
Method Article
Belirli ve verimli protein tükenmesi elde etmek için bozulma ayarlama
Bu Makalede
Özet
Burada, biz etkin bir protokol sunmak ve özellikle β-EST yardım sistemi kullanarak Maya Saccharomyces cerevisiae ilgi bir protein tüketmek.
Özet
Bitki oksin vardır bağlayıcı reseptör, TIR1, belirli bir oksin vardır indüklenebilir degron içeren proteinleri tanır (AID) oksin vardır varlığında motif, bozulma için onları hedefleyen. Bu sistem birçok non-Plant eukaryotes, böyle bir hedef protein, AID motif ile etiketlenmiş, oksin vardır ilavesi üzerine düşürülmüş olarak sömürülebilir. Düzey TIR1 ifade önemlidir; aşırı ifade, auxin yokluğunda bile AID etiketli protein bozulması yol açar, düşük ifade yavaş tükenme yol açar ise. Bir β-estradiol indüklenebilir yardım sistemi oluşturuldu, TIR1 ifadesinin altında, bir β-erradiol indüklenebilir promotör kontrolü. TIR1 seviyesi, oksin vardır ilavesi öncesinde β-estradiol ile inkübasyon süresini değiştirerek ayarlanabilir. Bu protokol, AıD sistemini kullanarak bir hedef proteini nasıl hızla bozacağını açıklar. Uygun β-estradiol inkübasyon süresi hedef protein bolluğuna bağlıdır. Bu nedenle, verimli tükenmesi de auxin bağımsız tükenmesi en aza indirir optimum zamanlamaya bağlıdır.
Giriş
Sıcaklığa duyarlı mutantlar gibi koşullu mutasyonlar, esansiyel proteinlerin çalışması için güçlü bir araçtır, buna izin veren durum altında hücre büyümesini sağlar, ancak karşı olmayan koşullar altında fonksiyon kaybına neden olur. Ancak, hücre metabolizması ciddi kusur teşvik etmek için gerekli büyüme koşullarında değişiklik tarafından tedirgin olabilir ve ayrıca off-hedef etkileri oluşturabilirsiniz. Çeşitli yöntemler geliştirilmiştir, hangi faiz proteini koşullu olarak1 veya onun ifadesi küçük bir molekül ilavesi ile2,3 kontrol edilir. Bu protokol, oksin vardır ve oksin vardır-indüklenebilir degron (AID) sistemini verimli bir şekilde hedef proteini tüketmek kullanır.
Yardım sistemi, bir oksin vardır (Bu protokol indole-3-asetik asit (IAA) kullanılan) bitkiler, kökeni vardır, TIR1 ile AUX/IAA protein etkileşimi uyarır, SCF U3 Ubiquitin ligaz kompleksi4bir üyesi. SCF kompleks etkileşimi, proteasom5,6tarafından bozulması ile sonuçlanır AUX/IAA aile proteinlerinin polyubiquitination neden olur.
Bu sistem daha önce Maya Saccharomyces cerevisiae7,8 ile TIR1 protein ifade ederek kullanılmak üzere adapte edildi oriza sativa (ostir) Maya hücrelerinde, endojen Maya ile etkileşim mümkün olduğu SCF kompleksi. İlgi protein AUX/ıAA protein IAA17 bir motif ile etiketlendi bozulması için hedef. IAA17 fonksiyonel truncations daha sonra, örneğin Yardım *8,9,10, Arabidopsis thaliana IAA17 gelen 43 amino asit auxin duyarlı motif içeren geliştirilmiştir, bir epitopu etiketi ile birlikte etkinleştirmek için Algılama.
Sistem ilk olarak tomurcuklanan Maya7,8 bir Maya GAL Promoter osTIR1 protein ifade kullanılmak üzere uyarlanmış. İfade, ne yazık ki, hücre metabolizması11geniş değişen bir diauxic vardiya sonuçları tek karbon kaynağı olarak galaktoz ile büyüme orta kayması gerektirir. Öte yandan, TIR1 'in kurucu ifadesinin auxin/ıAA12 yokluğunda hedef proteinin bozulmasına yol açmasının, ifade düzeyi yüksek Ise düşük TIR1 ifadesi verimsiz bir şekilde tükenmesine neden olduğu bildirilmiştir. Β-EST AıD adlı geliştirilmiş bir yardım sistemi, osTIR 'in, hücre metabolizmasında minimal etkiye sahip, hedef proteine uyacak şekilde ayarlanabilir bir indüklenebilir promotör kontrolü altında olduğu geliştirilmiştir. Bunu başarmak için, bir yapay transkripsiyon faktörü (ATF) hangi VP16 Viral transkripsiyon aktivatörü östrojen reseptör ve dört Zn parmaklar DNA bağlayıcı etkı (DBD) için erimiş inşa edilmiştir. Β-estradiol (östrojene) mevcutken ATF, çekirdeğe girebilir ve ostir transkripsiyonu, promotör (Z4EVpr)13,12' ye bağlayabilirler.
osTIR ifadesi genellikle β-estradiol12eklendikten sonra yaklaşık 20 dakika algılanabilir. Ancak, ostir ifadesinin optimum süresi oksin vardır ile etiketlenmiş protein verimli tükenmesi elde etmek için, oksin vardır ilavesi önce tükenmesi kaçınarak, her hedef protein için ampirik olarak belirlenmelidir. Bu pre-inkübasyon için yaklaşık bir süre Saccharomyces Genome veritabanı (SGD https://www.yeastgenome.org/) bolluk değerlerden tahmin edilebilir. Şekil 1' de görülebilir gibi, bol protein, Dcp1 (2880 için 4189 moleküller/hücre), gerektirir 40 dk ön-inkübasyon β-estradiol ile, hiçbir auxin bağımsız tükenmesi gözlenen. Çok daha az bol protein, Prp2 (172 için 211 moleküller/hücre), güçlü bir şekilde sadece 20 dakika pre-inkübasyon sonra tükenmiş. İki ek pre-inkübasyon süreleri test etmek için tavsiye edilir, 10 için 20 dakika önce veya sonra bu ilk tahmini süre (20 dakika önerilen minimum zaman). Optimum pre-inkübasyon süresi, oksin vardır eklemeden önce hedef protein tükenmez ve bir kez oksin vardır, tükenmesi kabul edilebilir veya protein seviyeleri mümkün olan en az yaklaşım eklenir zaman. Yani, Şekil 1B, Prp22 için pre-inkübasyon 30 dakika ile, düzeyleri oksin vardır ek sonra çok 10 dk reddetti değil. Bu ile karşılaştırıldığında 40 dk pre-inkübasyon ve 15 dk ile IAA, orada az ek tükenmesi, daha uzun oksin vardır ile kuluçşuradaki hiçbir yararı yoktur 10 dakika veya ön-daha uzun 30 dakika, özellikle orada olmayan-oksin vardır kanıtı 40 dk 'da bağımlı tükenme. 40 dk pre-inkübasyon ile Dcp1 için (protein seviyesinin yaklaşık% 100 olan son nokta oksin vardır ilavesi öncesinde), 15 ila 20 dk oksin vardır ile tükenmesi kabul edilebilir. Hücre metabolizmasında ikincil etkileri azaltmak için mümkün olduğunca kısa tükenmesi zaman tutmak için tavsiye edilir14.
Bu makalede, auxin eklemeden önce tükenmesi olmadan ıAA ek üzerine hızlı hedef protein tükenmesi elde etmek için osTIR ifadesi için β-estradiol inkübasyon zamanlaması optimize ederek β-EST yardım sisteminin nasıl kullanılacağı gösterilmektedir.
Protokol
Not: Bkz. Şekil 2 grafik Özeti için.
1. gerinim hazırlığı
-
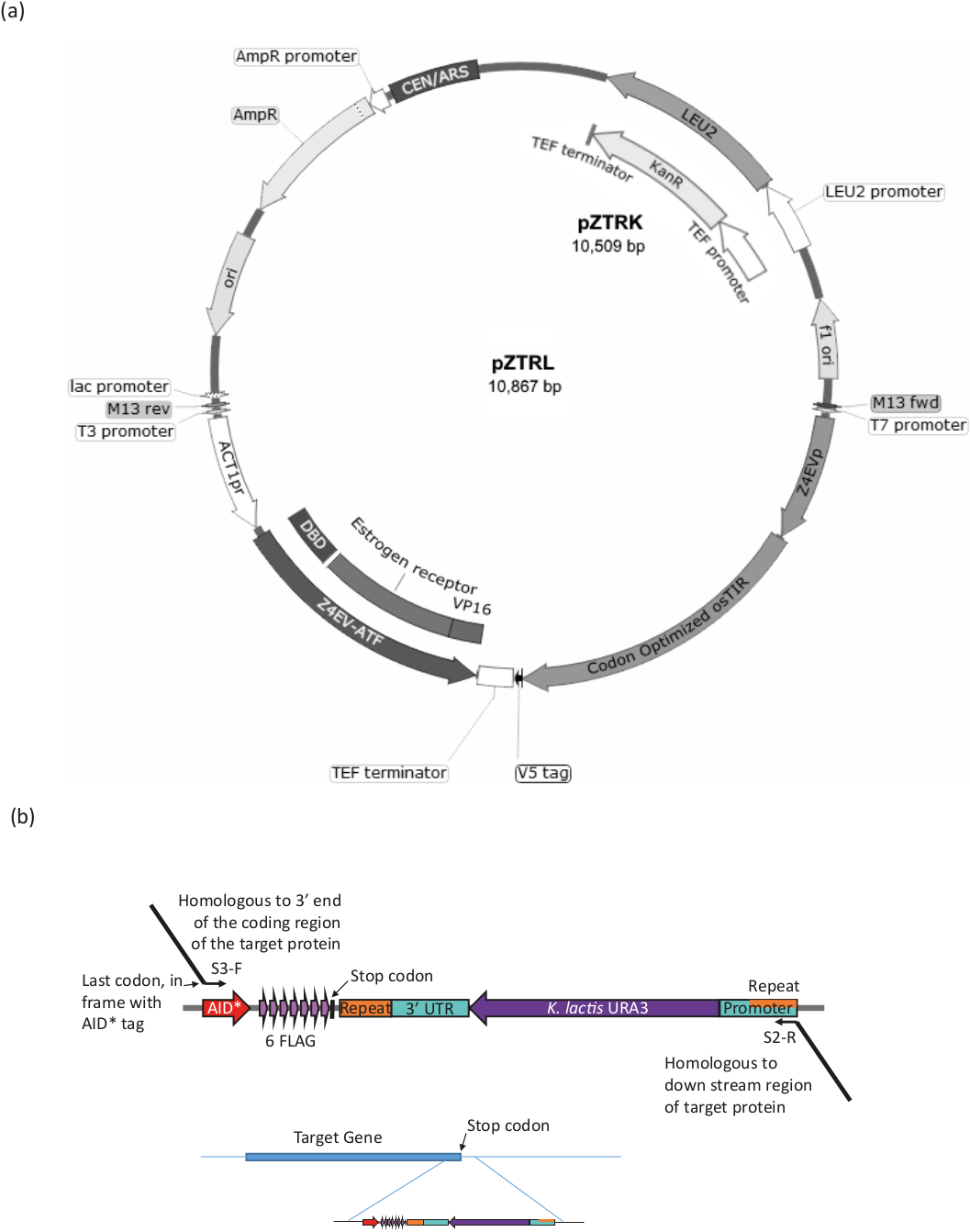
Bir ura3-strain kullanarak, β-EST yardım sistemi tanıtmak (yani, genler β-estradiol duyarlı transkripsiyon faktörü (ATF) ve ostir kodlaması) ve AID * etiket hedef protein (bkz Şekil 3 ve tablo 1 prosedürün bir özeti için).
- Dönüşüm15 ya pZTRK (G418 direnç Marker) veya pZTRL (LEU2 Marker) plazmid (Maya genetik kaynak merkezi 'nden kullanılabilir) ura3- Maya gerinim içine veya PLASMID genomik için PCR ürün üretmek için bir şablon olarak kullanın Entegrasyon.
- PCR ATF (Plasmid harita üzerinde işaretlenmiş Z4EVATF) ve osTIR plazmids pZTRK veya pZTRL ya bir yüksek sadakat polimeraz kullanarak güçlendirin. 50 ile astarlar kullanarak 60 baz 3 ' genomik bölgeye Homoloji ile uzatma, Homolog rekombinasyon ile doğrudan entegrasyon16. İki bileşenin ayrı veya birlikte genomik entegrasyonu için, astar ve koşullar için Tablo 1 ' i görün.
Not: Gerinim pZ4EV-NTR1, genomda entegre edilmiş bileşenlere sahiptir (Maya genetik Kaynak Merkezi, Japonya 'dan edinilebilir). - Hedef proteinin AıD * Longtine prosedürü17 (bkz. Şekil 3B ve Tablo 1) kullanılarak etiketlenmiş olduğundan emin olun.
- Yardım * etiketi büyüme etkiler ve adım 1,5 kullanım için büyüme oranını tahmin belirlemek için β-estradiol ve ıAA mevcut olmadan gerinim üzerinde bir büyüme analizi gerçekleştirin.
2. depletion için genel prosedür
- Tüm örneklerin toplanması için ne kadar kültürün gerekli olduğunu hesaplayın; Örneğin, 0,8 OD600 'de 10 ml kültür, tek bir numune için PROTEIN, RNA ve DNA ekstraksiyonu için yeterlidir, bu nedenle 6 numune için en az 60 ml kültür gereklidir.
- Bir gecede kültürden, OD600 0,1 ila 0,2 ve 30 °c ' de büyümeye bırakın yeterli yeni kültür kurmak. Diğer büyüme koşulları kullanılabilir olsa da, YPDA gibi zengin bir ortam önerilir:
Maya özü 10 g Pepton 20 g Glikoz 20 g Adenin sülfat 40 mg H2O için 1 L
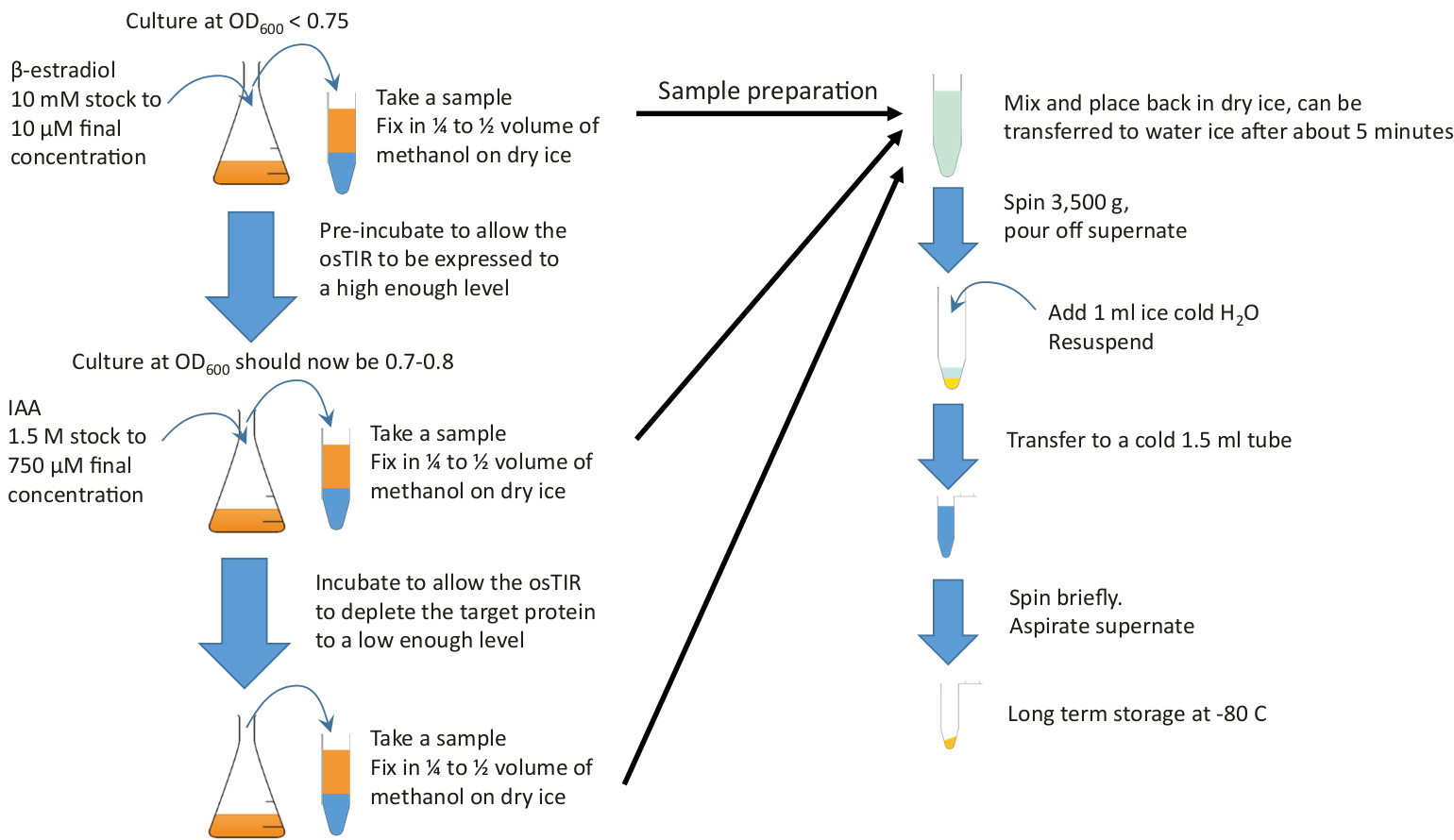
Not: Otoklav veya filtre sterilize; Filtre sterilizasyonu, numune koleksiyonunda kullanılan metanol içinde otoklavlama tarafından üretilen peptid/şeker kompleksler olarak tercih edilir. - Örnekleri almaya hazırlanın.
- Metanol amaçlanan numune hacminin bir tüp içine% 30-50% koyun. Örneğin, 10 mL örnek alınırsa, 15 mL şahin tüpüne 5 mL metanol koyun ve tüpü sıkıca kapatın. Kapalı bir kez, tüp etiket ve kuru buz veya at-80 °C soğuk koymak.
Dikkat: Metanol bir duman kaputu içinde dağıtın. - Etiket 1,5 uzun süreli depolama için mL tüpler ve soğuk buzda yer örnekleri.
- Buz üzerinde yeterli H2O (örnek başına en az 1 ml) serin.
- Metanol amaçlanan numune hacminin bir tüp içine% 30-50% koyun. Örneğin, 10 mL örnek alınırsa, 15 mL şahin tüpüne 5 mL metanol koyun ve tüpü sıkıca kapatın. Kapalı bir kez, tüp etiket ve kuru buz veya at-80 °C soğuk koymak.
- Kültürün büyümesini öngörün. Örnekleri toplamak için hedef OD yaklaşık 0,7 için 0,8, ancak pre-inkübasyon adım (β-estradiol ile inkübasyon osTIR teşvik etmek), böylece kültür yaklaşık doğru OD zaman numuneler ulaşacaktır daha önce başlaması gerekiyor Toplanan.
Not: Bu başlangıç OD tahmin edilebilir böylece deney kullanılmak üzere koşullarda bir büyüme eğrisi gerçekleştirmek için tavsiye edilir. - Ön kuluçka başlangıcı için hedef OD ulaşıldığında, bir örnek almak (genellikle 10 mL), soğuk metanol içeren önceden hazırlanmış tüp içine. Karıştırmak ve kuru buzda geri yerleştirmek için kısaca tersine çevir.
Not: Örnek, bunu yapmak için uygun ise, yaklaşık 5 dakika sonra su buzuna taşınabilir. - Hemen β-estradiol, 1 μL/mL Kültür (son konsantrasyon 10 μM) ekleyin; numune toplama ve β-estradiol ekleyerek arasında alınan süreyi azaltmak için, bir pipet içinde önceden ölçülen β-estradiol ön ölçülmüştür. Hızla karıştırarak güçlü bir şekilde karıştırmak.
- Daha önce olduğu gibi kültür büyümeye devam (adım 2,2), (Bu "Pre-inkübasyon" adım) en uygun zaman için β-estradiol ile (optimum ön kuluçl süresi belirlenmesi için bakın Şekil 1).
- IAA (auxin) eklemeye hazırlanın. Adım 2,10 için gerekli ıAA hacmini alın (örn., mL kültür başına ıAA 0,5 μL). Bu adım 2,20 daha hızlı hale getirir.
- Adım 2,5 olarak bir örnek toplayın.
- Hemen ıAA 0,5 μL/mL kültürü, adım 2,8 ' de hazırlanan 750 μM son konsantrasyonuna ekleyin. Hızla karıştırarak güçlü bir şekilde karıştırmak.
- Deneysel tasarım göre, adım 2,5 olarak, örnekleri toplayın. Ya tek bir örnek, bir anda ne zaman bu protein güvenilir bir şekilde tükenmiş olacak, ya da bir zaman ders tükenmesi birden fazla numune bekleniyor. Örneğin, 5 dk aralıklarla zamanlama için uygundur ve protein seviyeleri bir dizi sağlamak. En iyi duruma getirme stratejisi, Şekil 1' de gösterildiği gibi uygun zamanlarda bir belirti verecektir.
- Örnekleri işleme.
- Örnekleri buz üzerine yerleştirin, zaten yapılırsa. Numunelerin hiçbiri dondurulmadığından emin olun; varsa, yavaşça elle sıcak, böylece sıcaklık yerel olarak yükselmez sürekli tersine çevirme.
Not: Bu en iyi örnek sıcaklık değerlendirilebilir olarak el yapılır, her zaman soğuk hissedebilirsiniz. Buzun üzerine yerleştirin. Bu bir duraklama noktası değil-bir kez tüm örnekleri sıvı, sonraki adıma geçin. - Tüm örnekler toplanan ve artık dondurulmuş ise, 2 dk (mümkünse 4 °C) için 3.500 x g spin.
- Metanol/orta karışımı dökün ve buzun üzerine geri yerleştirin; Tüm sıvı kaldırılmışsa endişelenmeyin.
- 1 ml buz soğuk H2O (adım 2.3.3) hücre Pelet resuspend ve bir etiketli 1,5 ml tüp (adım 2.3.2 yılında hazırlanmış) buz üzerinde transfer.
- > 15000 x g 'de kısa sürede (örn. 10 s toplam zaman) hücreleri yeniden Pellet, buzun üzerine geri yerleştirin ve sıvıyı çıkarın.
- H2O aspirasyon ile çıkarın. Hücre pelalları uzun süreli depolama için-20 °C veya-80 °C ' de depolanabilir.
- Örnekleri buz üzerine yerleştirin, zaten yapılırsa. Numunelerin hiçbiri dondurulmadığından emin olun; varsa, yavaşça elle sıcak, böylece sıcaklık yerel olarak yükselmez sürekli tersine çevirme.
- Batı leke Analizi18ile protein tükendiği seviyesini kontrol edin.
Not: Nadir RNA türlerinin daha fazla numune hacmi gerektirmesine karşın, yeterli protein19 ve/veya nükleik asit tek bir hücre pelekinden çoğu amaçla çıkartılabilir.
Sonuçlar
Örnek tükenme örnekleri Şekil 1' de görüntülenir. Bu figürde sunulan üç deney, Prp2, Prp22 ve Dcp1 proteinlerinin tükenmesi için optimizasyon deneyleridir. Düşük bolluk, intronları Prp2 ve Prp22 proteinleri her ikisi de% 20 ' den az tükendikten sonra β-estradiol ile 40 min pre-inkübasyon sonra auxin ile 15 dakika takip. Daha uzun pre-inkübasyon süreleri daha hızlı tükenme yol ama aynı zamanda oksin vardır ilavesi önce istenmeyen protein tükenmesi göstermek. Buna kıyasla, daha bol Dcp1 sadece aynı tedavi ile yaklaşık% 30 tükenmiş, ancak 60 dk pre-inkübasyon% 13 ile aynı oksin vardır tedavisi ile, tükenme maliyeti, oksin vardır eklenmeden önce bitmiş sonuçlandı. Β-estradiol ve oksin vardır ile 15 dakika pre-inkübasyon en az 50 dk daha kısa bir zaman noktasında benzer sonuçlar elde olurdu ve bu yüzden daha optimal olurdu mümkündür.

Şekil 1: tükenme oranı β-estradiol pre-inkübasyon süresini modülasyon ile ayarlanabilir. Batı leke18 hedef proteinleri: (a ve b) Prp22-Aid *-6flag, (c ve d) Prp2-Aid *-6flag, ve (e ve f) Dcp1-Aid *-6ha, daha önce β-estradiol (β-EST) ile önceden inkübatlı kültürlerden 20, 30, 40, veya 60 min önce oksin vardır ek12. Her şeritte eşit miktarlarda toplam protein yüklendi. Pgk1 panel edışında bir görsel yükleme denetimi olarak algılanır, burada Pgk1 ve Dcp1 Co-migrate. A, c ve e panellerinde protein bandının ölçülmesini sırasıyla b, dve f panellerinde gösterilir. Tükenme hızı ölçüsü olarak, eğim (m) doğrusal bölüm için hesaplanır (dan 100% için 30% başlangıç değerlerinin) her eğri. Optimum pre-inkübasyon süresi, protein düzeylerinin hala un kaynaklı seviyelere (% 100) yakın olduğu zamandı. ve sonraki tükenme oranı hızlıdır. Dcp1 (f) için, 60 min pre-kuluçk çok uzun, protein auxin yokluğunda zayıflatmaya başladı gibi, 20 dakika çok kısa ise, protein bu zaman kursuna fark etmez gibi. 40 min pre-inkübasyon sonra, protein yaklaşık 70% tükenmiş olarak oksin vardır ile 15 dakika kullanılabilir ve, 20 dakika daha fazla tükenme neden olsa da, aynı zamanda ikincil etkilere neden olabilir. Hata çubukları iki biyolojik çoğaltır standart sapmasını temsil eder. Her deney için bir temsilci leke gösterilir. Bu rakam önceki yayın9' dan türetilmiştir. Bu figürün daha büyük bir versiyonunu görmek Için lütfen tıklayınız.
{kind=link}

Şekil 2: grafiksel Özet. Önceden inkübasyon başlatmak için zengin Orta ve gerekli sıcaklıkta büyüyen yeterli kültüre β-estradiol ekleyin. Tükenmeye başlamak için ıAA (auxin) eklemeden önce gerekli ön kuluçba zaman büyüme devam edin. Ön kuluçba ve tükenme süreleri proteinin tükenmesine bağlıdır, ancak pre-inkübasyon genellikle 20-60 dk aralığında ve tükenmesi süresi genellikle 10 ila 20 dk. 10 mL numuneler ön kuluçdak ve Durin başlangıcı ve sonunda alınmalıdır. g tükenmesi. Bu numuneler, peletleme ve depolama öncesinde soğuk metanol içinde hızla sabitlenir. Bu figürün daha büyük bir versiyonunu görmek Için lütfen tıklayınız.
{kind=link}

Şekil 3: B-EST sistemi Için gerinim üretimi. (a) yardım * sistemi ile bir Maya gerinim oluşturmak için, ya pZTRL (LEU2) veya pztrk (Kanamycin (G418) direnç), Plasmid gerinme içine tanıtılmalıdır veya, alternatif olarak, ATF ve ostir tarafından genom içine eklenebilir PCR tarafından 3 ' End astarlardan oluşturulan bir parçanın homolog yeniden kombinasyonu (bkz. Şekil 3B ve Tablo 1). (b) C-hedef proteinin Terminal etiketleme Plasmid PURA3-AID uygun bölgenin PCR amplifikasyon tarafından elde edilir *-6Flag (pURA3_AID *-6ha sadece etiketi farklıdır ve tam olarak aynı şekilde tedavi edilebilir), Longtine astar S3-F kullanarak ve 3 ' uzatma ile S2-R 3 ' hedef protein sonuna homolog. İleri primer uzatma yardım * etiketi başlangıcı ile çerçeve içinde son amino asit kodon içermelidir ve stop kodon içermemelidir. Ters primer uzantısı, kodlama bölgesinin aşağı doğru bir bölgeye olmalıdır. Bir kez GENOMA takılı, URA3 Marker (işaretçinin her iki ucunda bulunan özdeş bölgeler arasında Homolog rekombinasyon tarafından) kaybetti hücreleri 5-FOA ile büyüme tarafından seçilebilir, bu sayaç- URA3 hücreleri seçer. Bu figürün daha büyük bir versiyonunu görmek Için lütfen tıklayınız.
{kind=link}
| a. Primer dizileri | |||||||
| Hedef | Konum | Astar | Adı | Sıra | TM (°C) | ||
| (pZTRL) | 516 | F | pZTRL_F | <-Homoloji Bölgesi-> GCGACAGCATCACCGACTTCG | 61,23 | ||
| 7897 | R | pZTR_R | CGCCGCCTCTACCTTGCAGA <-Homoloji bölgesi (RC)-> | 61,30 | |||
| (pZTRK) | 9154 | F | pZTRK_F | <-Homoloji Bölgesi-> ACGTTGAGCCATTAGTATCAATTTGCTTACC | 59,40 | ||
| 5897 | R | pZTR_R | CGCCGCCTCTACCTTGCAGA <-Homoloji bölgesi (RC)-> | 61,30 | |||
| pURA3-AID *-6FLAG veya pURA3_AID *-6HA | F | S3-F | <-Homoloji Bölgesi-> CGTACGCTGCAGGTCGAC | 59,21 | |||
| R | S2-R | ATCGATGAATTCGAGCTCG <-Homoloji bölgesi (RC)-> | 52,76 | ||||
| pZRTL/K β-EST yardım sistemini güçlendirir | |||||||
| pURA3-Aid *-6flag/6ha yardım * ve epitopu etiketi hedef protein (lontine prosedürü) etiketlemek için yükseltmek için | |||||||
| <-Homoloji Bölgesi-> | Bölge, sistemin yerleştirileceği yer olan çevrelerden homolog. Bu bölgenin daha uzun olması, değişikliğin başarılı olması daha olasıdır; 50-100 üsleri tavsiye edilir. | ||||||
| <-Homoloji bölgesi (RC)-> | Bölge, sistemin yerleştirileceği Flanşli bölgelere homolog, ters tamamlayıcı kullanmayı unutmayın. Yukarıdaki gibi, bu bölge daha iyi. | ||||||
| TM (°C) | TM 50 mM NaCl ile% GC yöntemini kullanarak | ||||||
| b. PCR Mix | |||||||
| Bileşen | Hacim (μL) | ||||||
| Şablon | < 10 | ||||||
| NEB Phusion HF tampon (5x) * | 100 | ||||||
| İleri primer 100 μM | 2,5 | ||||||
| Ters primer 100 μM | 2,5 | ||||||
| her biri 10 mM dNTPs | 10 | ||||||
| H2O | 500 için | ||||||
| * NEB Phusion GC Buffer (5x) de kullanılabilir ancak tercih edilmez | |||||||
| Bu karışımı olun, 10 tüpler içine bölünmüş 50 μL her mix ve tablo 1 c olarak PCR gerçekleştirin. | |||||||
| PCR bir agaroz jel üzerinde çalışan tarafından çalıştı kontrol | |||||||
| Bir tüp ve etanol çöktürme tüm başarılı reaksiyonlar birleştirin | |||||||
| PCR tarafından üretilen tüm malzemeyle Maya dönüşümü | |||||||
| c. PCR koşulları | |||||||
| Adım | Sıcaklık (°C) | Zaman | |||||
| İlk denaturation | 98 | 30 sn | |||||
| 25-35 döngüleri | Denatüre | 98 | 10 sn | ||||
| Tavlama | 45 – 60 | 20 sn | |||||
| Uzantısı | 72 | 30 s/KB | |||||
| Final uzatma | 72 | 10 dak. | |||||
| Tutun | 8 | ||||||
| PZTRL/K astar için Lontine astar seti (S3-F ve S2-R) ve 60 °C için 45 °C ' de anneal | |||||||
| Lontine PCR için 3 dakika ve pZTRL/K için 3 dakika uzatın | |||||||
Tablo 1: Primer dizileri, PCR karışımı ve PCR koşulları.
Tartışmalar
İyi optimize edilmiş bir protokol, hedef proteinin hızlı ve verimli bir şekilde tükenmesine neden olabilir. Β-estradiol ile yaklaşık pre-inkübasyon süresini belirlemek önemlidir, bu da tükenme tekrarlanabilirlik artar, ancak ön kuluçba zaman küçük varyasyonlar tolere edilebilir. Diğer taraftan, protein seviyesi çok hızlı düşüşler gibi, oksin vardır ilavesi sonrasında zamanlama ile bakım alınmalıdır.
Bu yaklaşımın bir avantajı, β-estradiol ve ıAA inkübasyon süresi ile Pre-inkübasyon süresi kombinasyonlarını değiştirerek ayarlanan tükenme elde edilebilir olmasıdır. Örneğin, istenirse, hedef proteinin ön kuluçca süresini azaltarak daha yavaş tükenmiş olabilir.
Β-EST AıD sistemi, OsTIR 'in kurucu olarak ifade edildiği sistemlerde bazı avantajlar sunar. Örneğin, hedef protein viability için gerekli ise, osTIR düzenlenmiş ifade hedef protein erken tükenmesi önleyebilirsiniz. Dahası, osTIR ifadesi, hedef proteinin bolluğuna ve bozulmaya karşı duyarlılığını karşılamak için ayarlanabilir ve tükenmesi ya hızlı ya da yavaş olabilir. İki küçük molekül Efektörler, β-estradiol ve auxin, burada kullanılan koşullar altında Maya metabolizmasını Perturb değil, Rapamycin aksine, çapa-Away sistemi1kullanılır.
Bazı proteinlerin etiketlenmesi, herhangi bir hedeflenen tükenme sistemi ile ilgili bir sorun olan işlevlerini bozacak şekilde belirtilmelidir. Bu durumda, C-Terminal etiketi olmadığında bir N-terminal etiketi çalışabilir. Ayrıca, tüm proteinler verimli bir şekilde tükenmiş olacak; Örneğin, hedef proteinin AID-Tag osTIR protein erişilemez olabilir. Bu nedenle, yardım-etiketleme sonra, her hedef protein büyüme etiketinin herhangi bir etkisi için test edilmelidir, ve tükenme etkili olup olmadığını belirlemek için, β-estradiol pre-inkübasyon ve oksin vardır tedavi zamanlamaları optimize edilmiştir önce.
Bu AıD * sistemi çok basittir ve daha fazla büyüme içermeyen herhangi bir sonraki deneysel prosedür ile uyumludur, protein gibi, DNA veya RNA analizi veya mikroskopisi. Buna ek olarak, sistem iyi bir şekilde birlikte çalışırken, bu RNA20 ' siniarındırmak için ne zaman çalışır.
Bu sistem, Maya hücresinin metabolizmasını başka bir şekilde etkilemeden bir proteini tüketen etmek için hızlı, spesifik ve tekrarlanabilir bir yol sağlar.
Açıklamalar
Yazarların ifşa etmesi gereken hiçbir şey yok.
Teşekkürler
Bu programı başlattığı için Jane Reid sayesinde, geliştirme için Barbara Terlouw, "Ura Looper" yapıları için vahid Aslanzadeh ve birçok yararlı tartışmalar için Susana de Lucas. Bu çalışma Consejo Nacional de ciencia y Tecnología, Meksika (CONACYT) ve Edinburgh Üniversitesi Biyoloji Bilimleri, IEM [105256] için Wellcome doktora öğrencilik ve Wellcome fon [104648] JD Beggs gelen Gimo için bir burs tarafından desteklenmektedir . Hücre biyoloji Wellcome merkezi 'nde çalışma Wellcome çekirdek fon tarafından desteklenmektedir [092076].
Malzemeler
| Name | Company | Catalog Number | Comments |
| Adenine sulphate | Formedium | DOC0230 | |
| Agar | Formedium | AGA03 | |
| Β-estradiol | Sigma Aldrich | E2758-1G | 10 mM solution in ethanol. Store at -20 °C |
| DMSO | Alfa Aesar | 42780 | DMSO should be solid at 4 °C |
| Glucose | Fisher Scientific | G/0500/60 | |
| IAA 1H-Indole-3-acetic acid | Across Orgainics | 122150100 | Auxin analogue. 1.5 M in DMSO. The solution will be a russet colour and darken as time goes on; a deep red solution should be discarded and a new one made. Store at -20 °C. |
| Methanol | Fisher Scientific | M/4000/PC17 | CAUTION Toxic and flammable |
| Phusion High-Fidelity DNA Polymerase | NEB | M0530 | |
| Peptone | Formedium | PEP03 | |
| SCSM single drop-out –ura | Formedium | DSCS101 | |
| Yeast Extract | Formedium | YEA03 | |
| Yeast nitrogen base without amino acids with amonium sulphate | Formedium | CYN0410 |
Referanslar
- Haruki, H., Nishikawa, J., Laemmli, U. K. The Anchor-Away Technique: Rapid, Conditional Establishment of Yeast Mutant Phenotypes. Molecular Cell. 31, 925-932 (2008).
- Bellí, G., Garí, E., Piedrafita, L., Aldea, M., Herrero, E. An activator/repressor dual system allows tight tetracycline-regulated gene expression in budding yeast. Nucleic Acids Research. 26, 942-947 (1998).
- Alexander, R. D., et al. RiboSys, a high-resolution, quantitative approach to measure the in vivo kinetics of pre-mRNA splicing and 3′-end processing in Saccharomyces cerevisiae. RNA. 16, 2570-2580 (2010).
- Deshaies, R. J., Joazeiro, C. A. P. RING Domain E3 Ubiquitin Ligases. Annual Review of Biochemistry. 78, 399-434 (2009).
- Tan, X., et al. Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature. 446, 640-645 (2007).
- Teale, W. D., Paponov, I. A., Palme, K. Auxin in action: signalling, transport and the control of plant growth and development. Nature Reviews Molecular Cell Biology. 7, 847-859 (2006).
- Nishimura, K., Fukagawa, T., Takisawa, H., Kakimoto, T., Kanemaki, M. An auxin-based degron system for the rapid depletion of proteins in nonplant cells. Nature Methods. 6, 917-922 (2009).
- Morawska, M., Ulrich, H. D. An expanded tool kit for the auxin-inducible degron system in budding yeast. Yeast. 30, 341-351 (2013).
- Kubota, T., Nishimura, K., Kanemaki, M. T., Donaldson, A. D. The Elg1 Replication Factor C-like Complex Functions in PCNA Unloading during DNA Replication. Molecular Cell. 50, 273-280 (2013).
- Brosh, R., et al. A dual molecular analogue tuner for dissecting protein function in mammalian cells. Nature Communications. 7, 11742 (2016).
- DeRisi, J. L., Iyer, V. R., Brown, P. O. Exploring the metabolic and genetic control of gene expression on a genomic scale. Science. 278, 680-686 (1997).
- Mendoza-Ochoa, G. I., et al. A fast and tuneable auxin-inducible degron for depletion of target proteins in budding. Yeast. , (2018).
- McIsaac, R. S., et al. Synthetic gene expression perturbation systems with rapid, tunable, single-gene specificity in yeast. Nucleic Acids Res. 41, e57 (2013).
- Prusty, R., Grisafi, P., Fink, G. R. The plant hormone indoleacetic acid induces invasive growth in Saccharomyces cerevisiae. PNAS. 101, 4153-4157 (2004).
- Geitz, D., St Jean, A., Woods, R. A., Schiest, R. H. Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Research. 20, 1425 (1992).
- Widlund, P. O., Davis, T. N. A high-efficiency method to replace essential genes with mutant alleles in yeast. Yeast. 22, 769-774 (2005).
- Longtine, M. S., et al. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast. 14, 953-961 (1998).
- Eaton, S. L., et al. A Guide to Modern Quantitative Fluorescent Western Blotting with Troubleshooting Strategies. Journal of Visualized Experiments. , e52099 (2014).
- Volland, C., Urban-Grimal, D., Géraud, G., Haguenauer-Tsapis, R. Endocytosis and degradation of the yeast uracil permease under adverse conditions. Journal of Biological Chemistry. 269, 9833-9841 (1994).
- Barrass, J. D., et al. Transcriptome-wide RNA processing kinetics revealed using extremely short 4tU labeling. Genome Biology. 16, 282 (2015).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır