
Method Article
Ottimizzazione della degradazione per ottenere un esaurimento specifico ed efficiente delle proteine
In questo articolo
Riepilogo
Qui, presentiamo un protocollo per esaurire in modo efficace e specifico una proteina di interesse per il lievito Saccharomyces cerevisiae utilizzando il sistema AID di est.
Abstract
Il recettore del legame dell'auxina vegetale, TIR1, riconosce le proteine contenenti uno specifico motivo degron auxina inducibile (AID) in presenza di auxina, mirando a degradarle. Questo sistema è sfruttato in molti eucarioti non vegetali, in modo che una proteina bersaglio, etichettata con il motivo AID, sia degradata sull'aggiunta di auxina. Il livello di espressione TIR1 è critico; un'espressione eccessiva porta alla degradazione della proteina marcata con AID anche in assenza di auxina, mentre la bassa espressione porta a un lento esaurimento. È stato creato un sistema AID inducibile con z-estradiolo, con espressione di TIR1 sotto il controllo di un promotore inducibile di estradiolo. Il livello di TIR1 è regolabile modificando il tempo di incubazione con l'aggiunta di auxl prima dell'auxina. Questo protocollo descrive come esaurire rapidamente una proteina bersaglio utilizzando il sistema AID. L'appropriato tempo di incubazione estradiolo dipende dall'abbondanza della proteina bersaglio. Pertanto, un esaurimento efficiente dipende dalla tempistica ottimale che riduce al minimo l'esaurimento indipendente dall'auxina.
Introduzione
Le mutazioni condizionali, come i mutanti sensibili alla temperatura, sono un potente strumento per lo studio delle proteine essenziali, consentendo la crescita cellulare nella condizione permissiva, ma causando la perdita di funzione in condizioni non permissive. Tuttavia, metabolismo cellulare può essere seriamente turbato dal cambiamento nelle condizioni di crescita necessaria per indurre il difetto e può anche creare effetti off-target. Sono stati sviluppati diversi metodi, in cui la proteina di interesse è sequestrata in modo condizionale1 o la sua espressione è controllata2,3 mediante l'aggiunta di una piccola molecola. Questo protocollo utilizza l'auxina e il sistema di degron auxina (AID) per esaurire in modo efficiente una proteina bersaglio.
Il sistema AID ha origine nelle piante, dove viene utilizzata un'auxina (in questo protocollo indole-3-acetica acid (IAA), stimola l'interazione della proteina Aux/IAA con TIR1, membro del complesso di ligase di ubiquitina SCF U34. L'interazione complessa SCF provoca l'assoluzione poliilogica delle proteine familiari Aux/IAA, che si traduce nella loro degradazione da parte del proteasome5,6.
Questo sistema è stato precedentemente adattato per l'uso nel lievito Saccharomyces cerevisiae7,8 esprimendo la proteina TIR1 di Oriza sativa (osTIR) nelle cellule di lievito, dove è in grado di interagire con il lievito endogeno Complesso SCF. La proteina di interesse è stata etichettata con un motivo della proteina Aux/IAA IAA17 per indirizzarla alla degradazione. I troncazioni funzionali di IAA17 sono stati sviluppati più tardi, come AID8,9,10, contenente il motivo sensibile all'auxina da 43 aminoacidi dell'Arabidopsis thaliana IAA17, insieme a un tag epitope per consentire scoperta.
Il sistema inizialmente adattato per l'uso in lievito in erba7,8 ha espresso la proteina osTIR1 da un promotore GAL di lievito. Espressione richiede il passaggio al mezzo di crescita con galactose come unica fonte di carbonio, che, purtroppo, si traduce in un spostamento diauxico con ampi cambiamenti al metabolismo cellulare11. D'altra parte, è stato riferito che l'espressione costitutiva di TIR1 può portare alla degradazione della proteina bersaglio in assenza di auxin/IAA12 se il livello di espressione è alto, mentre una bassa espressione TIR1 provoca un esaurimento inefficiente. È stato sviluppato un sistema di AID migliorato, denominato AID di z-est, in cui l'osTIR è sotto il controllo di un promotore inducibile che è regolabile per soddisfare la proteina bersaglio, con un effetto minimo sul metabolismo cellulare. Per raggiungere questo obiettivo, è stato costruito un fattore di trascrizione artificiale (ATF) in cui l'attivatore di trascrizione virale VP16 viene fuso a un recettore degli estrogeni e a un dominio di legame del DNA delle dita di quattro zn (DBD). Quando è presente il z-estradiolo (un estrogeno), l'ATF può entrare nel nucleo e indurre la trascrizione osTIR legandosi al suo promotore (4EVpr)13,12.
L'espressione osTIR è solitamente rilevabile circa 20 min dopo l'aggiunta di estradiol12. Tuttavia, la durata ottimale dell'espressione osTIR per ottenere un'efficienza della proteina marcata con auxina, evitando l'esaurimento prima dell'aggiunta di auxina, deve essere determinata empiricamente per ogni proteina bersaglio. Un tempo approssimativo per questa pre-incubazione può essere stimato dai valori di abbondanza nel banca dati del genoma di Saccharomyces (SGD https://www.yeastgenome.org/). Come si può vedere nella Figura1, l'abbondante proteina, Dcp1 (2880 a 4189 molecole/cellule), richiede 40 min di pre-incubazione con zestradiolo, senza alcun esaurimento indipendente dall'auxina osservato. La proteina molto meno abbondante, Prp2 (172 a 211 molecole/cellule), è fortemente esaurita dopo soli 20 min di pre-incubazione. Si consiglia di testare due tempi pre-incubazione aggiuntivi, da 10 a 20 min prima o dopo questo tempo stimato iniziale (20 min è il tempo minimo consigliato). Il tempo ottimale di pre-incubazione è il momento in cui la proteina bersaglio non si è esaurita prima di aggiungere l'auxina e una volta aggiunta l'auxina è accettabile o i livelli di proteine si avvicinano al minimo possibile. Così, dalla figura 1b, per Prp22 con 30 min di pre-incubazione, i livelli non sono diminuiti di molto 10 min dopo l'aggiunta di auxina. Confrontando questo con 40 min di pre-incubazione e 15 min con iAA, dove c'è poco esaurimento aggiuntivo, non vi è alcun beneficio nell'incubazione con auxina più lunga di 10 min o pre-incubazione per più di 30 min, in particolare perché vi sono prove di l'esaurimento dipendente a 40 min. Per Dcp1 con 40 min di pre-incubazione (l'ultimo punto in cui il livello proteico è di circa il 100% prima dell'aggiunta di auxina), 15-20 min di esaurimento con auxina è accettabile. Si raccomanda di mantenere il tempo di esaurimento il più breve possibile per ridurre gli effetti secondari sul metabolismo cellulare14.
In questo articolo viene illustrato come utilizzare il sistema AID di tipo Est, ottimizzando i tempi di incubazione estradiolo per l'espressione osTIR per ottenere un rapido esaurimento delle proteine bersaglio sull'aggiunta di IAA senza esaurirsi prima di aggiungere l'auxina.
Protocollo
NOT: Vedere La Figura 2 per un riepilogo grafico.
1. Preparazione del deformazione
-
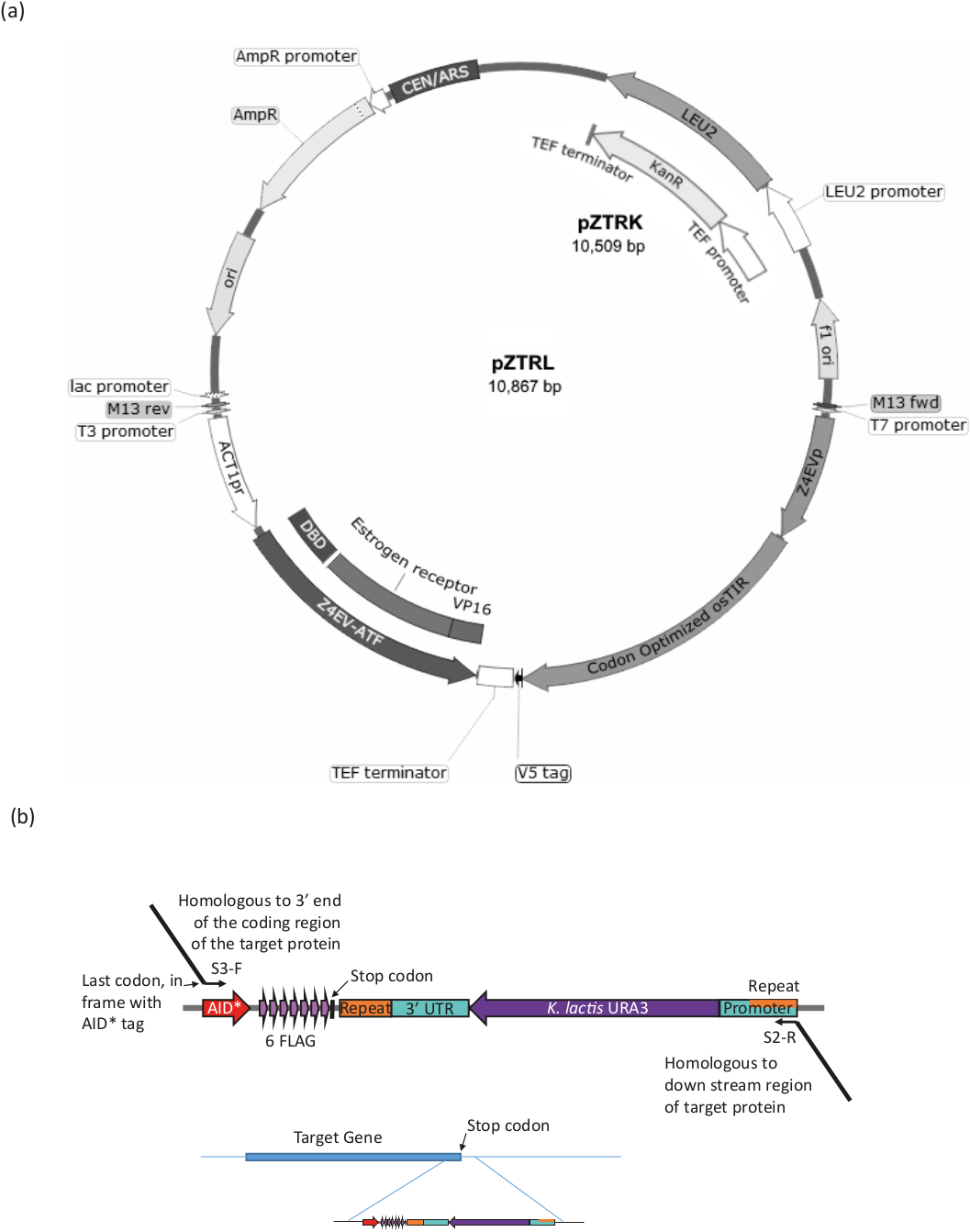
Utilizzando un ura3-strain, introdurre il sistema DI AID (cioè i geni che codificano il fattore di trascrizione reattivo-estradiolo (ATF) e l'osTIR) e l'AID, etichettare la proteina bersaglio (vedere la Figura 3 e la Tabella 1 per un riepilogo della procedura).
- Trasformare il plasmidedi 15 p-TRK (G418) o il plasmide (marcatoreLEU2) (disponibile presso il Lievito Genetic Ogmcomino) nel ceppo di lievito riccio3 o utilizzare il plasmide come modello per produrre il prodotto PCR per la genomica integrazione f.
- La PCR amplifica l'ATF (contrassegnata con la 4EVATF sulla mappa plasmide) e l'osTIR utilizzando una polimerasi ad alta fedeltà da uno dei plasmidi p-TRK o p-TRL. Utilizzare primer con 50 a 60 estensioni in base 3' con omologia alla regione genomica, per l'integrazione diretta con ricombinazione omologa16. Per l'integrazione genomica dei due componenti separatamente o insieme, vedere la tabella 1 per i primer e le condizioni.
NOT: Il ceppo p-4EV-NTR1 ha i componenti già integrati nel genoma (disponibile presso il Lievito Genetic Resource Centre, Giappone). - Assicurarsi che la proteina bersaglio sia etichettata con la procedura Longtine17 (vedere la figura 3b e la tabella 1).
- Eseguire un'analisi della crescita sul ceppo senza la presenza di z-estradiol e IAA per determinare se il tag AID influenzi la crescita e per prevedere il tasso di crescita per l'uso al punto 1,5.
2. Procedura generale per l'esaurimento
- Calcolare la quantità di cultura necessaria per la raccolta di tutti i campioni; ad esempio, 10 mL di coltura a OD600 di 0,8 sono sufficienti per l'estrazione di proteine, RNA e DNA per un singolo campione, quindi per 6 campioni sono necessari almeno 60 mL di coltura.
- Da una cultura notturna, impostare una nuova cultura sufficiente a OD600 0.1 a 0.2 e lasciare crescere a 30 gradi centigradi. Un mezzo ricco come YPDA è raccomandato, anche se altre condizioni di crescita possono essere utilizzati:
Estratto di lievito 10 g Peptone 20 g glucosio 20 g Solfato di Adenina 40 mg H2O per 1 L
NOT: Autoclave o filtro sterilizzare; la sterilizzazione filtrante è preferita come complessi peptidi/zucchero prodotti da precipitazioni autoclaving nel metanolo utilizzato nella raccolta dei campioni. - Prepararsi a ricevere i campioni.
- Mettere dal 30 al 50% del volume del campione previsto di metanolo in un tubo. Ad esempio, se deve essere prelevato un campione da 10 mL, mettere 5 mL di metanolo in un tubo falco da 15 mL e chiudere saldamente il tubo. Una volta chiuso, etichettare il tubo e mettere su ghiaccio secco o a -80 gradi per raffreddare.
ATTENZIONE: Distribuisci il metanolo in una cappa di fumi. - Etichettare i tubi da 1,5 mL per lo stoccaggio a lungo termine dei campioni e metterli nel ghiaccio per raffreddarle.
- Raffreddare abbastanza H2O (almeno 1 mL per campione) sul ghiaccio.
- Mettere dal 30 al 50% del volume del campione previsto di metanolo in un tubo. Ad esempio, se deve essere prelevato un campione da 10 mL, mettere 5 mL di metanolo in un tubo falco da 15 mL e chiudere saldamente il tubo. Una volta chiuso, etichettare il tubo e mettere su ghiaccio secco o a -80 gradi per raffreddare.
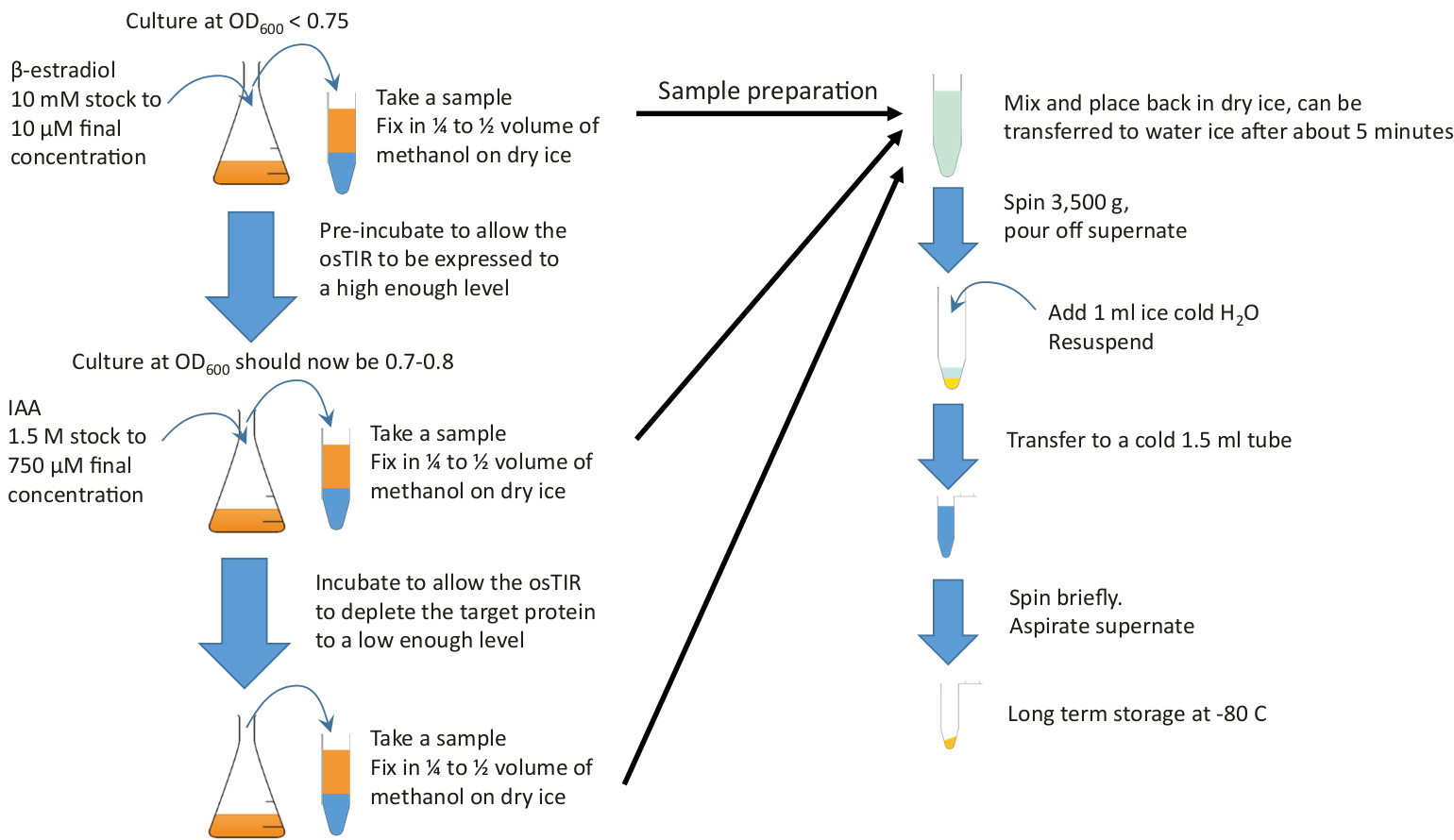
- Anticipare la crescita della cultura. L'OD target per la raccolta dei campioni è di circa 0,7 a 0,8, ma la fase di pre-incubazione (l'incubazione con il -estradiolo per indurre l'osTIR), deve essere avviata prima in modo che la coltura raggiunga approssimativamente l'OD destro nel momento in cui i campioni sono raccolto.
NOT: Si consiglia di eseguire una curva di crescita nelle condizioni da utilizzare nell'esperimento in modo che questo OD iniziale possa essere stimato. - Una volta raggiunto l'OD target per l'inizio della pre-incubazione, preleva un campione (di solito 10 mL), nel tubo pre-preparato contenente metanolo freddo. Invertire brevemente per mescolare e riporre nel ghiaccio secco.
NOT: Il campione può essere spostato sul ghiaccio d'acqua dopo circa 5 min, se conveniente farlo. - Aggiungere immediatamente il valore di z-estradiol, 1 l/mL di coltura (concentrazione finale di 10 M); hanno il modello z-estradiolo pre-misurato in una pipetta pronta per l'uso al fine di ridurre il tempo necessario tra la raccolta del campione e l'aggiunta del file estradiolo. Mescolare rapidamente vorticoso vigorosamente.
- Continuare a far crescere la coltura come prima (passaggio 2.2), incubare (questa è la fase di "pre-incubazione") con il percorso di z-estradiol per il tempo ottimale (per la determinazione del tempo ottimale pre-incubazione vedere Figura 1).
- Prepararsi ad aggiungere IAA (auxin). Riprendi il volume di IAA necessario per il passaggio 2.10 (cioè 0,5 - L di IAA per mL di coltura). Questo rende il passo 2.20 più veloce.
- Raccogliere un campione come passaggio 2.5.Collect a sample as step 2.5.
- Aggiungere immediatamente iAA di 0,5 gradi/mL di coltura ad una concentrazione finale di 750 M, come preparato al punto 2.8. Mescolare rapidamente vorticoso vigorosamente.
- Raccogliere i campioni, come passo 2.5, secondo il vostro progetto sperimentale. O un singolo campione, in un momento in cui si prevede che la proteina sarà esaurita in modo affidabile, o più campioni in un ciclo temporale di esaurimento. Ad esempio, intervalli di 5 min sono convenienti per la temporizzazione e forniscono una gamma di livelli di proteine. La strategia di ottimizzazione, come illustrato nella Figura 1, darà un'indicazione dei tempi adatti.
- Elaborare i campioni.
- Mettere i campioni sul ghiaccio, se non già fatto. Assicurarsi che nessuno dei campioni sia congelato; se hanno, delicatamente caldo in mano, invertendo costantemente in modo che la temperatura non aumenti localmente.
NOT: Questo è meglio fare in mano come la temperatura del campione può essere valutato, dovrebbe sempre sentire freddo. Mettere sul ghiaccio. Questo non è un punto di pausa - una volta che tutti i campioni sono fluidi, procedere al passaggio successivo. - Una volta che tutti i campioni sono stati raccolti e non sono più congelati, girare a 3.500 x g per 2 min (se possibile a 4 gradi centigradi).
- Versare il mix metanolo/medio e riporre il ghiaccio; non preoccuparti se non tutto il liquido è stato rimosso.
- Risospendere il pellet cellulare in 1 mL di ghiaccio freddo H2O (dal passo 2.3.3) e trasferirlo in un tubo etichettato da 1,5 mL (preparato al punto 2.3.2) sul ghiaccio.
- Girare brevemente (ad esempio, 10 s tempo totale) a >15,000 x g per re-pellet le cellule, riposizionare sul ghiaccio e rimuovere il liquido.
- Rimuovere H2O per aspirazione. I pellet cellulari possono essere immagazzinati a -20 gradi centigradi, o -80 gradi centigradi per lo stoccaggio a lungo termine.
- Mettere i campioni sul ghiaccio, se non già fatto. Assicurarsi che nessuno dei campioni sia congelato; se hanno, delicatamente caldo in mano, invertendo costantemente in modo che la temperatura non aumenti localmente.
- Controllare il livello al quale la proteina è stata esaurita dall'analisi macchia occidentale18.
NOT: Una proteina19 e/o acido nucleico può essere estratta da un pellet a singola cellula per la maggior parte degli scopi, anche se le specie di RNA rare potrebbero richiedere più volume campione.
Risultati
Esempi rappresentativi di esaurimento sono visualizzati nella figura 1. I tre esperimenti presentati in questa figura sono stati esperimenti di ottimizzazione per l'esaurimento delle proteine Prp2, Prp22 e Dcp1. La bassa abbondanza, le proteine silliceosomic e Prp22 entrambe si esaurivano a meno del 20% dopo 40 min pre-incubazione con z-estradiol seguito da 15 min con auxina. Tempi di pre-incubazione più lunghi portano ad un esaurimento più rapido, ma mostrano anche un esaurimento indesiderato delle proteine prima dell'aggiunta di auxina. In confronto, il più abbondante Dcp1 è stato esaurito solo a circa il 30% con lo stesso trattamento, ma 60 min di pre-incubazione ha portato all'esaurimento al 13% con lo stesso trattamento con auxina, a costo di esaurimento prima che venga aggiunta l'auxina. È possibile che 50 min di pre-incubazione con z-estradiol e 15 min con auxina avrebbero ottenuto risultati simili in un momento più breve e quindi sarebbe stato più ottimale.

Figura 1: Il tasso di esaurimento può essere sintonizzato modulando la durata dell'incubazione pre-estradiole. Blotoccidentale 18 delle proteine bersaglio: (a e b) Prp22-AID-6FLAG, (c e d) Prp2-AID-6FLAG, e (e e f) Dcp1-AID-6HA, da colture pre-incubate con zestradiolo ( 60 min prima dell'aggiunta di auxina12. Pari quantità di proteine totali sono state caricate in ogni corsia. Pgk1 viene rilevato come controllo di caricamento visivo, ad eccezione del pannello e, dove pgk1 e Dcp1 co-migrate. La quantificazione delle bande proteiche nei pannelli a, c ed e sono mostrate rispettivamente nei pannelli b, de f. Come misura del tasso di esaurimento, la pendenza (m) è stata calcolata per la sezione lineare (dal 100% al 30% dei valori iniziali) di ogni curva. Il tempo ottimale di pre-incubazione è il momento in cui i livelli proteici sono ancora vicini ai livelli non indotti (100%) e il successivo tasso di esaurimento è veloce. Per Dcp1 (f), 60 min di pre-incubazione è troppo lungo, poiché la proteina ha cominciato a degradarsi in assenza di auxina, mentre 20 min è troppo breve, in quanto la proteina non si esaurisce sensibilmente in questo corso di tempo. Dopo 40 min pre-incubazione, 15 min con auxina può essere utilizzato come la proteina è circa 70% impoverito e, anche se 20 min si tradurrebbe in un ulteriore esaurimento, potrebbe anche provocare effetti secondari. Le barre di errore rappresentano la deviazione standard di due repliche biologiche. Per ogni esperimento, viene mostrata una macchia rappresentativa. Questa cifra è derivata dalla precedente pubblicazione9. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Riepilogo grafico. Aggiungere lo z-estradiol a una cultura sufficiente crescendo nel mezzo ricco e alla temperatura richiesta al fine di iniziare l'incubazione pre-incubazione. Continuare la crescita per il tempo pre-incubazione richiesto prima di aggiungere IAA (auxin) per iniziare l'esaurimento. I tempi di pre-incubazione e di esaurimento dipendono dalla proteina da impoverire, ma la pre-incubazione è spesso compresa tra 20 e 60 min e il tempo di esaurimento è generalmente nell'ordine di 10-20 min. 10 mL devono essere prelevati all'inizio e alla fine della pre-incubazione e della durata g l'esaurimento. Questi campioni vengono rapidamente fissati nel metanolo freddo prima della pellettura e dell'immagazzinamento. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: Generazione di deformazione per il sistema B-est. (a) Per generare un ceppo di lievito con il sistema AID, sia il p-TRL (LEU2) o la resistenza p-TRK (kanamycin (G418), plasmide dovrebbe essere introdotto nel ceppo o, in alternativa, l'ATF e l'osTIR possono essere inseriti nel genoma da ricombinazione omologa di un frammento generato dalla PCR da primer 3'end (vedere la figura 3b e la tabella 1). (b) L'etichettatura del terminale C della proteina bersaglio è ottenuta dall'amplificazione PCR della regione appropriata del pLAsmide pURA3-AID-6FLAG (pURA3_AID-6HA differisce solo nel tag e può essere trattata esattamente allo stesso modo), utilizzando i primer longtine S3-F e S2-R con 3' estensioni omologhe all'estremità 3' della proteina bersaglio. L'estensione del primer in avanti deve includere l'ultimo codone aminoacido nel telaio con l'inizio del tag AID e non deve includere il codone di arresto. L'estensione del primer inverso deve essere in un'area a valle dell'area di codifica. Una volta inserite nel genoma, le cellule che hanno perso il marcatore URA3 (mediante ricombinazione omologa tra le regioni identiche trovate a entrambe le estremità del marcatore) possono essere selezionate dalla crescita con 5-FOA, che controseleziona le cellule URA3. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
| a. Sequenze Primer | |||||||
| bersaglio | posizione | testo elementare | nome | sequenza | Tm (C) | ||
| P-RATL (T. TRL) | 516 del sistema | F | F (in modo incui in base all'utilizzo di p | GCGACAGCATCACCGCTACTTG | 61.23 | ||
| 7897 | R | P.TR_R | CGCCGCCTCTCTCTTGCAGA | 61.30 | |||
| il nuovo p-TRK | 9154 | F | P-TRK_F | ACGTTGAGCCATTAGTATCAATCATTGCTTACC | Ore 59.40 | ||
| 5897 | R | P.TR_R | CGCCGCCTCTCTCTTGCAGA | 61.30 | |||
| pURA3-AID-6FLAG o pURA3_AID-6HA | F | S3-F | CGTACGCTGCAGGTCGAC | 59.21 | |||
| R | S2-R | ATCGATGAATTCGAGCTCG | 52,76 | ||||
| l'operazione di amplificare il sistema AID | |||||||
| pURA3-AID-6FLAG/6HA per amplificare l'AID e il tag epitope per etichettare la proteina bersaglio (procedura Lontine) | |||||||
| <-regione di omologia-> | Regione omologa alle regioni di fianco in cui deve essere inserito il sistema. Più a lungo questa regione è più probabile che la modifica abbia successo; 50 - 100 basi è raccomandato. | ||||||
| Regione omologa alle regioni di fianco in cui il sistema deve essere inserito, ricordarsi di utilizzare il complemento inverso. Come sopra, più a lungo questa regione è migliore. | |||||||
| Tm (C) | Tm utilizzando il metodo %GC con 50 mM NaCl | ||||||
| b. Miscela PCR | |||||||
| componente | Volume (L) | ||||||
| sagoma | <10 | ||||||
| NeB Phusion HF Buffer (5x) | 100 del sistema | ||||||
| Primer in avanti 100 M | 2.5 24,5 | ||||||
| Primer inverso 100 M | 2.5 24,5 | ||||||
| dNTPs 10 mM ciascuno | 10 del sistema | ||||||
| H2O | fino a 500 | ||||||
| Il NEB Phusion GC Buffer (5x) può anche essere utilizzato, ma non è preferito | |||||||
| Fare questa miscela, divisa in 10 tubi da 50, mescolati ciascuno ed eseguire la PCR come Tabella 1 c. | |||||||
| Controllare che la PCR abbia funzionato correndo su un gel di agarose | |||||||
| Combinare tutte le reazioni riuscite in un unico tubo e precipitato di etanolo | |||||||
| Trasformare il lievito con tutto il materiale prodotto dalla PCR | |||||||
| c. Condizioni PCR | |||||||
| passo | Temp (C) | ora | |||||
| Denaturazione iniziale | 98 (di base) | 30 s | |||||
| 25-35 cicli | Denatura | 98 (di base) | 10 s | ||||
| ricuocere | 45–60 anni | 20 s | |||||
| interno | 72 del 179 | 30 s/kb | |||||
| Estensione finale | 72 del 179 | 10 min | |||||
| tenere | 8 (IN vio | ||||||
| Anneal a 45 gradi centigradi per il set di primer Lontine (S3-F e S2-R) e 60 s per i primer p-TRL/K | |||||||
| Estendere per 3 minuti per la PCR Lontine e 3 minuti per p-TRL/K | |||||||
Tabella 1: sequenze Primer, mix PCR e condizioni PCR.
Discussione
Un protocollo ben ottimizzato può produrre un rapido ed efficiente esaurimento della proteina bersaglio. Determinare il tempo approssimativo pre-incubazione con il periodo di pre-incubazione è importante, in quanto ciò aumenta la riproducibilità dell'esaurimento, ma possono essere tollerate piccole variazioni nel tempo di pre-incubazione. D'altra parte, la cura deve essere presa con tempi dopo l'aggiunta di auxina, come il livello di proteine diminuisce molto rapidamente.
Un vantaggio di questo approccio è che l'esaurimento sintonizzato può essere ottenuto mediante combinazioni variabili di tempo di pre-incubazione con tempo di incubazione di z-estradiolo e IAA. Ad esempio, se lo si desidera, la proteina bersaglio può essere più lentamente esaurita riducendo il tempo di pre-incubazione.
Il sistema AID di est offre alcuni vantaggi rispetto ai sistemi in cui OsTIR è espresso in modo imposizione. Ad esempio, se la proteina bersaglio è essenziale per la vitalità, l'espressione regolata di osTIR può evitare l'esaurimento prematuro della proteina bersaglio. Inoltre, l'espressione di osTIR può essere regolata per soddisfare l'abbondanza della proteina bersaglio e la sua suscettibilità alla degradazione, e l'esaurimento può essere veloce o lento. I due piccoli effetti molecolari, estradiolo e auxina, non pertturbiscono il metabolismo del lievito nelle condizioni utilizzate qui, a differenza della rapamicina, utilizzata nel sistema di ancoraggio1.
Va notato che l'etichettatura di alcune proteine interrompe la loro funzione, che è un problema con qualsiasi sistema di deplezione mirato. In questo caso, un tag N-terminal può funzionare quando un tag C-terminal non funziona. Inoltre, non tutte le proteine saranno esaurite in modo efficiente; ad esempio, l'etichetta AID sulla proteina bersaglio potrebbe essere inaccessibile alla proteina osTIR. Pertanto, dopo l'etichettatura DELL'AID, ogni proteina bersaglio deve essere testata per qualsiasi effetto del tag sulla crescita e per determinare se l'esaurimento è efficace, prima che i tempi di pre-incubazione e trattamento dell'auxina siano ottimizzati.
Questo sistema AID è molto semplice ed è compatibile con qualsiasi procedura sperimentale successiva che non comporti ulteriore crescita, come l'analisi di proteine, DNA o RNA o microscopia. Inoltre, il sistema funziona bene se combinato con la tioetichettaling per purificare l'RNA nascente20.
Questo sistema fornisce un mezzo rapido, specifico e riproducibile per esaurire una proteina senza influenzare in altro modo il metabolismo della cellula di lievito.
Divulgazioni
Gli autori non hanno nulla da rivelare.
Riconoscimenti
Grazie a Jane Reid per aver dato l'avvio di questo programma, Barbara Terlouw per lo sviluppo, Vahid Aslanzadeh per i costrutti "ura looper" e Susana de Lucas per molte discussioni utili. Questo lavoro è stato sostenuto da una borsa di studio a GIMO dal Consejo Nacional de Ciencia y Tecnologa, Messico (CONACYT) e dalla Scuola di Scienze Biologiche dell'Università di Edimburgo, un dottorato di ricerca Wellcome a IEM [105256] e da Wellcome finanziamento [104648] a JD Beggs . Il lavoro nel Wellcome Centre for Cell Biology è supportato dal finanziamento del nucleo del Wellcome [092076].
Materiali
| Name | Company | Catalog Number | Comments |
| Adenine sulphate | Formedium | DOC0230 | |
| Agar | Formedium | AGA03 | |
| Β-estradiol | Sigma Aldrich | E2758-1G | 10 mM solution in ethanol. Store at -20 °C |
| DMSO | Alfa Aesar | 42780 | DMSO should be solid at 4 °C |
| Glucose | Fisher Scientific | G/0500/60 | |
| IAA 1H-Indole-3-acetic acid | Across Orgainics | 122150100 | Auxin analogue. 1.5 M in DMSO. The solution will be a russet colour and darken as time goes on; a deep red solution should be discarded and a new one made. Store at -20 °C. |
| Methanol | Fisher Scientific | M/4000/PC17 | CAUTION Toxic and flammable |
| Phusion High-Fidelity DNA Polymerase | NEB | M0530 | |
| Peptone | Formedium | PEP03 | |
| SCSM single drop-out –ura | Formedium | DSCS101 | |
| Yeast Extract | Formedium | YEA03 | |
| Yeast nitrogen base without amino acids with amonium sulphate | Formedium | CYN0410 |
Riferimenti
- Haruki, H., Nishikawa, J., Laemmli, U. K. The Anchor-Away Technique: Rapid, Conditional Establishment of Yeast Mutant Phenotypes. Molecular Cell. 31, 925-932 (2008).
- Bellí, G., Garí, E., Piedrafita, L., Aldea, M., Herrero, E. An activator/repressor dual system allows tight tetracycline-regulated gene expression in budding yeast. Nucleic Acids Research. 26, 942-947 (1998).
- Alexander, R. D., et al. RiboSys, a high-resolution, quantitative approach to measure the in vivo kinetics of pre-mRNA splicing and 3′-end processing in Saccharomyces cerevisiae. RNA. 16, 2570-2580 (2010).
- Deshaies, R. J., Joazeiro, C. A. P. RING Domain E3 Ubiquitin Ligases. Annual Review of Biochemistry. 78, 399-434 (2009).
- Tan, X., et al. Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature. 446, 640-645 (2007).
- Teale, W. D., Paponov, I. A., Palme, K. Auxin in action: signalling, transport and the control of plant growth and development. Nature Reviews Molecular Cell Biology. 7, 847-859 (2006).
- Nishimura, K., Fukagawa, T., Takisawa, H., Kakimoto, T., Kanemaki, M. An auxin-based degron system for the rapid depletion of proteins in nonplant cells. Nature Methods. 6, 917-922 (2009).
- Morawska, M., Ulrich, H. D. An expanded tool kit for the auxin-inducible degron system in budding yeast. Yeast. 30, 341-351 (2013).
- Kubota, T., Nishimura, K., Kanemaki, M. T., Donaldson, A. D. The Elg1 Replication Factor C-like Complex Functions in PCNA Unloading during DNA Replication. Molecular Cell. 50, 273-280 (2013).
- Brosh, R., et al. A dual molecular analogue tuner for dissecting protein function in mammalian cells. Nature Communications. 7, 11742 (2016).
- DeRisi, J. L., Iyer, V. R., Brown, P. O. Exploring the metabolic and genetic control of gene expression on a genomic scale. Science. 278, 680-686 (1997).
- Mendoza-Ochoa, G. I., et al. A fast and tuneable auxin-inducible degron for depletion of target proteins in budding. Yeast. , (2018).
- McIsaac, R. S., et al. Synthetic gene expression perturbation systems with rapid, tunable, single-gene specificity in yeast. Nucleic Acids Res. 41, e57 (2013).
- Prusty, R., Grisafi, P., Fink, G. R. The plant hormone indoleacetic acid induces invasive growth in Saccharomyces cerevisiae. PNAS. 101, 4153-4157 (2004).
- Geitz, D., St Jean, A., Woods, R. A., Schiest, R. H. Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Research. 20, 1425 (1992).
- Widlund, P. O., Davis, T. N. A high-efficiency method to replace essential genes with mutant alleles in yeast. Yeast. 22, 769-774 (2005).
- Longtine, M. S., et al. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast. 14, 953-961 (1998).
- Eaton, S. L., et al. A Guide to Modern Quantitative Fluorescent Western Blotting with Troubleshooting Strategies. Journal of Visualized Experiments. , e52099 (2014).
- Volland, C., Urban-Grimal, D., Géraud, G., Haguenauer-Tsapis, R. Endocytosis and degradation of the yeast uracil permease under adverse conditions. Journal of Biological Chemistry. 269, 9833-9841 (1994).
- Barrass, J. D., et al. Transcriptome-wide RNA processing kinetics revealed using extremely short 4tU labeling. Genome Biology. 16, 282 (2015).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati