
Method Article
Tuning Degradation, um eine spezifische und effiziente Proteinerschöpfung zu erreichen
In diesem Artikel
Zusammenfassung
Hier stellen wir ein Protokoll vor, um ein Protein von Interesse in der Hefe Saccharomyces cerevisiae mit dem AID-System effektiv und spezifisch zu erschöpfen.
Zusammenfassung
Der pflanzliche Auxin-Bindungsrezeptor TIR1 erkennt Proteine, die ein spezifisches Auxin-induzierbares Degron(AID)-Motiv in Gegenwart von Auxin enthalten, und zielt auf sie zum Abbau. Dieses System wird in vielen nicht-pflanzlichen Eukaryoten ausgenutzt, so dass ein Zielprotein, das mit dem AID-Motiv markiert ist, nach Auxin-Zugabe abgebaut wird. Die Ebene der TIR1-Expression ist entscheidend; übermäßige Expression führt zu einer Verschlechterung des AID-markierten Proteins auch ohne Auxin, während eine niedrige Expression zu einer langsamen Erschöpfung führt. Es wurde ein induzierbares AID-System mit dem Ausdruck TIR1 unter der Kontrolle eines induzierbaren Promotors von '-estradiol erstellt. Der TIR1-Gehalt kann abgeändert werden, indem die Inkubationszeit mit dem Estradiol vor der Auxin-Zugabe geändert wird. Dieses Protokoll beschreibt, wie ein Zielprotein mithilfe des AID-Systems schnell aufgebraucht werden kann. Die entsprechende Inkubationszeit von '-estradiol hängt von der Häufigkeit des Zielproteins ab. Daher hängt eine effiziente Erschöpfung von einem optimalen Timing ab, das auch die auxinunabhängige Erschöpfung minimiert.
Einleitung
Bedingte Mutationen, wie temperaturempfindliche Mutanten, sind ein leistungsfähiges Werkzeug für die Untersuchung essentieller Proteine, die das Zellwachstum unter dem freizügigen Zustand ermöglichen, aber unter nicht-freizügigen Bedingungen zum Verlust der Funktion führen. Jedoch, Zellstoffwechsel kann durch die Veränderung der Wachstumsbedingungen erforderlich, um den Defekt zu induzieren ernsthaft gestört werden und kann auch Off-Target-Effekte verursachen. Es wurden mehrere Methoden entwickelt, bei denen das Protein von Interesse bedingt1 sequestriert wird oder seine Expression2,3 durch Zugabe eines kleinen Moleküls gesteuert wird. Dieses Protokoll verwendet Auxin und das Auxin-induzierbare Degron(AID)-System, um ein Zielprotein effizient zu erschöpfen.
Das AID-System hat seinen Ursprung in Pflanzen, wo ein Auxin (in diesem Protokoll Indole-3-Essigsäure (IAA) verwendet wird), stimuliert die Interaktion des Aux/IAA-Proteins mit TIR1, einem Mitglied des SCF U3 Ubiquitin Ligase-Komplexes4. SCF-komplexe Wechselwirkung verursacht Polyubiquitination von Aux/IAA-Familie Proteine, die in ihrem Abbau durch das Proteasomführt 5,6.
Dieses System wurde zuvor für den Einsatz in der Hefe Saccharomyces cerevisiae7,8 angepasst, indem das TIR1-Protein aus Oriza sativa (osTIR) in Hefezellen exexziert wird, wo es in der Lage ist, mit der endogenen Hefe zu interagieren. SCF-Komplex. Das Protein von Interesse wurde mit einem Motiv aus dem Aux/IAA-Protein IAA17 getaggt, um es für den Abbau zu zielen. Später wurden funktionelle Abkürzungen von IAA17 entwickelt, wie AID*8,9,10, die das 43 Aminosäure-Auxin-sensitive Motiv von Arabidopsis thaliana IAA17 enthalten, zusammen mit einem Epitop-Tag, um entdeckung.
Das System, das ursprünglich für den Einsatz in der angehenden Hefe7angepasst war,8 exprimierte das osTIR1-Protein von einem Hefe-GAL-Promotor. Expression erfordert die Umstellung auf Wachstumsmedium mit Galaktose als einzige Kohlenstoffquelle, was leider zu einer diauxischen Verschiebung mit weitreichenden Veränderungen des Zellstoffwechsels führt11. Andererseits wurde berichtet, dass die konstitutive Expression von TIR1 ohne Auxin/IAA12 zu einer Verschlechterung des Zielproteins führen kann, wenn der Expressionspegel hoch ist, während eine niedrige TIR1-Expression eine ineffiziente Erschöpfung verursacht. Es wurde ein verbessertes AID-System mit dem Namen '-est AID entwickelt, bei dem das osTIR unter der Kontrolle eines induzierbaren Promotors steht, der auf das Zielprotein abgestimmt werden kann, mit minimaler Wirkung auf den Zellstoffwechsel. Um dies zu erreichen, wurde ein künstlicher Transkriptionsfaktor (ATF) konstruiert, bei dem der VIRUStranskriptionsaktivator VP16 mit einem Östrogenrezeptor und einer DNA-Bindungsdomäne (DBD) von vier Zn-Fingern verschmolzen ist. Wenn das Östradiol (ein Östrogen) vorhanden ist, kann die ATF in den Kern eindringen und die osTIR-Transkription durch Bindung an ihren Promotor (Z4EVpr)13,12induzieren.
osTIR-Expression ist in der Regel ca. 20 min nach Zugabe von '-estradiol12nachweisbar. Die optimale Dauer der osTIR-Expression, um eine effiziente Erschöpfung des markierten Proteins mit Auxin zu erreichen und gleichzeitig eine Erschöpfung vor auxinaddition zu vermeiden, muss jedoch empirisch für jedes Zielprotein bestimmt werden. Eine ungefähre Zeit für diese Vorinkubation kann anhand von Überflusswerten in der Saccharomyces Genome Database (SGD https://www.yeastgenome.org/) geschätzt werden. Wie in Abbildung 1zu sehen ist, benötigt das reichlich eisernreiche Protein Dcp1 (2880 bis 4189 Moleküle/Zelle) 40 min Vorinkubation mit '-estradiol, ohne dass eine auxinunabhängige Erschöpfung beobachtet wird. Das viel weniger reichlich eitel Protein Prp2 (172 bis 211 Moleküle/Zelle) ist nach nur 20 min Vorinkubation stark erschöpft. Es ist ratsam, zwei zusätzliche Vorinkubationszeiten zu testen, 10 bis 20 min vor oder nach dieser ersten geschätzten Zeit (20 min ist die empfohlene Mindestzeit). Die optimale Vorinkubationszeit ist die Zeit, in der das Zielprotein vor dem Hinzufügen von Auxin nicht erschöpft ist und nach Zugabe von Auxin ist die Erschöpfung akzeptabel oder der Proteingehalt nähert sich dem Minimal möglich. So, aus Abbildung 1b, für Prp22 mit 30 min Vorinkubation, die Ebenen haben nicht viel 10 min nach Auxin Addition gesunken. Vergleicht man dies mit 40 min Vorinkubation und 15 min mit IAA, wo es wenig zusätzliche Erschöpfung gibt, gibt es keinen Nutzen bei der Inkubation mit Auxin länger als 10 min oder vor der Inkubation länger als 30 min, insbesondere da es Hinweise auf Nicht-Auxin gibt. abhängige Erschöpfung bei 40 min. Für Dcp1 mit 40 min Vorinkubation (der letzte Punkt, an dem der Proteingehalt etwa 100% vor Auxin-Zugabe beträgt), sind 15 bis 20 min Erschöpfung mit Auxin akzeptabel. Es wird empfohlen, die Erschöpfungszeit so kurz wie möglich zu halten, um sekundäre Auswirkungen auf den Zellstoffwechsel zu reduzieren14.
Dieser Artikel zeigt, wie sie das AID-System verwenden kann, indem Sie das Timing der indizonlintisten inkubation für die osTIR-Expression optimieren, um eine schnelle Zielproteinerschöpfung bei IAA-Zugabe ohne Erschöpfung zu erreichen, bevor Auxin hinzugefügt wird.
Protokoll
HINWEIS: Eine grafische Zusammenfassung finden Sie in Abbildung 2.
1. Dehnungsvorbereitung
-
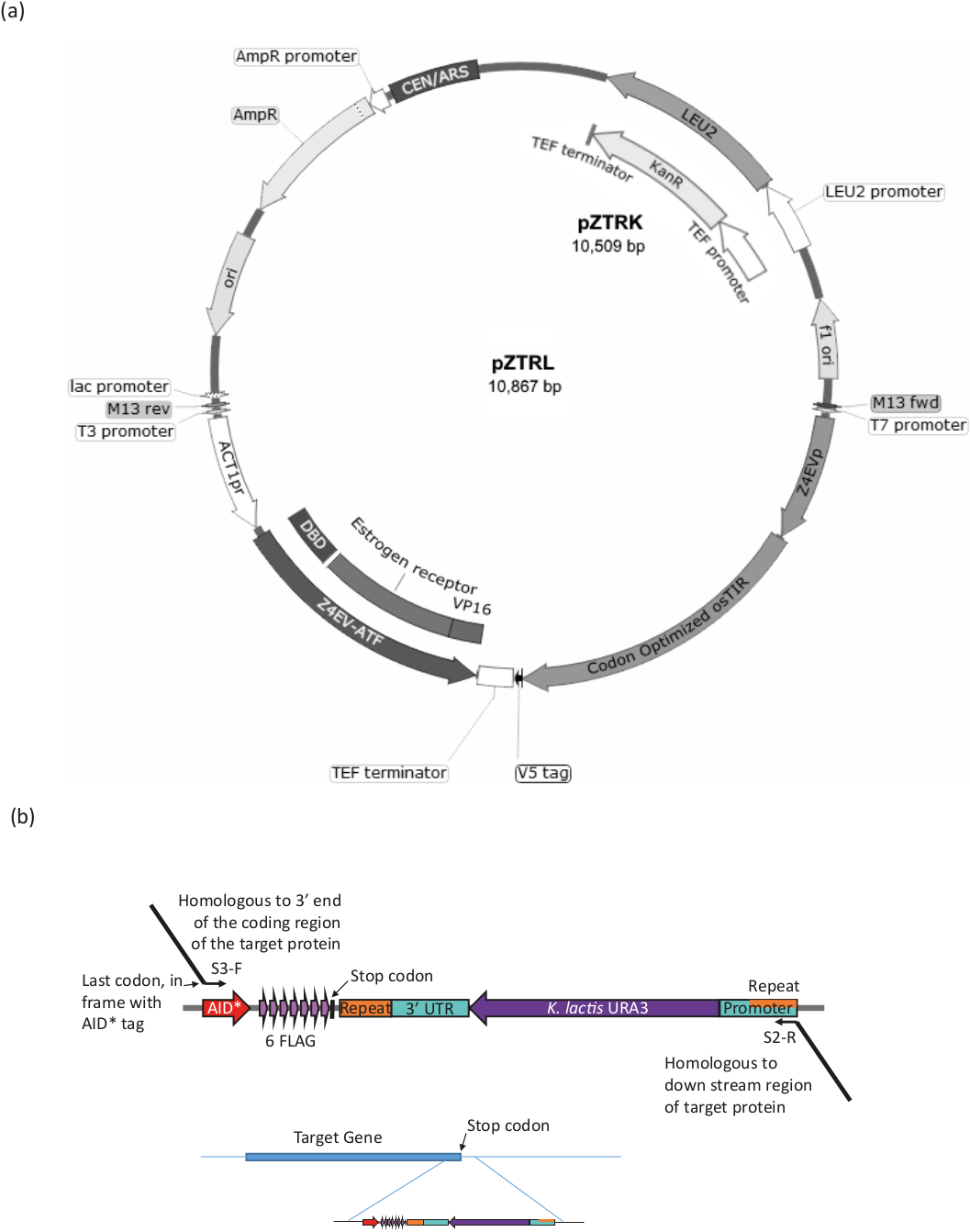
Führen Sie mit einem ura3-Stammdas AID-System ein (d. h. Gene, die den -estradiol-responsive nontiver Transkriptionsfaktor (ATF) und den osTIR kodieren) und AID* markieren das Zielprotein (siehe Abbildung 3 und Tabelle 1 für eine Zusammenfassung des Verfahrens).
- Verwandeln Sie15 entweder pZTRK (G418-Widerstandsmarker) oder pZTRL (LEU2-Marker) Plasmid (verfügbar vom Yeast Genetic Resource Centre) in den ura3-Hefestamm oder verwenden Sie das Plasmid als Vorlage, um das PCR-Produkt für die Genomik zu produzieren integration.
- PCR verstärken die ATF (auf der Plasmidkarte mit Z4EVATF gekennzeichnet) und osTIR mit einer Hochtreuepolymerase aus einem der Plasmide pZTRK oder pZTRL. Verwenden Sie Primer mit 50 bis 60 Base 3' Erweiterungen mit Homologie in die Genomregion, um die Integration durch homologe Rekombination zu lenken16. Zur genomischen Integration der beiden Komponenten entweder getrennt oder zusammen siehe Tabelle 1 für Primer und Bedingungen.
HINWEIS: Der Stamm pZ4EV-NTR1 enthält die Komponenten, die bereits im Genom integriert sind (erhältlich vom Yeast Genetic Resource Centre, Japan). - Stellen Sie sicher, dass das Zielprotein AID* mit dem Longtine-Verfahren17 markiert ist (siehe Abbildung 3b und Tabelle 1).
- Führen Sie eine Wachstumsanalyse des Stammes durch, ohne dass das Gerät ohne das vorhandene Estradiol und IAA vorhanden ist, um festzustellen, ob das AID*-Tag das Wachstum beeinflusst, und um die Wachstumsrate für die Verwendung bei Schritt 1.5 vorherzusagen.
2. Allgemeines Verfahren zur Erschöpfung
- Berechnen Sie, wie viel Kultur für die Erfassung aller Stichproben erforderlich ist. zum Beispiel reichen 10 ml Kultur bei OD600 von 0,8 für die Protein-, RNA- und DNA-Extraktion für eine einzelne Probe aus, so dass für 6 Proben mindestens 60 ml Kultur benötigt werden.
- Von einer Nachtkultur, richten Sie eine ausreichende neue Kultur bei OD600 0.1 bis 0.2 ein und lassen Sie es bei 30 °C wachsen. Ein reichhaltiges Medium wie YPDA wird empfohlen, obwohl andere Wachstumsbedingungen verwendet werden können:
Hefe-Extrakt 10 g Peptone 20 g glukose 20 g Adeninsulfat 40 mg H2O bis 1 L
HINWEIS: Autoklav oder Filter sterilisieren; Die Filtersterilisation wird als Peptid-/Zuckerkomplex bevorzugt, die durch Autoklavieren im bei der Probenentnahme verwendeten Methanol hergestellt werden. - Bereiten Sie sich auf den Erhalt der Proben vor.
- 30 bis 50 % des vorgesehenen Probenvolumens von Methanol in eine Röhre geben. Wenn beispielsweise eine 10 ml-Probe entnommen werden soll, legen Sie 5 ml Methanol in ein 15 ml Falkenrohr und schließen Sie das Rohr fest. Nach dem Schließen die Röhre beschriften und zum Abkühlen auf Trockeneis oder bei -80 °C legen.
ACHTUNG: Das Methanol in einer Dunstabzugshaube abgeben. - 1,5 ml Rohre für die Langzeitlagerung der Proben beschriften und in Eis abkühlen lassen.
- Genug H2O (mindestens 1 ml pro Probe) auf Eis abkühlen lassen.
- 30 bis 50 % des vorgesehenen Probenvolumens von Methanol in eine Röhre geben. Wenn beispielsweise eine 10 ml-Probe entnommen werden soll, legen Sie 5 ml Methanol in ein 15 ml Falkenrohr und schließen Sie das Rohr fest. Nach dem Schließen die Röhre beschriften und zum Abkühlen auf Trockeneis oder bei -80 °C legen.
- Antizipieren Sie das Wachstum der Kultur. Die Ziel-OD für das Sammeln der Proben ist etwa 0,7 bis 0,8, aber der Vorinkubationsschritt (die Inkubation mit dem '-estradiol, um die osTIR zu induzieren), muss früher gestartet werden, damit die Kultur ungefähr die richtige OD erreicht, wenn die Proben ruhig.
HINWEIS: Es ist ratsam, eine Wachstumskurve unter den Bedingungen durchzuführen, die im Experiment verwendet werden sollen, damit dieser Start-OD geschätzt werden kann. - Sobald das Ziel OD für den Beginn der Vorinkubation erreicht ist, nehmen Sie eine Probe (in der Regel 10 ml), in das vorgefertigte Rohr mit kaltem Methanol. Kurz umkehren und in Trockeneis zurücklegen.
HINWEIS: Die Probe kann nach ca. 5 min ins Wassereis bewegt werden, wenn dies zweckmäßig ist. - Sofort das '-estradiol, 1 l/ml Kultur (Endkonzentration von 10 'M); das in einer Pipette vormessene Sestradiol einsatzbereit haben, um die Zeit zwischen dem Sammeln der Probe und dem Hinzufügen des '-estradiol zu reduzieren. Schnell mischen, indem Sie kräftig wirbeln.
- Die Kultur wie bisher weiter ausbauen (Schritt 2.2), inkubieren (dies ist der "Pre-Inkubationsschritt") mit dem '-estradiol für die optimale Zeit (zur Bestimmung der optimalen Vorinkubationszeit siehe Abbildung 1).
- Bereiten Sie das Hinzufügen von IAA (Auxin) vor. Nehmen Sie das für Schritt 2.10 erforderliche IAA-Volumen auf (d. h. 0,5 l IAA pro ml Kultur). Das macht Schritt 2.20 schneller.
- Sammeln Sie eine Probe als Schritt 2.5.
- Fügen Sie sofort IAA 0,5 l/ml Kultur zu einer Endkonzentration von 750 m hinzu, wie in Schritt 2.8 vorbereitet. Schnell mischen, indem Sie kräftig wirbeln.
- Sammeln Sie Proben, wie Schritt 2.5, nach Ihrem experimentellen Design. Entweder eine einzelne Probe zu einem Zeitpunkt, zu dem erwartet wird, dass das Protein zuverlässig erschöpft wird, oder mehrere Proben in einem Zeitverlauf der Erschöpfung. Zum Beispiel sind 5 min Intervalle bequem für Timing und bieten eine Reihe von Proteinspiegeln. Die Optimierungsstrategie, wie in Abbildung 1dargestellt, gibt einen Hinweis auf geeignete Zeiten.
- Verarbeiten Sie die Proben.
- Legen Sie die Proben auf Eis, wenn nicht bereits getan. Stellen Sie sicher, dass keine der Proben eingefroren wurde. wenn sie haben, sanft warm in der Hand, ständig invertieren, so dass die Temperatur nicht lokal steigen.
HINWEIS: Dies geschieht am besten in der Hand, da die Temperatur der Probe beurteilt werden kann, sollte es sich immer kalt anfühlen. Auf Eis legen. Dies ist kein Pausenpunkt - sobald alle Proben flüssig sind, fahren Sie mit dem nächsten Schritt fort. - Sobald alle Proben entnommen wurden und nicht mehr eingefroren sind, drehen Sie bei 3.500 x g für 2 min (bei 4 °C, wenn möglich).
- Die Methanol/Medium-Mischung abgießen und wieder auf Eis legen; Keine Sorge, wenn nicht die gesamte Flüssigkeit entfernt wurde.
- Das Zellpellet in 1 ml eiskaltH2 O (ab Schritt 2.3.3) wieder aufhängen und auf Eis in ein markiertes 1,5 ml Rohr (vorbereitet in Schritt 2.3.2) geben.
- Drehen Sie kurz (z.B. 10 s Gesamtzeit) bei >15.000 x g, um die Zellen neu zu pellet, wieder auf Eis zu legen und die Flüssigkeit zu entfernen.
- Entfernen Sie die H2O durch Aspiration. Die Zellpellets können bei -20 °C oder -80 °C zur Langzeitlagerung gelagert werden.
- Legen Sie die Proben auf Eis, wenn nicht bereits getan. Stellen Sie sicher, dass keine der Proben eingefroren wurde. wenn sie haben, sanft warm in der Hand, ständig invertieren, so dass die Temperatur nicht lokal steigen.
- Überprüfen Sie den Gehalt, auf den das Protein durch Western Blot Analysis18erschöpft wurde.
HINWEIS: Ausreichendes Protein19 und/oder Nukleinsäure kann für die meisten Zwecke aus einem einzelzelligen Pellet extrahiert werden, obwohl seltene RNA-Arten möglicherweise mehr Probenvolumen erfordern.
Ergebnisse
Repräsentative Beispiele für Erschöpfung sind in Abbildung 1dargestellt. Die drei in dieser Abbildung vorgestellten Experimente waren Optimierungsexperimente zur Erschöpfung der Proteine Prp2, Prp22 und Dcp1. Die geringe Häufigkeit, spliceosomal Prp2 und Prp22 Proteine beide erschöpft auf weniger als 20% nach 40 min Pre-Inkubation mit '-estradiol gefolgt von 15 min mit Auxin. Längere Vorinkubationszeiten führen zu einer schnelleren Erschöpfung, zeigen aber auch eine unerwünschte Proteinerschöpfung vor der Auxin-Zugabe. Im Vergleich dazu wurde die reichlichere Dcp1 nur auf etwa 30% mit der gleichen Behandlung erschöpft, aber 60 min der Vorinkubation führte zu einer Erschöpfung auf 13% mit der gleichen Auxin-Behandlung, auf Kosten der Erschöpfung, bevor das Auxin hinzugefügt wird. Es ist möglich, dass 50 min Vorinkubation mit '-estradiol und 15 min mit Auxin zu einem kürzeren Zeitpunkt ähnliche Ergebnisse erzielt hätten und damit optimaler gewesen wären.

Abbildung 1: Die Erschöpfungsrate kann durch Modulation der Dauer der Vorinkubation von '-estradiol optimiert werden. Western Blot18 der Zielproteine: (a und b) Prp22-AID*-6FLAG, (c und d) Prp2-AID*-6FLAG, und (e und f) Dcp1-AID*-6HA, aus Kulturen, die mit dem Vorbebrüter mit '-estradiol', ('-est) für 20, 30, 40, oder 60 min vor Auxin-Zugabe12. In jeder Spur wurden gleiche Mengen an Gesamtprotein geladen. Pgk1 wird als visuelles Ladesteuerelement erkannt, mit Ausnahme des Panels e, in dem Pgk1 und Dcp1 gemeinsam migrieren. Die Quantifizierung von Proteinbändern in den Paneelen a, c und e wird in den Panels b, dbzw. f dargestellt. Als Maß für die Erschöpfungsrate wurde die Steigung (m) für den linearen Schnitt (von 100 % bis 30 % der Anfangswerte) jeder Kurve berechnet. Die optimale Vorinkubationszeit ist die Zeit, in der die Proteinspiegel noch nahe an den uninduzierten Werten liegen (100%) und die anschließende Erschöpfungsrate ist schnell. Für Dcp1 (f) sind 60 min Vorinkubation zu lang, da das Protein in Ermangelung von Auxin zu abbauen begonnen hat, während 20 min zu kurz ist, da das Protein in diesem Zeitverlauf nicht spürbar erschöpft ist. Nach 40 min Vorinkubation können 15 min mit Auxin verwendet werden, da das Protein etwa 70% erschöpft ist und obwohl 20 min zu einer weiteren Erschöpfung führen würde, könnte es auch zu sekundären Effekten führen. Fehlerbalken stellen die Standardabweichung von zwei biologischen Replikationen dar. Für jedes Experiment wird ein repräsentativer Fleck angezeigt. Diese Zahl ist aus der vorherigen Veröffentlichung9abgeleitet. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

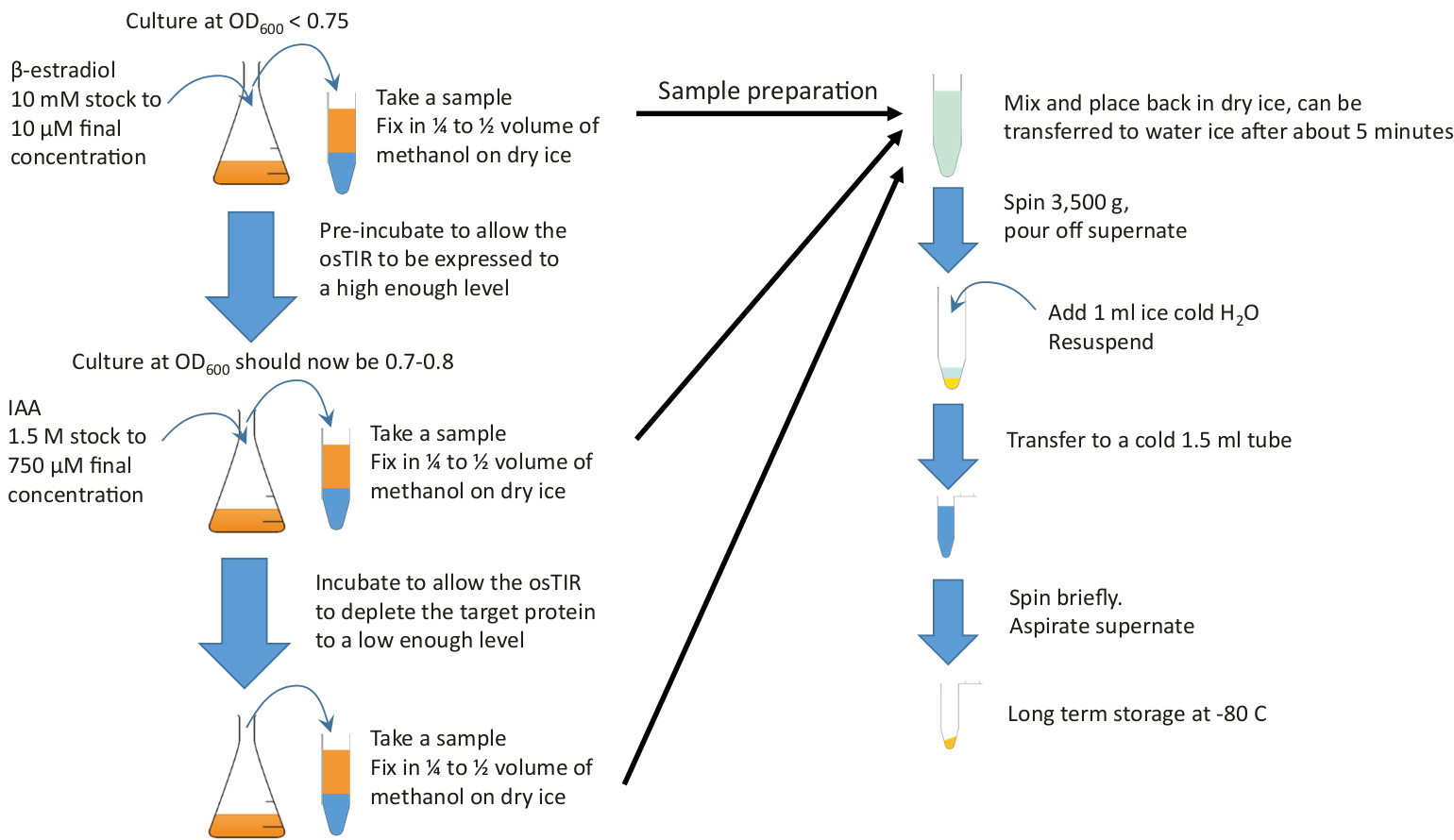
Abbildung 2: Grafische Zusammenfassung. Fügen Sie die Ausreichende Kultur, die in einem reichhaltigen Medium und bei der erforderlichen Temperatur wächst, um mit der Vorinkubation zu beginnen, zu einer ausreichenden Kultur hinzu. Setzen Sie das Wachstum für die erforderliche Vorinkubationszeit fort, bevor Sie IAA (Auxin) hinzufügen, um mit der Erschöpfung zu beginnen. Die Vorinkubations- und Erschöpfungszeiten hängen vom zu erschöpfenden Protein ab, aber die Vorinkubation liegt oft im Bereich von 20-60 min und die Erschöpfungszeit liegt in der Regel in der Größenordnung von 10 bis 20 min. 10 ml Proben sollten zu Beginn und am Ende der Vorinkubation entnommen werden und Durin g die Erschöpfung. Diese Proben werden vor dem Granulieren und Lagern schnell in kaltem Methanol fixiert. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 3: Dehnungsgenerierung für das B-est-System. (a) Um einen Hefestamm mit dem AID*-System zu erzeugen, entweder das pZTRL (LEU2) oder das pZTRK (Kanamycin (G418) Resistenz), sollte Plasmid in den Stamm eingeführt werden, oder alternativ können ATF und osTIR homologe Rekombination eines von PCR erzeugten Fragments aus 3-End-Primern (siehe Abbildung 3b und Tabelle 1). (b) C-Terminal-Tagging des Zielproteins wird durch PCR-Verstärkung des geeigneten Bereichs des Plasmids pURA3-AID*-6FLAG erreicht (pURA3_AID*-6HA unterscheidet sich nur im Tag und kann exakt auf die gleiche Weise behandelt werden), mit Longtingrundiern S3-F und S2-R mit 3' Erweiterungen homologe bis zum 3' Ende des Zielproteins. Die Vorwärtsprimer-Erweiterung sollte das letzte Aminosäure-Codon im Rahmen mit dem Start des AID*-Tags enthalten und darf das Stop-Codon nicht enthalten. Die umgekehrte Primererweiterung sollte sich auf eine Region unterhalb der Codierungsregion befinden. Einmal in das Genom eingeführt, können Zellen, die den URA3-Marker verloren haben (durch homologe Rekombination zwischen den identischen Regionen an beiden Enden des Markers), durch Wachstum mit 5-FOA ausgewählt werden, das URA3-Zellen gegenwählt. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
| a. Primer-Sequenzen | |||||||
| zielscheibe | standort | leitfaden | name | folge | Tm (°C) | ||
| pZTRL | 516 | f | pZTRL_F | <-Region der Homologie->GCGACAGCATCACCGACTTCG | 61,23 | ||
| 7897 | R | pZTR_R | CGCCGCCTCTCCTTGCAGA<-Region der Homologie (RC)-> | 61,30 Uhr | |||
| pZTRK | 9154 | f | pZTRK_F | <-Region der Homologie->ACGTTGAGCCATTAGTATCAATTTGCTTACC | 59,40 | ||
| 5897 | R | pZTR_R | CGCCGCCTCTCCTTGCAGA<-Region der Homologie (RC)-> | 61,30 Uhr | |||
| pURA3-AID*-6FLAG oder pURA3_AID*-6HA | f | S3-F | <-Region der Homologie->CGTACGCTGCAGGTCGAC | 59,21 | |||
| R | S2-R | ATCGATGAATTCGAGCTCG<-Region der Homologie (RC)-> | 52,76 | ||||
| pZRTL/K soll das AID-System | |||||||
| pURA3-AID*-6FLAG/6HA zur Verstärkung des AID*- und Epitop-Tags zur Kennzeichnung des Zielproteins (Lontinverfahren) | |||||||
| <-Region der Homologie-> | Region homologe zu den flankierenden Regionen, in die das System eingefügt werden soll. Je länger diese Region ist, desto wahrscheinlicher ist es, dass die Änderung erfolgreich ist; 50 - 100 Basen werden empfohlen. | ||||||
| <-Region der Homologie (RC)-> | Region homologe zu den flankierenden Regionen, in die das System eingefügt werden soll, denken Sie daran, die umgekehrte Ergänzung zu verwenden. Je länger diese Region ist, desto besser. | ||||||
| Tm (°C) | Tm mit der %GC Methode mit 50 mM NaCl | ||||||
| b. PCR-Mix | |||||||
| teil | Volumen (L) | ||||||
| schablone | <10 | ||||||
| NEB Phusion HF Puffer (5x)* | 100 | ||||||
| Vorwärtsgrundierung 100 m | 2,5 | ||||||
| Reverse Primer 100 m | 2,5 | ||||||
| dNTPs je 10 mM | 10 | ||||||
| H2O | bis 500 | ||||||
| * Der NEB Phusion GC Buffer (5x) kann auch verwendet werden, wird aber nicht bevorzugt | |||||||
| Machen Sie diese Mischung, aufgeteilt in 10 Röhren von je 50 l mischen und führen Sie die PCR als Tabelle 1 c. | |||||||
| Überprüfen Sie, ob die PCR funktioniert hat, indem Sie auf einem Agarose-Gel ausgeführt werden | |||||||
| Kombinieren Sie alle erfolgreichen Reaktionen in einer Röhre und Ethanol ausgefällt | |||||||
| Transformieren Sie die Hefe mit dem gesamten Material, das von der PCR produziert wird | |||||||
| c. PCR-Bedingungen | |||||||
| schritt | Temperatur (°C) | zeit | |||||
| Anfängliche Denaturierung | 98 | 30 s | |||||
| 25-35 Zyklen | Denature | 98 | 10 s | ||||
| Anneal | 45–60 | 20 s | |||||
| anbau | 72 | 30 s/kb | |||||
| Endgültige Verlängerung | 72 | 10 Min. | |||||
| halten | 8 | ||||||
| Neal bei 45 °C für das Lontine Primer Set (S3-F und S2-R) und 60 °C für die pZTRL/K Primer | |||||||
| Verlängern Sie für die Lontine PCR 3 Minuten und für pZTRL/K 3 Minuten | |||||||
Tabelle 1: Primersequenzen, PCR-Mix und PCR-Bedingungen.
Diskussion
Ein gut optimiertes Protokoll kann zu einer schnellen und effizienten Erschöpfung des Zielproteins führen. Die Bestimmung der ungefähren Vorinkubationszeit mit dem Estradiol ist wichtig, da dies die Reproduzierbarkeit der Erschöpfung erhöht, aber kleine Schwankungen der Vorinkubationszeit toleriert werden können. Auf der anderen Seite ist beim Timing nach Auxin-Zugabe Vorsicht geboten, da der Proteinspiegel sehr schnell abnimmt.
Ein Vorteil dieses Ansatzes besteht darin, dass eine abgestimmte Erschöpfung durch unterschiedliche Kombinationen der Vorinkubationszeit mit der Inkubationszeit von '-estradiol und iAA erreicht werden kann. Wenn gewünscht, kann das Zielprotein z. B. langsamer aufgebraucht werden, indem die Vorinkubationszeit reduziert wird.
Das AID-System bietet bestimmte Vorteile gegenüber Systemen, bei denen OsTIR konstitutiv ausgedrückt wird. Wenn das Zielprotein beispielsweise für die Lebensfähigkeit unerlässlich ist, kann eine regulierte Expression von osTIR eine vorzeitige Erschöpfung des Zielproteins vermeiden. Darüber hinaus kann die Expression von osTIR auf die Fülle des Zielproteins und seine Erniedrigungsanfälligkeit abgestimmt werden, und die Erschöpfung kann entweder schnell oder langsam sein. Die beiden kleinen Moleküleffektoren, '-estradiol und auxin, stören den Hefestoffwechsel unter den hier verwendeten Bedingungen nicht, im Gegensatz zu Rapamycin, das im Anker-Auswärtssystem verwendet wird1.
Es sollte beachtet werden, dass das Markieren einiger Proteine ihre Funktion stört, was ein Problem mit jedem gezielten Erschöpfungssystem ist. In diesem Fall kann ein N-Terminal-Tag funktionieren, wenn ein C-Terminal-Tag dies nicht tut. Außerdem werden nicht alle Proteine effizient aufgebraucht sein; Beispielsweise kann das AID-Tag auf dem Zielprotein für das osTIR-Protein unzugänglich sein. Daher sollte nach dem AID-Tagging jedes Zielprotein auf jede Wirkung des Tags auf das Wachstum getestet werden, und um festzustellen, ob die Erschöpfung wirksam ist, bevor die Timings der Vorinkubation und Auxin-Behandlung optimiert werden.
Dieses AID*-System ist sehr einfach und mit allen nachfolgenden experimentellen Verfahren kompatibel, die kein weiteres Wachstum mit sich bringen, wie Protein-, DNA- oder RNA-Analyse oder Mikroskopie. Darüber hinaus funktioniert das System gut in Kombination mit Thiolabelling, um entstehende RNA20zu reinigen.
Dieses System bietet ein schnelles, spezifisches und reproduzierbares Mittel zur Erschöpfung eines Proteins, ohne den Stoffwechsel der Hefezelle anderweitig zu beeinträchtigen.
Offenlegungen
Die Autoren haben nichts zu verraten.
Danksagungen
Vielen Dank an Jane Reid für die Initiierung dieses Programms, Barbara Terlouw für die Entwicklung, Vahid Aslanzadeh für die "ura looper" Konstrukte und Susana de Lucas für viele hilfreiche Diskussionen. Diese Arbeit wurde durch ein Stipendium an GIMO vom Consejo Nacional de Ciencia y Tecnologa, Mexiko (CONACYT) und der University of Edinburgh School of Biological Sciences, eine Wellcome PhD-Studentschaft an IEM [105256] und durch Wellcome-Förderung [104648] an JD Beggs unterstützt. . Die Arbeit im Wellcome Centre for Cell Biology wird durch Wellcome-Kernförderung unterstützt [092076].
Materialien
| Name | Company | Catalog Number | Comments |
| Adenine sulphate | Formedium | DOC0230 | |
| Agar | Formedium | AGA03 | |
| Β-estradiol | Sigma Aldrich | E2758-1G | 10 mM solution in ethanol. Store at -20 °C |
| DMSO | Alfa Aesar | 42780 | DMSO should be solid at 4 °C |
| Glucose | Fisher Scientific | G/0500/60 | |
| IAA 1H-Indole-3-acetic acid | Across Orgainics | 122150100 | Auxin analogue. 1.5 M in DMSO. The solution will be a russet colour and darken as time goes on; a deep red solution should be discarded and a new one made. Store at -20 °C. |
| Methanol | Fisher Scientific | M/4000/PC17 | CAUTION Toxic and flammable |
| Phusion High-Fidelity DNA Polymerase | NEB | M0530 | |
| Peptone | Formedium | PEP03 | |
| SCSM single drop-out –ura | Formedium | DSCS101 | |
| Yeast Extract | Formedium | YEA03 | |
| Yeast nitrogen base without amino acids with amonium sulphate | Formedium | CYN0410 |
Referenzen
- Haruki, H., Nishikawa, J., Laemmli, U. K. The Anchor-Away Technique: Rapid, Conditional Establishment of Yeast Mutant Phenotypes. Molecular Cell. 31, 925-932 (2008).
- Bellí, G., Garí, E., Piedrafita, L., Aldea, M., Herrero, E. An activator/repressor dual system allows tight tetracycline-regulated gene expression in budding yeast. Nucleic Acids Research. 26, 942-947 (1998).
- Alexander, R. D., et al. RiboSys, a high-resolution, quantitative approach to measure the in vivo kinetics of pre-mRNA splicing and 3′-end processing in Saccharomyces cerevisiae. RNA. 16, 2570-2580 (2010).
- Deshaies, R. J., Joazeiro, C. A. P. RING Domain E3 Ubiquitin Ligases. Annual Review of Biochemistry. 78, 399-434 (2009).
- Tan, X., et al. Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature. 446, 640-645 (2007).
- Teale, W. D., Paponov, I. A., Palme, K. Auxin in action: signalling, transport and the control of plant growth and development. Nature Reviews Molecular Cell Biology. 7, 847-859 (2006).
- Nishimura, K., Fukagawa, T., Takisawa, H., Kakimoto, T., Kanemaki, M. An auxin-based degron system for the rapid depletion of proteins in nonplant cells. Nature Methods. 6, 917-922 (2009).
- Morawska, M., Ulrich, H. D. An expanded tool kit for the auxin-inducible degron system in budding yeast. Yeast. 30, 341-351 (2013).
- Kubota, T., Nishimura, K., Kanemaki, M. T., Donaldson, A. D. The Elg1 Replication Factor C-like Complex Functions in PCNA Unloading during DNA Replication. Molecular Cell. 50, 273-280 (2013).
- Brosh, R., et al. A dual molecular analogue tuner for dissecting protein function in mammalian cells. Nature Communications. 7, 11742 (2016).
- DeRisi, J. L., Iyer, V. R., Brown, P. O. Exploring the metabolic and genetic control of gene expression on a genomic scale. Science. 278, 680-686 (1997).
- Mendoza-Ochoa, G. I., et al. A fast and tuneable auxin-inducible degron for depletion of target proteins in budding. Yeast. , (2018).
- McIsaac, R. S., et al. Synthetic gene expression perturbation systems with rapid, tunable, single-gene specificity in yeast. Nucleic Acids Res. 41, e57 (2013).
- Prusty, R., Grisafi, P., Fink, G. R. The plant hormone indoleacetic acid induces invasive growth in Saccharomyces cerevisiae. PNAS. 101, 4153-4157 (2004).
- Geitz, D., St Jean, A., Woods, R. A., Schiest, R. H. Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Research. 20, 1425 (1992).
- Widlund, P. O., Davis, T. N. A high-efficiency method to replace essential genes with mutant alleles in yeast. Yeast. 22, 769-774 (2005).
- Longtine, M. S., et al. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast. 14, 953-961 (1998).
- Eaton, S. L., et al. A Guide to Modern Quantitative Fluorescent Western Blotting with Troubleshooting Strategies. Journal of Visualized Experiments. , e52099 (2014).
- Volland, C., Urban-Grimal, D., Géraud, G., Haguenauer-Tsapis, R. Endocytosis and degradation of the yeast uracil permease under adverse conditions. Journal of Biological Chemistry. 269, 9833-9841 (1994).
- Barrass, J. D., et al. Transcriptome-wide RNA processing kinetics revealed using extremely short 4tU labeling. Genome Biology. 16, 282 (2015).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten