el almacenamiento a largo plazo de las muestras y colocar en hielo para enfriar.
- Enfríe lo suficiente H2O (al menos 1 ml por muestra) sobre hielo.
- Poner 30 a 50% del volumen de muestra de metanol previsto en un tubo. Por ejemplo, si se va a tomar una muestra de 10 ml, coloque 5 ml de metanol en un tubo de halcón de 15 ml y cierre el tubo firmemente. Una vez cerrado, etiquete el tubo y colóquelo en hielo seco o a -80 oC para enfriar.
- Anticipar el crecimiento de la cultura. El OD objetivo para la recolección de las muestras es aproximadamente 0,7 a 0,8, pero el paso de pre-incubación (la incubación con el estradiol para inducir el osTIR), debe iniciarse antes para que el cultivo alcance aproximadamente el Dod adecuado en el momento en que las muestras Recogidos.
NOTA: Es aconsejable realizar una curva de crecimiento en las condiciones a utilizar en el experimento para que se pueda estimar esta DO inicial. - Una vez que se haya alcanzado la DO objetivo para el inicio de la preincubación, tome una muestra (normalmente 10 ml), en el tubo pre-preparado que contiene metanol frío. Invierta brevemente para mezclar y volver a colocar en hielo seco.
NOTA: La muestra se puede mover al hielo de agua después de unos 5 minutos, si es conveniente hacerlo. - Añadir inmediatamente el estradiol, 1 L/ml de cultivo (concentración final de 10 m); tener el estradiol pre-medido en una pipeta lista para su uso con el fin de reducir el tiempo que se toma entre la recolección de la muestra y la adición del estradiol. Mezcle rápidamente girando vigorosamente.
- Continuar creciendo el cultivo como antes (paso 2.2), incubar (este es el paso de "pre-incubación") con el estradiol para el tiempo óptimo (para la determinación del tiempo óptimo de pre-incubación ver Figura 1).
- Prepárese para agregar IAA (auxin). Tome el volumen de IAA necesario para el paso 2.10 (es decir, 0,5 l de IAA por ml de cultivo). Esto hace que el paso 2.20 sea más rápido.
- Recoja una muestra como paso 2.5.
- Añadir inmediatamente IAA 0,5 l/ml de cultivo a una concentración final de 750 m según se prepare en el paso 2.8. Mezcle rápidamente girando vigorosamente.
- Recoger muestras, como paso 2.5, de acuerdo con su diseño experimental. Ya sea una sola muestra, en un momento en que se espera que la proteína se agote de forma fiable, o varias muestras en un curso de tiempo de agotamiento. Por ejemplo, los intervalos de 5 minutos son convenientes para el cronometraje y proporcionan una gama de niveles de proteínas. La estrategia de optimización, como se muestra en la Figura1, dará una indicación de los tiempos adecuados.
- Procesar las muestras.
- Coloque las muestras sobre hielo, si no se ha hecho ya. Asegúrese de que ninguna de las muestras se haya congelado; si tienen, suavemente caliente en la mano, invirtiendo constantemente para que la temperatura no suba localmente.
NOTA: Esto se hace mejor en la mano, ya que la temperatura de la muestra se puede evaluar, siempre debe sentirfrío. Colocar en hielo. Esto no es un punto de pausa - una vez que todas las muestras son fluidas, proceda al siguiente paso. - Una vez recogidas todas las muestras y que ya no se congelan, gire a 3.500 x g durante 2 min (a 4 oC si es posible).
- Vierta la mezcla de metanol/medio y vuelva a colocarla en hielo; no se preocupe si no se ha eliminado todo el líquido.
- Resuspenda el pellet celular en 1 ml de hielo frío H2O (a partir del paso 2.3.3) y transfiera a un tubo etiquetado de 1,5 ml (preparado en el paso 2.3.2) sobre hielo.
- Gire brevemente (por ejemplo, 10 s de tiempo total) a >15.000 x g para volver a pelañizar las células, vuelva a colocarlas sobre hielo y retire el líquido.
- Retire el H2O por aspiración. Los pellets de células se pueden almacenar a -20 oC, o -80 oC para el almacenamiento a largo plazo.
- Coloque las muestras sobre hielo, si no se ha hecho ya. Asegúrese de que ninguna de las muestras se haya congelado; si tienen, suavemente caliente en la mano, invirtiendo constantemente para que la temperatura no suba localmente.
- Compruebe el nivel al que se ha agotado la proteína por el análisis de manchas occidentales18.
NOTA: Suficiente proteína19 y/o ácido nucleico se pueden extraer de un solo pellet celular para la mayoría de los propósitos, aunque las especies raras de ARN pueden requerir más volumen de muestra.
Resultados
En la Figura 1se muestran ejemplos representativos de agotamiento. Los tres experimentos presentados en esta figura fueron experimentos de optimización para el agotamiento de las proteínas Prp2, Prp22 y Dcp1. Las proteínas prp2 y Prp22 de baja abundancia, spliceosomales, se agotaron a menos del 20% después de 40 minutos de preincubación con é-estradiol seguido de 15 min con auxina. Los tiempos de preincubación más largos conducen a un agotamiento más rápido, pero también muestran un agotamiento de proteínas indeseable antes de la adición de auxina. En comparación, el Dcp1 más abundante sólo se agotó a aproximadamente el 30% con el mismo tratamiento, pero 60 minutos de pre-incubación resultaron en agotamiento al 13% con el mismo tratamiento de auxina, a costa del agotamiento antes de que se añada la auxina. Es posible que 50 minutos de pre-incubación con el estradiol y 15 min con auxina hubieran logrado resultados similares en un momento más corto y por lo tanto habría nado más óptimo.

Figura 1: La tasa de agotamiento se puede ajustar modulando la duración de la preincubación de estradiol. Manchaoccidental 18 de proteínas diana: (a y b) Prp22-AID*-6FLAG, (c y d) Prp2-AID*-6FLAG, y (e y f) Dcp1-AID*-6HA, de cultivos preincubados con é-estradiol (-est) para 20, 30, 40 o 60 min antes de la adición de auxin12. Se cargaron cantidades iguales de proteína total en cada carril. Pgk1 se detecta como un control de carga visual, excepto el panel e, donde pgk1 y Dcp1 co-migran. La cuantificación de las bandas proteicas en los paneles a , c y e se muestran en los paneles b, dy f, respectivamente. Como medida de la tasa de agotamiento, la pendiente (m) se calculó para la sección lineal (del 100% al 30% de los valores iniciales) de cada curva. El tiempo óptimo de pre-incubación es el momento en el que los niveles de proteína sin dejar de estar cerca de los niveles no inducidos (100%) y la tasa de agotamiento subsiguiente es rápida. Para Dcp1 (f), 60 min de pre-incubación es demasiado largo, ya que la proteína ha comenzado a degradarse en ausencia de auxina, mientras que 20 min es demasiado corto, ya que la proteína no se agota sensiblemente en este curso de tiempo. Después de 40 minutos de pre-incubación, 15 minutos con auxina se puede utilizar ya que la proteína está aproximadamente 70% agotada y, aunque 20 min resultaría en un mayor agotamiento, también podría resultar en efectos secundarios. Las barras de error representan la desviación estándar de dos réplicas biológicas. Para cada experimento, se muestra una mancha representativa. Esta cifra se deriva de la publicación anterior9. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: Resumen gráfico. Añadir el estradiol a un cultivo suficiente que crece en un medio rico y a la temperatura requerida para iniciar la preincubación. Continuar el crecimiento durante el tiempo de pre-incubación requerido antes de agregar IAA (auxin) para iniciar el agotamiento. Los tiempos de preincubación y agotamiento dependen de la proteína a agotar, pero la preincubación suele estar en el rango de 20-60 min y el tiempo de agotamiento suele ser del orden de 10 a 20 min. 10 ml se deben tomar muestras al inicio y al final de la preincubación y la durinidad g el agotamiento. Estas muestras se fijan rápidamente en metanol en frío antes de peletizar y almacenar. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: Generación de tensión para el sistema B-est. (a ) Para generar una cepa de levadura con el sistema AID*, ya sea el pZTRL (LEU2) o el pZTRK (resistencia a la kanamicina (G418), el plásmido debe introducirse en la cepa o, alternativamente, el ATF y el osTIR pueden insertarse en el genoma recombinación homóloga de un fragmento generado por el PCR a partir de imprimaciones de 3 extremos (véase la figura 3b y el cuadro 1). (b) El etiquetado C-terminal de la proteína diana se logra mediante la amplificación pcR de la región apropiada del plásmido pURA3-AID*-6FLAG (pURA3_AID*-6HA difiere sólo en la etiqueta y se puede tratar exactamente de la misma manera), utilizando imprimaciones Longtine S3-F y S2-R con extensiones de 3' homólogas al extremo de 3' de la proteína diana. La extensión de imprimación delantera debe incluir el último codón de aminoácidos en el marco con el inicio de la etiqueta AID* y no debe incluir el codón de parada. La extensión de imprimación inversa debe ser a una región aguas abajo de la región de codificación. Una vez insertadas en el genoma, las células que han perdido el marcador URA3 (por recombinación homóloga entre las regiones idénticas que se encuentran en ambos extremos del marcador) se pueden seleccionar por crecimiento con 5-FOA, que contraselecciona las células URA3. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
| a. Secuencias de primerplano | |||||||
| Objetivo | Ubicación | Cartilla | Nombre | Secuencia | Tm (C) | ||
| pZTRL | 516 | F | pZTRL_F | <-región de homología->GCGACAGCATCACCGACTTCG | 61.23 | ||
| 7897 | R | pZTR_R | CGCCGCCTCTCTCCTTGCAGA<-región de la homología (RC)-> | 61.30 | |||
| pZTRK | 9154 | F | pZTRK_F | <-región de homología->ACGTTGAGCCATTAGTATCAATTTGCTTACC | 59.40 | ||
| 5897 | R | pZTR_R | CGCCGCCTCTCTCCTTGCAGA<-región de la homología (RC)-> | 61.30 | |||
| pURA3-AID*-6FLAG o pURA3_AID*-6HA | F | S3-F | <-región de homología->CGTACGCTGCAGGTCGAC | 59.21 | |||
| R | S2-R | ATCGATGAATTCGAGCTCG<-región de homología (RC)-> | 52.76 | ||||
| pZRTL/K es para amplificar el sistema de AID | |||||||
| pURA3-AID*-6FLAG/6HA para amplificar la etiqueta AID* y epítopo para etiquetar la proteína diana (procedimiento de lontina) | |||||||
| <-región de la homología-> | Región homóloga a las regiones flanqueantes donde se va a insertar el sistema. Cuanto más larga sea esta región, más probable es que la modificación tenga éxito; Se recomiendan 50 - 100 bases. | ||||||
| <-región de la homología (RC)-> | Región homóloga a las regiones de flanqueo donde se va a insertar el sistema, recuerde utilizar el complemento inverso. Como se mencionó anteriormente, cuanto más larga sea esta región, mejor. | ||||||
| Tm (C) | Tm usando el método %GC con 50 mM NaCl | ||||||
| b. Mezcla de PCR | |||||||
| Componente | Volumen (L) | ||||||
| Plantilla | <10 | ||||||
| Zona de influencia DE HF de PHUsion NEB (5x)* | 100 | ||||||
| Primer delantero 100 m | 2.5 | ||||||
| Imprimación inversa 100 m | 2.5 | ||||||
| dNTPs 10 mM cada uno | 10 | ||||||
| H2O | a 500 | ||||||
| * El búfer NEB Phusion GC (5x) también se puede utilizar, pero no se prefiere | |||||||
| Haga esta mezcla, dividida en 10 tubos de mezcla de 50 l cada uno y realice la PCR como Tabla 1 c. | |||||||
| Compruebe que el PCR ha funcionado funcionando con un gel de agarosa | |||||||
| Combine todas las reacciones exitosas en un solo tubo y el etanol se precipite | |||||||
| Transformar la levadura con todo el material producido por el PCR | |||||||
| c. Condiciones de la PCR | |||||||
| Paso | Temperatura (C) | hora | |||||
| Desnaturalización inicial | 98 | 30 s | |||||
| 25-35 Ciclos | Desnaturalizar | 98 | 10 s | ||||
| Recueza | 45–60 | 20 s | |||||
| Extensión | 72 | 30 s/kb | |||||
| Extensión final | 72 | 10 min | |||||
| Mantener | 8 | ||||||
| Anneal a 45oC para el juego de imprimación Lontine (S3-F y S2-R) y 60oC para los imprimadores pZTRL/K | |||||||
| Extender durante 3 minutos para el Lontine PCR y 3 minutos para pZTRL/K | |||||||
Tabla 1: Secuencias de imprimación, mezcla de PCR y condiciones de PCR.
Discusión
Un protocolo bien optimizado puede producir un agotamiento rápido y eficiente de la proteína objetivo. Determinar el tiempo aproximado de pre-incubación con el estradiol es importante, ya que esto aumenta la reproducibilidad del agotamiento, pero se pueden tolerar pequeñas variaciones en el tiempo de pre-incubación. Por otro lado, se debe tener cuidado con el tiempo después de la adición de auxina, ya que el nivel de proteína disminuye muy rápidamente.
Una ventaja de este enfoque es que el agotamiento ajustado se puede lograr mediante diferentes combinaciones de tiempo de pre-incubación con el tiempo de incubación de la IAA y el estradiol. Por ejemplo, si se desea, la proteína diana puede agotarse más lentamente reduciendo el tiempo de preincubación.
El sistema AID ofrece ciertas ventajas sobre los sistemas en los que OsTIR se expresa de forma constitutiva. Por ejemplo, si la proteína diana es esencial para la viabilidad, la expresión regulada de osTIR puede evitar el agotamiento prematuro de la proteína diana. Además, la expresión de osTIR se puede ajustar para adaptarse a la abundancia de la proteína diana y su susceptibilidad a la degradación, y el agotamiento puede ser rápido o lento. Los dos pequeños efectores de moléculas, el estradiol y la auxina, no perturban el metabolismo de la levadura en las condiciones utilizadas aquí, a diferencia de la rapamicina, utilizada en el sistema de anclaje1.
Cabe señalar que el etiquetado de algunas proteínas interrumpe su función, que es un problema con cualquier sistema de agotamiento dirigido. En este caso, una etiqueta N-terminal puede funcionar cuando una etiqueta C-terminal no funciona. Además, no todas las proteínas se agotarán eficientemente; por ejemplo, la etiqueta AID de la proteína diana puede ser inaccesible para la proteína osTIR. Por lo tanto, después del etiquetado AID, cada proteína diana debe ser probada para cualquier efecto de la etiqueta en el crecimiento, y para determinar si el agotamiento es eficaz, antes de que se optimizan los plazos de pre-incubación de é-estradiol y tratamiento de auxina.
Este sistema AID* es muy simple y es compatible con cualquier procedimiento experimental posterior que no implique un mayor crecimiento, como el análisis de proteínas, ADN o ARN o la microscopía. Además, el sistema funciona bien cuando se combina con tiolabelling para purificar el ARN naciente20.
Este sistema proporciona un medio rápido, específico y reproducible de agotar una proteína sin afectar de otro modo el metabolismo de la célula de levadura.
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
Gracias a Jane Reid por iniciar este programa, Barbara Terlouw para el desarrollo, Vahid Aslanzadeh para las construcciones "ura looper" y Susana de Lucas para muchas discusiones útiles. Este trabajo fue apoyado por una beca para GIMO del Consejo Nacional de Ciencia y Tecnología de México (CONACYT) y la Escuela de Ciencias Biológicas de la Universidad de Edimburgo, una beca de doctorado wellcome a IEM [105256] y por Wellcome funding [104648] a JD Beggs . El trabajo en el Wellcome Centre for Cell Biology está respaldado por la financiación básica de Wellcome [092076].
Materiales
| Name | Company | Catalog Number | Comments |
| Adenine sulphate | Formedium | DOC0230 | |
| Agar | Formedium | AGA03 | |
| Β-estradiol | Sigma Aldrich | E2758-1G | 10 mM solution in ethanol. Store at -20 °C |
| DMSO | Alfa Aesar | 42780 | DMSO should be solid at 4 °C |
| Glucose | Fisher Scientific | G/0500/60 | |
| IAA 1H-Indole-3-acetic acid | Across Orgainics | 122150100 | Auxin analogue. 1.5 M in DMSO. The solution will be a russet colour and darken as time goes on; a deep red solution should be discarded and a new one made. Store at -20 °C. |
| Methanol | Fisher Scientific | M/4000/PC17 | CAUTION Toxic and flammable |
| Phusion High-Fidelity DNA Polymerase | NEB | M0530 | |
| Peptone | Formedium | PEP03 | |
| SCSM single drop-out –ura | Formedium | DSCS101 | |
| Yeast Extract | Formedium | YEA03 | |
| Yeast nitrogen base without amino acids with amonium sulphate | Formedium | CYN0410 |
Referencias
- Haruki, H., Nishikawa, J., Laemmli, U. K. The Anchor-Away Technique: Rapid, Conditional Establishment of Yeast Mutant Phenotypes. Molecular Cell. 31, 925-932 (2008).
- Bellí, G., Garí, E., Piedrafita, L., Aldea, M., Herrero, E. An activator/repressor dual system allows tight tetracycline-regulated gene expression in budding yeast. Nucleic Acids Research. 26, 942-947 (1998).
- Alexander, R. D., et al. RiboSys, a high-resolution, quantitative approach to measure the in vivo kinetics of pre-mRNA splicing and 3′-end processing in Saccharomyces cerevisiae. RNA. 16, 2570-2580 (2010).
- Deshaies, R. J., Joazeiro, C. A. P. RING Domain E3 Ubiquitin Ligases. Annual Review of Biochemistry. 78, 399-434 (2009).
- Tan, X., et al. Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature. 446, 640-645 (2007).
- Teale, W. D., Paponov, I. A., Palme, K. Auxin in action: signalling, transport and the control of plant growth and development. Nature Reviews Molecular Cell Biology. 7, 847-859 (2006).
- Nishimura, K., Fukagawa, T., Takisawa, H., Kakimoto, T., Kanemaki, M. An auxin-based degron system for the rapid depletion of proteins in nonplant cells. Nature Methods. 6, 917-922 (2009).
- Morawska, M., Ulrich, H. D. An expanded tool kit for the auxin-inducible degron system in budding yeast. Yeast. 30, 341-351 (2013).
- Kubota, T., Nishimura, K., Kanemaki, M. T., Donaldson, A. D. The Elg1 Replication Factor C-like Complex Functions in PCNA Unloading during DNA Replication. Molecular Cell. 50, 273-280 (2013).
- Brosh, R., et al. A dual molecular analogue tuner for dissecting protein function in mammalian cells. Nature Communications. 7, 11742 (2016).
- DeRisi, J. L., Iyer, V. R., Brown, P. O. Exploring the metabolic and genetic control of gene expression on a genomic scale. Science. 278, 680-686 (1997).
- Mendoza-Ochoa, G. I., et al. A fast and tuneable auxin-inducible degron for depletion of target proteins in budding. Yeast. , (2018).
- McIsaac, R. S., et al. Synthetic gene expression perturbation systems with rapid, tunable, single-gene specificity in yeast. Nucleic Acids Res. 41, e57 (2013).
- Prusty, R., Grisafi, P., Fink, G. R. The plant hormone indoleacetic acid induces invasive growth in Saccharomyces cerevisiae. PNAS. 101, 4153-4157 (2004).
- Geitz, D., St Jean, A., Woods, R. A., Schiest, R. H. Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Research. 20, 1425 (1992).
- Widlund, P. O., Davis, T. N. A high-efficiency method to replace essential genes with mutant alleles in yeast. Yeast. 22, 769-774 (2005).
- Longtine, M. S., et al. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast. 14, 953-961 (1998).
- Eaton, S. L., et al. A Guide to Modern Quantitative Fluorescent Western Blotting with Troubleshooting Strategies. Journal of Visualized Experiments. , e52099 (2014).
- Volland, C., Urban-Grimal, D., Géraud, G., Haguenauer-Tsapis, R. Endocytosis and degradation of the yeast uracil permease under adverse conditions. Journal of Biological Chemistry. 269, 9833-9841 (1994).
- Barrass, J. D., et al. Transcriptome-wide RNA processing kinetics revealed using extremely short 4tU labeling. Genome Biology. 16, 282 (2015).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados