
Method Article
Culture arricchite di coni dalla retina degli embrioni di pollo per studiare le interazioni cellulari tra bastone e cono
In questo articolo
Riepilogo
Descriviamo un metodo per ottenere colture primarie di fotorecettori a cono dalla retina degli embrioni di pollo e il suo uso per lo screening ad alto contenuto.
Abstract
La visione umana diurna si basa sulla funzione dei fotorecettori a cono al centro della retina, la fovea. I pazienti affetti dalla forma più diffusa di degenerazione retinica ereditaria, retinite pigmentosa, perdono la visione notturna a causa della perdita guidata dalla mutazione dei fotorecettori dell'asta, un fenomeno seguito da una progressiva perdita di funzione e morte di coni che porta alla cecità. I genetisti hanno identificato molti geni con mutazioni che causano questa malattia, ma le prime mutazioni identificate hanno messo in discussione i meccanismi della degenerazione secondaria del cono e come una mutazione dominante nella codifica genica della rodopsina per il pigmento visivo espresso esclusivamente nelle aste possa innescare la degenerazione del cono.
Questo risultato dei trapianti in un modello genetico della malattia ha portato al concetto di interazioni cellulari tra canne e coni e di degenerazione autonoma non cellulare dei coni in tutte le forme genetiche di retinite pigmentosa.
I coni comprendono il 5% di tutti i fotorecettori nell'uomo e solo il 3% nel topo, quindi il loro studio è difficile in queste specie, ma i coni superano le aste nelle specie di uccelli. Abbiamo adattato 96 piastre di pozzo a precursori retinali di coltura dalla retina degli embrioni di pollo nella fase 29 del loro sviluppo. In queste colture primarie, i coni rappresentano l'80% delle cellule dopo la differenziazione in vitro. Le cellule degenerano per un periodo di una settimana in assenza di siero. Qui descriviamo i metodi e la sua standardizzazione.
Questo sistema di coltura arricchito di coni è stato utilizzato per identificare il fattore di vitalità del cono derivato dall'epitelio (EdCVF) mediante screening ad alto contenuto di una libreria di cDNA normalizzata di epitelio pigmentato retinico di ratto. L'EdCVF ricombinante impedisce la degenerazione dei coni.
Introduzione
La retina delle specie di vertebrati è doppia, con fotorecettori a bastone per una visione fioca della luce e fotorecettori a cono per la visione della luce del giorno, del colore e dell'acuità. L'acuità visiva dei primati si basa su una regione al centro della retina, chiamata fovea, che si arricchisce di coni, ma nel complesso i coni rappresentano solo il 5% di tutti i fotorecettori. Di conseguenza, l'analisi dei coni nella retina dei primati e in particolare la cultura dei coni sono tecnicamente difficili. Tutte le altre specie di mammiferi non hanno fovea e la percentuale di coni è bassa per i roditori che sono più comunemente usati nella ricerca retinica. Non è così per le specie aviari, per le quali i coni dominano la retina di queste specie di uccelli ben vedendo. I dinosauri, che hanno dominato l'ecosistema quando i mammiferi sono apparsi per la prima volta durante l'evoluzione, sono all'origine filogenetica degli uccelli1. Come conseguenza di tale competizione tra dinosauri e primi mammiferi, i mammiferi sono per lo più notturni con retine dominate da aste. Solo più tardi durante l'evoluzione la visione diurna di alcune specie di mammiferi, tra cui i primati appartengono, divenne un vantaggio evolutivo. Tuttavia, il periodo ancestrale rimane come atavismo del collo di bottiglia notturno nell'evoluzione della visione dei mammiferi2,3.
Mentre studiavano la differenziazione cellulare retinica, Adler e Hatlee hanno dimostrato che i fotorecettori rappresentano circa il 70% delle cellule differenziate retiniche nelle colture derivate dal pollo al giorno embrionale (ED) 6 o stadio 294. A causa della prevalenza di coni nella retina di pollo, colture di cellule retiniche provenienti da embrioni di pollo ED6 sono state sviluppate come colture arricchite di coni5.
L'importanza dell'acuità visiva mediata dal cono per l'uomo è un truismo. Le persone affosse di malattie genetiche o dell'invecchiamento che alterano la funzione del cono sono notevolmente svantaggiate. Questo ha promosso un corpo molto ampio di studi sulle degenerazioni retiniche ereditarie (IRD) con l'obiettivo di trovare trattamenti per queste malattieaccecante 6,7. Il primo successo, ottenuto utilizzando un vettore adeno-associato ricombinante (AAV) per la terapia di una forma grave di amaurosi congenita leber IRD (LCA), è una prova del concetto per la terapia genica8. L'identificazione dei geni le cui mutazioni innescano l'IRD apre la possibilità di curare queste malattie utilizzando la terapia genica. Tuttavia, queste malattie sono il risultato di mutazioni in più di 200 geni distinti9. Anche nel caso di forme autosomiche recessive di IRD, quando la reintroduzione della copia normale del gene morboso potrebbe ripristinare la funzione visiva, il costo economico di ogni singolo sviluppo favorisce quelli più diffusi a scapito di quelli meno comuni e di quelli per i quali l'origine genetica rimane sconosciuta. Questo fatto ha portato i ricercatori a pensare a terapie più generali. La morte cellulare apoptotica è apparsa come una via comune e un bersaglio terapeutico di queste malattie che progrediscono per degenerazione di fotorecettori, anche per le forme autosomidominanti 10,11. Tuttavia, mancano i successi di tale approccio. Per la forma più comune di IRD, retinite pigmentosa (RP), la via comune è la perdita secondaria della funzione alla fine seguita dalla degenerazione dei coni12,13. Prevenire la perdita della funzione del cono conserverà la visione centrale dei fovea indipendentemente dalle mutazioni causali14.
Nella fase iniziale della RP, la perdita di aste innesca una riduzione dell'espressione del fattore di vitalità del cono derivato dall'asta (RdCVF), codificato dal gene nucleoredossina-simile a 1(NXNL1),che interrompe la segnalazione metabolica e redox tra aste e coni15. La somministrazione di un AAV ricombinante che codifica per i due prodotti del gene NXNL1, il fattore trolico RdCVF e l'enzima tioredossina RdCVFL, potrebbe teoricamente prevenire la perdita della vista del cono in tutte le forme genetiche di RP16. Abbiamo dimostrato che il prodotto genetico NXNL1, RdCVFL, è espresso in colture arricchite con coni dipollo 17 e dove svolge un ruolo protettivo18. RdCVF e il gene NXNL1 sono stati identificati mediante screening ad alto contenuto di una libreria cDNA retinica utilizzando la sopravvivenza delle cellule da una coltura arricchita di coni come readout19. Abbiamo proiettato l'equivalente di 210.000 singoli cloni della libreria usando 8 test paralleli per ogni clone. Ciò rappresenta un gran numero di test che richiedono un facile accesso al materiale biologico, le retine degli embrioni di pollo. Abbiamo scoperto che era relativamente facile ottenere uova di gallina embrionale su base settimanale perché sono ampiamente prodotte per l'agroindustria per galline ovaiole e polli da carne. Dopo un'attenta standardizzazione delle colture arricchite di coni, il sistema fornisce un modo semplice, robusto e riproducibile per testare migliaia di molecole per la loro capacità di preservare la vitalità del cono. Queste cellule sono anche suscettibili alle manipolazioni genetiche20 che beneficiano dello studio della trasduzione del segnale e delle analisibiochimiche 21,22,23.
I ricercatori retinali hanno sviluppato metodi alternativi come l'uso della linea cellulare del cono 661W24,25,26. Tuttavia, l'identità di questa linea cellulare rimane controversa27,28. Le cellule 661W sono state clonate da tumori retinali di una linea transgenica di topo che esprime il grande antigene T SV40 sotto il controllo del promotore umano di proteine leganti il retinolo interfotorecettore. SV40 grande antigene T media la trasformazione cellulare e l'immortalizzazione. Di conseguenza, la via di segnalazione identificata utilizzando cellule da 661W deve essere segnalata nel contesto di una linea cellulare trasformata e immortalata che è distinta per molti versi dai coni in situ. A questo proposito, il sistema di coltura arricchito di coni è composto da neuroni primari, i coni che sono più fisiologicamente rilevanti.
Mentre è possibile ottenere una coltura pura di fotorecettori utilizzando la sessatura vibratoma della retina del topo, la bassissimo percentuale di coni nella retina esterna dei roditori rende questo approccio inadatto per produrre colture arricchite di coni29. La retina di maiale non contiene fovea ma ha una regione chiamata area centralis che è molto arricchita in coni30. L'alta percentuale di coni nei roditori diurni della retina, come Arvicanthis ansorgei e Psammomys obsesus31,32, offre una possibile soluzione ma richiede l'allevamento di tali specie esotiche. Gli occhi adulti di maiale, raccolti dai macelli locali, possono essere utilizzati per produrre una coltura mista di aste e coni che sono stati utilizzati per studiare la sopravvivenza del fotorecettore33. Una soluzione elegante è la pre-purificazione dei coni dalla retina di maiale utilizzando la panning con lectina di arachidi agglutinina (PNA), che si lega selettivamente ai coni34. Tuttavia, questo metodo è difficile da implementare su larga scala a causa della sua complessità.
Le cellule staminali pluripotenti indotte dall'uomo (iPS) offrono l'approccio più promettente per ottenere una popolazione cellulare fotorecettoria a cono che può essere utilizzata per il trapianto retinale ma che può anche essere adattata alla coltura arricchita di coni35,36. Poiché il fattore di trascrizione NRL è richiesto per i fotorecettori a canna37, il topo Nrl-/- ha una retina dominata da coni ad onda corta (Coni S). L'inattivazione potrebbe essere utilizzata per produrre una preparazione arricchita con cono di S differenziando l'uomo da iPS38,39. Un altro possibile approccio è quello di promuovere la differenziazione del cono utilizzando l'ormonetiroideo segnalazione 40. Mentre stanno emergendo nuovi metodi per produrre colture arricchite di coni da iPS umani, gli embrioni di pollo forniscono un metodo collaudato19.
La cultura arricchita di coni è stata determinante per l'identificazione di RdCVF mediante clonazione diespressione 19. Questo sistema è stato anche utilizzato con successo per dimostrare che RdCVF stimola l'assorbimento del glucosio e il suo metabolismo mediante glicolisi aerobica22. Inoltre, la coltura arricchita di coni è stata utilizzata per convalidare il ruolo protettivo di RdCVFL, il secondo prodotto del gene NXNL1 23. Più recentemente, questo sistema è stato utilizzato per dimostrare l'esistenza di molecole protettive secrete da cellule epiteliali pigmentate retiniche trasdotto con OTX241.
Protocollo
Il protocollo è stato approvato dal Comitato per l'etica degli esperimenti sugli animali dell'Università Pierre e Marie Curie e dal ministero francese della Ricerca (Numero di permesso: APAFIS#1028 2015070211275177). Gli esperimenti sugli animali furono eseguiti con la seguente autorizzazione: "Certificat d'autorisation d'expérimenter sur les animaux vertébrés A-75-1863. Préfecture de Police de Paris (9 novembre 2011 -8 novembre 2016)".
1. Incubazione di uova fecondate
- Raccogliere uova fecondate settimanali (ceppo I 657, etichetta rossa), ottenute naturalmente, in un vivaio industriale.
- Mantenere le uova fecondate a 17 °C (il loro zero biologico) in laboratorio dopo essere state "deposte" dalla gallina.
- Per ogni coltura, incubare sette uova fecondate per 24 ore a 20 °C e poi 136 ore a 37 °C con reversione intermittente dell'inclinazione (un movimento progressivo per 2 ore da un lato all'altro, un ciclo di 4 ore) delle uova in una camera umidificata.
2. Recupero degli embrioni di pollo
- Lavare la superficie delle sette uova con disinfettante (ad esempio, Pursept A express).
- Per rompere il guscio d'uovo, fai un buco nella parte superiore del guscio con grandi pinze dritte. Quindi tagliare il guscio per rimuovere il cappello dall'uovo come un uovo sodo.
- Estrarre delicatamente ogni embrione dal guscio d'uovo con forcelle curve, quindi trasferirlo in una piastra di Petri contenente tampone di fosfato sterile saline (PBS) precedentemente riscaldata a 37 °C. Rimuovere delicatamente l'involucro che circonda gli embrioni (il corore o la membrana corioallantoica).
- Verificare lo stadio di sviluppo di ogni embrione tramite confronto visivo con Hamburger e Hamilton42.
- Selezionare due embrioni nella 29afase di sviluppo (Figura 1). Le ali si piegano ai gomiti. Il colletto si distingue visibilmente. Il disegno di legge è più importante che alla28esima tappa.
- Enucleare gli occhi di questi embrioni selezionati e trasferirli in mezzo indipendente dalla CO2(tecnologie Life).
ATTENZIONE: È molto importante che l'embrione si trova nella fase 29 e non nelle fasi 28 o 30(figura 1); ecco perché è necessario incubare almeno 7 uova, anche se solo due saranno finalmente utilizzate. Il corione è molto sottile, così difficile da distinguere, ma è molto vicino all'embrione, quindi deve essere rimosso senza toccare l'embrione.
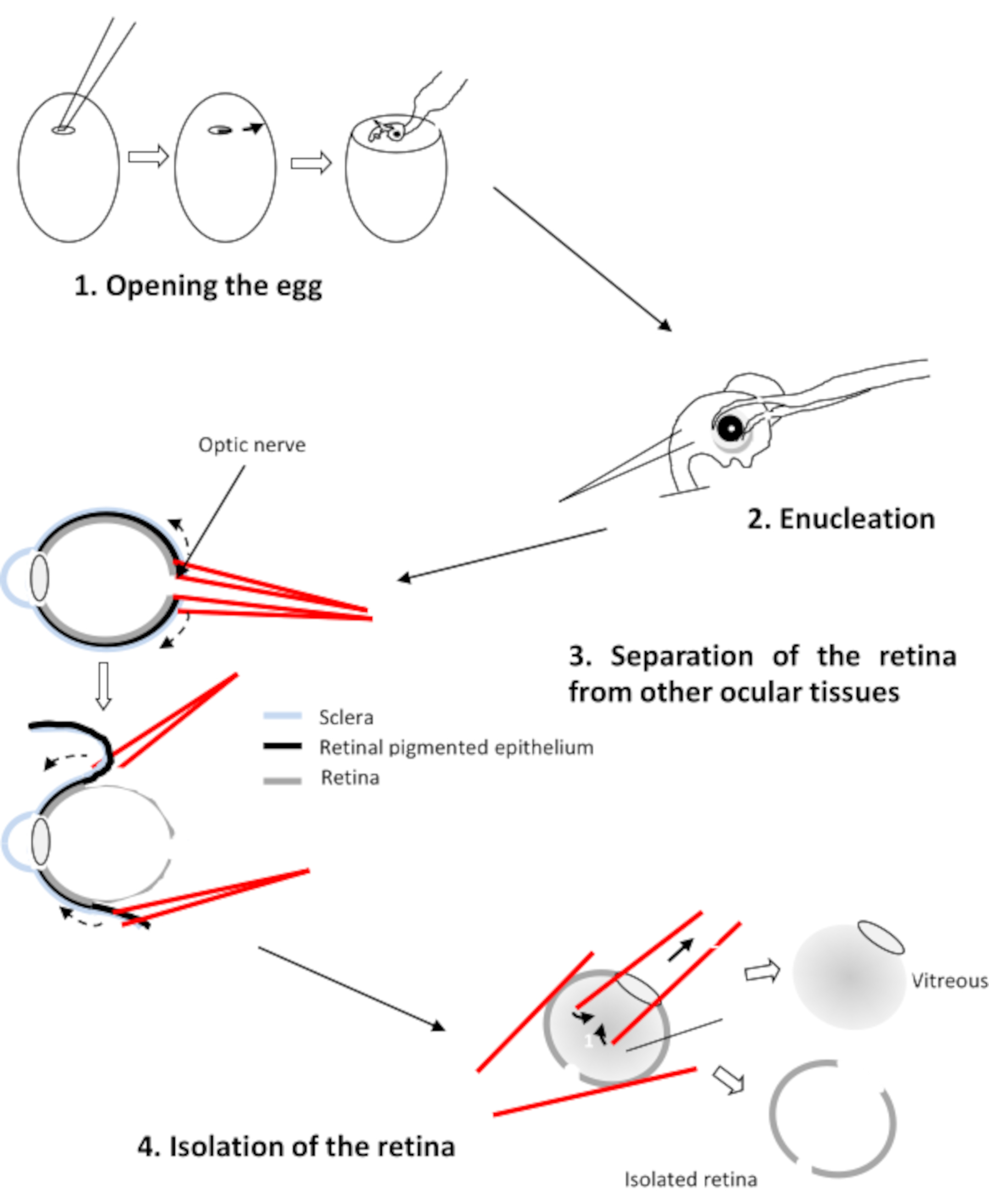
3. Dissezione delle retine
- Decapitare ed enucleare gli embrioni selezionati con forcep curve.
- Trasferire i quattro occhi in un mezzo indipendente dalla CO2. Questo mezzo contiene 0,9 mM CaCl2 e 0,65 mM MgCl2.
- Posizionare l'occhio con la cornea a faccia in giù, il nervo ottico rivolto verso lo sperimentatore. Praticare un foro nel nervo ottico usando due forcette dritte.
- Inserire un ramo di ogni forcep tra la retina e l'epitelio pigmentato (Figura 2). Tirare su ogni forcep e ruotare l'occhio per staccare l'epitelio dalla retina. Rimuovere la cornea seguita dalla lente e dal vitreo.
- Trasferire le quattro retine in una piastra di Petri contenente il mezzo di Ringer a pH 7.2.
ATTENZIONE: Assicurarsi che rimanga solo la retina e rimuovere eventuali tracce di epitelio pigmentato vitreo e rimanente retinico.
4. Preparazione della sospensione cellulare retinica
- Tagliare le quattro retine in pezzi molto piccoli usando due pinze dritte.
- Lavare i pezzi della retina due volte con il mezzo di Ringer.
- Dopo il secondo lavaggio con il mezzo di Ringer, lasciare cadere i pezzi di retina sul fondo del tubo e rimuovere il mezzo della suoneria. Trattare i pezzi retinali per 20 minuti a 37 °C con una soluzione di tripsidenza (0,25% w/v).
- Disperdere la soluzione dopo 10 minuti mediante successiva aspirazione. Scaricare utilizzando una pipetta Pasteur e verificare la dissociazione dei pezzi retinali. Interrompere la reazione aggiungendo mezzi di coltura integrati con siero fetale del vitello inattivato al 10%.
- Incubare la sospensione cellulare con 0,05 mg di DNasi I. Dissociare i cluster cellulari e il DNA mediante successiva aspirazione e scarico utilizzando una pipetta Pasteur subito dopo l'aggiunta della DNasi.
- Lavare la sospensione cellulare retinica due volte con un mezzo di coltura definito chimicamente (CDCM): un volume uguale del mezzo aquila modificato di Dulbecco e del supporto M199 integrato con acido linoleico da 100 μg/mL/BSA, 0,86 μM di insulina, 0,07 μM transferrina, 2,0 μM progesterone, 0,28 μM di prostaglandina, 0,29 μM Na2SeO3, 182 μM putrescina, taurina da 3 mM, 4,7 μM di citidina 5′-difosfocolina, 2,7 μM citidina 5′-difosfoetanolammina, 0,55 μM idrocortisone, 0,03 μM triiodotironina, 1 mM di piruvato di sodio e 20 μM di gentamicina.
5. Semina di cellule retiniche
- Trattare due piastre di coltura nere da 96 polietilene con fondo trasparente per 2 ore a 37 °C con poli-L-lisina a 32,25 μg/cm2.
- Risciacquare questi piatti due volte con il mezzo di coltura M199. Rimosi il pellet di cella in 1 mL di CDCM.
- Aggiungere ad un'aliquota di 10 μL della sospensione cellulare trypan blu per macchiare le cellule viventi. Aggiungere il campione di sospensione delle cellule a un emocitometro (camera di conteggio cellulare di Malassez).
- Al microscopio, contare le celle macchiate per quattro file dell'emocitometro (cioè 40 quadrati), quindi calcolare il numero medio di cellule per una riga. Calcolare la concentrazione di cellule della sospensione applicando il seguente metodo: Numero di celle/mL di sospensione = numero medio di cellule in una riga (10 quadrati) x 10 il numero totale di quadrati dell'emocitometro x diluizione con tripano blu x 1.000 (per esprimere il risultato come celle/mL).
- Portare la sospensione cellulare a due concentrazioni (5,6 x 104 celle/mL e 1,12 x 105 celle/mL) corrispondenti alle due densità di placcatura (1 x 105 celle/cm2 e 2 x 105 celle/cm 2)utilizzando cdcm.
- Seme 50 μL delle due sospensioni cellulari nelle due piastre di coltura nere pretrattate da 96 pozzi. Distribuire le celle nelle piastre con una pipetta multicanale da destra della piastra a sinistra, omogeneizzando tra ogni colonna, in modo che la distribuzione delle cellule sia omogenea.
- Aggiungere 50 μL della libreria di molecole (ad esempio, il supporto condizionato di una libreria cDNA, vedi sotto) da migliorare utilizzando un modello predefinito (Tabella 1).
- Incubare le piastre per sette giorni a 37°C sotto il 5% di CO2 senza alcun cambiamento di supporto.
6. Conteggio di cellule vitali
- Ad ogni pozzo della piastra aggiungere 2,7 μM di calceina AM e 0,3 mM di omodimero di etidio.
NOTA: La calceina penetra nelle cellule impermeabili alle grandi molecole come omodimero di etidio. Nel citoplasma delle cellule viventi, la calceina viene idrolizzata da esterasi endogeni, diventa fluorescente emettendo a 520 nm quando eccitata a 485 nm. L'omodimero dell'etidio si lega al DNA sulle cellule morte. Il legame DNA-omodimero dell'etidio fa sì che la fluorescenza rossa venga emessa a 635 nm dopo l'eccitazione a 520 nm. - Incubare le piastre per 1 ora a temperatura ambiente in assenza di luce.
- Leggere la fluorescenza su un lettore automatico di lastre composto da un microscopio invertito dotato di una lampada a mercurio con due filtri di eccitazione a 485 e 520 nm, due filtri di emissione a 520 e 635 nm, un obiettivo (x10), uno stadio motorizzato controllato da un processore e una telecamera del dispositivo ad accoppiamento di carica (CCD). Questa piattaforma di conteggio è controllata dal software Metamorph19. Permette di acquisire contemporaneamente immagini a fluorescenza di cellule viventi e cellule morte in ogni pozzo della piastra da 96 pozza, partendo dai pozzi A1 verso il pozzo A12, quindi B12 verso B1, C1 verso C12 e così via(Tabella I).
- Calcolare l'area media A di una singola cella utilizzando i 18 pozzi dei controlli negativi (Tabella I).
- Contare le cellule in ogni pozzo della piastra e applicare la seguente formula empirica A x 29 / 20.7 per evitare che i farsa delle cellule (raggruppamento di due celle) siano contati. Segnare l'effetto protettivo dei coni per molecole come rapporto tra il numero medio di cellule nei 4 pozzi in cui abbiamo testato la molecola rispetto al numero medio di cellule nei 18 pozzi del controllo negativo (vedi tabella 1 e figura complementare 1).
- Unire i risultati della piastra seminata a 1 x 105 celle/cm2 con quella seminata a 2 x 105 celle/cm2 per valutare la protezione potenziale (rapporto di vitalità) da parte di ciascuna molecola schermata.
Risultati
Descriviamo qui come il sistema di coltura arricchito di coni può essere utilizzato per identificare nuove proteine che proteggono il cono. Abbiamo usato questo protocollo per schermare una libreria cDNA normalizzata fatta di epitelio pigmentato coroide e retinico da 400 occhi di ratti Long-Evans di 8 settimane43.
Questa libreria contiene 6,0 x 106 colonie indipendenti che formano unità (CNO) e ha una dimensione media di inserto clonata di 2,1 kilobase (kb), con oltre il 99% dei cloni ricombinanti. Le pozze di 100 cloni di quella libreria sono state transiently trasfette (0,1 μg di DNA plasmide) in cellule COS-1 e il mezzo condizionato (CM) di COS-1 è stato raccolto dopo l'incubazione per 48 ore in DMEM senza siero. Membrane o esosomi non sono stati rimossi dall'ultracentrifugazione. Cinquanta microlitri di ogni CM sono stati aggiunti a 4 pozzi di due piastre da 96 pozzi: uno seminato a 2 x 105 celle /cm2,l'altro a 4 x 105 celle/cm2. Cm da cellule COS-1 trasfette con il vettore vuoto pcDNA3.1 utilizzato per costruire la libreria è stato utilizzato come controllo negativo (Tabella 1). Un totale di 2.112 set di 100 cloni corrispondenti a 211.200 cloni individuali sono stati valutati in quattro pozzi di coltura e per due condizioni di semina di colture arricchite di coni. Le due condizioni corrispondono a due densità di inoculazione leggermente diverse che consente di valutare più precisamente l'attività protettiva, per un totale di 1.689.600 pozzi di coltura.
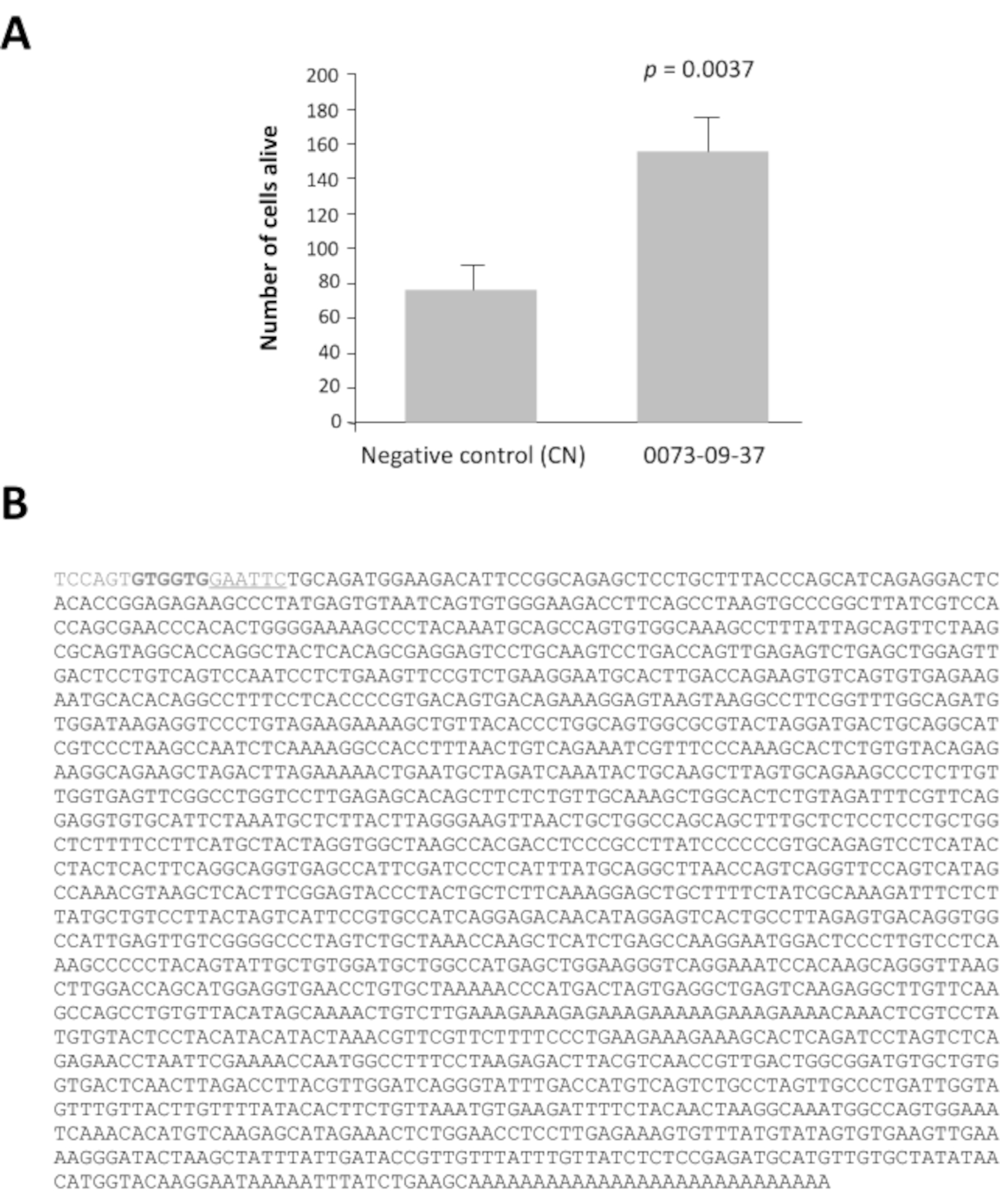
Tra i 42 pool di cloni con un rapporto maggiore di 2, i pool 0080 e 0073 hanno un rapporto di vitalità 16 e 14 volte superiore dopo 7 giorni di coltura rispetto al controllo negativo, pcDNA3.1 (Figura complementare 1). Questa analisi è essenziale per identificare i pool di interesse. Ogni pool selezionato di 100 cloni era suddiviso in 16 set di 10 cloni dal loro stock di glicerolo. Questi sottocomiti sono stati preparati e testati secondo lo stesso metodo in un secondo ciclo di screening (cioè un totale di 3.200 pozzi di coltura). Il sotto-pool 0073-09 ha dato il più forte rapporto di vitalità (Figura supplementare 2A) ed è stato suddiviso per produrre 16 cloni individuali che sono stati testati in un terzo ciclo di screening su colture arricchite di coni. Il clone 0073-09-37 si distingue chiaramente dagli altri con un rapporto di vitalità pari a 2,5 (Figura supplementare 2B). L'asse y ha una scala diversa anche se la densità di semina era la stessa che nella figura complementare 2A. Lo abbiamo visto comunemente quando i test vengono ripetuti settimanalmente per mesi. Dopo l'analisi, questi risultati confermano che il clone 0073-09-37 ha un effetto robusto e riproducibile sulla sopravvivenza del cono. La prova è stata ripetuta in modo indipendente (Figura 3A) e l'inserto di 1,8 kb è stato sequenziato (Figura 3B).
Un'analisi bioinformatica ha rivelato che il clone 0073-09-37, che abbiamo chiamato fattore di vitalità del cono derivato dall'epitelio (EdCVF), contiene tre telai di lettura aperti (ORF), quello che il più a monte (ORF1) codifica per 84 residui della parte C-terminale della proteina del ratto zinco finger proteina-180 (ZFP180, NP_653358) di 727 amminoacidi44. Gli altri due ORF (ORF2 e ORF3) sono molto meno ben conservati nei topi e assenti in altri mammiferi. Se testato in modo indipendente, solo ORF1 esercita un effetto protettivo sui coni (Figura complementare 3). L'ORF1 è stato prodotto come proteina di fusione glutatione S-transferasi (GST)(Figura 4A). La proteina EdCVF è stata purificata e il tag GST rimosso (Figura 4B). EdCVF è in grado di prevenire la degenerazione del cono nel sistema di coltura arricchito di coni (Figura 4C).

Figura 1: Embrioni di pollonelle fasi 28,29e 30dello sviluppo. Frecce W: ala, C: colletto e B: becco. Barra di scala 1 cm.
{kind=link}

Figura 2: Dissezione della retina dell'embrione di pollo Fare clic qui per visualizzare una versione più ampia di questa cifra.
{kind=link}

Figura 3: Fattore di vitalità del cono derivato dall'epitelio (EdCVF), clone 0073-09-37. A. La commissione per l'a Maggiore vitalità sulla cultura arricchita di coni. B. La commissione per l' Sequenza del clone cDNA 0073-09-37. La sequenza sottolineata GAATTC è il sito di restrizione EcoRI utilizzato per costruire la libreria. I due codoni GTG in grassetto sono siti di iniziazione alla traduzione non comuni per EdCVF provenienti dal vettore. Analisi statistica per test dello Studente. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Attività di EdCVF ricombinante. A. Sequenza di EdCVF nella fusione con glutatione S-transferasi (GST). B. La commissione per l' La proteina EdCVF ricombinante purificata. C. La commissione per l' Attività trofica di GST-EdCVF sul cono in coltura. Analisi statistica utilizzando il test di Tukey. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}
Figura complementare 1: Rapporto tra il numero di cella medio per un pool di cDNA e il controllo negativo, il mezzo condizionato delle cellule COS-1 trasfettato con il vettore vuoto pcDNA3.1 durante il primo ciclo di screening. Clicca qui per scaricare questo file.
Domanda complementare Figura 2: Numero di cellule vive. A. La commissione per l'a Il secondo giro di screening con sotto-piscine di piscina 0073. B. La commissione per l' Il terzo round di screening con cloni isolati. CN: il mezzo condizionato delle cellule COS-1 trasfette con il vettore vuoto, pcDNA3.1. Clicca qui per scaricare questo file.
Figura complementare 3: Attività trofica dei tre fotogrammi di lettura aperti del clone isolato 0073-09-37. Analisi statistica utilizzando il test di Dunnett. Clicca qui per scaricare questo file.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
| un | 1 | 1 | 1 | 1 | 2 | Nc | 2 | 2 | 2 | 3 | 3 | Nc |
| B | Nc | 3 | 3 | 4 | 4 | 4 | Nc | 4 | 5 | 5 | 5 | 5 |
| C | 6 | Nc | 6 | 6 | 6 | 7 | 7 | Nc | 7 | 7 | 8 | 8 |
| re | 8 | 8 | Nc | 9 | 9 | 9 | 9 | 10 | Nc | 10 | 10 | 10 |
| ecstasy | 11 | 11 | 11 | Nc | 11 | 12 | 12 | 12 | 12 | Nc | 13 | 13 |
| fa | 13 | 13 | 14 | 14 | Nc | 14 | 14 | 15 | 15 | 15 | Nc | 15 |
| sol | 16 | 16 | 16 | 16 | 17 | Nc | 17 | 17 | 17 | 18 | 18 | Nc |
| H | 18 | 18 | 19 | 19 | 19 | 19 | Nc | 20 | 20 | 20 | Nc | 20 |
| Controlli negativi NC | ||||||||||||
| 1-20 posizioni delle piscine testate in quadruplicazione | ||||||||||||
Tabella 1: Piano della piastra da 96 pozzi per lo screening ad alto contenuto
Discussione
Tra i molti parametri che potrebbero limitare la produzione di una coltura arricchita di coni da embrioni di pollo, il primo passo critico è identificare con precisione lo stadio di sviluppo degli embrioni nelle uova tratteggiate. È stato osservato che la coltura di cellule dalla retina degli embrioni a ED8 (34° stadio) produce solo il 35% di fotorecettori, con il restante 65% sono fatti di altri neuroni4. Qualunque sia la logistica applicata per ottenere le uova tratteggiate, è necessario mettere a punto la temperatura e il tempo di incubazione, e esaminare attentamente gli embrioni rispetto alle immagini di riferimento di tutte le fasi disviluppo 42,45.
Originariamente, il sistema di coltura arricchito con coni è stato sviluppato utilizzando il ceppo di LivornoBianco 4. Il colore bianco delle uova di quella varietà non è particolarmente apprezzato in Francia, quindi abbiamo usato un ceppo di pollo che produce uova marroni. Abbiamo utilizzato il ceppo I 657, che è fatto incrociando I 66 galli con galliNE JA575. Siamo stati in grado di riprodurre le caratteristiche delle culture originali. Ciò dimostra che il background genetico del pollo non è fondamentale per ottenere colture arricchite di coni.
Non abbiamo testato l'effetto della rimozione individuale degli integratori nel mezzo di coltura, ma abbiamo osservato che l'insulina svolge un ruolo critico in conformità con l'effetto dell'insulina sulla sopravvivenza dei coni nel topo rd1, un modello di RP autosomico recessivo46. La triiodotironina (T3) può anche partecipare alla differenziazione delle cellule precursori retiniche dell'embrione di pollo in coni in base al ruolo del recettore dell'ormone tiroideo nel destino cellulare retinico durante losviluppo 40. Di conseguenza, il sistema di coltura arricchito di coni non può essere utilizzato per identificare l'insulina medianteclonazione di espressione 46.
Il sistema di coltura arricchito di coni si basa sulla coltura dei neuroni primari ed è molto più appropriato per i metodi che si basano sull'uso di cellule immortalate come linea cellulare 661W24,25,26.
Il metodo qui descritto può essere modificato eseguendo una precedente elettroporazione con DNA plasmide20. Prima di preparare la sospensione cellulare retinica, l'intera retina viene posta nella camera di un elettroporatore su misura con 120 μL di 0,5 μg/μL di DNA plasmide in 10 mM Tris-HCl pH 8.0, 1 mM EDTA. Cinque impulsi da 15 V per 50 ms ciascuno sono applicati separati da 950 ms intervallo22. I tentativi di fornire RNA interferente (RNAi) utilizzando i retrovirus RCAS (Avian Splice) competenti per la replicazione in colture arricchite di coni non hannoavuto successo 47. Ciò è certamente dovuto al fatto che in assenza di siero e in colture a bassa densità, le cellule precursori retiniche non sono replicative, un requisito per la propagazione dei retrovirus.
Abbiamo sviluppato il sistema di coltura arricchito di coni per identificare i fattori trotrofici che promuovono la sopravvivenza del cono usando laclonazione dell'espressione 19. Per renderlo fattibile, abbiamo eseguito un primo passo di screening ad alto contenuto utilizzando mezzo condizionato da pool di 100 cloni. Anche se i cDNA della libreria sono espressi sotto il controllo di un forte promotore CMV dopo la trasfezione delle cellule COS-1, non offre una garanzia che tutte le proteine codificate dai singoli cDNA raggiungano una concentrazione sufficiente per essere valutate positive dal saggio di vitalità. Si tratta di una limitazione importante. In questo senso, qualsiasi screening non è realmente esaustivo. Inoltre, anche se le proteine membranose non sono state rimosse dal mezzo condizionato, la configurazione del saggio è sfavorevole all'identificazione di fattori non diffusivi. Un'alternativa sarebbe quello di schermare i singoli cloni dopo aver ottenuto la sequenza dei cDNA al fine di evitare duplicazioni nel saggiare molte volte la stessa proteina candidata. Questo è stato avviato sequenziando le librerie cDNA retiniche che abbiamo usato43. Sebbene razionale, questo approccio ha anche i suoi limiti. L'analisi bioinformatica delle sequenze cDNA imporrà irresistibilmente, oltre alla riduzione della ridondanza, la priorità dello screening di alcuni cloni in base alla conoscenza. Ciò non sarà dannoso se, infine, l'intera libreria verrà schermata anche se il tempo necessario per farlo sarà significativamente allungato. Ma invariabilmente, l'identità della sequenza influenzerà il nostro modo di guardare i risultati. Ciò non sarà neutro, poiché l'interpretazione della sequenza sarà naturalmente in concorrenza con i dati sperimentali.
L'identificazione di EdCVF dimostra anche che lo screening ad alto contenuto comporta limitazioni tecniche. Fin dalla prima tornata di screening, la Commissione ha identificato due pool ad alta attività(figura complementare 1). Il pool 0073 ha portato alla riuscita identificazione di EdCVF, mentre il pool 0080 non ha condotto a tale scoperta. Non abbiamo risolto il problema che potrebbe derivare dalla perdita del clone attivo durante la preparazione dei sotto-pool. In alternativa, non è escluso, anche se non statisticamente favorevole, che tra i cDNA del pool 0080, due proteine agivano sinergisticamente e la loro attività non poteva essere osservata come singoli cloni.
L'identificazione di molecole che proteggono i coni attraverso lo screening di piccole molecole è una futura applicazione del sistema di coltura arricchito di coni. Tali molecole saranno preziose per il trattamento di patologie retiniche per le quali la terapia genica non è l'approccio più appropriato come degenerazione maculare legata all'età.
Divulgazioni
TL, J-AS e VF detengono un brevetto sull'uso di EdCVF per il trattamento delle degenerazioni retiniche [WO2009071659 (A1). 12 giugno 2007].
Riconoscimenti
Gli autori ringraziano Jacques Bellalou, Lorelei Fournier, Emmanuelle Clérin, Frédéric Blond e l'entreprise agricole à responsabilité limitée (EARL Morizeau Dangers, Francia) per il loro prezioso aiuto. Questo lavoro è stato sostenuto da Inserm, Università della Sorbona, Agence Nationale pour la Recherche (ANR, Labex Lifesenses), Foundation Fighting Blindness (USA) e IHU FOReSIGHT [ANR-18-IAHU-0001] supportati da fondi statali francesi gestiti dall'ANR nell'ambito del programma Investissements d'Avenir.
Materiali
| Name | Company | Catalog Number | Comments |
| 96 black plates (Clear Button with lid Tissue culture treated) | Corning | 3603 | |
| Calcein AM | Thermo-Fisher scientific | C1430 | |
| CCD Camera | Photometrics | CoolSnap FX HQ | |
| CDPC (Cytidine 5′-diphosphocholine sodium salt dihydrate) | Sigma-Aldrich | C0256 | |
| CO2 Independant | Thermo-Fisher scientific | 18045-054 | |
| Curved forceps | Dutscher | 005093 | |
| DMEM Media | Thermo-Fisher scientific | 41966-029 | |
| DNAse | Sigma-Aldrich | D4263 | |
| Eggs incubator | FarmLine | M08 01 3100 | |
| Ethidium Homodimer | Thermo-Fisher scientific | E1169 | |
| Fœtal bovine serum | Thermo-Fisher scientific | 10270-098 | |
| Gentamycin | Thermo-Fisher scientific | 15710-049 | |
| Hydrocortisone | Sigma-Aldrich | H0880 | |
| ITS (insulin Transferine selenium) | Sigma-Aldrich | I1884 | |
| large straight pliers | Dutscher | 005074 | |
| Linoleic acid | Sigma-Aldrich | L8384 | |
| M199 medium | Thermo-Fisher scientific | 31150-022 | |
| Metamorph software | Metamorph | ||
| Microscope | NIKON | Eclipse TE2000 | |
| Motorized stage | Martzauzer | Mutlicontrol 2000 | |
| Optical filter switch | Shutter Instrument company | Lambda 10-2 | |
| PBS 1X | Thermo-Fisher scientific | 14190-086 | |
| Poly-L-lysine | Sigma-Aldrich | P6282 | |
| Progesterone | Sigma-Aldrich | P7556 | |
| Pursept A express | Fisher scientific | 11814110 | |
| Putriscine | Sigma-Aldrich | P5780 | |
| Sodium pyruvate | Sigma-Aldrich | S8636 | |
| straight forceps | Dutscher | 005092 | |
| Taurine | Sigma-Aldrich | T8691 | |
| Triiodothyronine | Sigma-Aldrich | T6397 | |
| Trypan blue | Thermo-Fisher scientific | 15250-061 | |
| Trypsine 0.25 % | Thermo-Fisher scientific | 25200-056 |
Riferimenti
- Brownstein, C. D. . The Implicit Mind. , (2019).
- Heesy, C. P., Hall, M. I. The nocturnal bottleneck and the evolution of mammalian vision. Brain, Behavior and Evolution. 75 (3), 195-203 (2010).
- Borges, R., et al. Adaptive genomic evolution of opsins reveals that early mammals flourished in nocturnal environments. BMC Genomics. 19 (1), 121 (2018).
- Adler, R., Hatlee, M. Plasticity and differentiation of embryonic retinal cells after terminal mitosis. Science. 243 (4889), 391-393 (1989).
- Fintz, A. C., et al. Partial characterization of retina-derived cone neuroprotection in two culture models of photoreceptor degeneration. Investigative Ophthalmology & Visual Science. 44 (2), 818-825 (2003).
- Roska, B., Sahel, J. A. Restoring vision. Nature. 557 (7705), 359-367 (2018).
- Duncan, J. L., et al. Inherited Retinal Degenerations: Current Landscape and Knowledge Gaps. Translational Vision Science & Technology. 7 (4), 6 (2018).
- Russell, S., et al. Efficacy and safety of voretigene neparvovec (AAV2-hRPE65v2) in patients with RPE65-mediated inherited retinal dystrophy: a randomised, controlled, open-label, phase 3 trial. Lancet. 390 (10097), 849-860 (2017).
- RetNet. . Retinal Information Network. , (2020).
- Doonan, F., Donovan, M., Cotter, T. G. Activation of multiple pathways during photoreceptor apoptosis in the rd mouse. Investigative Ophthalmology & Visual Science. 46 (10), 3530-3538 (2005).
- Comitato, A., et al. Dominant and recessive mutations in rhodopsin activate different cell death pathways. Human Molecular Genetics. 25 (13), 2801-2812 (2016).
- Cronin, T., Leveillard, T., Sahel, J. A. Retinal degenerations: from cell signaling to cell therapy; pre-clinical and clinical issues. Current Gene Therapy. 7 (2), 121-129 (2007).
- Baumgartner, W. A., Baumgartner, A. M. Accounting for disagreements on average cone loss rates in retinitis pigmentosa with a new kinetic model: Its relevance for clinical trials. Medical Hypotheses. 89, 107-114 (2016).
- Leveillard, T., Sahel, J. A. Rod-derived cone viability factor for treating blinding diseases: from clinic to redox signaling. Science Translational Medicine. 2 (26), (2010).
- Leveillard, T., Sahel, J. A. Metabolic and redox signaling in the retina. Cellular and Molecular Life Sciences. 74 (20), 3649-3665 (2017).
- Clerin, E., Marussig, M., Sahel, J. A., Leveillard, T. Metabolic and Redox Signaling of the Nucleoredoxin-Like-1 Gene for the Treatment of Genetic Retinal Diseases. International Journal of Molecular Sciences. 21 (5), (2020).
- Fridlich, R., et al. The Thioredoxin-like Protein Rod-derived Cone Viability Factor (RdCVFL) Interacts with TAU and Inhibits Its Phosphorylation in the Retina. Molecular & Cellular Proteomics. 8 (6), 1206-1218 (2009).
- Mei, X., et al. The Thioredoxin Encoded by the Rod-Derived Cone Viability Factor Gene Protects Cone Photoreceptors Against Oxidative Stress. Antioxidants, Redox Signaling. 24 (16), 909-923 (2016).
- Leveillard, T., et al. Identification and characterization of rod-derived cone viability factor. Nature Genetics. 36 (7), 755-759 (2004).
- Vergara, M. N., Gutierrez, C., Canto-Soler, M. V. Efficient Gene Transfer in Chick Retinas for Primary Cell Culture Studies: An Ex-ovo Electroporation Approach. Journal of Visualized Experiments. (105), e52002 (2015).
- Fridlich, R., et al. The thioredoxin-like protein rod-derived cone viability factor (RdCVFL) interacts with TAU and inhibits its phosphorylation in the retina. Molecular & Cellular Proteomics. 8 (6), 1206-1218 (2009).
- Ait-Ali, N., et al. Rod-derived cone viability factor promotes cone survival by stimulating aerobic glycolysis. Cell. 161 (4), 817-832 (2015).
- Mei, X., et al. The Thioredoxin Encoded by the Rod-Derived Cone Viability Factor Gene Protects Cone Photoreceptors Against Oxidative Stress. Antioxidants & Redox Signaling. 24 (16), 909-923 (2016).
- Tan, E., et al. Expression of cone-photoreceptor-specific antigens in a cell line derived from retinal tumors in transgenic mice. Investigative Ophthalmology & Visual Science. 45 (3), 764-768 (2004).
- Wang, X. W., Tan, B. Z., Sun, M., Ho, B., Ding, J. L. Thioredoxin-like 6 protects retinal cell line from photooxidative damage by upregulating NF-kappaB activity. Free Radical Biology and Medicine. 45 (3), 336-344 (2008).
- Perron, N. R., Beeson, C., Rohrer, B. Early alterations in mitochondrial reserve capacity; a means to predict subsequent photoreceptor cell death. Journal of Bioenergetics and Biomembranes. 45 (1-2), 101-109 (2013).
- Krishnamoorthy, R. R., Clark, A. F., Daudt, D., Vishwanatha, J. K., Yorio, T. A forensic path to RGC-5 cell line identification: lessons learned. Investigative Ophthalmology & Visual Science. 54 (8), 5712-5719 (2013).
- Al-Ubaidi, M. R. RGC-5: are they really 661W? The saga continues. Experimental Eye Research. 119, 115 (2014).
- Clerin, E., et al. Vibratome sectioning mouse retina to prepare photoreceptor cultures. Journal of Visualized Experiments. (94), (2014).
- Hendrickson, A., Hicks, D. Distribution and density of medium- and short-wavelength selective cones in the domestic pig retina. Experimental Eye Research. 74 (4), 435-444 (2002).
- Bobu, C., Craft, C. M., Masson-Pevet, M., Hicks, D. Photoreceptor organization and rhythmic phagocytosis in the nile rat Arvicanthis ansorgei: a novel diurnal rodent model for the study of cone pathophysiology. Investigative Ophthalmology & Visual Science. 47 (7), 3109-3118 (2006).
- Saidi, T., Mbarek, S., Chaouacha-Chekir, R. B., Hicks, D. Diurnal rodents as animal models of human central vision: characterisation of the retina of the sand rat Psammomys obsesus. Graefes Arch Clin Exp Ophthalmol. 249 (7), 1029-1037 (2011).
- Traverso, V., Kinkl, N., Grimm, L., Sahel, J., Hicks, D. Basic fibroblast and epidermal growth factors stimulate survival in adult porcine photoreceptor cell cultures. Investigative Ophthalmology & Visual Science. 44 (10), 4550-4558 (2003).
- Balse, E., et al. Purification of mammalian cone photoreceptors by lectin panning and the enhancement of their survival in glia-conditioned medium. Investigative Ophthalmology & Visual Science. 46 (1), 367-374 (2005).
- Gagliardi, G., et al. Characterization and Transplantation of CD73-Positive Photoreceptors Isolated from Human iPSC-Derived Retinal Organoids. Stem Cell Reports. 11 (3), 665-680 (2018).
- Gagliardi, G., Ben M'Barek, K., Goureau, O. Photoreceptor cell replacement in macular degeneration and retinitis pigmentosa: A pluripotent stem cell-based approach. Progress in Retinal and Eye Research. 71, 1-25 (2019).
- Mears, A. J., et al. Nrl is required for rod photoreceptor development. Nature Genetics. 29 (4), 447-452 (2001).
- Swaroop, A., Kim, D., Forrest, D. Transcriptional regulation of photoreceptor development and homeostasis in the mammalian retina. Nature Reviews Neuroscience. 11 (8), 563-576 (2010).
- Kallman, A., et al. Investigating cone photoreceptor development using patient-derived NRL null retinal organoids. Nature Communications. 3 (1), 82 (2020).
- Eldred, K. C., et al. Thyroid hormone signaling specifies cone subtypes in human retinal organoids. Science. 362 (6411), (2018).
- Kole, C., et al. Otx2-Genetically Modified Retinal Pigment Epithelial Cells Rescue Photoreceptors after Transplantation. Molecular Therapy. 26 (1), 219-237 (2018).
- Hamburger, V., Hamilton, H. L. A series of normal stages in the development of the chick embryo. Journal of Morphology. 88 (1), 49-92 (1951).
- Kole, C., et al. Identification of an Alternative Splicing Product of the Otx2 Gene Expressed in the Neural Retina and Retinal Pigmented Epithelial Cells. PLoS One. 11 (3), 0150758 (2016).
- Shannon, M., Hamilton, A. T., Gordon, L., Branscomb, E., Stubbs, L. Differential expansion of zinc-finger transcription factor loci in homologous human and mouse gene clusters. Genome Research. 13 (6), 1097-1110 (2003).
- Stern, C. D., Holland, P. W. . Essential developmental biology: a practical approach. , (1993).
- Punzo, C., Kornacker, K., Cepko, C. L. Stimulation of the insulin/mTOR pathway delays cone death in a mouse model of retinitis pigmentosa. Nature Neuroscience. 12 (1), 44-52 (2009).
- Harpavat, S., Cepko, C. L. RCAS-RNAi: a loss-of-function method for the developing chick retina. BMC Developmental Biology. 6, 2 (2006).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati