
Method Article
Estimulación y análisis de la neurogénesis adulta en el cerebro central de Drosophila
En este artículo
Resumen
Este artículo proporciona protocolos detallados para infligir lesión cerebral traumática penetrante (PTBI) a Drosophila adulta y examinar la neurogénesis resultante.
Resumen
Los mecanismos moleculares y celulares que subyacen a la neurogénesis en respuesta a una enfermedad o lesión no se comprenden bien. Sin embargo, comprender estos mecanismos es crucial para desarrollar terapias regenerativas neurales. Drosophila melanogaster es un modelo líder para estudios de desarrollo neuronal, pero históricamente no se ha explotado para investigar la regeneración cerebral adulta. Esto se debe principalmente a que el cerebro adulto exhibe una actividad mitótica muy baja. Sin embargo, la lesión cerebral traumática penetrante (PTBI) en el cerebro central de Drosophila adulta desencadena la generación de nuevas neuronas y nuevas glías. Las poderosas herramientas genéticas disponibles en Drosophila combinadas con el protocolo de lesión simple pero riguroso descrito aquí ahora hacen que el cerebro adulto de Drosophila sea un modelo robusto para la investigación de la regeneración neuronal. Aquí se proporcionan instrucciones detalladas para (1) lesiones penetrantes en el cerebro central adulto y (2) disección, inmunohistoquímica e imágenes posteriores a la lesión. Estos protocolos producen resultados altamente reproducibles y facilitarán estudios adicionales para diseccionar los mecanismos subyacentes a la regeneración neuronal.
Introducción
El daño al cerebro y al sistema nervioso es una de las principales causas de muerte y discapacidad en todo el mundo. Aproximadamente 1.5 millones de estadounidenses sufren lesiones cerebrales traumáticas (LCT) cada año1, mientras que se estima que 6 millones de personas solo en los Estados Unidos sufren de enfermedades neurodegenerativas, como la enfermedad de Parkinson y la enfermedad de Alzheimer2. Tanto la enfermedad como la lesión cerebral pueden causar degeneración neuronal, lo que lleva a defectos sensoriales, cognitivos y motores3. Desarrollar estrategias terapéuticas para la reparación del cerebro humano ha sido difícil debido a la compleja fisiología del cerebro. Los organismos modelo como Drosophila melanogaster proporcionan un sistema simple para identificar los mecanismos fundamentales subyacentes a la neurodegeneración y las posibles dianas terapéuticas4.
La mosca de la fruta Drosophila melanogaster ha sido un poderoso organismo modelo durante más de un siglo, avanzando en los campos de la genética, la biología del desarrollo y la neurociencia5,6. El cerebro de Drosophila comprende solo ~ 90,000 neuronas7, un millón de veces menos que el cerebro humano promedio8, sin embargo, tienen muchas similitudes. Tanto los cerebros humanos como los de mosca utilizan los neurotransmisores GABA, glutamato, acetilcolina y las aminas biogénicas dopamina y serotonina9. Drosophila y las neuronas humanas también funcionan de manera similar, con una arquitectura sináptica compartida y tipos de células neuronales análogas10. El menor tamaño del cerebro de Drosophila y la disponibilidad de técnicas genéticas avanzadas, en combinación con la conservación de los mecanismos moleculares, celulares y fisiológicos entre Drosophila y los mamíferos, permite a los investigadores de Drosophila hacer preguntas que son poco prácticas o difíciles de responder en modelos de mamíferos.
Nuestra comprensión actual de la neurogénesis adulta en Drosophila, tanto durante la homeostasis como después de una lesión, sigue siendo limitada. Se sabe más sobre la neurogénesis durante el desarrollo normal. Por ejemplo, las neuronas y la glía se crean durante el desarrollo a partir de células precursoras, llamadas neuroblastos10,11. Se han distinguido al menos tres tipos diferentes de neuroblastos en el cerebro central. Tanto los neuroblastos de linaje tipo I como los tipo II salen del ciclo celular ~20-30 h después de la formación del pupario12. Por el contrario, los neuroblastos del cuerpo de los hongos son los últimos en terminar la división celular y lo hacen a través de la apoptosis dependiente de Reaper ~ 85-90 h después de la formación del pupario13. Después de la eclosión, el cerebro adulto de Drosophila tiene pocas células en división (~1 célula/cerebro), predominantemente glia14. Los lóbulos ópticos adultos poseen neuroblastos de ciclo lento capaces de neurogénesis15, mientras que el cerebro central adulto no tiene neuroblastos conocidos. La escasez de progenitores neurales y la proliferación celular limitada se asemejan mucho a la situación en el cerebro de los mamíferos adultos, lo que subraya la relevancia potencial de los mecanismos de neurogénesis adulta en Drosophila para los humanos.
El descubrimiento de bajos niveles de neurogénesis adulta en los lóbulos ópticos adultos de Drosophila después de una lesión15 llevó a la hipótesis de que el cerebro central de Drosophila adulto también podría ser capaz de neurogénesis adulta16. Este protocolo describe la creación de un modelo riguroso y reproducible de lesión cerebral central en Drosophila adulta que se puede utilizar para investigar la neurogénesis en el cerebro central adulto. Dadas las similitudes entre la arquitectura y la función del cerebro humano y Drosophila , estos descubrimientos podrían conducir a la identificación de objetivos críticos para la neurogénesis terapéutica en cerebros humanos lesionados y enfermos.
Protocolo
Este protocolo sigue las pautas de cuidado animal de UW-Madison.
1. Generación de Drosophila adulta para PTBI
- Para la cruz estándar, coloque 20 virgin y[1] w[1]; UAS-mCD8-GFP;; OK107-GAL417 hembras adultas y 10 y[1] w[1]17 moscas macho adultas juntas en viales (ver Tabla de Materiales) que contienen alimentos. Para tener un gran número de descendientes sincrónicos, configure 10-20 cruces al mismo tiempo. El cruce estándar da lugar a la progenie F1 del genotipo: y[1] w[1]; UAS-mCD8-GFP/+;; OK107-GAL4/+.
- Coloque los viales a 25 °C para el apareamiento y la puesta de huevos. Para maximizar la progenie, transfiera a los padres a nuevos viales cada 2-4 días, manteniendo los viales anteriores a 25 ° C. Deseche cada vial 18 días después de que los padres fueron colocados por primera vez en él para asegurarse de que no haya progenie F2.
NOTA: Se pueden generar 3 conjuntos de crías ("crías") a partir de cada conjunto de padres. - Verifique después de ~ 10 días, cuando la progenie de F1 comenzará a eclosionar.
- Coloque los viales a 25 °C para el apareamiento y la puesta de huevos. Para maximizar la progenie, transfiera a los padres a nuevos viales cada 2-4 días, manteniendo los viales anteriores a 25 ° C. Deseche cada vial 18 días después de que los padres fueron colocados por primera vez en él para asegurarse de que no haya progenie F2.
- Para los estudios de linaje, utilizar machos F1 perma-gemelos15 del genotipo: w; FRT40A, UAS-CD2-RFP, UAS-GFP-Mir/FRT40A, UAS-CD8-GFP, UAS-CD2-Mir; tub-GAL80ts/act-GAL4 UAS-flp. Para mayor coherencia, haga esta cruz en la misma dirección cada vez.
- Para generar machos perma-gemelos, coloque 20 w virgen; FRT40A, UAS-CD2-RFP, UAS-GFP-Mir; tub-GAL80ts/TM6B hembras y 10 w; FRT40A, UAS-CD8-GFP, UAS-CD2-Mir; act-GAL4 UAS-flp/TM6B15 machos adultos juntos en viales que contienen alimentos.
- Coloque los viales a 17 °C para el apareamiento y la puesta de huevos. Transfiera a los padres a viales de alimentos nuevos cada 7 días, manteniendo todos los viales a 17 °C. Deseche cada vial 35 días después de que los padres se colocaron por primera vez en él para asegurarse de que no haya progenie F2.
- Verifique después de ~ 21 días, cuando la progenie de F1 comenzará a eclosionar.
2. Lesión cerebral traumática penetrante (PTBI; Figura 1)
- Clasifica las moscas F1 recién eclosionadas. Seleccione machos jóvenes dentro de las 6 h posteriores a la eclosión. Coloque estos machos en viales limpios que contengan alimentos, con 40 o menos moscas por vial.
NOTA: Esto se logra más fácilmente a media mañana anestesiando moscas en la almohadilla de CO2 e identificando machos adultos que todavía tienen meconio (visible como una mancha verdosa oscura a través de la pared abdominal) en sus intestinos. - Prealimentación con 5-etinil-2'-desoxiuridina (EdU) (ver Tabla de Materiales) durante 6 h antes de la lesión si planea etiquetar las células en división con EdU. Consulte el paso 3 para obtener más información.
- Desinfecte los pines minutien (consulte la Tabla de materiales) durante al menos 5 minutos colocando ~ 100 pines en un tubo de microcentrífuga de 1.5 ml lleno de etanol al 70%.
- Desinfecte la almohadilla de CO2 y un pincel rociando etanol al 70% y seque con un pañuelo limpio y sin pelusa. Una vez que las herramientas estén limpias y secas, transfiera 40 o menos machos F1 clasificados de nuevo a la almohadilla limpia.
- Separe los machos F1 en 2 grupos en la plataforma de mosca. Un grupo servirá como un control de moscas ilesas. El segundo grupo experimental será sometido a PTBI.
- Usando fórceps, saque 4-5 nuevos pines Minutien del tubo de la microcentrífuga y colóquelos cerca del borde de la almohadilla de CO2 . Debajo del visor de disección, elija un pasador Minutien recto con un punto afilado.
NOTA: El uso de un solo pin Minutien para un experimento reducirá la variabilidad. Reutiliza pasadores afilados. Coloque los pasadores dañados o romos en un tubo de microcentrífuga separado de 1,5 ml que contenga 70% de etanol para una eliminación segura. - Si trabaja con moscas de la cruz estándar, encienda la lámpara de diodo emisor de luz (LED) del microscopio estereoscópico equipada con los filtros de excitación y emisión apropiados para la proteína de fluorescencia verde (GFP). Esto permite la excitación a 440-460 nM y permite la visualización de 500-560 nM.
NOTA: Con este juego de lámparas y filtros, los cuerpos celulares del cuerpo del hongo se fluorescen de color verde a través de la cutícula de la cabeza (Figura 1C). Para las moscas perma-gemelas o moscas de otros genotipos, se puede usar un iluminador de luz blanca estándar para el microscopio estereoscópico para visualizar puntos de referencia en la cutícula de la cabeza para apuntar a la lesión en el cuerpo del hongo (Figura 1C). - Use las pinzas para recoger y sostener el pin Minutien seleccionado en una mano (para las personas diestras, esta suele ser la mano derecha) y el pincel en la otra mano (generalmente izquierda). Elija una mosca del grupo experimental y coloque la mosca de tal manera que tenga una vista dorsal de la cápsula de la cabeza con la cabeza de la mosca a la derecha. Coloque el cepillo en la parte anterior del tórax dorsal y empuje suavemente hacia abajo para estabilizar la mosca.
- Apunte la punta del alfiler Minutien a los cuerpos celulares del cuerpo del hongo en el lado derecho de la cabeza y penetre la cápsula de la cabeza. Si usa puntos de referencia, apunte a la cutícula dorsal de la cabeza entre los ocelos y el borde dorsal del ojo (Figura 1).
- Después de completar la lesión, use el pincel para empujar la cabeza suavemente fuera del pasador Minutien.
- Si usa el cerebro para RNA-Seq o qRT-PCR, haga una segunda lesión en el lado izquierdo de la cabeza.
- Repita los pasos 2.8-2.10 para lesionar a todas las moscas del grupo experimental.
- Una vez que todas las moscas hayan sido lesionadas, coloque el control y las moscas lesionadas en viales separados etiquetados que contengan alimentos. Coloque los viales horizontalmente (es decir, de lado) mientras las moscas se recuperan de la anestesia y durante el envejecimiento posterior para evitar que las moscas queden atrapadas en los alimentos.
- Coloque las moscas cruzadas estándar a 25 ° C, y las moscas perma-twin a 30 ° C para envejecer.
- Para moscas de más de 24 h, colóquelas en alimentos limpios cada 1-2 días.
3. Etiquetado EdU
- Preparar un caldo de 10 mM 5-etinil-2'-desoxiuridina (EdU) en dimetilsulfóxido (DMSO). Esto se puede almacenar a -20 ° C durante un máximo de 12 meses.
- Preparar 200 μL de 50 μM de EdU en sacarosa al 10%. Pre-alimentación de moscas con EdU durante 6 h antes de PTBI.
- Coloque 200 μL de EdU de 50 μM en sacarosa al 10% sobre un papel de filtro redondo de grado 3 de 23 mm (consulte la Tabla de materiales) en un vial vacío.
- Coloque las moscas en el vial. Luego selle el vial con un tapón de algodón.
- Coloque el vial horizontalmente en una incubadora humidificada a 25 °C durante 6 h.
- Lleve a cabo PTBI como se describe en los pasos 2.3-2.10.
- Coloque las moscas de nuevo en el vial que contiene EdU. Selle el vial con un tapón de algodón.
- Coloque el vial horizontalmente en una incubadora humidificada a 25 °C durante un máximo de 24 h. Siga uno de los pasos que se describen a continuación (paso 3.8.1-3.8.3).
- Diseccionar y reparar cerebros como se describe en el paso 4 usando fijador, tampón de lavado y tampón de bloqueo sin azida y con la reacción de detección de EdU llevada a cabo antes de la tinción de anticuerpos.
- Transfiera las moscas a un vial limpio que contenga un nuevo papel de filtro y 200 μL de 50 μM EdU en sacarosa al 10%. Coloque el vial horizontalmente en una incubadora de 25 °C. Repetir cada 24 h durante la duración del etiquetado. Luego, diseccione y arregle los cerebros como se describe en el paso 4 usando tampones sin azida con la reacción de detección de EdU llevada a cabo antes de la tinción de anticuerpos.
- Para etiquetar la persecución de pulsos con EdU, alimente EdU durante el período de pulso, transfiera las moscas cada 24 h a un vial limpio que contenga un nuevo papel de filtro y 200 μL de 50 μM en sacarosa al 10%. Después del período de pulso (por ejemplo, 4 días), transfiera las moscas a un vial que contenga alimentos estándar de Drosophila. Coloque el vial de lado en una incubadora de 25 °C durante 3 días adicionales. Luego, diseccione y arregle los cerebros como se describe en el paso 4 usando tampones sin azida con la reacción de detección de EdU llevada a cabo antes de la tinción de anticuerpos.
4. Disección, inmunohistoquímica y montaje
- Preparar tubos de microcentrífuga de 1,5 mL con 100 μL de fijador: formaldehído al 4% en PEM (100 mM piperazina-N,N'-bis(ácido 2-etanosulfónico) [PIPES], 1 mM EGTA, 1 mM MgSO4, pH 7.0) (ver Tabla de Materiales) y colocar sobre hielo.
NOTA: Hasta 20 cerebros de un solo genotipo y condición pueden ser procesados en un solo tubo. - Prepare una placa de disección con una pequeña piscina (~100 μL) de etanol al 70% y tres pequeñas piscinas de solución salina tamponada con fosfato (PBS; 100 mM de K2HPO4, 140 mM de NaCl, pH 7.0).
- Anestesiar ~ 10 moscas de control o experimentales en una almohadilla de CO2 que ha sido desinfectada con etanol al 70%.
- Separe la cabeza del tronco de cada mosca con un bisturí.
- Recoja las cabezas con un pincel humedecido en etanol al 70% y coloque las cabezas durante 2-5 minutos en la piscina de etanol en la placa de disección.
NOTA: Esto deshidrata ligeramente los cerebros y los hace más fáciles de diseccionar lejos de la cutícula de la cabeza. - Transfiera las cabezas a un grupo de ~ 100 μL de PBS y diseccione los cerebros, moviendo cada cerebro a un grupo limpio de ~ 100 μL de PBS. Realice esto usando dos pares de fórceps Watchmakers para abrir la parte posterior de la cutícula de la cabeza y sosteniendo la cutícula con un par de fórceps mientras extrae suavemente el cerebro de la cutícula usando la punta cerrada del segundo par de fórceps.
- Transfiera los cerebros disecados a un tubo de microcentrífuga que contiene 100 μL de la solución de fijación utilizando un pipettor P200 equipado con una punta de plástico que ha sido cortada y biselada para permitir la entrada de los cerebros.
- Fijar durante 20-25 min a temperatura ambiente. Retire con cuidado la solución con un P200 o una pipeta de vidrio.
- Lave los cerebros fijos cuatro veces con 1 ml de 'PT' (PBS más 0.1% Triton X-100), permitiendo que los cerebros se asienten durante varios minutos entre cada lavado.
NOTA: Si los cerebros no se asientan rápidamente entre los lavados, es probable que todavía tengan cuerpo gordo y / o tráquea adherida. - Bloquee las muestras en 1 ml de PBS más Tritón X-100 al 0,1% y albúmina sérica bovina (PBT) al 2% durante ~1 h a temperatura ambiente.
- Retire la solución de bloqueo e incube las muestras con 100 μL de solución primaria de anticuerpos durante la noche a 4 °C en PBT. Los principales anticuerpos utilizados en este estudio son anti-PH3 de conejo (1:500) y anti-Fasiclin II de ratón (1:20) (ver Tabla de Materiales).
- Lave las muestras cinco veces con 1 ml de PT, permitiendo que los cerebros se asienten durante varios minutos entre cada lavado.
- Retire el lavado final e incube en 100 μL de solución secundaria de anticuerpos durante la noche a 4 °C. Los anticuerpos secundarios utilizados aquí son anti-conejo 568 (1:400) y anti-ratón Cy5 (1:100) (ver Tabla de Materiales).
- Lave las muestras cinco veces con 1 ml de PT, permitiendo que los cerebros se asienten durante varios minutos entre cada lavado. Durante el lavado final, agregue 4',6-Diamidino-2-Phenylindole, Dihydrochloride (DAPI; 1:100 de una solución de 10 μM) durante 10 min para teñir los núcleos. Retire el lavado, dejando 50-100 μL en cada tubo.
- Prepare portaobjetos de microscopio colocando una sola etiqueta de refuerzo autoadhesiva en el centro de cada portaobjetos y una gota de 50 μL de medios de montaje antidesvanecimiento en el centro de cada etiqueta (consulte la Tabla de materiales). La etiqueta de refuerzo sostiene el latícula ligeramente por encima de las diapositivas y evita que los cerebros montados se aplanen demasiado.
- Transfiera cuidadosamente los cerebros de cada tubo de microcentrífuga a un portaobjetos preparado con el menor búfer de lavado posible utilizando un P200 equipado con una punta de plástico cortada y biselada. Se pueden montar hasta 10 cerebros en una sola diapositiva.
- Use fórceps para reposicionar suavemente los cerebros antes de colocar un cubrehojas en cada diapositiva y sellar la diapositiva con esmalte de uñas. Oriente los cerebros con el lado posterior hacia arriba o el lado anterior hacia arriba, pero no debe tocarse entre sí.
NOTA: Debido a que es difícil obtener imágenes confocales de alta calidad a través de todo un cerebro, algunos cerebros deben montarse en cada orientación. - Guarde los portaobjetos preparados planos y en la oscuridad a 4 °C hasta la toma de imágenes. Para un almacenamiento a más largo plazo (hasta 1 año), las diapositivas pueden almacenarse a -20 °C.
5. Imágenes confocales
NOTA: Imagine cerebros utilizando un microscopio confocal de escaneo láser con láser láser y cubos de filtro de emisión apropiados para DAPI y los anticuerpos secundarios fluorescentes (es decir, 405 nm, 488 nm y 568 nm, 633 nm, respectivamente).
- Encienda la alimentación del microscopio, los láseres, el controlador y la computadora.
- Abra el software adquisición.
- En el modo Adquisición, seleccione hasta cuatro canales y establezca el parámetro para el escaneo secuencial de los canales deseados. Aumente la potencia del láser para cada canal al 5-10%.
- Coloque un portaobjetos en el microscopio.
- Elija un cerebro para ser fotografiado usando el accesorio de epifluorescencia (ver Tabla de Materiales).
- En el modo Adquisición, seleccione 1024 x 1024 píxeles como dimensión del fotograma.
- En el modo Adquisición, seleccione la opción pila Z, indique que la pila Z debe tomarse en secciones de 2 μm y enfoque a través de la muestra, seleccionando los planos focales superior e inferior que se van a visualizar.
- Recopile imágenes de pila Z de cerebros enteros utilizando un objetivo 20x y regiones cerebrales específicas como el cuerpo del hongo usando un objetivo 60x.
6. Análisis de datos
- Cuantificar las células proliferantes y el número de clones perma-gemelos manualmente y/o utilizando el software de análisis de imágenes (ver Tabla de Materiales). Cuando utilice software, seleccione regiones de interés (ROI) con áreas de al menos 10 μm.
Resultados
PTBI estimula la proliferación celular
Para determinar el grado de neurogénesis después de un PTBI cerebral central, se midió la respuesta proliferativa en varones adultos jóvenes recogidos y lesionados dentro de las 6 h de la eclosión. Se observó un aumento significativo en la proliferación 24 h después de la lesión utilizando antifosfofistona 3 (PH3), un marcador para las células sometidas activamente a mitosis. Aproximadamente 3 células PH3+ en cerebros centrales de control y 11 células PH3+ en los cerebros centrales lesionados se observan 24 h después de PTBI (Figura 2A-D). La mayoría de las células en división se encuentran cerca del sitio de la lesión. Se utilizó un segundo ensayo para la división celular para cuantificar la proliferación celular acumulada de una sola lesión y para evaluar el grado en que las células recién creadas sobrevivieron. La 5-etinil-2'-desoxiuridina (EdU) es un análogo de la timidina que se puede incorporar al ADN recién sintetizado y etiquetar permanentemente las células que se han sometido a la síntesis de ADN. A las moscas se les dio un pulso de EdU de 4 días, seguido de una persecución de 3 días. Esto reveló que las células marcadas eran viables y sobrevivieron al menos 3 días después de la proliferación. A los 7 días, había un promedio de 2 células EdU+ en los cerebros centrales de control y un promedio de 11 células EdU+ en los cerebros centrales lesionados, respectivamente (Figura 2E). Esto es similar a los resultados obtenidos 24 h después de la lesión utilizando el anticuerpo PH3. Cuando la proliferación celular se mide a los 14 días, los controles no lesionados promediaron 1 célula EdU + por cerebro central, mientras que los cerebros lesionados promediaron 29 células EdU + (Figura 2E), lo que demuestra que la proliferación celular continúa al menos hasta la segunda semana después de un PTBI.
La proliferación celular depende de la edad
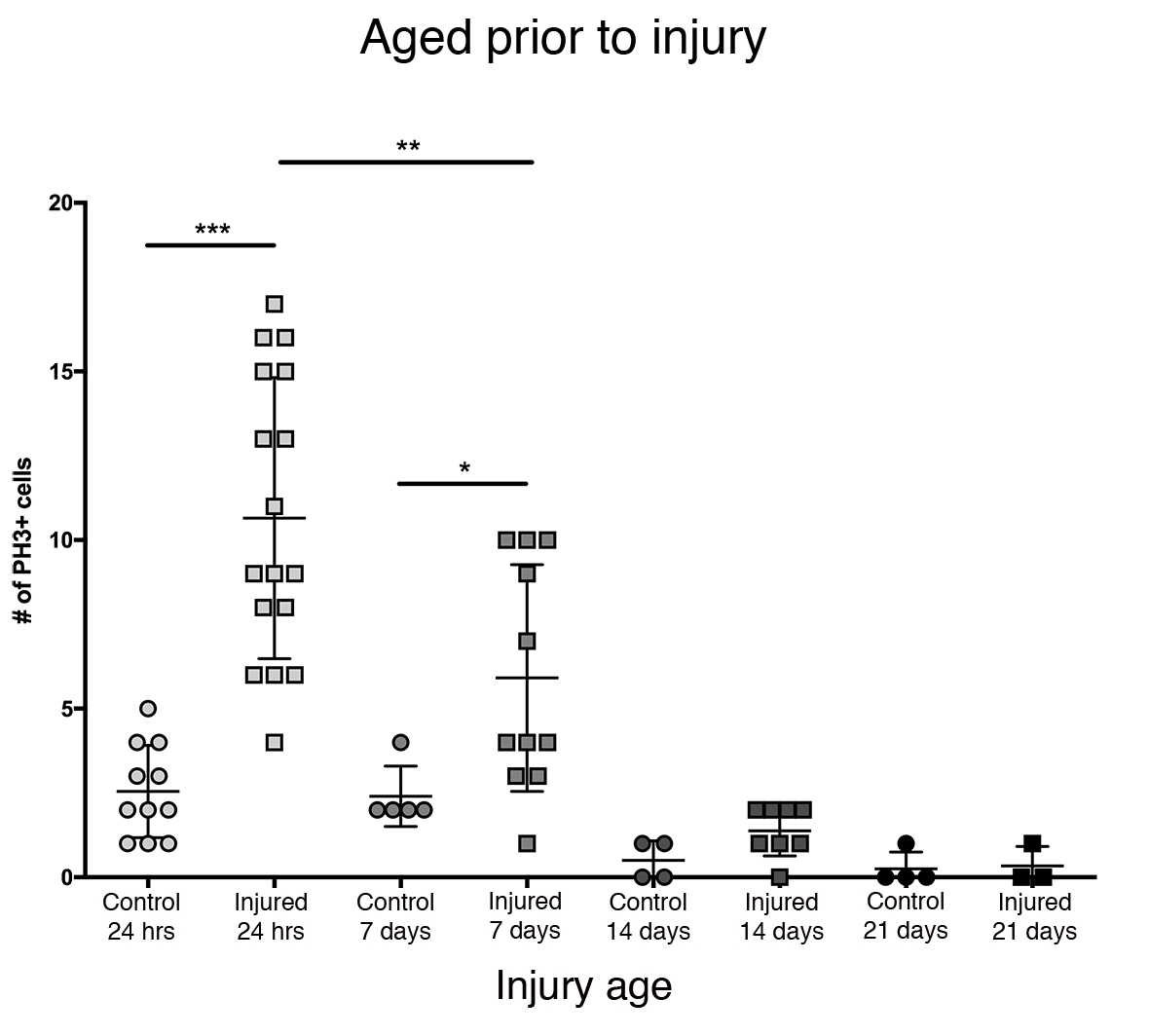
La mayor respuesta proliferativa en el cerebro central se observó dentro de las primeras 24 h después de la eclosión (Figura 3). A los 7 días después de la eclosión, una lesión penetrante todavía causa un aumento significativo en la proliferación, con un promedio de 6 células PH3 + por cerebro central. Aún así, a los 14 días después de la eclosión, la capacidad de las células para dividirse después de PTBI disminuye significativamente a 1 célula en división, similar a la de los cerebros de control (Figura 3). Por lo tanto, el potencial de proliferación celular después de PTBI depende de la edad.
Las neuronas recién creadas pueden proyectarse para corregir las áreas objetivo
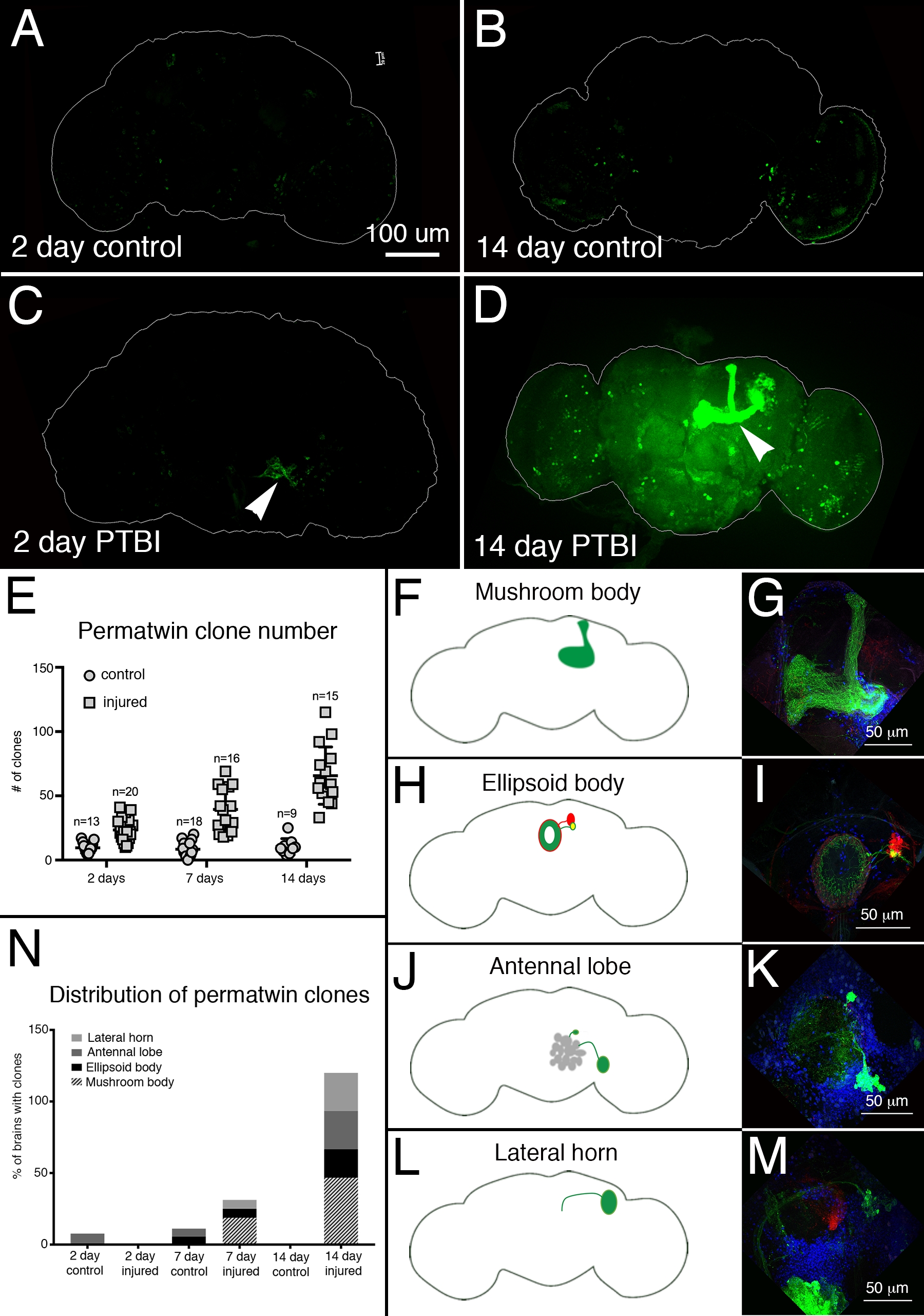
Para evaluar la regeneración neuronal post-PTBI se utilizó el sistema de etiquetado perma-twin15. El rastreo del linaje perma-gemelo etiqueta permanentemente las células en división y su progenie con una proteína fluorescente verde (GFP) o una proteína fluorescente roja (RFP)15. Se detectaron más clones de perma-gemelos en muestras lesionadas, a los 2 días y 2 semanas, que en los controles (Figura 4A-E). En particular, hubo nuevas neuronas del cuerpo de hongos en ~ 50% de los cerebros de PTBI 2 semanas después de la lesión (Figura 4N). Estas nuevas neuronas proyectaron sus dendritas apropiadamente al cáliz del cuerpo del hongo y sus axones apropiadamente a los lóbulos del cuerpo del hongo (Figura 4D, F, G). Esto indica que las células recién creadas pueden ser neuronas funcionales involucradas en la reparación de los cuerpos dañados de los hongos. Otras áreas del cerebro que parecieron regenerarse incluyen el cuerpo elipsoide (EB) (Figura 4H, I), los lóbulos antenales (AL) (Figura 4J, K) y el cuerno lateral (LH) (Figura 4L, M) que poseían clones grandes aproximadamente el 26%, 26% y 20% del tiempo, respectivamente (Figura 4N). Estos resultados subrayan la utilidad de este sistema para la investigación de la neurogénesis adulta. En la Figura 5 se muestra un modelo propuesto para la secuencia de eventos que siguen al PTBI y conducen a la generación de nuevas neuronas.

Figura 1: Lesión cerebral traumática penetrante (PTBI) en el cerebro central de Drosophila adulta. (A) Esquema del exterior de una cabeza de mosca adulta. Esta es una vista frontal. Por lo tanto, el lado derecho del animal está a la izquierda del espectador. (B) Esquema del interior de una cabeza adulta de Drosophila con la trayectoria de la lesión indicada en gris. Esta es una vista posterior. Por lo tanto, en esta imagen y figuras posteriores, el lado derecho del cerebro está a la derecha. El PTBI cerebral central afecta múltiples estructuras cerebrales, incluido el cuerpo de hongos (verde) y los tejidos fuera del cerebro, incluido el cuerpo graso (azul) y los hemocitos (rojo). CB = región central del cerebro. OL= región del lóbulo óptico. (C) Vista dorsal de una cabeza adulta viva en la que los cuerpos de hongos (puntas de flecha) están etiquetados con una proteína fluorescente verde (GFP). Este es el "genotipo estándar" (ver texto para más detalles). El protocolo PTBI resulta reproduciblemente en lesiones en los cuerpos de los hongos. Esta figura ha sido adaptada de Reference16. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: PtBI estimula la proliferación celular. Esquemas de control no lesionado (A) y PTBI (B). Los cuadros azules en las esquinas superiores derechas indican las regiones cerebrales que se muestran con mayor aumento en los paneles (C) y (D). (C,D) El anticuerpo PH3 (rojo) se utilizó para evaluar la proliferación celular 24 h después de la lesión. En los cerebros de control (C), hay pocas células PH3 + y ninguna cerca del MB. Sin embargo, en los cerebros PTBI (D), hay células PH3 + cerca del MB. (E) Cuantificación de células proliferantes. Los números reflejan la proliferación de células en todo el cerebro, no solo en las cercanías del cuerpo del hongo. A las 24 h, los cerebros de control no lesionados tenían un promedio de 3 células PH3 + / cerebro (n = 11 cerebros, 28 células), mientras que 24 h después de PTBI, los cerebros tenían un promedio de 11 células PH3 + / cerebro (n = 17 cerebros, 181 células). A los 7 días, los controles no lesionados tienen pocas células EdU +, con un promedio de 2 células EdU + / cerebro (n = 15 cerebros, 24 células), mientras que los cerebros post-PTBI de 7 días tenían un promedio de 11 células EdU + / cerebro (n = 22 cerebros, 238 células). A los 14 días, los controles no lesionados tienen un promedio de 1 célula / cerebro EdU + (n = 8 cerebros, 11 células), mientras que los cerebros post-PTBI de 14 días tienen un promedio de 29 células EdU + / cerebro (n = 14 cerebros, 400 células). Para este conjunto de experimentos, se utilizaron machos adultos jóvenes dentro de las 6 h de la eclosión. Pruebas t no apareadas de control y muestras de PTBI en los valores de rendimiento de 3 puntos de tiempo de p<0.0001, p<0.0001 y p<0.0002, respectivamente. Las barras de error reflejan la desviación estándar (DE). Esta figura ha sido adaptada de Reference16. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: La respuesta proliferativa al PTBI disminuye con la edad. Para explorar si la edad afecta la cantidad de proliferación celular que ocurre después de la lesión, los machos adultos recién eclosados se compararon con animales de 7 días, 14 días y 28 días antes de PTBI, utilizando anti-PH3 para evaluar la proliferación celular 24 h después de la lesión. Las moscas lesionadas dentro de las 6 h de la eclosión tenían un promedio de 11 células PH3+ / cerebro (n = 17 cerebros, 182 células) en comparación con un promedio de 3 células PH3 + / cerebro en controles de la misma edad (n = 11 cerebros, 28 células). Las moscas de hasta 7 días de edad, luego sometidas a PTBI, tenían un promedio de 6 células PH3 + / cerebro (n = 11 cerebros, 65 células) en comparación con los controles de la edad con un promedio de 2 células PH3 + / cerebro (n = 5 cerebros, 12 células). Cuando las moscas envejecieron hasta 14 días antes del PTBI y se ensayaron 24 h después, hubo un promedio de 1 célula / cerebro PH3 + (n = 8 cerebros, 11 células) similar a los controles de la misma edad, que también promediaron 1 célula PH3 + / cerebro (n = 4 cerebros, 2 células). Las moscas de control no lesionadas de 28 días (n = 4, 1 célula) y PTBI (n = 3, 1 célula) promediaron 0 PH3 + células / cerebro. Las pruebas t no apareadas para PTBI para controlar las comparaciones en estos 4 puntos temporales son p<0.0001, p<0.04, p<0.07 y p<0.84, respectivamente. Esta figura ha sido adaptada de Reference16. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: El rastreo del linaje perma-gemelo demuestra la regeneración cerebral y la orientación adecuada de los axones después de PTBI. Se utilizó el sistema de rastreo de linaje perma-gemelo15 para analizar la neurogénesis después de PTBI. Este sistema etiqueta permanentemente las células en división y la progenie con una proteína fluorescente verde (GFP) o una proteína fluorescente roja (RFP). Las moscas se criaron a 17 ° C para mantener el sistema apagado durante el desarrollo. Los machos F1 portadores de transgenes perma-gemelos se recolectaron en la eclosión, luego se lesionaron y se colocaron a 30 ° C para recuperarse durante 2 o 14 días. (A) En controles no lesionados de 2 días, hay algunas células GFP + dispersas por todo el cerebro. (B) A los 14 días, hay relativamente pocas células GFP + presentes en el cerebro central de control. (C) En comparación, los cerebros lesionados de 2 días tienen más células GFP + que tienden a agruparse cerca de la lesión (punta de flecha). (D) A los 14 días después de la lesión, hay clones grandes cerca del sitio de la lesión. Algunos de estos clones tienen axones que se proyectan a lo largo de los tractos del cuerpo del hongo (punta de flecha). Aquí solo se muestra el canal GFP; hubo clones de RFP+ similares en las muestras de PTBI. (E) El número de clones aumenta con el tiempo después del PTBI.Los cerebros no lesionados de control (n = 13) tienen un promedio de 10 clones a los 2 días, mientras que los cerebros PTBI de 2 días (n = 20) tienen un promedio de 23 clones (p<0.00002). A los 7 días, los cerebros de control tenían un promedio de 9 clones por cerebro (n = 18), mientras que los cerebros PTBI de 7 días tenían un promedio de 39 clones por cerebro (n = 16) (valor p<0.00000002). Esto es significativamente mayor que el número de clones observados a los 2 días después de la lesión (valor p<0.0009). En los cerebros de control de 14 días, hay un promedio de 10 clones por cerebro, lo que no es significativamente diferente de los controles de 2 y 7 días. Sin embargo, a los 14 días después de PTBI, hay un promedio de 66 clones de GFP +, que es significativamente más que los controles de edad emparejados (p<0.0000003) o los cerebros post-PTBI de 2 días (valor p<0.0001). Las barras de error reflejan SD. (F-M) PTBI estimula la formación de clones en múltiples regiones del cerebro. Los paneles en el lado izquierdo son esquemas de regiones cerebrales donde se encontraron clones grandes 14 días después de PTBI (A, H, J, L). Los paneles de la derecha muestran grandes aumentos de cerebros representativos (G, I, K, M). Muchos cerebros de 14 días tenían clones que se proyectaban a áreas objetivo particulares. Estos incluyeron el cuerpo del hongo (MB) (F, G), el cuerpo elipsoide (EB) (H, I), el lóbulo antenal (AL) (J, K) y el cuerno lateral (LH) (L, M). (N) Tanto el número de clones como el tamaño de los clones aumentan con el tiempo posterior al PTBI.Las proporciones de regiones cerebrales con clones grandes se calcularon a los 2, 7 y 14 días en los controles y cerebros lesionados. A los 2 días, ~ 8% de los cerebros de control (n = 13) mostraron clones de AL, mientras que no hubo clones de AL en cerebros lesionados de 2 días (n = 20). En cerebros de control de 7 días (n = 18), el 6% tenía AL y el 6% tenía clones de EB. A los 7 días después de PTBI (n = 16), el 6% de los cerebros también tenían clones de AL, el 6% tenía clones de EB y el 19% tenía clones de MB grandes. A los 14 días, los cerebros de control (n = 9) no exhibieron ninguna área específica con clones, mientras que el 47% de los cerebros de PTBI (n = 15) tenían clones de MB, el 20% de los cerebros de PTBI tenían clones de AL y el 27% de los cerebros de PTBI tenían clones de EB, y el 27% tenían clones de LH. Esta figura ha sido adaptada de Reference16. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

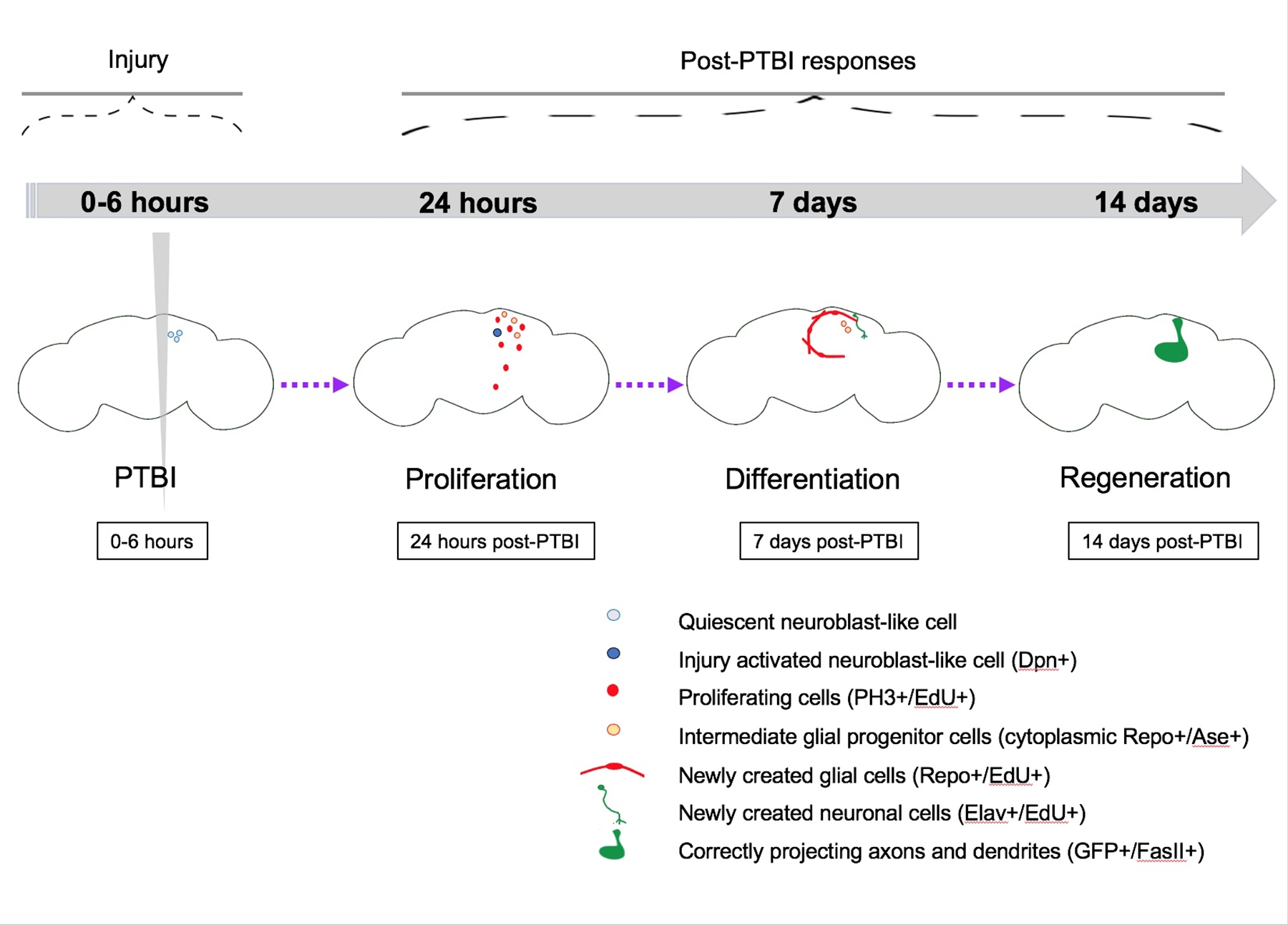
Figura 5: Modelo resumido para la regeneración después de una lesión cerebral traumática penetrante (PTBI). En Drosophila adulta joven, hay células inactivas similares a neuroblastos dentro del cerebro central que carecen de expresión de genes canónicos de neuroblastos. A las 24 horas después del PTBI, las células inactivas similares a los neuroblastos se activan, expresan genes de neuroblastos y han comenzado a proliferar. Tanto a las 4 h como a las 24 h post-PTBI, se produce una ola de muerte celular16. A los 7 días, la tasa de proliferación sigue siendo alta, y muchas de las nuevas células han adoptado identidades celulares maduras, convirtiéndose en neuronas o glía. A los 14 días post-PTBI, grandes clones de nuevas neuronas con axones y dendritas se proyectan correctamente a sus respectivas áreas objetivo. Los defectos locomotores también se restauran a los 14 días, lo que sugiere que la Drosophila adulta puede regenerarse funcional y estructuralmente. Esta figura ha sido adaptada de Reference16. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Discusión
Aunque anteriormente se han descrito lesiones penetrantes en el cerebro adulto de Drosophila15,17,18, estas lesiones se centraron en los lóbulos ópticos y no en el cerebro central. Además, hasta ahora faltan instrucciones detalladas sobre cómo llevar a cabo las lesiones. Este protocolo describe un modelo para la lesión penetrante en el cerebro central adulto de Drosophila que reproduce evidencia estadísticamente significativa para la neurogénesis adulta después de PTBI.
La reproducibilidad de este protocolo PTBI se debe, en parte, al cuerpo del hongo como la región objetivo de la lesión. El cuerpo del hongo es grande, que consiste en ~ 2200 neuronas con dendrita compleja y cenadores de axones en matrices grandes y altamente estereotipadas18. Los cuerpos celulares de las neuronas del cuerpo de hongos se encuentran cerca de la superficie del cerebro y se pueden visualizar a través de la cutícula de la cabeza utilizando la expresión de proteína fluorescente verde (GFP) (Figura 1C). Los precursores del cuerpo de hongos son las últimas células madre neurales en sufrir apoptosis durante el desarrollo13,12,19. Por lo tanto, muchas neuronas del cuerpo de hongos son bastante jóvenes en el momento de la eclosión. Esto llevó a la hipótesis de que el cuerpo del hongo podría tener más potencial mitótico que otras regiones del cerebro16. Además, el cuerpo de hongo es fundamental para el aprendizaje y la memoria18. Esto permite preguntarse si la neurogénesis desencadenada por PTBI conduce a la recuperación funcional.
Otros factores que contribuyen a la reproducibilidad de los resultados incluyen el uso de moscas cruzadas de genotipos consistentes, la realización de cruces en la misma dirección cada vez, el control preciso de las temperaturas de cría y envejecimiento, y el análisis de machos y hembras por separado. El uso de moscas F1 de un cruce reduce la probabilidad de analizar cerebros homocigotos en busca de mutaciones espontáneas. La cruz estándar de y[1] w[1]; UAS-mCD8-GFP;; OK107-GAL4 hembras adultas a y[1] w[1] moscas macho adultas resulta en progenie F1 del genotipo y[1] w[1]; UAS-mCD8-GFP/+;; OK107-GAL4/+. OK107-GAL4 se expresa en todas las neuronas intrínsecas del cuerpo del hongo e impulsa la expresión del reportero unido a la membrana UAS-mCD8-GFP que permite la visualización de los cuerpos del hongo y sus proyecciones. Para los cruces perma-gemelos, los cruces deben permanecer a 17 ° C en todo momento para mantener el sistema de rastreo de linaje apagado. Esto asegura que no se etiqueten las células en división durante el desarrollo y que solo se etiqueten las neuronas y la glía nacidas en adultos. Con este fin, la sala de moscas también se puede mantener a 17 ° C. Aunque la descripción inicial del sistema perma-twin15 recomendaba la cría de moscas a 18 °C, esto puede conducir a un etiquetado de fondo significativo.
Para mayor consistencia, también se recomienda mantener el control de moscas ilesas en la almohadilla de CO2 mientras se lleva a cabo el PTBI. Esto asegura que ambos conjuntos de moscas tengan una exposición anestésica idéntica. Además, es deseable que la reproducibilidad penetre completamente en la cabeza. Sin embargo, se debe tener cuidado de no doblar la punta del pasador contra la almohadilla, dejándola inutilizable para futuras lesiones. Para los profesionales calificados, hay poco daño involuntario a las moscas PTBI. Sin embargo, presionar demasiado fuerte en el tórax para estabilizar la mosca durante la lesión puede ser letal. Una forma de evaluar el alcance de la lesión no intencionada es cuantificar la mortalidad de las moscas PTBI 24 h después de la lesión. Para las moscas lesionadas unilateralmente, esto puede ser del 50% o más para los principiantes. Por lo tanto, para garantizar que los resultados observados se deban a PTBI y no a lesiones no intencionales, se recomienda que los principiantes practiquen la administración de PTBI en ~ 20 moscas diarias durante varias semanas y no analicen los cerebros resultantes hasta que la mortalidad de 24 h sea consistentemente <10%.
Para cuantificar la cantidad de proliferación estimulada por el PTBI central del cerebro, se puede emplear tanto la inmunotinción antifosticois H3 (PH3) como la incorporación de 5-etinil-2'-desoxiuridina (EdU). Anti-PH3 etiqueta las células antes y durante toda la metafase, limitando la detección a solo una fracción de las células que se dividen activamente. Por lo tanto, la tinción anti-PH3 proporciona solo una visión parcial de la proliferación. EdU es un análogo de timidina que se puede incorporar en el ADN recién sintetizado. Al alimentar a las moscas EdU antes y después de la lesión, es posible obtener una imagen más completa de las células que se están dividiendo o que se han dividido después de la lesión. El hecho de que cualquier célula que se divida esté marcada permanentemente es útil tanto para la identificación de células de ciclo lento como para evaluar la supervivencia de las células después de la proliferación inicial. Por razones poco claras, pero tal vez debido a la permeabilidad limitada de la barrera hematoencefálica, el etiquetado EdU es ineficiente y no informa la proliferación celular en el cerebro adulto. Esto se evidencia por el número similar de células PH3 + y EdU + en cerebros de control y experimentales a las 24 h posteriores a PTBI y al observar que solo un subconjunto de nuevas células en clones perma-gemelos incorporan EdU16. Para un etiquetado máximo, es esencial prealimentar a las moscas con EdU porque las moscas lesionadas no reanudan la alimentación durante varias horas después de PTBI. La alimentación se evaluó mediante la adición de colorante alimentario a la solución de EdU y el monitoreo de la cantidad de colorante en el intestino a través de la cutícula abdominal16.
Cabe señalar que, si bien hemos proporcionado un protocolo de disección cerebral en el paso 4, se pueden utilizar técnicas alternativas. Varios de ellos están disponibles en protocolos publicados anteriormente20,21,22. Drosophila melanogaster ofrece un modelo de bajo costo con potentes herramientas genéticas y moleculares que se pueden utilizar para estudiar los mecanismos subyacentes a la regeneración de múltiples tejidos, incluidos el intestino y los componentes del sistema nervioso. Aquí se describe un modelo de lesión novedoso y reproducible que se puede utilizar para estudiar la respuesta a la lesión cerebral. Los datos obtenidos utilizando estos protocolos apoyan la idea de que el cerebro central adulto de Drosophila conserva la capacidad proliferativa, generando nuevas neuronas en respuesta a una lesión. Estas observaciones justifican una mayor investigación tanto del alcance de la neurogénesis adulta como de sus mecanismos moleculares subyacentes. Una vez que los componentes involucrados en la regeneración neuronal se identifican en este sistema, podemos convertir nuestro conocimiento de la neurogénesis de Drosophila adulta en humanos.
Divulgaciones
Los autores no tienen conflictos de intereses.
Agradecimientos
Agradecemos a Stacey Rimkus y Becky Katzenberger por la asistencia técnica y a Eduardo Moreno por compartir las existencias de perma-twin. Nos gustaría agradecer a Barry Ganetzky y David Wassarman por las animadas discusiones que sin duda mejoraron la ciencia y a Kent Mok, Cayla Guerra y Bailey Spiegelberg por sus contribuciones al laboratorio. Los anticuerpos FasII fueron desarrollados por Corey Goodman y obtenidos del Developmental Studies Hybridoma Bank, creado por el NICHD de los NIH y mantenido en la Universidad de Iowa, Departamento de Biología, Iowa City, IA 52242. La mayoría de las cepas de Drosophila utilizadas en este estudio se obtuvieron del Bloomington Drosophila Stock Center (BDSC; NIH P40OD018537). Este trabajo fue apoyado por NIH T32 GM007133 (KLC); NIH NS090190 (GBF); NIH NS102698 (GBF); la Escuela de Graduados de la Universidad de Wisconsin (GBF); y el UW-Madison Women in Science and Engineering Leadership Institute (WISELI) (GBF).
Materiales
| Name | Company | Catalog Number | Comments |
| #11 disposable scalpels | Santa Cruz Biotechnology | sc-395923 | used for separating Drosophila heads from trunks prior to brain dissection |
| 150 mm diameter black Sylgard dishes | Dow | 1696157 | made in the laboratory with reagents from Dow; used for brain dissection |
| 18 mm coverslips | any | for mounting brains on microscope slides | |
| 4',6-Diamidino-2-Phenylindole, Dihydrochloride (DAPI) | ThermoFisher | D1306 | for immunohistochemistry |
| 70% Ethanol | made from 95% ethanol sourced variously | ||
| anti-mouse Cy5 | Jackson ImmunoResearch | 715-175-151 | for immunohistochemistry |
| anti-rabbit 568 | ThermoFisher | A11036 | for immunohistochemistry |
| bovine serum albumin (BSA) | SIgma Aldrich | A7030 | for immunohistochemisty |
| Clear nail polish | any | for sealing coverslips | |
| Click-It EdU labeling kit | InVitrogen | C10640 | to detect newly synthesized DNA |
| CO2 bubbler | Genesee Scientific | 59-181 | for anesthesia |
| CO2 pad | Genesee Scientific | 59-114 | for anesthesia |
| CO2 regulator and supply | any | for anesthesia | |
| Confocal microscope | any | for imaging fixed, stained and mounted brains | |
| cotton plugs | Genesee Scientific | 51-101 | for EdU labeling |
| Drosophila vials | Genesee Scientific | 32-109 | for EdU labeling |
| Fix buffer (Pipes, EGTA, Magnesium; PEM) | components sourced from various companies | for fixing adult brains; 100 mM piperazine-N,N’-bis(2-ethanesulfonic acid) [PIPES], 1 mM EGTA, 1 mM MgSO4, pH 7.0 | |
| Formaldehyde | Sigma Aldrich | 252549 | for fixing adult brains, added to PEM |
| Grade 3 round Whatman filters, 23 mm round | Tisch Scientific | 1003-323 | for EdU labeling |
| Microfuge tubes | any | for fixing and staining reactions and for storing Minutien pins | |
| Microscope slides | any | for mounting brains | |
| Minutien pins | Fine Science Tools | 26002-10 | for brain injury; 12.5 μm diameter tip and 100 μm diameter rod |
| mouse anti-Fasiclin II | Developmental Studies Hybridoma Bank | 1D4-s | for immunohistochemistry |
| NIGHTSEA stereo microscope fluorescence adaptor | Electron Microscopy Sciences | SFA-GR | fluorescence setup for dissecting microscope |
| P20, P200 and P1000 pipettors and tips | any | for measuring solutions | |
| phosphate buffered saliine (PBS) | components sourced from various companies | for dissecting brains and making immunohistochemistry blocking and washing solutions; 100 mM of K2HPO4, 140 mM of NaCl, pH 7.0 | |
| phosphate buffered saline with 0.1% Triton X-100 (PT) | components sourced from various companies | for washing dissected brains | |
| phosphate buffered saline with 0.1% Triton X-100 + 2% bovine serum albumin (PBT) | components sourced from various companies | blocking solution for immunohistochemistry and for diluting antibodies | |
| rabbit anti-PH3 | Santa Cruz Biotechnology, Inc | sc-8656-R | for immunohistochemistry |
| Reinforcement labels | Avery | 5721 | to maintain space between the microscope slide and the coverslip |
| Size 0 paintbrushes | any | to manipulate and stabilize adult Drosophila during injury | |
| Triton X-100 | Sigma Aldrich | 93443 | |
| Two pair of #5 watchmakers forceps | Fine Science Tools | 11255-20 | used to hold the Minutien pins and for brain dissections |
| Vectashield | Vector Laboratories | H-1000 | mounting medium for microscope slides |
Referencias
- . Report to Congress: Traumatic brain injury in the United States Available from: https://www.cdc.gov/traumaticbraininjury/pubs/tbi_report_to_congress.html (1999)
- . NIEHS Available from: https://www.nih.gov/research/supported/health/neurodegenerative/index.cfm (2021)
- Morton, N. V., Wehman, P. Psychosocial and emotional sequelae of individuals with traumatic brain injury: A literature review and recommendations. Brain Injury. 9 (1), 81-92 (2017).
- Bonini, N. M., Berger, S. L. The sustained impact of model organisms-in genetics and epigenetics. Genetics. 205, 1-4 (2017).
- Brace, E. J., DiAntonio, A. Models of axon regeneration in Drosophila. Experimental Neurology. 287, 310-317 (2017).
- Hao, Y., Collins, C. Intrinsic mechanisms for axon regeneration: insights from injured axons in Drosophila. Current Opinion in Genetics & Development. 44, 84-91 (2017).
- Chiang, A. S., et al. Three-dimensional reconstruction of brain-wide wiring networks in Drosophila at single-cell resolution. Current Biology. 21 (1), 1-11 (2011).
- Meinertzhagen, I. A. The organisation of invertebrate brains: cells, synapses and circuits. Acta Zoologica. 91 (1), 64-71 (2010).
- Bellen, H. J., Tong, C., Tsuda, H. 100 years of Drosophila research and its impact on vertebrate neuroscience: A history lesson for the future. Nature Reviews Neuroscience. 11 (7), 514-522 (2010).
- Lessing, D., Bonini, N. M. Maintaining the brain: insight into human neurodegeneration from Drosophila melanogaster mutants. Nature Reviews Genetics. 10 (6), 359-370 (2009).
- Boone, J. Q., Doe, C. Q. Identification of Drosophila type II neuroblast lineages containing transit amplifying ganglion mother cells. Developmental Neurobiology. 68 (9), 1185-1195 (2008).
- Ito, K., Hotta, Y. Proliferation pattern of postembryonic neuroblasts in the brain of Drosophila melanogaster. Developmental Biology. 149 (1), 134-148 (1992).
- Siegrist, S. E., Haque, N. S., Chen, C. H., Hay, B. A., Hariharan, I. K. Inactivation of both Foxo and reaper promotes long-term adult neurogenesis in Drosophila. Current Biology. 20 (7), 643-648 (2010).
- von Trotha, J. W., Egger, B., Brand, A. H. Cell proliferation in the Drosophila adult brain revealed by clonal analysis and bromodeoxyuridine labelling. Neural Development. 4, 9 (2009).
- Fernandez-Hernandez, I., Rhiner, C., Moreno, E. Adult neurogenesis in Drosophila. Cell Reports. 3 (6), 1857-1865 (2013).
- Crocker, K. L., et al. Neurogenesis in the adult Drosophila brain. Genetics. , (2021).
- Plavicki, J., Mader, S., Pueschel, E., Peebles, P., Boekhoff-Falk, G. Homeobox gene distal-less is required for neuronal differentiation and neurite outgrowth in the Drosophila olfactory system. Proceedings of the National Academy of Sciences of the United States of America. 109 (5), 1578-1583 (2012).
- Aso, Y. The neuronal architecture of the mushroom body provides a logic for associative learning. Elife. 3, 04577 (2014).
- Ito, K., Awano, W., Suzuki, K., Hiromi, Y., Yamamoto, D. The Drosophila mushroom body is a quadruple structure of clonal units each of which contains a virtually identical set of neurones and glial cells. Development. 124 (4), 761-771 (1997).
- Tito, A. J., Cheema, S., Jiang, M., Zhang, S. A. Simple one-step dissection protocol for whole-mount preparation of adult Drosophila brains. Journal of Visualized Experiments. (118), e55128 (2016).
- Kelly, S. M., Elchert, A., Kahl, M. Dissection and immunofluorescent staining of mushroom body and photoreceptor neurons in adult Drosophila melanogaster brains. Journal of Visualized Experiments. (129), e56174 (2017).
- Arain, U., Valentino, P., Islam, I. M., Erclik, T. Dissection, immunohistochemistry and mounting of larval and adult Drosophila brains for optic lobe visualization. Journal of Visualized Experiments. (170), e61273 (2021).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoExplorar más artículos
This article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados