
Method Article
Cultivos Enriquecidos con Conos De La Retina De Embriones De Pollo Para Estudiar Las Interacciones Celulares De Varilla A Cono
En este artículo
Resumen
Describimos un método para obtener cultivos primarios de fotorreceptores de cono de la retina de embriones de pollo y su uso para la detección de alto contenido.
Resumen
La visión diurna humana se basa en la función de los fotorreceptores de cono en el centro de la retina, la fóvea. Los pacientes que sufren de la forma más frecuente de degeneración retiniana heredada, retinosis pigmentaria, pierden la visión nocturna debido a la pérdida impulsada por mutaciones de fotorreceptores de varilla, un fenómeno seguido por una pérdida progresiva de la función y la muerte de los conos que conducen a la ceguera. Los genetistas han identificado muchos genes con mutaciones causantes de esta enfermedad, pero las primeras mutaciones identificadas cuestionaban los mecanismos de degeneración del cono secundario y cómo una mutación dominante en el gen de la rodopsina que codifica para el pigmento visual expresado exclusivamente en varillas puede desencadenar la degeneración del cono.
Este resultado de trasplantes en un modelo genético de la enfermedad llevó al concepto de interacciones celulares entre varillas y conos y de degeneración autónoma no celular de conos en todas las formas genéticas de retinosis pigmentaria.
Los conos comprenden el 5% de todos los fotorreceptores en los seres humanos y solo el 3% en el ratón, por lo que su estudio es difícil en estas especies, pero los conos superan en número a las varillas en las especies de aves. Hemos adaptado placas de 96 pozos para cultivar precursores de retina de la retina de embriones de pollo en la etapa 29 de su desarrollo. En estos cultivos primarios, los conos representan el 80% de las células después de la diferenciación in vitro. Las células degeneran durante un período de una semana en ausencia de suero. Aquí, describimos los métodos y su estandarización.
Este sistema cono-enriquecido de la cultura fue utilizado para identificar el factor epitelio-derivado de la viabilidad del cono (EdCVF) por la investigación de alto contenido de una biblioteca pigmentada retiniana del cDNA del epitelio de la rata normalizada. EdCVF recombinante previene la degeneración de los conos.
Introducción
La retina de las especies de vertebrados es dual, con fotorreceptores de varilla para la visión de luz tenue y fotorreceptores de cono para la luz del día, el color y la visión de agudeza. La agudeza visual de los primates se basa en una región en el centro de la retina, llamada fóvea, que está enriquecida en conos, pero en general, los conos representan solo el 5% de todos los fotorreceptores. En consecuencia, el análisis de los conos en la retina de los primates y especialmente el cultivo de conos son técnicamente difíciles. Todas las demás especies de mamíferos no tienen fóvea y el porcentaje de conos es bajo para los roedores que se utilizan más comúnmente en la investigación de la retina. Este no es el caso de las especies aviares, para las que los conos dominan la retina de estas especies de aves que ven bien. Los dinosaurios, que han dominado el ecosistema cuando los mamíferos aparecieron por primera vez durante la evolución, están en el origen filogenético de las aves1. Como consecuencia de tal competencia entre los dinosaurios y los primeros mamíferos, los mamíferos son en su mayoría nocturnos con retinas dominadas por varillas. Sólo más tarde durante la evolución la visión diurna de algunas especies de mamíferos, entre las que pertenecen los primates, se convirtió en una ventaja evolutiva. Sin embargo, el período ancestral permanece como un atavismo del cuello de botella nocturno en la evolución de la visión de los mamíferos2,3.
Mientras estudiaban la diferenciación celular retiniana, Adler y Hatlee demostraron que los fotorreceptores representan aproximadamente el 70% de las células diferenciadas de la retina en cultivos derivados del pollo en el día embrionario (DE) 6 o en la etapa 294. Debido a la prevalencia de conos en la retina del pollo, se han desarrollado cultivos de células retinianas a partir de embriones de pollo ED6 como cultivos enriquecidos con conos5.
La importancia de la agudeza visual mediada por cono para el ser humano es una obviedad. Las personas afectadas por enfermedades genéticas o de envejecimiento que alteran la función del cono están muy perjudicadas. Esto ha promovido un cuerpo muy amplio de estudios sobre degeneraciones hereditarias de la retina (IRD) con el objetivo de encontrar tratamientos para estas enfermedades cegantes6,7. El primer éxito, obtenido utilizando un vector adenoasociado recombinante (AAV) para la terapia de una forma grave de amaurosis congénita de LEBER (ACV) ird, es una prueba de concepto para la terapiagénica 8. La identificación de los genes cuyas mutaciones desencadenan la IRD abre la posibilidad de curar estas enfermedades mediante terapia génica. Sin embargo, estas enfermedades son el resultado de mutaciones en más de 200 genes distintos9. Incluso en el caso de las formas autosómicas recesivas de la IRD, cuando la reintroducción de la copia normal del gen mórbido podría restaurar la función visual, el coste económico de cada desarrollo individual favorece a los más prevalentes en detrimento de los menos comunes y a aquellos para los que el origen genético sigue siendo desconocido. Este hecho llevó a los investigadores a pensar en terapias más generales. La muerte celular apoptótica apareció como una vía común, y una diana terapéutica de estas enfermedades que progresan por la degeneración de los fotorreceptores, incluso para las formas autosómicas dominantes10,11. Sin embargo, faltan los éxitos de este enfoque. Para la forma más común de IRD, la retinosis pigmentaria (RP), la vía común es la pérdida secundaria de la función seguida en última instancia por la degeneración de los conos12,13. La prevención de la pérdida de la función cono preservará la visión central de la fóvea independientemente de las mutaciones causantes14.
En la etapa temprana de la RP, la pérdida de varillas desencadena una reducción en la expresión del factor de viabilidad del cono derivado de la varilla (RdCVF), codificado por el gen 1(NXNL1)similar a la nucleoredoxina, que interrumpe la señalización metabólica y redox entre las varillas y los conos15. La administración de un AAV recombinante que codifica los dos productos del gen NXNL1, el factor trófico RdCVF y la enzima tiorredoxina RdCVFL, podría teóricamente prevenir la pérdida de la visión cónica en todas las formas genéticas de RP16. Hemos demostrado que el producto del gen NXNL1, RdCVFL, se expresa en cultivos enriquecidos con cono de pollo17 y donde desempeña un papel protector18. RdCVF y el gen NXNL1 fueron identificados por la detección de alto contenido de una biblioteca de ADNc retiniano utilizando la supervivencia de células de un cultivo enriquecido con cono como lectura19. Se cribó el equivalente a 210.000 clones individuales de la biblioteca utilizando 8 pruebas paralelas para cada clon. Esto representa un gran número de pruebas que requieren un fácil acceso al material biológico, las retinas de los embriones de pollo. Descubrimos que era relativamente fácil obtener huevos de gallina embrionados semanalmente porque se producen ampliamente para la agroindustria de gallinas ponedoras de huevos y pollos productores de carne. Después de una cuidadosa estandarización de los cultivos enriquecidos con conos, el sistema proporciona una forma fácil, robusta y reproducible de probar miles de moléculas por su capacidad para preservar la viabilidad del cono. Estas células también son susceptibles a manipulaciones genéticas20 que benefician al estudio de la transducción de señales y a análisis bioquímicos21,22,23.
Investigadores de retina han desarrollado métodos alternativos como el uso de la línea celular de cono 661W24,25,26. Sin embargo, la identidad de esta línea celular sigue siendo controvertida27,28. Las células 661W fueron reproducidas de tumores retinianos de una línea transgénica del ratón que exprese el antígeno grande SV40 de T bajo control del promotor retinol-obligatorio humano de la proteína del interphotoreceptor. El antígeno T grande SV40 media la transformación celular y la inmortalización. Como consecuencia, la vía de señalización identificada mediante células de 661W debe informarse en el contexto de una línea celular transformada e inmortalizada que es distinta de muchas maneras de los conos in situ. En ese sentido, el sistema de cultivo enriquecido con cono está compuesto por neuronas primarias, los conos que son más relevantes fisiológicamente.
Si bien es posible obtener un cultivo puro de fotorreceptores utilizando el seccionamiento de vibratomas de la retina del ratón, el muy bajo porcentaje de conos en la retina externa de los roedores hace que este enfoque no sea adecuado para producir cultivos enriquecidos con conos29. La retina del cerdo no contiene fóvea pero tiene una región llamada área central que está muy enriquecida en conos30. La alta proporción de conos en la retina diurna roedores, como Arvicanthis ansorgei y Psammomys obsesus31,32,ofrece una posible solución, pero requiere la cría de este tipo de especies exóticas. Los ojos de cerdo adulto, recogidos de mataderos locales, pueden ser utilizados para producir un cultivo mixto de varillas y conos que han sido utilizados para estudiar la supervivencia de los fotorreceptores33. Una solución elegante es pre-purificar los conos de la retina del cerdo usando el paneo con la lectina de la aglutinina del cacahuete (PNA), que se une selectivamente a los conos34. Sin embargo, este método es difícil de implementar a gran escala debido a su complejidad.
Las células madre pluripotentes inducidas por el hombre (iPS) ofrecen el enfoque más prometedor para obtener una población de células fotorreceptoras de cono que se puede utilizar para el trasplante de retina, pero que también se puede adaptar al cultivo enriquecido con cono35,36. Dado que el factor de transcripción NRL es necesario para los fotorreceptores de varilla37,el ratón Nrl/-tiene una retina dominada por conos de onda corta (S-conos). La inactivación podría utilizarse para producir una preparación enriquecida con S-cono por diferenciación de humanos de iPS38,39. Otro posible enfoque es promover la diferenciación de conos utilizando la señalización de hormona tiroidea40. Mientras que están surgiendo nuevos métodos para producir cultivos enriquecidos con conos a partir de iPS humanas, los embriones de pollo proporcionan un método probado actual19.
El cultivo enriquecido con cono fue instrumental en la identificación de RdCVF por clonación de expresiones19. Este sistema también fue utilizado con éxito para demostrar que rdCVF estimula la captación de glucosa y su metabolismo por glucólisis aeróbica22. Además, se utilizó un cultivo enriquecido con cono para validar el papel protector de RdCVFL, el segundo producto del gen NXNL1 23. Más recientemente, este sistema fue utilizado para demostrar la existencia de moléculas protectoras secretadas por células epiteliales pigmentadas de la retina transducidas con OTX241.
Protocolo
El protocolo fue aprobado por el Comité de Ética de los Experimentos con Animales de la Universidad Pierre y Marie Curie y el Ministerio de Investigación francés (Número de permiso: APAFIS #1028 2015070211275177). Los experimentos con animales se realizaron bajo la siguiente autorización: "Certificat d'autorisation d'expérimenter sur les animaux vertébrés A-75-1863. Préfecture de Police de Paris (9 de noviembre de 2011-8 de noviembre de 2016)".
1. Incubación de huevos fertilizados
- Recoger semanalmente los huevos fertilizados (cepa I 657, etiqueta roja), obtenidos de forma natural, en una planta de incubación industrial.
- Mantener los huevos fertilizados a 17°C (su cero biológico) en el laboratorio después de que sean "puestos" por la gallina.
- Para cada cultivo, incubar siete huevos fertilizados durante 24 horas a 20 °C y luego 136 horas a 37 °C con reversión intermitente de la inclinación (un movimiento progresivo durante 2 horas de un lado a su opuesto, un ciclo de 4 horas) de los huevos en una cámara humidificada.
2. Recuperación de los embriones de pollo
- Lave la superficie de los siete huevos con desinfectante (por ejemplo, Pursept A express).
- Para romper la cáscara de huevo, haga un agujero en la parte superior de la cáscara con grandes alicates rectos. Luego corte la cáscara para quitar el sombrero del huevo como un huevo hervido suavemente.
- Extraiga suavemente cada embrión de la cáscara de huevo con fórceps curvos, y luego transfilícelo a una placa de Petri que contenga solución salina tampón de fosfato estéril (PBS) previamente calentada a 37 °C. Suavemente, retire la envoltura que rodea a los embriones (el corion o la membrana corioalantólica).
- Verificar la etapa de desarrollo de cada embrión mediante comparación visual con Hamburger y Hamilton42.
- Seleccione dos embriones en la 29ª etapa de desarrollo (Figura 1). Las alas se doblan en los codos. El collar destaca visiblemente. El proyecto de ley es más prominente que en la etapa 28.
- Enuclear los ojos de estos embriones seleccionados y transferirlos en co2-medio independiente (Tecnologías de vida).
PRECAUCIÓN: Es muy importante que el embrión esté en la etapa 29, y no en la etapa 28 o 30(Figura 1); es por eso que es necesario incubar al menos 7 huevos, incluso si solo se usarán finalmente dos. El corion es muy delgado, tan difícil de distinguir, pero está muy cerca del embrión, por lo que tiene que ser retirado sin tocar el embrión.
3. Disección de las retinas
- Decapitar y enuclear los embriones seleccionados con fórceps curvos.
- Transfiera los cuatro ojos al medio independiente de CO2. Este medio contiene 0,9 mM de CaCl2 y 0,65 mM de MgCl2.
- Coloque el ojo con la córnea boca abajo, el nervio óptico frente al experimentador. Perfore un agujero en el nervio óptico usando dos tórceps rectos.
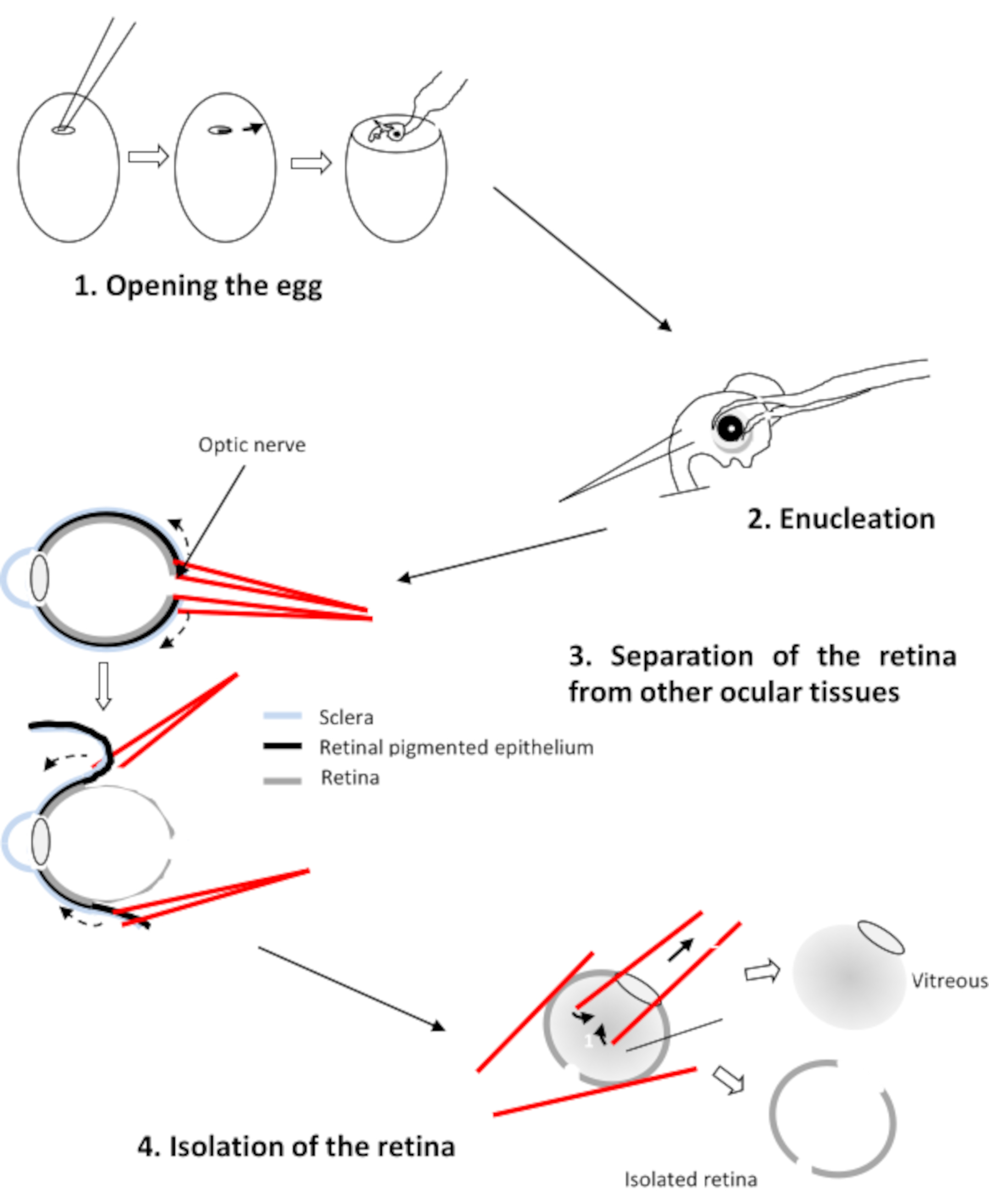
- Insertar una rama de cada fórceps entre la retina y el epitelio pigmentario (Figura 2). Tire de cada estratego y gire el ojo para separar el epitelio de la retina. Retire la córnea seguida del cristalino y el vítreo.
- Transfiera las cuatro retinas en una placa de Petri que contenga el medio de Ringer a pH 7.2.
PRECAUCIÓN: Asegúrese de que sólo queda la retina y eliminar cualquier rastro de epitelio pigmentado vítreo y restante de la retina.
4. Preparación de la suspensión celular de la retina
- Cortar las cuatro retinas en trozos muy pequeños con dos alicates rectos.
- Lave las piezas retinianas dos veces con el medio de Ringer.
- Después del segundo lavado con el medio del Campanero, deje que los pedazos de retina caigan en la parte inferior del tubo y retire el medio del Campanero. Trate las piezas retinianas durante 20 minutos a 37 °C con una solución de tripsina (0,25% p/v).
- Dispersar la solución después de 10 minutos por succión sucesiva. Descarga con una pipeta Pasteur y comprueba la disociación de las piezas retinianas. Detenga la reacción agregando medios de cultivo complementados con suero de ternero fetal inactivado al 10%.
- Incubar la suspensión celular con 0,05 mg de DNasa I. Disociar los grupos celulares y el ADN mediante succión y descarga sucesivas utilizando una pipeta Pasteur inmediatamente después de añadir la DNasa.
- Lave la suspensión de células retinianas dos veces con un medio de cultivo definido químico (CDCM): un volumen igual de medio eagle modificado de Dulbecco y medios M199 suplementados con ácido linoleico de 100 μg/mL/BSA, 0,86 μM de insulina, 0,07 μM de transferrina, 2,0 μM de progesterona, 0,28 μM de prostaglandina, 0,29 μM Na2SeO3,182 μM de putrescina, 3 mM de taurina, 4,7 μM de citidina 5′-difosfocolina, 2,7 μM citidina 5′-difosfoetanolamina, 0,55 μM de hidrocortisona, 0,03 μM de triyodotironina, piruvato de sodio de 1 mM y gentamicina de 20 μM.
5. Siembra de células retinianas
- Trate dos placas de cultivo negras de 96 pocillos con fondo transparente durante 2 horas a 37 °C con poli-L-lisina a 32,25 μg/cm2.
- Enjuague estas placas dos veces con el medio de cultivo M199. Resuspend la pelotilla de la célula en 1 mL de CDCM.
- Añadir a una alícuota de 10 μL de la suspensión celular trypan azul para teñir las células vivas. Añadir la muestra de suspensión de células a un hemocitómetro (cámara de recuento de células de Malassez).
- Bajo un microscopio, cuente las células teñidas para cuatro filas del hemocitómetro (es decir, 40 cuadrados) y luego calcule el número promedio de celdas para una fila. Calcular la concentración de células de la suspensión aplicando el siguiente método: Número de células/mL de suspensión = número medio de células seguidas (10 cuadrados) x 10 el número total de cuadrados del hemocitómetro x dilución con tripano azul x 1.000 (para expresar el resultado como células/mL).
- Llevar la suspensión celular a dos concentraciones (5,6 x 104 células/mL y 1,12 x 105 células/mL) correspondientes a las dos densidades de chapado (1 x 105 células/cm2 y 2 x 105 células/cm2) utilizando CDCM.
- Semilla de 50 μL de las suspensiones de dos células en las dos placas de cultivo de 96 pozos negras pretratadas. Distribuir las células de las placas con una pipeta multicanal desde la derecha de la placa hacia la izquierda, homogeneizando entre cada columna, de forma que la distribución de las células sea homogénea.
- Añadir 50 μL de la biblioteca de moléculas (por ejemplo, el medio acondicionado de una biblioteca de ADNc, ver más abajo) para ser tamizado utilizando un patrón predefinido (Tabla 1).
- Incubar las placas durante siete días a 37°C bajo 5% de CO2 sin cambio de medio.
6. Contar células viables
- A cada pozo de la placa, añadir 2,7 μM de calcina AM y 0,3 mM de homodímero de etidio.
NOTA: La calcina penetra en las células que son impermeables a moléculas grandes como el homodímero de etidio. En el citoplasma de las células vivas, la calcina es hidrolizada por esterasas endógenas, se vuelve fluorescente al emitir a 520 nm cuando se excita a 485 nm. El homodímero del etidio se une al ADN en las células muertas. La unión ADN-homodímero de etidio hace que la fluorescencia roja se emita a 635 nm después de la excitación a 520 nm. - Incubar las placas durante 1 hora a temperatura ambiente en ausencia de luz.
- Lea la fluorescencia en un lector de placas automatizado compuesto por un microscopio invertido equipado con una lámpara de mercurio con dos filtros de excitación a 485 y 520 nm, dos filtros de emisión a 520 y 635 nm, un objetivo (x10), una etapa motorizada controlada por un procesador y una cámara de dispositivo de carga acoplada (CCD). Esta plataforma de conteo está controlada por el software Metamorph19. Permite adquirir imágenes de fluorescencia de células vivas y células muertas simultáneamente en cada pozo de la placa de 96 pozos, a partir de los pozos A1 hacia el pozo A12, luego B12 hacia B1, C1 hacia C12 y así sucesivamente (Tabla I).
- Calcular el área media A de una sola celda utilizando los 18 pozos de los controles negativos (Tabla I).
- Cuente las celdas en cada pozo de la placa y aplique la siguiente fórmula empírica A x 29 / 20.7 para evitar que se cuenten los dobletes celulares (agrupación de dos celdas). Puntuar el efecto protector de los conos por moléculas como la relación entre el número medio de células en los 4 pocillos donde probamos la molécula frente al número medio de células en los 18 pocillos del control negativo (ver Tabla 1 y Figura Suplementaria 1).
- Combinar los resultados de la placa sembrada a 1 x 105 células/cm2 con la sembrada a 2 x 105 células/cm2 para evaluar la protección potencial (relación de viabilidad) por cada molécula tamizado.
Resultados
Describimos aquí cómo el sistema de cultivo enriquecido con cono se puede utilizar para identificar nuevas proteínas protectoras de conos. Utilizamos este protocolo para examinar una biblioteca normalizada del cDNA hecha del epitelio pigmentado coroides y retiniano a partir de 400 ojos de 8 ratas de Largo-Evans de las semanas43.
Esta biblioteca contiene 6,0 x 106 colonias independientes formando unidades (UFC) y tiene un tamaño medio de inserción clonada de 2,1 kilobases (kb), con más del 99% de clones recombinantes. Las piscinas de 100 copias de esa biblioteca fueron transfectadas transitorio (0,1 μg de la DNA del plásmido) en las células COS-1 y el medio condicionado (CM) de COS-1 fue cosechado después de la incubación por 48 horas en DMEM sin el suero. Las membranas o los exosomas no fueron quitados por la ultracentrifugación. Se añadieron cincuenta microlitros de cada CM a 4 pozos de dos placas de 96 pozos: uno sembrado a 2 x 105 células/cm2, el otro a 4 x 105 células/cm2. Cm de células COS-1 transfectadas con el vector vacío pcDNA3.1 utilizado para construir la biblioteca se utilizó como control negativo (Tabla 1). Se evaluaron un total de 2.112 conjuntos de 100 clones correspondientes a 211.200 clones individuales en cuatro pozos de cultivo y para dos condiciones de siembra de cultivos enriquecidos con conos. Las dos condiciones corresponden a dos densidades de inoculación ligeramente diferentes, lo que permite evaluar la actividad protectora con mayor precisión, para un total de 1.689.600 pozos de cultivo.
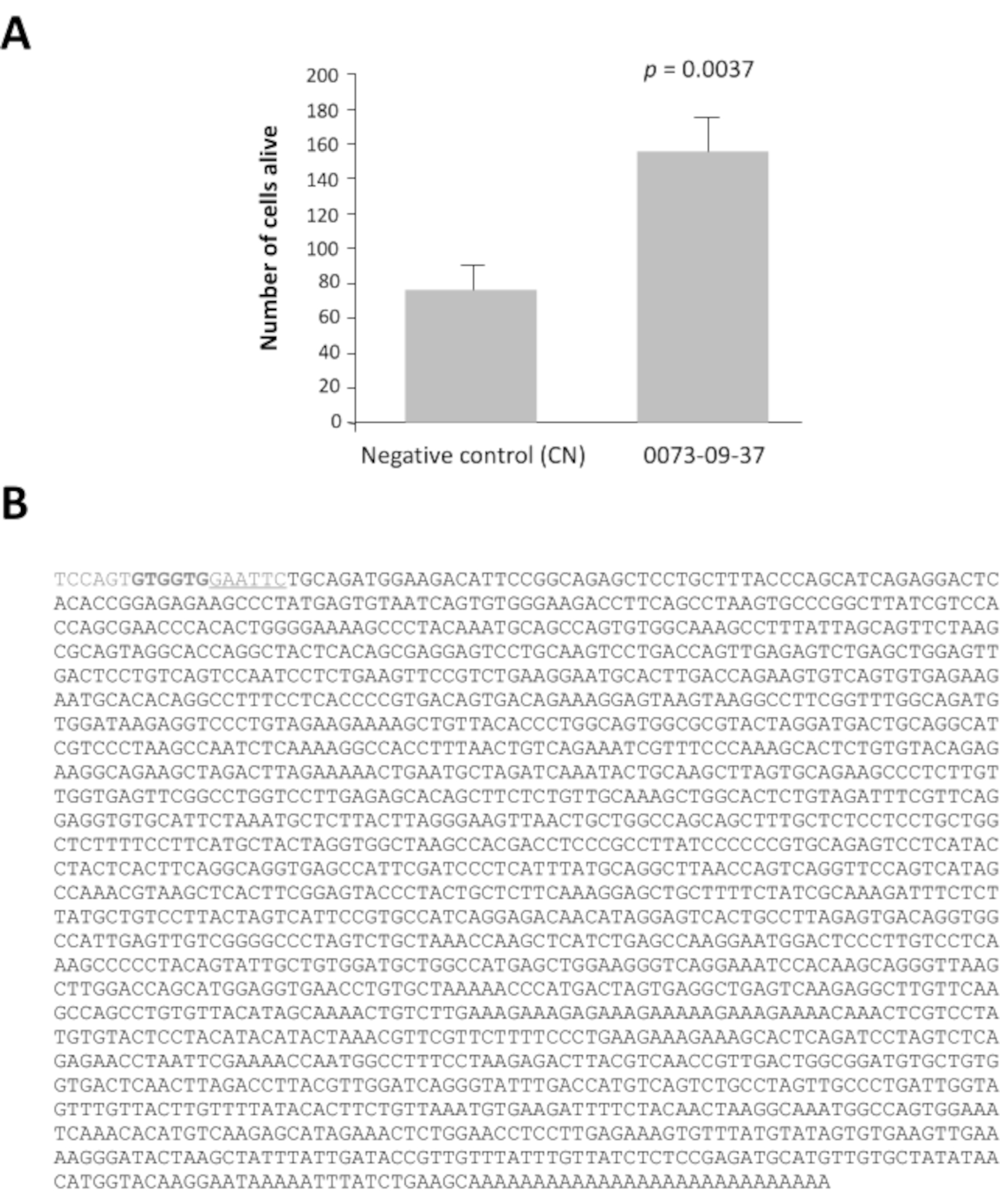
Entre las 42 piscinas de clones con una relación superior a 2, las piscinas 0080 y 0073 tienen una relación de viabilidad 16 y 14 veces mayor después de 7 días de cultivo que el control negativo, pcDNA3.1(Figura suplementaria 1). Este análisis es esencial para identificar los fondos de interés. Cada grupo seleccionado de 100 clones se subdividió en 16 conjuntos de 10 clones de su stock de glicerol. Estos subgrupos se prepararon y probaron de acuerdo con el mismo método en una segunda ronda de selección (es decir, un total de 3.200 pozos de cultivo). El subgrupo 0073-09 dio la relación de viabilidad más fuerte(Figura suplementaria 2A)y se subdividió para producir 16 clones individuales que se probaron en una tercera ronda de cribado en cultivos enriquecidos con conos. El clon 0073-09-37 se destaca claramente de los demás con una relación de viabilidad igual a 2,5 (Figura suplementaria 2B). El eje y tiene una escala diferente incluso si la densidad de siembra era la misma que en la figura suplementaria 2A. Hemos visto esto comúnmente cuando los ensayos se repiten semanalmente durante meses. Después del análisis, estos resultados confirman que el clon 0073-09-37 tiene un efecto robusto y reproducible sobre la supervivencia del cono. La prueba se repitió de forma independiente (Figura 3A), y se secuencia el inserto de 1,8 kb (Figura 3B).
Un análisis bioinformático reveló que el clon 0073-09-37, que llamamos factor de viabilidad de cono derivado del epitelio (EdCVF), contiene tres marcos de lectura abiertos (ORFs), el que más aguas arriba (ORF1) codifica para 84 residuos de la parte C-terminal de la proteína de rata zinc finger protein-180 (ZFP180, NP_653358) de 727 aminoácidos44. Los otros dos ORF (ORF2 y ORF3) están mucho menos bien conservados en ratones y ausentes en otros mamíferos. Cuando se prueba de forma independiente, sólo ORF1 ejerce un efecto protector sobre los conos (Figura suplementaria 3). ORF1 se produjo como una proteína de fusión de glutatión S-transferasa (GST)(Figura 4A). Se purificó la proteína EdCVF y se eliminó la etiqueta GST (Figura 4B). EdCVF es capaz de prevenir la degeneración del cono en el sistema de cultivo enriquecido con cono (Figura 4C).

Figura 1:Embriones depollo en las etapas 28,29y 30de desarrollo. Flechas W: ala, C: cuello y B: pico. Barra de escala 1 cm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2:Disección de la retina del embrión de pollo Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3:Factor deviabilidad del cono derivado del epitelio (EdCVF), clon 0073-09-37. A. Aumento de la viabilidad en el cultivo enriquecido con cono. B. La secuencia del clon de ADNc 0073-09-37. La secuencia subrayada GAATTC es el sitio de restricción EcoRI utilizado para construir la biblioteca. Los dos codones GTG en negrita son sitios poco comunes de iniciación de traducción para EdCVF que se originan en el vector. Análisis estadístico por prueba de Estudiante. Haga clic aquí para ver una versión más amplia de esta figura.
{kind=link}

Figura 4:Actividad recombinante de EdCVF. A. Secuencia de EdCVF en la fusión con glutatión S-transferasa (GST). B. La proteína EdCVF recombinante purificada. C. Actividad trófica de GST-EdCVF en el cono en cultura. Análisis estadístico mediante la prueba de Tukey. Haga clic aquí para ver una versión más amplia de esta figura.
{kind=link}
Figura suplementaria 1: Relación entre el número medio de células para un grupo de ADNc y el control negativo, el medio condicionado de las células COS-1 transfectadas con el vector vacío pcDNA3.1 durante la primera ronda de cribado. Haga clic aquí para descargar este archivo.
Complementarios Figura 2: Número de células vivas. A. La segunda ronda de selección con sub-piscinas de 0073 piscina. B. La tercera ronda de cribado con clones aislados. CN: el medio condicionado de células COS-1 transfectadas con el vector vacío, pcDNA3.1. Haga clic aquí para descargar este archivo.
Figura suplementaria 3: Actividad trófica de los tres marcos de lectura abiertos del clon aislado 0073-09-37. Análisis estadístico utilizando la prueba de Dunnett. Haga clic aquí para descargar este archivo.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
| un | 1 | 1 | 1 | 1 | 2 | Nc | 2 | 2 | 2 | 3 | 3 | Nc |
| B | Nc | 3 | 3 | 4 | 4 | 4 | Nc | 4 | 5 | 5 | 5 | 5 |
| C | 6 | Nc | 6 | 6 | 6 | 7 | 7 | Nc | 7 | 7 | 8 | 8 |
| D | 8 | 8 | Nc | 9 | 9 | 9 | 9 | 10 | Nc | 10 | 10 | 10 |
| E | 11 | 11 | 11 | Nc | 11 | 12 | 12 | 12 | 12 | Nc | 13 | 13 |
| F | 13 | 13 | 14 | 14 | Nc | 14 | 14 | 15 | 15 | 15 | Nc | 15 |
| G | 16 | 16 | 16 | 16 | 17 | Nc | 17 | 17 | 17 | 18 | 18 | Nc |
| h | 18 | 18 | 19 | 19 | 19 | 19 | Nc | 20 | 20 | 20 | Nc | 20 |
| Controles NC negativos | ||||||||||||
| 1-20 posiciones de las piscinas probadas en cuadruplicado | ||||||||||||
Tabla 1: Plan de la placa de 96 pozos para cribado de alto contenido
Discusión
Entre los muchos parámetros que podrían limitar la producción de un cultivo enriquecido con cono a partir de embriones de pollo, el primer paso crítico es identificar con precisión la etapa de desarrollo de los embriones en los huevos eclosionados. Se ha observado que el cultivo de células de las retinas de los embriones en ED8 (34ª etapa) produce sólo el 35% de los fotorreceptores, mientras que el 65% restante están hechos de otras neuronas4. Cualquiera que sea la logística aplicada para obtener los huevos eclosionados, es necesario afinar la temperatura y el tiempo de incubación, y examinar cuidadosamente los embriones en comparación con las imágenes de referencia de todas las etapas del desarrollo42,45.
Originalmente, el sistema de cultivo enriquecido con cono se desarrolló utilizando la cepa White Leghorn4. El color blanco de los huevos de esa cepa no es particularmente apreciado en Francia, por lo que utilizamos una cepa de pollo que producen huevos marrones. Utilizamos la cepa I 657, que se hace cruzando I 66 gallos con gallinas JA575. Pudimos reproducir las características de las culturas originales. Esto demuestra que el fondo genético del pollo no es crítico para obtener cultivos enriquecidos con cono.
No hemos probado el efecto de la eliminación individual de los suplementos en el medio de cultivo, pero hemos observado que la insulina juega un papel crítico de acuerdo con el efecto de la insulina sobre la supervivencia de los conos en el ratón rd1, un modelo de RP autosómico recesivo46. La triyodotironina (T3) también puede participar en la diferenciación de las células precursoras retinianas del embrión de pollo en conos según el papel del receptor de la hormona tiroidea en el destino de las células de la retina durante el desarrollo40. En consecuencia, el sistema de cultivo enriquecido con cono no puede ser utilizado para identificar la insulina por clonación de expresión46.
El sistema de cultivo enriquecido con cono se basa en el cultivo de neuronas primarias y es mucho más apropiado tha métodos que se basan en el uso de células inmortalizadas como la línea celular 661W24,25,26.
El método aquí descrito puede ser modificado mediante la realización de una electroporación previa con ADN plásmido20. Antes de preparar la suspensión celular de la retina, toda la retina se coloca en la cámara de un electroporador a medida con 120 μL de 0,5 μg/μL de ADN plásmido en 10 mM Tris-HCl pH 8.0, 1 mM EDTA. Se aplican cinco impulsos de 15 V durante 50 ms cada uno separados por un intervalo de 950 ms22. Los intentos de suministrar ARN interferente (ARNi) utilizando los retrovirus de empalme aviar competente en replicación (RCAS) en cultivos enriquecidos con conos fueron infructuosos47. Esto se debe sin duda al hecho de que en ausencia de suero y en cultivos de baja densidad, las células precursoras de la retina no son replicativas, un requisito para la propagación de retrovirus.
Desarrollamos el sistema de cultivo enriquecido con cono para identificar los factores tróficos que promueven la supervivencia del cono utilizando la clonación de expresión19. Para hacerlo factible, realizamos un primer paso de cribado de alto contenido utilizando medio acondicionado de piscinas de 100 clones. Incluso si los cDNAs de la biblioteca se expresan bajo el control de un promotor fuerte de CMV después de la transfección de las células COS-1, no ofrece una garantía de que todas las proteínas codificadas por cDNAs individuales alcancen una concentración suficiente para ser puntuadas positivamente por el ensayo de viabilidad. Esta es una limitación importante. En ese sentido, cualquier cribado no es realmente exhaustivo. Además, incluso si las proteínas membranosas no se eliminaron del medio acondicionado, la configuración del ensayo es desfavorable a la identificación de factores no difusivos. Una alternativa sería examinar clones individuales después de haber obtenido la secuencia de los cDNAs para evitar duplicaciones en el ensayo de muchas veces la misma proteína candidata. Esto fue iniciada secuenciando las bibliotecas retinianas del cDNA que utilizamos43. Si bien es racional, este enfoque también tiene sus limitaciones. El análisis bioinformático de las secuencias de ADNc impondrá irresistiblemente, además de la reducción de la redundancia, la priorización del cribado de ciertos clones basados en el conocimiento. Esto no será perjudicial si finalmente, toda la biblioteca se examinara, incluso si el tiempo necesario para hacerlo se alarga significativamente. Pero invariablemente, la identidad de la secuencia influirá en nuestra forma de ver los resultados. Esto no será neutral ya que la interpretación de la secuencia estará naturalmente en competencia con los datos experimentales.
La identificación de EdCVF también muestra que la detección de alto contenido conlleva limitaciones técnicas. A partir de la primera ronda de cribado, se identificaron dos grupos con alta actividad(Figura Suplementaria 1). El grupo 0073 condujo a la identificación exitosa de EdCVF, mientras que el grupo 0080 no llevó a cabo tal hallazgo. No hemos resuelto el problema que podría resultar de la pérdida del clon activo durante la preparación de sub-pools. Alternativamente, no se excluye, aunque no sea estadísticamente favorable, que entre los cDNAs del pool 0080, dos proteínas actuaban sinérgicamente y su actividad no podía ser observada como clones individuales.
La identificación de moléculas que protegen los conos mediante el cribado de moléculas pequeñas es una aplicación futura del sistema de cultivo enriquecido con cono. Tales moléculas serán inestimables para el tratamiento de las patologías retinianas para las cuales la terapia génica no es el acercamiento más apropiado como degeneración macular relativa a la edad.
Divulgaciones
TL, J-AS y VF poseen una patente sobre el uso de EdCVF para tratar degeneraciones retinianas [WO2009071659 (A1). 12 de junio de 2007].
Agradecimientos
Los autores agradecen a Jacques Bellalou, Lorelei Fournier, Emmanuelle Clérin, Frédéric Blond y a la empresa agricole à responsabilité limitée (EARL Morizeau Dangers, Francia) su inestimable ayuda. Este trabajo fue apoyado por inserm, universidad de la Sorbona, la Agence Nationale pour la Recherche (ANR, Labex Lifesenses), Fundación que lucha contra la ceguera (EE.UU.) y la IHU FOReSIGHT [ANR-18-IAHU-0001] con el apoyo de fondos estatales franceses gestionados por la ANR dentro del programa Investissements d'Avenir.
Materiales
| Name | Company | Catalog Number | Comments |
| 96 black plates (Clear Button with lid Tissue culture treated) | Corning | 3603 | |
| Calcein AM | Thermo-Fisher scientific | C1430 | |
| CCD Camera | Photometrics | CoolSnap FX HQ | |
| CDPC (Cytidine 5′-diphosphocholine sodium salt dihydrate) | Sigma-Aldrich | C0256 | |
| CO2 Independant | Thermo-Fisher scientific | 18045-054 | |
| Curved forceps | Dutscher | 005093 | |
| DMEM Media | Thermo-Fisher scientific | 41966-029 | |
| DNAse | Sigma-Aldrich | D4263 | |
| Eggs incubator | FarmLine | M08 01 3100 | |
| Ethidium Homodimer | Thermo-Fisher scientific | E1169 | |
| Fœtal bovine serum | Thermo-Fisher scientific | 10270-098 | |
| Gentamycin | Thermo-Fisher scientific | 15710-049 | |
| Hydrocortisone | Sigma-Aldrich | H0880 | |
| ITS (insulin Transferine selenium) | Sigma-Aldrich | I1884 | |
| large straight pliers | Dutscher | 005074 | |
| Linoleic acid | Sigma-Aldrich | L8384 | |
| M199 medium | Thermo-Fisher scientific | 31150-022 | |
| Metamorph software | Metamorph | ||
| Microscope | NIKON | Eclipse TE2000 | |
| Motorized stage | Martzauzer | Mutlicontrol 2000 | |
| Optical filter switch | Shutter Instrument company | Lambda 10-2 | |
| PBS 1X | Thermo-Fisher scientific | 14190-086 | |
| Poly-L-lysine | Sigma-Aldrich | P6282 | |
| Progesterone | Sigma-Aldrich | P7556 | |
| Pursept A express | Fisher scientific | 11814110 | |
| Putriscine | Sigma-Aldrich | P5780 | |
| Sodium pyruvate | Sigma-Aldrich | S8636 | |
| straight forceps | Dutscher | 005092 | |
| Taurine | Sigma-Aldrich | T8691 | |
| Triiodothyronine | Sigma-Aldrich | T6397 | |
| Trypan blue | Thermo-Fisher scientific | 15250-061 | |
| Trypsine 0.25 % | Thermo-Fisher scientific | 25200-056 |
Referencias
- Brownstein, C. D. . The Implicit Mind. , (2019).
- Heesy, C. P., Hall, M. I. The nocturnal bottleneck and the evolution of mammalian vision. Brain, Behavior and Evolution. 75 (3), 195-203 (2010).
- Borges, R., et al. Adaptive genomic evolution of opsins reveals that early mammals flourished in nocturnal environments. BMC Genomics. 19 (1), 121 (2018).
- Adler, R., Hatlee, M. Plasticity and differentiation of embryonic retinal cells after terminal mitosis. Science. 243 (4889), 391-393 (1989).
- Fintz, A. C., et al. Partial characterization of retina-derived cone neuroprotection in two culture models of photoreceptor degeneration. Investigative Ophthalmology & Visual Science. 44 (2), 818-825 (2003).
- Roska, B., Sahel, J. A. Restoring vision. Nature. 557 (7705), 359-367 (2018).
- Duncan, J. L., et al. Inherited Retinal Degenerations: Current Landscape and Knowledge Gaps. Translational Vision Science & Technology. 7 (4), 6 (2018).
- Russell, S., et al. Efficacy and safety of voretigene neparvovec (AAV2-hRPE65v2) in patients with RPE65-mediated inherited retinal dystrophy: a randomised, controlled, open-label, phase 3 trial. Lancet. 390 (10097), 849-860 (2017).
- RetNet. . Retinal Information Network. , (2020).
- Doonan, F., Donovan, M., Cotter, T. G. Activation of multiple pathways during photoreceptor apoptosis in the rd mouse. Investigative Ophthalmology & Visual Science. 46 (10), 3530-3538 (2005).
- Comitato, A., et al. Dominant and recessive mutations in rhodopsin activate different cell death pathways. Human Molecular Genetics. 25 (13), 2801-2812 (2016).
- Cronin, T., Leveillard, T., Sahel, J. A. Retinal degenerations: from cell signaling to cell therapy; pre-clinical and clinical issues. Current Gene Therapy. 7 (2), 121-129 (2007).
- Baumgartner, W. A., Baumgartner, A. M. Accounting for disagreements on average cone loss rates in retinitis pigmentosa with a new kinetic model: Its relevance for clinical trials. Medical Hypotheses. 89, 107-114 (2016).
- Leveillard, T., Sahel, J. A. Rod-derived cone viability factor for treating blinding diseases: from clinic to redox signaling. Science Translational Medicine. 2 (26), (2010).
- Leveillard, T., Sahel, J. A. Metabolic and redox signaling in the retina. Cellular and Molecular Life Sciences. 74 (20), 3649-3665 (2017).
- Clerin, E., Marussig, M., Sahel, J. A., Leveillard, T. Metabolic and Redox Signaling of the Nucleoredoxin-Like-1 Gene for the Treatment of Genetic Retinal Diseases. International Journal of Molecular Sciences. 21 (5), (2020).
- Fridlich, R., et al. The Thioredoxin-like Protein Rod-derived Cone Viability Factor (RdCVFL) Interacts with TAU and Inhibits Its Phosphorylation in the Retina. Molecular & Cellular Proteomics. 8 (6), 1206-1218 (2009).
- Mei, X., et al. The Thioredoxin Encoded by the Rod-Derived Cone Viability Factor Gene Protects Cone Photoreceptors Against Oxidative Stress. Antioxidants, Redox Signaling. 24 (16), 909-923 (2016).
- Leveillard, T., et al. Identification and characterization of rod-derived cone viability factor. Nature Genetics. 36 (7), 755-759 (2004).
- Vergara, M. N., Gutierrez, C., Canto-Soler, M. V. Efficient Gene Transfer in Chick Retinas for Primary Cell Culture Studies: An Ex-ovo Electroporation Approach. Journal of Visualized Experiments. (105), e52002 (2015).
- Fridlich, R., et al. The thioredoxin-like protein rod-derived cone viability factor (RdCVFL) interacts with TAU and inhibits its phosphorylation in the retina. Molecular & Cellular Proteomics. 8 (6), 1206-1218 (2009).
- Ait-Ali, N., et al. Rod-derived cone viability factor promotes cone survival by stimulating aerobic glycolysis. Cell. 161 (4), 817-832 (2015).
- Mei, X., et al. The Thioredoxin Encoded by the Rod-Derived Cone Viability Factor Gene Protects Cone Photoreceptors Against Oxidative Stress. Antioxidants & Redox Signaling. 24 (16), 909-923 (2016).
- Tan, E., et al. Expression of cone-photoreceptor-specific antigens in a cell line derived from retinal tumors in transgenic mice. Investigative Ophthalmology & Visual Science. 45 (3), 764-768 (2004).
- Wang, X. W., Tan, B. Z., Sun, M., Ho, B., Ding, J. L. Thioredoxin-like 6 protects retinal cell line from photooxidative damage by upregulating NF-kappaB activity. Free Radical Biology and Medicine. 45 (3), 336-344 (2008).
- Perron, N. R., Beeson, C., Rohrer, B. Early alterations in mitochondrial reserve capacity; a means to predict subsequent photoreceptor cell death. Journal of Bioenergetics and Biomembranes. 45 (1-2), 101-109 (2013).
- Krishnamoorthy, R. R., Clark, A. F., Daudt, D., Vishwanatha, J. K., Yorio, T. A forensic path to RGC-5 cell line identification: lessons learned. Investigative Ophthalmology & Visual Science. 54 (8), 5712-5719 (2013).
- Al-Ubaidi, M. R. RGC-5: are they really 661W? The saga continues. Experimental Eye Research. 119, 115 (2014).
- Clerin, E., et al. Vibratome sectioning mouse retina to prepare photoreceptor cultures. Journal of Visualized Experiments. (94), (2014).
- Hendrickson, A., Hicks, D. Distribution and density of medium- and short-wavelength selective cones in the domestic pig retina. Experimental Eye Research. 74 (4), 435-444 (2002).
- Bobu, C., Craft, C. M., Masson-Pevet, M., Hicks, D. Photoreceptor organization and rhythmic phagocytosis in the nile rat Arvicanthis ansorgei: a novel diurnal rodent model for the study of cone pathophysiology. Investigative Ophthalmology & Visual Science. 47 (7), 3109-3118 (2006).
- Saidi, T., Mbarek, S., Chaouacha-Chekir, R. B., Hicks, D. Diurnal rodents as animal models of human central vision: characterisation of the retina of the sand rat Psammomys obsesus. Graefes Arch Clin Exp Ophthalmol. 249 (7), 1029-1037 (2011).
- Traverso, V., Kinkl, N., Grimm, L., Sahel, J., Hicks, D. Basic fibroblast and epidermal growth factors stimulate survival in adult porcine photoreceptor cell cultures. Investigative Ophthalmology & Visual Science. 44 (10), 4550-4558 (2003).
- Balse, E., et al. Purification of mammalian cone photoreceptors by lectin panning and the enhancement of their survival in glia-conditioned medium. Investigative Ophthalmology & Visual Science. 46 (1), 367-374 (2005).
- Gagliardi, G., et al. Characterization and Transplantation of CD73-Positive Photoreceptors Isolated from Human iPSC-Derived Retinal Organoids. Stem Cell Reports. 11 (3), 665-680 (2018).
- Gagliardi, G., Ben M'Barek, K., Goureau, O. Photoreceptor cell replacement in macular degeneration and retinitis pigmentosa: A pluripotent stem cell-based approach. Progress in Retinal and Eye Research. 71, 1-25 (2019).
- Mears, A. J., et al. Nrl is required for rod photoreceptor development. Nature Genetics. 29 (4), 447-452 (2001).
- Swaroop, A., Kim, D., Forrest, D. Transcriptional regulation of photoreceptor development and homeostasis in the mammalian retina. Nature Reviews Neuroscience. 11 (8), 563-576 (2010).
- Kallman, A., et al. Investigating cone photoreceptor development using patient-derived NRL null retinal organoids. Nature Communications. 3 (1), 82 (2020).
- Eldred, K. C., et al. Thyroid hormone signaling specifies cone subtypes in human retinal organoids. Science. 362 (6411), (2018).
- Kole, C., et al. Otx2-Genetically Modified Retinal Pigment Epithelial Cells Rescue Photoreceptors after Transplantation. Molecular Therapy. 26 (1), 219-237 (2018).
- Hamburger, V., Hamilton, H. L. A series of normal stages in the development of the chick embryo. Journal of Morphology. 88 (1), 49-92 (1951).
- Kole, C., et al. Identification of an Alternative Splicing Product of the Otx2 Gene Expressed in the Neural Retina and Retinal Pigmented Epithelial Cells. PLoS One. 11 (3), 0150758 (2016).
- Shannon, M., Hamilton, A. T., Gordon, L., Branscomb, E., Stubbs, L. Differential expansion of zinc-finger transcription factor loci in homologous human and mouse gene clusters. Genome Research. 13 (6), 1097-1110 (2003).
- Stern, C. D., Holland, P. W. . Essential developmental biology: a practical approach. , (1993).
- Punzo, C., Kornacker, K., Cepko, C. L. Stimulation of the insulin/mTOR pathway delays cone death in a mouse model of retinitis pigmentosa. Nature Neuroscience. 12 (1), 44-52 (2009).
- Harpavat, S., Cepko, C. L. RCAS-RNAi: a loss-of-function method for the developing chick retina. BMC Developmental Biology. 6, 2 (2006).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados