
Method Article
鶏胚のレチナから、ロッドからコーン細胞相互作用を研究するコーンエンリッチ培養
要約
鶏胚のレチナからコーン感光体の一次培養物を得るための方法と、高含有スクリーニングの使用について述べた。
要約
人間の昼間の視力は、レチナ、窩の中心にあるコーン感光体の機能に依存しています。最も一般的な形態の遺伝性網膜変性、網膜色素変性症に罹患している患者は、ロッド感光体の突然変異駆動喪失のために夜間視力を失い、その後、機能の進行性喪失および失明につながるコーンの死を引き起こした。遺伝学者は、この疾患を引き起こす突然変異を有する多くの遺伝子を同定したが、最初に同定された突然変異は、二次コーン変性のメカニズムと、ロッドで排他的に発現する視覚色素をコードするロドプシン遺伝子の支配的な突然変異がコーン変性を引き起こす方法を疑問視した。
この疾患の遺伝的モデルにおける移植の結果は、棒と錐体の間の細胞相互作用の概念と、網膜色素変性症のすべての遺伝的形態における錐体の非細胞自律変性の概念につながった。
コーンはヒトのすべての感光体の5%、マウスの3%しか含まないので、これらの種では研究は難しいが、コーンは鳥類の棒を上回る。我々は、その発達の段階29で鶏胚のレティナから培養レチン前駆体に96ウェルプレートを適応させました。これらの一次培養において、円錐はインビトロ分化後の細胞の80%を表す。細胞は、血清がない場合に1週間の期間にわたって退化する。ここでは、メソッドとその標準化について説明します。
このコーン濃縮培養系は、ラット網膜色素上皮正規化cDNAライブラリーの高含有スクリーニングにより上皮由来コーン生存率因子(EdCVF)を同定するために使用された。組換えEdCVFは、コーンの変性を防ぎます。
概要
脊椎動物種のレチナは二重で、薄暗い光ビジョン用のロッド感光体と、日光、色、視力ビジョンのためのコーン感光体を備えています。霊長類の視力は、錐体で濃縮された窩と呼ばれるレチナの中心にある領域に依存するが、全体的に、コーンはすべての感光体の5%しか表さない。その結果、霊長類のレチナにおけるコーンの分析、特にコーンの培養は技術的に困難である。他のすべての哺乳類種はfoveaを持たないし、肛門の研究で最も一般的に使用されるげっ歯類のコーンの割合は低い。これは、これらのよく見る鳥の種のレティナを支配する鳥類種には当てはまりません。進化の際に哺乳類が最初に出現したときに生態系を支配してきた恐竜は、鳥1の系統的起源にある。恐竜と初期哺乳類の間のこのような競争の結果として、哺乳類は主に棒によって支配されるレチナを有する夜行性である。進化の後になって初めて、霊長類が属するいくつかの哺乳類種の日次ビジョンが進化的な利点となった。それにもかかわらず、祖先期は哺乳類ビジョン2、3の進化における夜行性ボトルネックのアタビズムとして残っている。
副腎細胞分化を研究している間、アドラーとハトリーは、感光体が胚の日(ED)6またはステージ294で鶏由来の培養において、約70%のレチナル分化細胞を表していることを示した。鶏のレチナにおける円錐の有病率のために、ED6鶏胚からのレチナル細胞の培養は円錐を豊富に含む培養物として開発された5。
人間にとってコーンを介した視力の重要性は、トラウマである。コーン機能を変化させる遺伝性疾患や老化性疾患の影響を受ける人は、大きな障害を持っています。これは、これらの盲検疾患の治療法を見つけることを目的として遺伝性の遺伝性退化(IRD)に関する研究の非常に大規模な体を促進しました6,7.最初の成功は、IRDレーバー先天性アマウロシス(LCA)の重篤な形態の治療のために組換えアデノ関連ベクター(AAV)を用いて得られた、遺伝子治療8の概念実証である。変異がIRDを引き起こす遺伝子の同定は、遺伝子治療を使用してこれらの疾患を治癒する可能性を開きます。それにもかかわらず、これらの疾患は、200以上の異なる遺伝子9の突然変異から生じる。IRDの常染色体劣性形態の場合でも、病的遺伝子の正常なコピーの再導入が視覚機能を回復することができるとき、個々の開発の経済的コストは、あまり一般的なものではなく、遺伝的起源が不明のままであるものに対して最も一般的なものを支持する。この事実は、研究者がより一般的な治療法について考えるきっかけになった。アポトーシス細胞死は、一般的な経路として現れ、常染色体優勢形態10,11を含む感光体の変性によって進行するこれらの疾患の治療対象である。しかし、そのようなアプローチの成功は欠けています。IRDの最も一般的な形態については、網膜色素変性症(RP)は、共通経路が最終的にコーン12、13の変性に続く機能の二次的損失である。コーン機能の喪失を防ぐことは、原因変異14とは無関係に窩の中心視力を維持する。
RPの初期段階では、ロッドの喪失は、棒とコーン15の間の代謝および酸化還元シグナル伝達を遮断するヌクレドキシン様1(NXNL1)遺伝子によってコードされるロッド由来コーン生存率因子(RdCVF)の発現の減少を引き起こす。NXNL1遺伝子の2つの産物をコードする組換えAAVの投与により、栄養因子RdCVFとチオレドキシン酵素RdCVFLは、RP16の全遺伝形態におけるコーン視力喪失を理論的に防止できる。我々は、NXNL1遺伝子産物RdCVFLがチキンコーン濃縮培養17で発現され、保護的な役割を果たす場所18であることを示した。RdCVFおよびNXNL1遺伝子は、読出し19としてコーン濃縮培養物からの細胞の生存を用いたretinal cDNAライブラリーの高含有スクリーニングによって同定された。各クローンに対して8つの並列テストを使用して、ライブラリのクローン210,000個相当をスクリーニングしました。これは、生物学的材料、鶏の胚のレティナへの容易なアクセスを必要とする非常に多くのテストを表しています。産卵鶏や肉を産む鶏の農業産業向けに広く生産されているため、週単位で胚化鶏卵を得るのは比較的簡単であることがわかりました。コーンエンリッチ培養の慎重な標準化の後、システムはコーンの生存率を維持する能力のために何千もの分子をテストする簡単で堅牢で再現可能な方法を提供します。これらの細胞はまた、シグナル伝達の研究と生化学的分析21、22、23に利益をもたらす遺伝子操作20に適している。
研究家は、コーン細胞株661W24,25,26の使用として代替方法を開発した。それにもかかわらず、この細胞株のアイデンティティは議論の余地がある27、28.661W細胞を、ヒト光受容体間レチノール結合タンパク質プロモーターの制御下でSV40大T抗原を発現するトランスジェニックマウスラインのレチナル腫瘍からクローン化した。SV40大T抗原は細胞の形質転換と不死化を媒介する。その結果、661W細胞を用いて同定されたシグナル伝達経路は、その中のコーンとは多くの点で明確である形質転換および不死化細胞株の文脈に報告されなければならない。その点、コーンエンリッチ培養系は、一次ニューロン、より生理学的に関連するコーンで構成される。
マウスのレチナのビブラート断面を用いて感光体の純粋な培養を得ることは可能であるが、げっ歯類の外性のレチナにおけるコーンの非常に低い割合は、このアプローチをコーン濃縮培養29を作り出すためには不適当である。豚のretinaは、foveaを含んでいませんが、コーン30で非常に豊かにされたエリア中央と呼ばれる領域を持っています。アルビカンティス・アンソルゲイとプサンモミース・オブセッサス31、32のように、レナ・ディナル・げっ歯類のコーンの高い割合は、可能な解決策を提供するが、そのようなエキゾチックな種の繁殖を必要とする。地元の食肉処理場から採取された成虫の豚の目は、感光体生存33を研究するために使用されてきたロッドとコーンの混合培養物を作り出すために使用することができる。エレガントな解決策は、ピナツナグルチニン(PNA)レクチンをパンニングを使用してブタのレチナからコーンを予精製し、コーン34に選択的に結合する。しかし、このメソッドは複雑であるため、大規模に実装することは困難です。
ヒト誘導多能性幹細胞(iPS)は、腎移植に使用できるが、コーンエンリッチ培養35,36にも適応できるコーン感光体細胞集団を得るための最も有望なアプローチを提供する。転写因子NRLはロッド感光体37に必要であるため、Nrl-/-マウスは短波コーン(Sコーン)によって支配されるレチナを有する。不活性化は、iPS38,39からのヒトの分化によってS-コーン濃縮製剤を生成するために使用することができる。もう一つの可能なアプローチは、甲状腺ホルモンシグナル伝達40を使用して円錐分化を促進することです。ヒトiPSからコーンを豊富に含む培養を作製する新しい方法が出現しているが、鶏の胚は現在実証済みの方法19を提供する。
コーンリッチ培養は、発現クローニング19によるRdCVFの同定に役立った。このシステムは、RdCVFが好気性解糖分解22によってグルコース取り込みおよびその代謝を刺激することを実証するためにもうまく使用された。さらに、コーンエンリッチ培養は 、NXNL1 遺伝子23の第2の産物であるRdCVFLの保護役割を検証するために使用された。さらに最近では、OTX241を導入した網膜色素上皮細胞によって分泌される保護分子の存在を実証するためにこのシステムが用いられた。
プロトコル
この議定書は、ピエール大学とマリー・キュリー大学の動物実験倫理委員会とフランス研究省(許可番号:APAFIS#1028 2015070211275177)によって承認されました。動物実験は次の認可の下で行われました: "セルティフィカト・ド・オートリゼーション・ド・デ・エクスペリメンター・シュル・レ・アニモー・ヴェルテブレA-75-1863。パリ警察のプレフェクチャー(2011年11月9日~2016年11月8日)」。
1. 受精卵のインキュベーション
- 産業孵化場で、自然に得られる毎週の受精卵(株I 657、赤いラベル)を収集します。
- 受精卵は鶏によって「産まれた」後、実験室で17°C(生物学的ゼロ)で維持します。
- 各培養物に対して、7個の受精卵を20°Cで24時間、37°Cで136時間培養し、加湿室で卵の傾斜(片側から反対側へ2時間の進行動、4時間周期)を間欠的に反転させます。
2. 鶏胚の回収
- 7個の卵の表面を消毒剤で洗います(例えば、Pursept A急行)。
- 卵殻を壊すために、大きなまっすぐなペンチでシェルの上部に穴を開けます。その後、殻を切って卵から帽子を柔らかいゆで卵として取り除きます。
- 卵殻から各胚を緩やかに引き出し、37°Cに加熱した無菌リン酸緩衝液生理塩水(PBS)を含むペトリ皿に移す。 胚(絨毛膜または絨毛管膜)を取り囲むエンベロープをそっと取り除きます。
- ハンブルガーとハミルトン42と視覚的に比較して、各胚の発達段階を検証する。
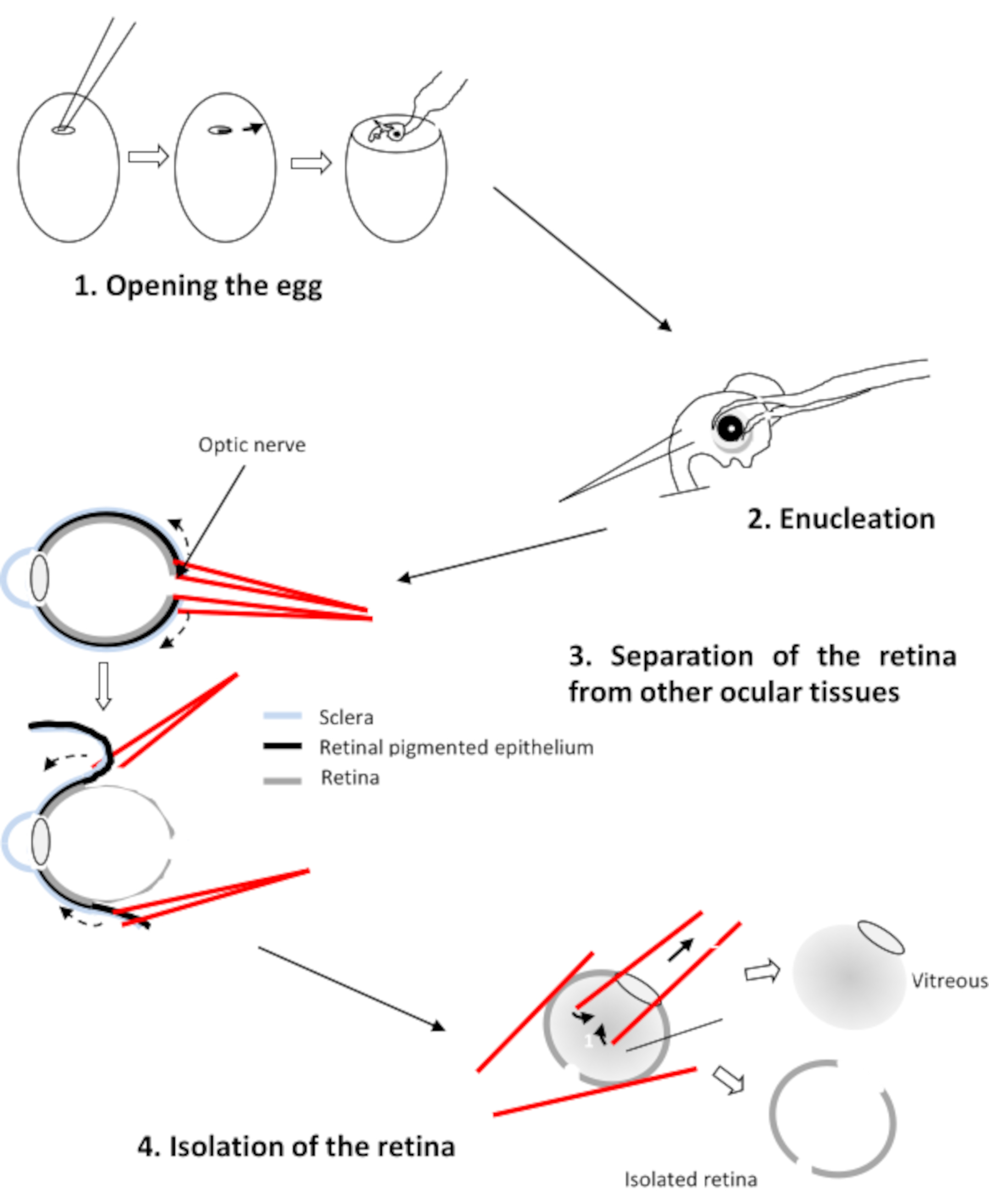
- 開発の第29段階 で2つの胚を選択します(図1)。翼は肘で曲がる。襟が目に見えて目立ちます。法案は28番目 の段階よりも顕著です。
- これらの選択した胚の目を核とし、CO2-独立培地(生命技術)でそれらを移す。
注意: 胚がステージ 29 にあり、ステージ 28 または 30 (図1)にないことを非常に重要です。2個しか使わなくても、少なくとも7個の卵をインキュベートする必要があるのはそのためです。絨毛は非常に薄く、区別するのが難しいが、胚と非常に近いので、胚に触れることなく取り除かねばなっている。
3. レティナの解剖
- 切断し、湾曲した鉗子で選択した胚を核とします。
- 4つの目をCO2-独立培地に移します。この媒体は0.9 mM CaCl2 および0.65 mM MgCl2を含んでいる。
- 角膜を下にして眼を下に、実験者に向かって視神経を置く。2つのストレート鉗子を使用して視神経に穴を開けます。
- 網膜と色素上皮の間に各鉗子の分岐を挿入する(図2)。各鉗子を引っ張り、眼を回転させて網膜から上皮を取り外します。角膜を取り除き、レンズとガラス膜を取り除きます。
- pH 7.2でリンガーの媒体を含むペトリ皿に4つのレチナを移します。
注意:網膜だけが残っていることを確認し、色素性および残存網膜色素上皮の痕跡を取り除く。
4. 細胞の細胞懸濁液の準備
- 2つのまっすぐなペンチを使用して非常に小さな部分に4つのレチナをカットします。
- リンガーの媒体で2回、レチンの部分を洗います。
- リンガーの媒体で2回目の洗浄を行った後、レティナの破片をチューブの底に落とし、リンガーの媒体を取り除きます。トリプシン(0.25%w/v)の溶液で37°Cで20分間、レチナルピースを処理します。
- 連続吸引により10分後に溶液を分散させる。パスツールピペットを使用して排出し、レチナルピースの解離を確認します。10%不活性化胎児子牛血清を添加した培養培地を添加して反応を停止する。
- DNase Iの0.05mgで細胞懸濁液をインキュベートし、DNaseを添加した直後にパスツールピペットを使用して連続吸引および排出することによって細胞クラスターとDNAを解震させる。
- レチナル細胞懸濁液を化学的定義培養培地(CDCM)で2回洗浄する:100 μg/mLリノール酸/BSAを添加したダルベッコの修飾イーグル培地およびM199培地の等量 0.86 μMインスリン、0.07 μMトランスフェリン、2.0 μMプロゲステロン、0.28 μMプロスタグランジン、0.29 μM Na2SeO 3、182 μM プトレシン、3 mMタウリン、 4.7 μMシチジン5′ジホスホコリン、2.7 μMシチジン5'ジホスホエタノールアミン、0.55 μMヒドロコルチゾン、0.03 μMトリヨードチロニン、1 mMナトリウムピルビン酸、20μMゲンタマイシン。
5. レティナル細胞の播種
- 37°Cで2時間透明な底を持つ2つの黒い96ウェル培養プレートをポリL-リジンで32.25 μg/cm2で扱います。
- これらのプレートをM199培地で2回リンスします。CDCMの1 mLで細胞ペレットを再懸濁する。
- 細胞懸濁液トリパンブルーの10μLのアリコートに加えて、生きている細胞を染色します。細胞懸濁試料を血球計(マラッセの細胞計数室)に加える。
- 顕微鏡下で、血球計の4列(すなわち40四角)の染色された細胞を数え、1行の平均細胞数を計算する。懸濁液の細胞の濃度を計算するには、次の方法を適用します: 細胞/mLの懸濁液の数 = 行内の平均セル数 (10 四方) x 10 トリパンブルー x 1,000 での血球減少の総平方数を 1,000 (セル/mL として結果を表す)
- CDCMを使用して、2つのめっき密度(1 x 105細胞/cm2および2 x 10 5細胞/cm2)に対応する2つの濃度(5.6 x 104細胞/mLおよび1.12 x 105細胞/mL)に細胞懸濁液を持って来なさい。
- 2つの細胞懸濁液の種子50μLを、2つの前処理された黒96ウェル培養プレートに入った。プレートの右側から左側にマルチチャンネルピペットを使用してプレート内の細胞を分配し、各カラム間で均質化し、細胞の分布が均質になるようにします。
- あらかじめ定義されたパターンを用いてスクリーニングされる分子のライブラリの50μL(例えば、cDNAライブラリからのコンディションされたメディア、下記参照)を加える(表1)。
- 培地交換なしで、37°CのCO2 の下で7日間プレートをインキュベートします。
6. 生存可能な細胞を数える
- プレートの各ウェルに、カルセインAMの2.7 μMとエチジウムホモモマーの0.3 mMを加えます。
注:カルセインは、エチジウムホモメータとして大きな分子に不浸透性である細胞を貫通します。生細胞の細胞質では、カルセインは内因性エステラーゼによって加水分解され、485nmで励起されると520nmで発光することにより蛍光化する。エチジウムホモジマーは死んだ細胞上のDNAに結合する。エチジウムDNA-ホモジマー結合は、520nmで励起した後、635 nmで赤色蛍光を放出する。 - 光が当たらなくて室温でプレートを1時間インキュベートします。
- 485と520 nmの2つの励起フィルターを備えた水銀ランプを備えた反転顕微鏡、520および635 nmの2つの発光フィルター、目的(x10)、プロセッサと電荷結合デバイスカメラ(CCD)によって制御される電動ステージで構成された自動プレートリーダーで蛍光を読み取ります。このカウントプラットフォームは、メタモルフソフトウェア19によって制御されます。96ウェルプレートの各ウェルで生きている細胞と死細胞の蛍光画像を同時に取得し、ウェルA1からウェルA12に向かい、次いでB12からB1に向かって、C12に向かってC12などを得ることを可能にする(表I)。
- 負のコントロールの 18 ウェルを使用して、単一セルの平均領域 A を計算します (表 I)。
- 各プレートウェル内のセルをカウントし、次の経験的な式A x 29 / 20.7を適用して、セルダブレット(2つのセルのグループ化)がカウントされるのを防ぎます。分子による円錐の保護効果を、負の対照の18ウェル内の分子対平均細胞数を試験した4つのウェルにおける平均細胞数との比率としてスコアを付けます( 表1 および 補足図1を参照)。
- 1 x 105 細胞/cm2 で播種したプレートの結果と2 x 105 細胞/cm2 で播種したものを組み合わせて、スクリーニングされた各分子による潜在的保護(生存率)を評価します。
結果
ここでは、コーン濃縮培養システムを使用して、新規のコーン保護タンパク質を同定する方法を説明します。このプロトコルを使用して、8週齢のLong-Evansラット43の400眼から脈絡膜および網膜色素上皮で作られた正規化されたcDNAライブラリをスクリーニングしました。
このライブラリには、6.0 x 106 個の独立したコロニー形成ユニット (CFUs) が含まれ、平均クローン挿入サイズは 2.1 キロベース (kb) で、組み換えクローンの 99% を超えています。そのライブラリーから100クローンのプールを一過性にトランスフェクトし(0.1μgのプラスミドDNA)をCOS-1細胞に、COS-1のコンディション培地(CM)を、血清なしでDMEMで48時間培養した後に収穫した。膜またはエキソソームは、超遠心分離によって除去されなかった。各CMの50マイクロリットルは、2つの96ウェルプレートの4つのウェルに加えられました:1つは2 x 105 細胞/cm2で播種し、もう1つは4 x 105 細胞/cm2で播種した。ライブラリを構築するために使用される空のベクターpcDNA3.1にトランスフェクトされたCOS-1細胞からのCMを、陰性制御として用いた(表1)。211,200個のクローンに対応する100クローンの合計2,112セットを4つの培養ウェルおよびコーン濃縮培養の2つの播種条件について評価した。2つの条件は、2つのわずかに異なる接種密度に対応し、合計1,689,600の培養井戸に対して、保護活性をより正確に評価することを可能にする。
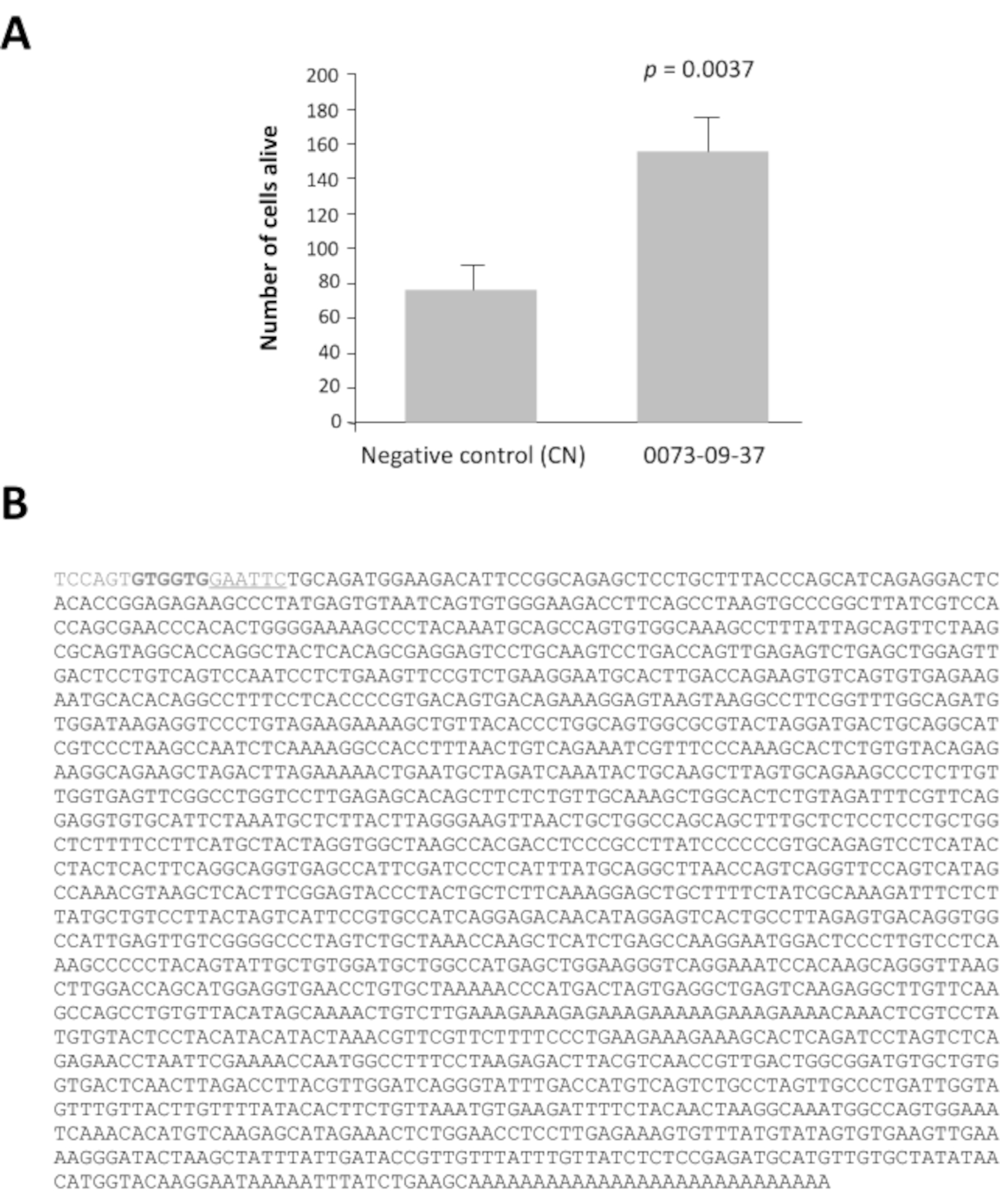
2より大きい比率を有するクローンの42プールのうち、プール0080および0073は、負の対照よりも培養の7日後に生存率比が16および14倍高い、pcDNA3.1(補足図1)。この分析は、関心のあるプールを特定するために不可欠です。100クローンの各選択されたプールは、グリセロールストックから10クローンの16セットに細分化された。これらのサブプールは、スクリーニングの第2ラウンド(すなわち、合計3,200の培養井戸)で同じ方法に従って調製し、試験した。サブプール0073-09は最も強い生存率を与え(補足図2A)、コーンエンリッチ培養物のスクリーニングの第3ラウンドで試験した16個のクローンを作製するために細分化された。クローン0073-09-37クローンは明らかに2.5に等しい生存率を持つ他のクローンから際立っています (補足図2B)。なお、y軸は、補助図2Aとはシード密度が同じであった場合でも、スケールが異なる。私たちは、アッセイが何ヶ月も毎週繰り返されるとき、これを一般的に見てきました。分析後、これらの結果はクローン0073-09-37が円錐生存に強く、再現性のある効果を有することを確認した。テストは独立して繰り返され(図3A)、挿入された1.8kbはシーケンス化された(図3B)。
バイオインフォマティクス分析の結果、上皮由来コーン生存率因子(EdCVF)と名付けられたクローン0073-09-37には、ラットタンパク質亜鉛フィンガープロテイン-180(ZFP180、NP_653358 470)のC末端部分の84残基に対して最も上流(ORF1)がコードする3つのオープンリーディングフレーム(ORF)が含まれていることが明らかになった。他の2つのORF(ORF2およびORF3)は、マウスではあまり保存されず、他の哺乳類には存在しない。独立して試験した場合、ORF1だけがコーンに対して保護効果を発揮する(補助図3)。ORF1はグルタチオンSトランスファーゼ(GST)融合タンパク質として産生した(図4A)。このEdCVFタンパク質を精製し、GSTタグを除去した(図4B)。EdCVFは、コーンエンリッチ培養システムにおけるコーン変性を防ぐことができる(図4C)。

図1:鶏の胚を発生期28、29日、30日に矢印W:翼、C:襟とB:法案。スケールバー1cm.この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図2: 鶏胚の残性の解剖この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図3:上皮由来コーン生存率因子(EdCVF)、クローン0073-09-37。A. コーンエンリッチ培養の実行可能性の向上。 B. cDNAクローン0073-09-37の配列。下線付きの列GAATTCは、ライブラリを構築するために使用されるEcoRI制限サイトです。太字の2つのコドンGTGは、ベクトルに由来するEdCVFの珍しい翻訳開始部位です。学生のテストによる統計分析。 この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図4:組換えEdCVF活性. A.グルタチオンS-トランストランスナーゼ(GST)との融合におけるEdCVFの配列。B.精製組換えEdCVFタンパク質。C.培養におけるコーン上のGST-EdCVFの栄養活性Tukeyの検定を用いた統計分析。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}
補足図1:cDNAプールと陰性対照の平均細胞数の比率は、第1回スクリーニング中に空のベクターpcDNA3.1にトランスフェクトされたCOS-1細胞のコンディション培地である。 このファイルをダウンロードするには、ここをクリックしてください。
補足図2:生きている細胞の数。A. 0073プールのサブプールによるスクリーニングの第2ラウンド。B.分離クローンによるスクリーニングの第3ラウンド。CN:空のベクターにトランスフェクトしたCOS-1細胞のコンディション培地であるpcDNA3.1。このファイルをダウンロードするには、ここをクリックしてください。
補助図3:単離クローン0073-09-37の3つの開いた読み取りフレームの栄養活性。 ダネットの検定を用いた統計解析。このファイルをダウンロードするには、ここをクリックしてください。
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
| A | 1 | 1 | 1 | 1 | 2 | NC | 2 | 2 | 2 | 3 | 3 | NC |
| ロ | NC | 3 | 3 | 4 | 4 | 4 | NC | 4 | 5 | 5 | 5 | 5 |
| C | 6 | NC | 6 | 6 | 6 | 7 | 7 | NC | 7 | 7 | 8 | 8 |
| ニ | 8 | 8 | NC | 9 | 9 | 9 | 9 | 10 | NC | 10 | 10 | 10 |
| E | 11 | 11 | 11 | NC | 11 | 12 | 12 | 12 | 12 | NC | 13 | 13 |
| ヘ | 13 | 13 | 14 | 14 | NC | 14 | 14 | 15 | 15 | 15 | NC | 15 |
| G | 16 | 16 | 16 | 16 | 17 | NC | 17 | 17 | 17 | 18 | 18 | NC |
| H | 18 | 18 | 19 | 19 | 19 | 19 | NC | 20 | 20 | 20 | NC | 20 |
| NC ネガティブコントロール | ||||||||||||
| 4倍にテストされたプールの1-20の位置 | ||||||||||||
表1:高内容スクリーニングのための96ウェルプレートの計画
ディスカッション
鶏の胚からのコーン濃縮培養の産生を制限する可能性のある多くのパラメータの中で、最初の重要なステップは、孵化卵中の胚の発達段階を正確に同定することです。ED8(第34段階)における胚のレチナからの細胞の培養は35%の感光体しか産生しない、残りの65%は他のニューロン4で作られていることが観察されている。孵卵を得るために適用されるロジスティックが何であれ、温度とインキュベーション時間を微調整し、開発42、45のすべての段階の参照写真と比較して胚を注意深く調べる必要がある。
もともと、コーンを豊かにした培養システムは、ホワイトレグホーン株4を用いて開発された。その株の卵の白い色はフランスでは特に評価されていないので、茶色の卵を生産する鶏肉の株を使用しました。JA57鶏5とI 66の雄鶏を交わして作られたI 657株を利用しました。オリジナル文化の特徴を再現することができました。これは、鶏肉の遺伝的背景がコーン濃縮培養を得るために重要ではないことを示しています。
我々は、培地中のサプリメントの個々の除去の効果を試験していないが、 インスリンがrd1 マウスにおけるコーンの生存に及ぼすインスリンの効果に従って重要な役割を果たしていることを観察した、常染色体劣性RP46のモデル。トリヨードサイロニン(T3)は、発達40の間のレチナル細胞運命における甲状腺ホルモン受容体の役割に従って、鶏胚の前駆体細胞の円錐への分化にも関与し得る。その結果、コーン濃縮培養システムは、発現クローニング46によってインスリンを同定するために使用することができない。
コーンエンリッチ培養システムは、一次ニューロンの培養に依存し、細胞株661W24、25、26として不死化細胞の使用に依存するはるかに適切なthaメソッドである。
ここで説明する方法は、プラスミドDNA20で前のエレクトロポレーションを行うことにより修飾することができる。レチナル細胞懸濁液を調製する前に、レチナ全体を、10 mM Tris-HCl pH 8.0,1 mM EDTAで0.5 μG/μLのプラスミドDNAの120 μLのカスタムメイドエレクトロポレーターのチャンバーに配置します。50 msのそれぞれ15Vの5つのパルスは950ミリ秒間隔22で分けて適用される。コンネ富化培養物への複製有能な鳥類スプライス(RCAS)レトロウイルスを用いた干渉RNA(RNAi)の送達試行は47に失敗した。これは確かに血清の不在や低密度の培養において、レトロウイルスの伝播に必要なレチナル前駆細胞は複製されないという事実によるものです。
我々は、発現クローニング19を用いてコーン生存を促進する栄養因子を同定するために、コーンを豊かにした培養システムを開発した。それを実現するために、100クローンのプールからコンディションされた培地を用いた高含有スクリーニングの第一歩を行った。ライブラリーからのcDNAがCOS-1細胞のトランスフェクション後に強力なCMVプロモーターの制御下で発現したとしても、個々のcDNAによってコードされるすべてのタンパク質が生存アッセイによって陽性にスコア付けされるのに十分な濃度に達するという保証を提供しない。これは大きな制限事項です。その意味で、スクリーニングは本当に網羅的ではありません。また、膜タンパク質をコンディション培地から除去しなくても、非拡散因子の同定にはアッセイの構成が好ましくない。別の方法は、同じ候補タンパク質を何度もアッセイする重複を避けるためにcDNAの配列を得た後に個々のクローンをスクリーニングすることです。これは、43を使用した retinal cDNA ライブラリのシーケンスによって開始されました。合理的ですが、このアプローチには限界もあります。cDNA配列のバイオインフォマティクス解析は、冗長性の減少に加えて、知識に基づいて特定のクローンをスクリーニングすることの優先順位付けを、たまらなく課す。最終的に、ライブラリ全体が大幅に長くなっても、ライブラリ全体がスクリーニングされる場合、これは有害ではありません。しかし、常に、シーケンスのアイデンティティは、結果を見る私たちの方法に影響を与えます。シーケンスの解釈は自然に実験データと競合するため、これは中立ではありません。
EdCVFの同定は、高いコンテンツスクリーニングには技術的な制限が伴うことも示している。スクリーニングの最初のラウンドから、我々は高い活性を有する2つのプールを同定した(補足図1)。プール0073はEdCVFの識別に成功しましたが、プール0080はそのような発見には行わなかった。サブプールの準備中にアクティブクローンが失われた場合に発生する可能性のある問題は解決していません。あるいは、統計的に良好でなくても、プール0080のcDNAの中で、2つのタンパク質が相乗的に作用しており、その活性を個々のクローンとして観察することができなかったことを排除しない。
小分子をスクリーニングしてコーンを保護する分子の同定は、コーン濃縮培養系の将来の応用である。このような分子は、遺伝子治療が加齢黄斑変性症として最も適切なアプローチではない眼科病変の治療に非常に貴重である。
開示事項
TL、J-ASおよびVFは、EdCVFを使用して、レチン系退性退化を治療するための特許を取得しています[WO2009071659(A1)。2007年6月12日]。
謝辞
著者らは、ジャック・ベラロウ、ローレライ・フルニエ、エマニュエル・クレラン、フレデリック・ブロンド、そして彼らの貴重な助けに対する責任あるア・レスポンサビリテ・リミテ(EARL Morizeauの危険性、フランス)に感謝する。この研究は、インサーム、ソルボンヌ大学、アジェンス・ナショナル・プール・ラ・レシェルシュ(ANR、Labex Lifesenses)、財団ファイティング・ブラインドネス(米国)、IHU FOReSIGHT[ANR-18-IAHU-0001]によって支えられました。
資料
| Name | Company | Catalog Number | Comments |
| 96 black plates (Clear Button with lid Tissue culture treated) | Corning | 3603 | |
| Calcein AM | Thermo-Fisher scientific | C1430 | |
| CCD Camera | Photometrics | CoolSnap FX HQ | |
| CDPC (Cytidine 5′-diphosphocholine sodium salt dihydrate) | Sigma-Aldrich | C0256 | |
| CO2 Independant | Thermo-Fisher scientific | 18045-054 | |
| Curved forceps | Dutscher | 005093 | |
| DMEM Media | Thermo-Fisher scientific | 41966-029 | |
| DNAse | Sigma-Aldrich | D4263 | |
| Eggs incubator | FarmLine | M08 01 3100 | |
| Ethidium Homodimer | Thermo-Fisher scientific | E1169 | |
| Fœtal bovine serum | Thermo-Fisher scientific | 10270-098 | |
| Gentamycin | Thermo-Fisher scientific | 15710-049 | |
| Hydrocortisone | Sigma-Aldrich | H0880 | |
| ITS (insulin Transferine selenium) | Sigma-Aldrich | I1884 | |
| large straight pliers | Dutscher | 005074 | |
| Linoleic acid | Sigma-Aldrich | L8384 | |
| M199 medium | Thermo-Fisher scientific | 31150-022 | |
| Metamorph software | Metamorph | ||
| Microscope | NIKON | Eclipse TE2000 | |
| Motorized stage | Martzauzer | Mutlicontrol 2000 | |
| Optical filter switch | Shutter Instrument company | Lambda 10-2 | |
| PBS 1X | Thermo-Fisher scientific | 14190-086 | |
| Poly-L-lysine | Sigma-Aldrich | P6282 | |
| Progesterone | Sigma-Aldrich | P7556 | |
| Pursept A express | Fisher scientific | 11814110 | |
| Putriscine | Sigma-Aldrich | P5780 | |
| Sodium pyruvate | Sigma-Aldrich | S8636 | |
| straight forceps | Dutscher | 005092 | |
| Taurine | Sigma-Aldrich | T8691 | |
| Triiodothyronine | Sigma-Aldrich | T6397 | |
| Trypan blue | Thermo-Fisher scientific | 15250-061 | |
| Trypsine 0.25 % | Thermo-Fisher scientific | 25200-056 |
参考文献
- Brownstein, C. D. . The Implicit Mind. , (2019).
- Heesy, C. P., Hall, M. I. The nocturnal bottleneck and the evolution of mammalian vision. Brain, Behavior and Evolution. 75 (3), 195-203 (2010).
- Borges, R., et al. Adaptive genomic evolution of opsins reveals that early mammals flourished in nocturnal environments. BMC Genomics. 19 (1), 121 (2018).
- Adler, R., Hatlee, M. Plasticity and differentiation of embryonic retinal cells after terminal mitosis. Science. 243 (4889), 391-393 (1989).
- Fintz, A. C., et al. Partial characterization of retina-derived cone neuroprotection in two culture models of photoreceptor degeneration. Investigative Ophthalmology & Visual Science. 44 (2), 818-825 (2003).
- Roska, B., Sahel, J. A. Restoring vision. Nature. 557 (7705), 359-367 (2018).
- Duncan, J. L., et al. Inherited Retinal Degenerations: Current Landscape and Knowledge Gaps. Translational Vision Science & Technology. 7 (4), 6 (2018).
- Russell, S., et al. Efficacy and safety of voretigene neparvovec (AAV2-hRPE65v2) in patients with RPE65-mediated inherited retinal dystrophy: a randomised, controlled, open-label, phase 3 trial. Lancet. 390 (10097), 849-860 (2017).
- RetNet. . Retinal Information Network. , (2020).
- Doonan, F., Donovan, M., Cotter, T. G. Activation of multiple pathways during photoreceptor apoptosis in the rd mouse. Investigative Ophthalmology & Visual Science. 46 (10), 3530-3538 (2005).
- Comitato, A., et al. Dominant and recessive mutations in rhodopsin activate different cell death pathways. Human Molecular Genetics. 25 (13), 2801-2812 (2016).
- Cronin, T., Leveillard, T., Sahel, J. A. Retinal degenerations: from cell signaling to cell therapy; pre-clinical and clinical issues. Current Gene Therapy. 7 (2), 121-129 (2007).
- Baumgartner, W. A., Baumgartner, A. M. Accounting for disagreements on average cone loss rates in retinitis pigmentosa with a new kinetic model: Its relevance for clinical trials. Medical Hypotheses. 89, 107-114 (2016).
- Leveillard, T., Sahel, J. A. Rod-derived cone viability factor for treating blinding diseases: from clinic to redox signaling. Science Translational Medicine. 2 (26), (2010).
- Leveillard, T., Sahel, J. A. Metabolic and redox signaling in the retina. Cellular and Molecular Life Sciences. 74 (20), 3649-3665 (2017).
- Clerin, E., Marussig, M., Sahel, J. A., Leveillard, T. Metabolic and Redox Signaling of the Nucleoredoxin-Like-1 Gene for the Treatment of Genetic Retinal Diseases. International Journal of Molecular Sciences. 21 (5), (2020).
- Fridlich, R., et al. The Thioredoxin-like Protein Rod-derived Cone Viability Factor (RdCVFL) Interacts with TAU and Inhibits Its Phosphorylation in the Retina. Molecular & Cellular Proteomics. 8 (6), 1206-1218 (2009).
- Mei, X., et al. The Thioredoxin Encoded by the Rod-Derived Cone Viability Factor Gene Protects Cone Photoreceptors Against Oxidative Stress. Antioxidants, Redox Signaling. 24 (16), 909-923 (2016).
- Leveillard, T., et al. Identification and characterization of rod-derived cone viability factor. Nature Genetics. 36 (7), 755-759 (2004).
- Vergara, M. N., Gutierrez, C., Canto-Soler, M. V. Efficient Gene Transfer in Chick Retinas for Primary Cell Culture Studies: An Ex-ovo Electroporation Approach. Journal of Visualized Experiments. (105), e52002 (2015).
- Fridlich, R., et al. The thioredoxin-like protein rod-derived cone viability factor (RdCVFL) interacts with TAU and inhibits its phosphorylation in the retina. Molecular & Cellular Proteomics. 8 (6), 1206-1218 (2009).
- Ait-Ali, N., et al. Rod-derived cone viability factor promotes cone survival by stimulating aerobic glycolysis. Cell. 161 (4), 817-832 (2015).
- Mei, X., et al. The Thioredoxin Encoded by the Rod-Derived Cone Viability Factor Gene Protects Cone Photoreceptors Against Oxidative Stress. Antioxidants & Redox Signaling. 24 (16), 909-923 (2016).
- Tan, E., et al. Expression of cone-photoreceptor-specific antigens in a cell line derived from retinal tumors in transgenic mice. Investigative Ophthalmology & Visual Science. 45 (3), 764-768 (2004).
- Wang, X. W., Tan, B. Z., Sun, M., Ho, B., Ding, J. L. Thioredoxin-like 6 protects retinal cell line from photooxidative damage by upregulating NF-kappaB activity. Free Radical Biology and Medicine. 45 (3), 336-344 (2008).
- Perron, N. R., Beeson, C., Rohrer, B. Early alterations in mitochondrial reserve capacity; a means to predict subsequent photoreceptor cell death. Journal of Bioenergetics and Biomembranes. 45 (1-2), 101-109 (2013).
- Krishnamoorthy, R. R., Clark, A. F., Daudt, D., Vishwanatha, J. K., Yorio, T. A forensic path to RGC-5 cell line identification: lessons learned. Investigative Ophthalmology & Visual Science. 54 (8), 5712-5719 (2013).
- Al-Ubaidi, M. R. RGC-5: are they really 661W? The saga continues. Experimental Eye Research. 119, 115 (2014).
- Clerin, E., et al. Vibratome sectioning mouse retina to prepare photoreceptor cultures. Journal of Visualized Experiments. (94), (2014).
- Hendrickson, A., Hicks, D. Distribution and density of medium- and short-wavelength selective cones in the domestic pig retina. Experimental Eye Research. 74 (4), 435-444 (2002).
- Bobu, C., Craft, C. M., Masson-Pevet, M., Hicks, D. Photoreceptor organization and rhythmic phagocytosis in the nile rat Arvicanthis ansorgei: a novel diurnal rodent model for the study of cone pathophysiology. Investigative Ophthalmology & Visual Science. 47 (7), 3109-3118 (2006).
- Saidi, T., Mbarek, S., Chaouacha-Chekir, R. B., Hicks, D. Diurnal rodents as animal models of human central vision: characterisation of the retina of the sand rat Psammomys obsesus. Graefes Arch Clin Exp Ophthalmol. 249 (7), 1029-1037 (2011).
- Traverso, V., Kinkl, N., Grimm, L., Sahel, J., Hicks, D. Basic fibroblast and epidermal growth factors stimulate survival in adult porcine photoreceptor cell cultures. Investigative Ophthalmology & Visual Science. 44 (10), 4550-4558 (2003).
- Balse, E., et al. Purification of mammalian cone photoreceptors by lectin panning and the enhancement of their survival in glia-conditioned medium. Investigative Ophthalmology & Visual Science. 46 (1), 367-374 (2005).
- Gagliardi, G., et al. Characterization and Transplantation of CD73-Positive Photoreceptors Isolated from Human iPSC-Derived Retinal Organoids. Stem Cell Reports. 11 (3), 665-680 (2018).
- Gagliardi, G., Ben M'Barek, K., Goureau, O. Photoreceptor cell replacement in macular degeneration and retinitis pigmentosa: A pluripotent stem cell-based approach. Progress in Retinal and Eye Research. 71, 1-25 (2019).
- Mears, A. J., et al. Nrl is required for rod photoreceptor development. Nature Genetics. 29 (4), 447-452 (2001).
- Swaroop, A., Kim, D., Forrest, D. Transcriptional regulation of photoreceptor development and homeostasis in the mammalian retina. Nature Reviews Neuroscience. 11 (8), 563-576 (2010).
- Kallman, A., et al. Investigating cone photoreceptor development using patient-derived NRL null retinal organoids. Nature Communications. 3 (1), 82 (2020).
- Eldred, K. C., et al. Thyroid hormone signaling specifies cone subtypes in human retinal organoids. Science. 362 (6411), (2018).
- Kole, C., et al. Otx2-Genetically Modified Retinal Pigment Epithelial Cells Rescue Photoreceptors after Transplantation. Molecular Therapy. 26 (1), 219-237 (2018).
- Hamburger, V., Hamilton, H. L. A series of normal stages in the development of the chick embryo. Journal of Morphology. 88 (1), 49-92 (1951).
- Kole, C., et al. Identification of an Alternative Splicing Product of the Otx2 Gene Expressed in the Neural Retina and Retinal Pigmented Epithelial Cells. PLoS One. 11 (3), 0150758 (2016).
- Shannon, M., Hamilton, A. T., Gordon, L., Branscomb, E., Stubbs, L. Differential expansion of zinc-finger transcription factor loci in homologous human and mouse gene clusters. Genome Research. 13 (6), 1097-1110 (2003).
- Stern, C. D., Holland, P. W. . Essential developmental biology: a practical approach. , (1993).
- Punzo, C., Kornacker, K., Cepko, C. L. Stimulation of the insulin/mTOR pathway delays cone death in a mouse model of retinitis pigmentosa. Nature Neuroscience. 12 (1), 44-52 (2009).
- Harpavat, S., Cepko, C. L. RCAS-RNAi: a loss-of-function method for the developing chick retina. BMC Developmental Biology. 6, 2 (2006).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved