
Method Article
Stimulation und Analyse der adulten Neurogenese im zentralen Gehirn von Drosophila
In diesem Artikel
Zusammenfassung
Dieser Artikel enthält detaillierte Protokolle zur Zufügung einer penetrierenden traumatischen Hirnverletzung (PTBI) bei erwachsenen Drosophila und zur Untersuchung der daraus resultierenden Neurogenese.
Zusammenfassung
Die molekularen und zellulären Mechanismen, die der Neurogenese als Reaktion auf Krankheiten oder Verletzungen zugrunde liegen, sind nicht gut verstanden. Das Verständnis dieser Mechanismen ist jedoch entscheidend für die Entwicklung neuronaler regenerativer Therapien. Drosophila melanogaster ist ein führendes Modell für Studien zur neuronalen Entwicklung, wurde aber in der Vergangenheit nicht zur Untersuchung der Regeneration des Gehirns von Erwachsenen genutzt. Dies liegt in erster Linie daran, dass das erwachsene Gehirn eine sehr geringe mitotische Aktivität aufweist. Nichtsdestotrotz löst die durchdringende traumatische Hirnverletzung (PTBI) im erwachsenen Drosophila-Zentralhirn die Bildung neuer Neuronen und neuer Gliazellen aus. Die leistungsstarken genetischen Werkzeuge, die in Drosophila verfügbar sind, kombiniert mit dem einfachen, aber strengen Verletzungsprotokoll, das hier beschrieben wird, machen das erwachsene Drosophila-Gehirn zu einem robusten Modell für die neuronale Regenerationsforschung. Hier finden Sie detaillierte Anweisungen für (1) penetrierende Verletzungen des erwachsenen Zentralhirns und (2) Dissektion, Immunhistochemie und Bildgebung nach verletzungen. Diese Protokolle liefern hochgradig reproduzierbare Ergebnisse und werden zusätzliche Studien zur Analyse der Mechanismen der neuronalen Regeneration erleichtern.
Einleitung
Schäden am Gehirn und Nervensystem sind weltweit eine der Hauptursachen für Tod und Behinderung. Etwa 1,5 Millionen Amerikaner erleiden jedes Jahr traumatische Hirnverletzungen (TBI1), während schätzungsweise 6 Millionen Menschen allein in den Vereinigten Staaten an neurodegenerativen Erkrankungen wie Parkinson und Alzheimer leiden2. Sowohl Krankheiten als auch Verletzungen des Gehirns können eine neuronale Degeneration verursachen, die zu sensorischen, kognitiven und motorischen Defekten führt3. Die Entwicklung therapeutischer Strategien für die Reparatur des menschlichen Gehirns war aufgrund der komplexen Physiologie des Gehirns schwierig. Modellorganismen wie Drosophila melanogaster bieten ein einfaches System zur Identifizierung der grundlegenden Mechanismen, die der Neurodegeneration und potenziellen therapeutischen Zielen zugrunde liegen4.
Die Fruchtfliege Drosophila melanogaster ist seit mehr als einem Jahrhundert ein mächtiger Modellorganismus, der die Bereiche Genetik, Entwicklungsbiologie und Neurowissenschaften voranbringt5,6. Das Drosophila-Gehirn umfasst nur ~ 90.000 Neuronen7, eine Million Mal weniger als das durchschnittliche menschliche Gehirn8, aber sie haben viele Ähnlichkeiten. Sowohl menschliche als auch fliegende Gehirne nutzen die Neurotransmitter GABA, Glutamat, Acetylcholin und die biogenen Amine Dopamin und Serotonin9. Drosophila und menschliche Neuronen funktionieren ebenfalls ähnlich, mit einer gemeinsamen synaptischen Architektur und analogen neuronalen Zelltypen10. Die kleinere Gehirngröße von Drosophila und die Verfügbarkeit fortschrittlicher genetischer Techniken in Kombination mit der Erhaltung molekularer, zellulärer und physiologischer Mechanismen zwischen Drosophila und Säugetieren ermöglichen es drosophila-Forschern, Fragen zu stellen, die in Säugetiermodellen unpraktisch oder schwer zu beantworten sind.
Unser derzeitiges Verständnis der adulten Neurogenese bei Drosophila, sowohl während der Homöostase als auch nach einer Verletzung, bleibt begrenzt. Über die Neurogenese während der normalen Entwicklung ist mehr bekannt. Zum Beispiel werden Neuronen und Gliazellen während der Entwicklung aus Vorläuferzellen, neuroblasten genannt, erzeugt10,11. Mindestens drei verschiedene Arten von Neuroblasten wurden im zentralen Gehirn unterschieden. Sowohl Typ-I- als auch Typ-II-Linien-Neuroblasten verlassen den Zellzyklus ~ 20-30 h nach der Pupariumsbildung12. Im Gegensatz dazu sind die Neuroblasten des Pilzkörpers die letzten, die die Zellteilung beenden, und zwar über reaper-abhängige Apoptose ~85-90 h nach Pupariumsbildung13. Nach der Eklosion hat das erwachsene Drosophila-Gehirn nur wenige sich teilende Zellen (~ 1 Zelle / Gehirn), überwiegend Glia14. Die adulten Sehlappen besitzen langsam zyklische Neuroblasten, die zur Neurogenese fähig sind15, während das erwachsene Zentralgehirn keine bekannten Neuroblasten hat. Der Mangel an neuronalen Vorläufern und die begrenzte Zellproliferation ähneln stark der Situation im erwachsenen Säugetiergehirn und unterstreichen die potenzielle Relevanz der Mechanismen der adulten Neurogenese in Drosophila für den Menschen.
Die Entdeckung eines niedrigen Niveaus der adulten Neurogenese in den adulten Drosophila-Optiklappen nach einer Verletzung15 führte zu der Hypothese, dass das erwachsene Drosophila-Zentralhirn auch zur adulten Neurogenese fähig sein könnte16. Dieses Protokoll beschreibt die Erstellung eines rigorosen, reproduzierbaren Modells der zentralen Hirnverletzung bei erwachsenen Drosophila , das zur Untersuchung der Neurogenese im erwachsenen Zentralgehirn verwendet werden kann. Angesichts der Ähnlichkeiten zwischen der Architektur und Funktion des menschlichen und des Drosophila-Gehirns könnten diese Entdeckungen zur Identifizierung kritischer Ziele für die therapeutische Neurogenese in verletzten und erkrankten menschlichen Gehirnen führen.
Protokoll
Dieses Protokoll folgt den Tierpflegerichtlinien von UW-Madison.
1. Generierung von Drosophila für Erwachsene für PTBI
- Für das Standardkreuz platziere 20 jungfräulich y[1] w[1]; UAS-mCD8-GFP;; OK107-GAL417 erwachsene Weibchen und 10 y[1] w[1]17 erwachsene männliche Fliegen zusammen in Fläschchen (siehe Tabelle der Materialien), die Nahrung enthalten. Um eine große Anzahl von synchronen Nachkommen zu haben, richten Sie 10-20 Kreuze gleichzeitig ein. Die Standardkreuzung führt zu F1-Nachkommen des Genotyps: y[1] w[1]; UAS-mCD8-GFP/+;; OK107-GAL4/+.
- Stellen Sie die Durchstechflaschen zur Paarung und Eiablage bei 25 °C ab. Um die Nachkommenschaft zu maximieren, übertragen Sie die Eltern alle 2-4 Tage in neue Fläschchen, wobei die früheren Durchstechflaschen bei 25 ° C bleiben. Entsorgen Sie jede Durchstechflasche 18 Tage, nachdem die Eltern zum ersten Mal hineingelegt wurden, um sicherzustellen, dass es keine F2-Nachkommen gibt.
HINWEIS: Aus jeder Gruppe von Eltern können 3 Gruppen von Nachkommen ("Bruten") erzeugt werden. - Überprüfen Sie nach ~ 10 Tagen, wann die F1-Nachkommen anfangen zu ekleinieren.
- Stellen Sie die Durchstechflaschen zur Paarung und Eiablage bei 25 °C ab. Um die Nachkommenschaft zu maximieren, übertragen Sie die Eltern alle 2-4 Tage in neue Fläschchen, wobei die früheren Durchstechflaschen bei 25 ° C bleiben. Entsorgen Sie jede Durchstechflasche 18 Tage, nachdem die Eltern zum ersten Mal hineingelegt wurden, um sicherzustellen, dass es keine F2-Nachkommen gibt.
- Verwenden Sie für Abstammungsstudien F1 Perma-Zwillingsmännchen15 des Genotyps: w; FRT40A, UAS-CD2-RFP, UAS-GFP-Mir/FRT40A, UAS-CD8-GFP, UAS-CD2-Mir; tub-GAL80ts/act-GAL4 UAS-flp. Aus Konsistenzgründen führen Sie dieses Kreuz jedes Mal in die gleiche Richtung durch.
- Um Perma-Zwillingsmännchen zu erzeugen, platzieren Sie 20 jungfräuliche w; FRT40A, UAS-CD2-RFP, UAS-GFP-Mir; tub-GAL80ts/TM6B Buchsen und 10 w; FRT40A, UAS-CD8-GFP, UAS-CD2-Mir; act-GAL4 UAS-flp/TM6B15 erwachsene Männer zusammen in Fläschchen mit Nahrung.
- Stellen Sie die Durchstechflaschen zur Paarung und Eiablage bei 17 °C auf. Legen Sie die Eltern alle 7 Tage in Fläschchen mit neuen Lebensmitteln, wobei alle Durchstechflaschen bei 17 ° C gehalten werden. Entsorgen Sie jede Durchstechflasche 35 Tage, nachdem die Eltern zum ersten Mal hineingelegt wurden, um sicherzustellen, dass es keine F2-Nachkommen gibt.
- Überprüfen Sie nach ~ 21 Tagen, wann die F1-Nachkommen anfangen zu ekleinieren.
2. Penetrierende traumatische Hirnverletzung (PTBI; Abbildung 1)
- Sortieren Sie neu emittierte F1-Fliegen. Wählen Sie junge Männchen innerhalb von 6 h nach der Eklosion aus. Legen Sie diese Männchen in saubere Fläschchen mit Futter, mit 40 oder weniger Fliegen pro Fläschchen.
HINWEIS: Dies wird am einfachsten am Vormittag erreicht, indem Fliegen auf dem CO2-Pad betäubt und erwachsene Männchen identifiziert werden, die noch Mekonium (sichtbar als dunkelgrünlicher Fleck durch die Bauchdecke) in ihren Eingeweiden haben. - Vorfütterung mit 5-Ethynyl-2'-desoxyuridin (EdU) (siehe Materialtabelle) für 6 h vor der Verletzung, wenn geplant ist, sich teilende Zellen mit EdU zu markieren. Weitere Informationen finden Sie in Schritt 3.
- Desinfizieren Sie Minutien-Pins (siehe Materialtabelle) für mindestens 5 minuten, indem Sie ~ 100 Pins in ein 1,5 ml Mikrozentrifugenrohr legen, das mit 70% Ethanol gefüllt ist.
- Desinfizieren Sie das CO2-Pad und einen Pinsel, indem Sie 70% Ethanol sprühen und mit einem sauberen fusselfreien Taschentuch trocken wischen. Sobald die Werkzeuge sauber und trocken sind, übertragen Sie 40 oder weniger sortierte F1-Männchen zurück auf das saubere Pad.
- Teilen Sie die F1-Männchen auf dem Fliegenkissen in 2 Gruppen auf. Eine Gruppe wird als Kontrolle dienen, unverletzte Fliegen. Die zweite experimentelle Gruppe wird dem PTBI unterzogen.
- Ziehen Sie mit einer Pinzette 4-5 neue Minutien-Stifte aus dem Mikrozentrifugenrohr und platzieren Sie sie in der Nähe des Randes des CO2-Pads . Wählen Sie unter dem Sezierbereich einen geraden Minutien-Stift mit einer scharfen Spitze.
HINWEIS: Die Verwendung eines einzelnen Minutien-Pins für ein Experiment reduziert die Variabilität. Verwenden Sie scharfe Stifte wieder. Legen Sie beschädigte oder stumpfe Stifte in ein separates 1,5 ml Mikrozentrifugenröhrchen mit 70% Ethanol zur sicheren Entsorgung. - Wenn Sie mit Fliegen aus dem Standardkreuz arbeiten, schalten Sie die Stereomikroskop-Leuchtdiodenlampe (LED) ein, die mit den entsprechenden Anregungs- und Emissionsfiltern für grünes Fluoreszenzprotein (GFP) ausgestattet ist. Dies ermöglicht die Anregung bei 440-460 nM und ermöglicht die Visualisierung von 500-560 nM.
HINWEIS: Mit diesem Lampen- und Filterset fluoreszieren die Zellkörper des Pilzkörpers grün durch die Kopfkutikula (Abbildung 1C). Für Perma-Zwillingsfliegen oder Fliegen anderer Genotypen kann ein Standard-Weißlichtbeleuchter für das Stereomikroskop verwendet werden, um Landmarken auf der Kopfkutikula zu visualisieren, um die Verletzung des Pilzkörpers anzuvisieren (Abbildung 1C). - Verwenden Sie die Pinzette, um den ausgewählten Minutien-Stift in einer Hand (für Rechtshänder ist dies normalerweise die rechte Hand) und den Pinsel in der anderen (normalerweise linken) Hand aufzunehmen und zu halten. Wählen Sie eine Fliege aus der Versuchsgruppe und positionieren Sie die Fliege so, dass Sie einen Rückenblick auf die Kopfkapsel mit dem Kopf der Fliege nach rechts haben. Legen Sie die Bürste auf das Vorderteil des dorsalen Thorax und drücken Sie sie sanft nach unten, um die Fliege zu stabilisieren.
- Richten Sie die Spitze des Minutien-Stifts auf die Zellkörper des Pilzkörpers auf der rechten Seite des Kopfes und dringen Sie in die Kopfkapsel ein. Wenn Sie Landmarken verwenden, zielen Sie auf die dorsale Kopfkutikula zwischen den Ocelli und dem dorsalen Rand des Auges (Abbildung 1).
- Nachdem Sie die Verletzung abgeschlossen haben, verwenden Sie den Pinsel, um den Kopf sanft vom Minutien-Stift zu drücken.
- Wenn Sie das Gehirn für RNA-Seq oder qRT-PCR verwenden, machen Sie eine zweite Verletzung auf der linken Seite des Kopfes.
- Wiederholen Sie die Schritte 2.8-2.10, um alle Fliegen der Versuchsgruppe zu verletzen.
- Sobald alle Fliegen verletzt wurden, legen Sie die Kontroll- und verletzten Fliegen in markierte, separate Fläschchen mit Nahrung. Legen Sie die Fläschchen horizontal (d.h. auf ihre Seiten), während sich die Fliegen von der Narkose und während der anschließenden Alterung erholen, um zu verhindern, dass sich Fliegen in der Nahrung verfangen.
- Platzieren Sie Standard-Kreuzfliegen bei 25 ° C und Perma-Zwillingsfliegen bei 30 ° C bis zum Alter.
- Für Fliegen, die länger als 24 h gealtert sind, legen Sie sie alle 1-2 Tage auf saubere Nahrung.
3. EdU-Kennzeichnung
- Bereiten Sie einen Vorrat von 10 mM 5-Ethynyl-2'-desoxyuridin (EdU) in Dimethylsulfoxid (DMSO) vor. Diese kann bei -20 °C bis zu 12 Monate gelagert werden.
- 200 μL 50 μM EdU in 10% Saccharose zubereiten. Pre-Feed-Fliegen mit EdU für 6 h vor PTBI.
- Geben Sie 200 μL 50 μM EdU in 10% Saccharose auf ein 23 mm rundes Filterpapier der Klasse 3 (siehe Materialtabelle) in eine ansonsten leere Durchstechflasche.
- Legen Sie die Fliegen in die Durchstechflasche. Verschließen Sie dann die Durchstechflasche mit einem Baumwollpfropfen.
- Legen Sie die Durchstechflasche horizontal in einen befeuchteten Inkubator bei 25 °C für 6 h.
- Führen Sie PTBI wie in den Schritten 2.3-2.10 beschrieben durch.
- Legen Sie die Fliegen wieder in die EdU-haltige Durchstechflasche. Verschließen Sie die Durchstechflasche mit einem Baumwollpfropfen.
- Legen Sie die Durchstechflasche horizontal in einen befeuchteten Inkubator bei 25 °C für bis zu 24 h. Führen Sie einen der unten beschriebenen Schritte aus (Schritt 3.8.1-3.8.3).
- Sezieren und fixieren Sie Gehirne wie in Schritt 4 beschrieben mit Fixiermittel, Waschpuffer und Blockierpuffer ohne Azid und mit der EdU-Nachweisreaktion, die vor der Antikörperfärbung durchgeführt wird.
- Die Fliegen in eine saubere Durchstechflasche mit einem neuen Filterpapier und 200 μL 50 μM EdU in 10% Saccharose geben. Legen Sie die Durchstechflasche horizontal in einen 25 °C großen Inkubator. Wiederholen Sie dies während der Dauer der Etikettierung alle 24 h. Dann sezieren und fixieren Sie Gehirne wie in Schritt 4 beschrieben unter Verwendung von Puffern ohne Azid mit der EdU-Nachweisreaktion, die vor der Antikörperfärbung durchgeführt wird.
- Um das Pulse-Chase-Etikett mit EdU zu verfolgen, EdU für die Pulsperiode zuzuführen, übertragen Sie die Fliegen alle 24 h in eine saubere Durchstechflasche, die ein neues Filterpapier und 200 μL 50 μM in 10% Saccharose enthält. Nach der Pulsperiode (z. B. 4 Tage) die Fliegen in eine Durchstechflasche mit Standard-Drosophila-Futter geben. Legen Sie die Durchstechflasche für weitere 3 Tage auf die Seite in einen 25 °C-Inkubator. Dann sezieren und fixieren Sie Gehirne wie in Schritt 4 beschrieben unter Verwendung von Puffern ohne Azid mit der EdU-Nachweisreaktion, die vor der Antikörperfärbung durchgeführt wird.
4. Dissektion, Immunhistochemie und Montage
- 1,5 mL Mikrozentrifugenröhrchen mit 100 μL Fixiermittel: 4% Formaldehyd in PEM (100 mM Piperazin-N,N'-bis(2-ethansulfonsäure) [PIPES], 1 mM EGTA, 1 mM MgSO4, pH 7,0) (siehe Materialtabelle) herstellen und auf Eis legen.
HINWEIS: Bis zu 20 Gehirne eines einzigen Genotyps und Zustands können in einem einzigen Röhrchen verarbeitet werden. - Bereiten Sie eine Dissektionsplatte mit einem kleinen Pool (~ 100 μL) aus 70% Ethanol und drei kleinen Pools aus phosphatgepufferter Kochsalzlösung (PBS; 100 mM K2HPO4, 140 mM NaCl, pH 7,0) vor.
- Betäuben Sie ~ 10 Kontroll- oder Versuchsfliegen auf einem CO2-Pad , das mit 70% Ethanol desinfiziert wurde.
- Trennen Sie den Kopf mit einem Skalpell vom Rumpf jeder Fliege.
- Sammeln Sie die Köpfe mit einem mit 70% Ethanol benetzten Pinsel und legen Sie die Köpfe für 2-5 min in den Ethanolpool auf die Sezierplatte.
HINWEIS: Dies dehydriert die Gehirne leicht und macht es einfacher, sie von der Kopfkutikula weg zu sezieren. - Übertragen Sie die Köpfe in einen ~ 100 μL-Pool von PBS und sezieren Sie die Gehirne, wobei jedes Gehirn zu einem sauberen ~ 100 μL-Pool von PBS bewegt wird. Führen Sie dies mit zwei Paar Watchmakers-Pinzetten durch, um die Rückseite der Kopfkutikula zu öffnen, und halten Sie die Nagelhaut mit einer Pinzette fest, während Sie das Gehirn mit der geschlossenen Spitze des zweiten Zangenpaares vorsichtig aus der Nagelhaut herausheben.
- Übertragen Sie die sezierten Gehirne in ein Mikrozentrifugenröhrchen, das 100 μL der Fixierlösung enthält, indem Sie einen P200-Pipettor verwenden, der mit einer Kunststoffspitze ausgestattet ist, die geschnitten und abgeschrägt wurde, um den Eintritt in das Gehirn zu ermöglichen.
- Für 20-25 min bei Raumtemperatur fixieren. Entfernen Sie die Fixierung vorsichtig mit einer P200 oder einer Glaspipette.
- Waschen Sie die fixierten Gehirne viermal mit 1 ml "PT" (PBS plus 0,1% Triton X-100), so dass sich das Gehirn zwischen jeder Wäsche mehrere Minuten niederlassen kann.
HINWEIS: Wenn sich das Gehirn zwischen den Waschungen nicht schnell absetzt, haben sie wahrscheinlich noch Fettkörper und / oder Luftröhre. - Blockproben in 1 ml PBS plus 0,1% Triton X-100 und 2% Rinderserumalbumin (PBT) für ~1 h bei Raumtemperatur.
- Entfernen Sie die Blocklösung und inkubieren Sie die Proben mit 100 μL primärer Antikörperlösung über Nacht bei 4 °C in PBT. Die primären Antikörper, die in dieser Studie verwendet werden, sind Kaninchen-Anti-PH3 (1:500) und Maus-Anti-Fasiclin II (1:20) (siehe Materialtabelle).
- Waschen Sie Proben fünfmal mit 1 ml PT, so dass sich das Gehirn zwischen jeder Wäsche mehrere Minuten niederlassen kann.
- Die letzte Wäsche entfernen und in 100 μL sekundärer Antikörperlösung über Nacht bei 4 °C inkubieren. Die hier verwendeten sekundären Antikörper sind Anti-Kaninchen 568 (1:400) und Anti-Maus Cy5 (1:100) (siehe Materialtabelle).
- Waschen Sie Proben fünfmal mit 1 ml PT, so dass sich das Gehirn zwischen jeder Wäsche mehrere Minuten niederlassen kann. Während der letzten Wäsche 4',6-Diamidino-2-Phenylindol, Dihydrochlorid (DAPI; 1:100 einer 10 μM-Lösung) für 10 min hinzufügen, um die Kerne zu färben. Entfernen Sie die Wäsche und lassen Sie 50-100 μL in jedem Röhrchen.
- Bereiten Sie Objektträger vor, indem Sie ein einzelnes, selbstklebendes Verstärkungsetikett auf die Mitte jedes Objektträgers und einen 50-μL-Tröpfchen mit Anti-Fade-Montagemedien in der Mitte jedes Etiketts legen (siehe Materialtabelle). Das Verstärkungsetikett hält den Deckstab leicht über den Dias und verhindert, dass die montierten Gehirne übermäßig abgeflacht werden.
- Übertragen Sie die Gehirne vorsichtig von jedem Mikrozentrifugenröhrchen auf einen vorbereiteten Objektträger mit möglichst wenig Waschpuffer mit einem P200, der mit einer geschnittenen und abgeschrägten Kunststoffspitze ausgestattet ist. Bis zu 10 Gehirne können auf einem einzigen Objektträger montiert werden.
- Verwenden Sie eine Pinzette, um das Gehirn vorsichtig neu zu positionieren, bevor Sie einen Deckglas auf jeden Objektträger legen und den Objektträger mit Nagellack versiegeln. Orientieren Sie das Gehirn entweder mit der hinteren Seite nach oben oder mit der vorderen Seite nach oben, sollten sich aber nicht berühren.
HINWEIS: Da es schwierig ist, qualitativ hochwertige konfokale Bilder durch ein ganzes Gehirn zu erhalten, sollten einige Gehirne in jeder Ausrichtung montiert werden. - Lagern Sie vorbereitete Objektträger flach und im Dunkeln bei 4 °C bis zur Bildgebung. Bei längerfristiger Lagerung (bis zu 1 Jahr) können Objektträger bei -20 °C gelagert werden.
5. Konfokale Bildgebung
HINWEIS: Bildgehirne unter Verwendung eines konfokalen Laserscanning-Mikroskops mit Anregungslasern und Emissionsfilterwürfeln, die für DAPI und die fluoreszierenden sekundären Antikörper geeignet sind (d. h. 405 nm, 488 nm bzw. 568 nm, 633 nm).
- Schalten Sie das Mikroskop, die Laser, den Controller und den Computer ein.
- Öffnen Sie die Akquisitionssoftware.
- Wählen Sie im Erfassungsmodus bis zu vier Kanäle aus und stellen Sie den Parameter für das sequenzielle Scannen der gewünschten Kanäle ein. Erhöhen Sie die Laserleistung für jeden Kanal auf 5-10%.
- Legen Sie einen Objektträger auf das Mikroskop.
- Wählen Sie ein Gehirn aus, das mit dem Epifluoreszenzaufsatz abgebildet werden soll (siehe Materialtabelle).
- Wählen Sie im Aufnahmemodus 1024 x 1024 Pixel als Frame-Dimension aus.
- Wählen Sie im Erfassungsmodus die Option Z-Stack, geben Sie an, dass der Z-Stack mit 2 μm-Abschnitten aufgenommen werden soll, und fokussieren Sie durch die Probe, indem Sie die obere und untere Brennebene auswählen, die abgebildet werden soll.
- Sammeln Sie Z-Stack-Bilder von ganzen Gehirnen mit einem 20-fachen Objektiv und bestimmten Gehirnregionen wie dem Pilzkörper mit einem 60-fachen Objektiv.
6. Datenanalyse
- Quantifizieren Sie die sich ausbreitenden Zellen und die Anzahl der Perma-Zwillings-Klone manuell und/oder mit der Bildanalysesoftware (siehe Materialtabelle). Wählen Sie bei der Verwendung von Software Regions of Interest (ROIs) mit Bereichen von mindestens 10 μm aus.
Ergebnisse
PTBI stimuliert die Zellproliferation
Um das Ausmaß der Neurogenese nach einer PTBI im zentralen Gehirn zu bestimmen, wurde die proliferative Reaktion bei jungen erwachsenen Männern gemessen, die innerhalb von 6 Stunden nach der Eklosion gesammelt und verletzt wurden. Ein signifikanter Anstieg der Proliferation wurde 24 h nach der Verletzung unter Verwendung von Antiphosphohton 3 (PH3), einem Marker für Zellen, die sich aktiv einer Mitose unterziehen, beobachtet. Etwa 3 PH3+-Zellen in Kontroll-Zentralhirnen und 11 PH3+-Zellen in den verletzten Zentralhirnen werden 24 h nach PTBI beobachtet (Abbildung 2A-D). Die Mehrheit der sich teilenden Zellen befindet sich in der Nähe der Verletzungsstelle. Ein zweiter Assay zur Zellteilung wurde verwendet, um die kumulative Zellproliferation aus einer einzigen Verletzung zu quantifizieren und zu beurteilen, inwieweit die neu geschaffenen Zellen überlebten. 5-Ethynyl-2'-desoxyuridin (EdU) ist ein Thymidin-Analogon, das in neu synthetisierte DNA eingebaut werden kann und Zellen, die einer DNA-Synthese unterzogen wurden, dauerhaft markiert. Die Fliegen erhielten einen 4-tägigen Puls von EdU, gefolgt von einer 3-tägigen Verfolgungsjagd. Dies zeigte, dass die markierten Zellen lebensfähig waren und mindestens 3 Tage nach der Proliferation überlebten. Nach 7 Tagen gab es durchschnittlich 2 EdU+ Zellen in zentralen Kontrollgehirnen bzw. durchschnittlich 11 EdU+ Zellen in den verletzten Zentralhirnen (Abbildung 2E). Dies ist vergleichbar mit den Ergebnissen, die 24 h nach der Verletzung mit dem PH3-Antikörper erzielt wurden. Wenn die Zellproliferation nach 14 Tagen gemessen wird, haben die unverletzten Kontrollen durchschnittlich 1 EdU + -Zelle pro zentralem Gehirn, während die verletzten Gehirne durchschnittlich 29 EdU + -Zellen (Abbildung 2E) hatten, was zeigt, dass die Zellproliferation mindestens in der zweiten Woche nach einer PTBI anhält.
Zellproliferation ist altersabhängig
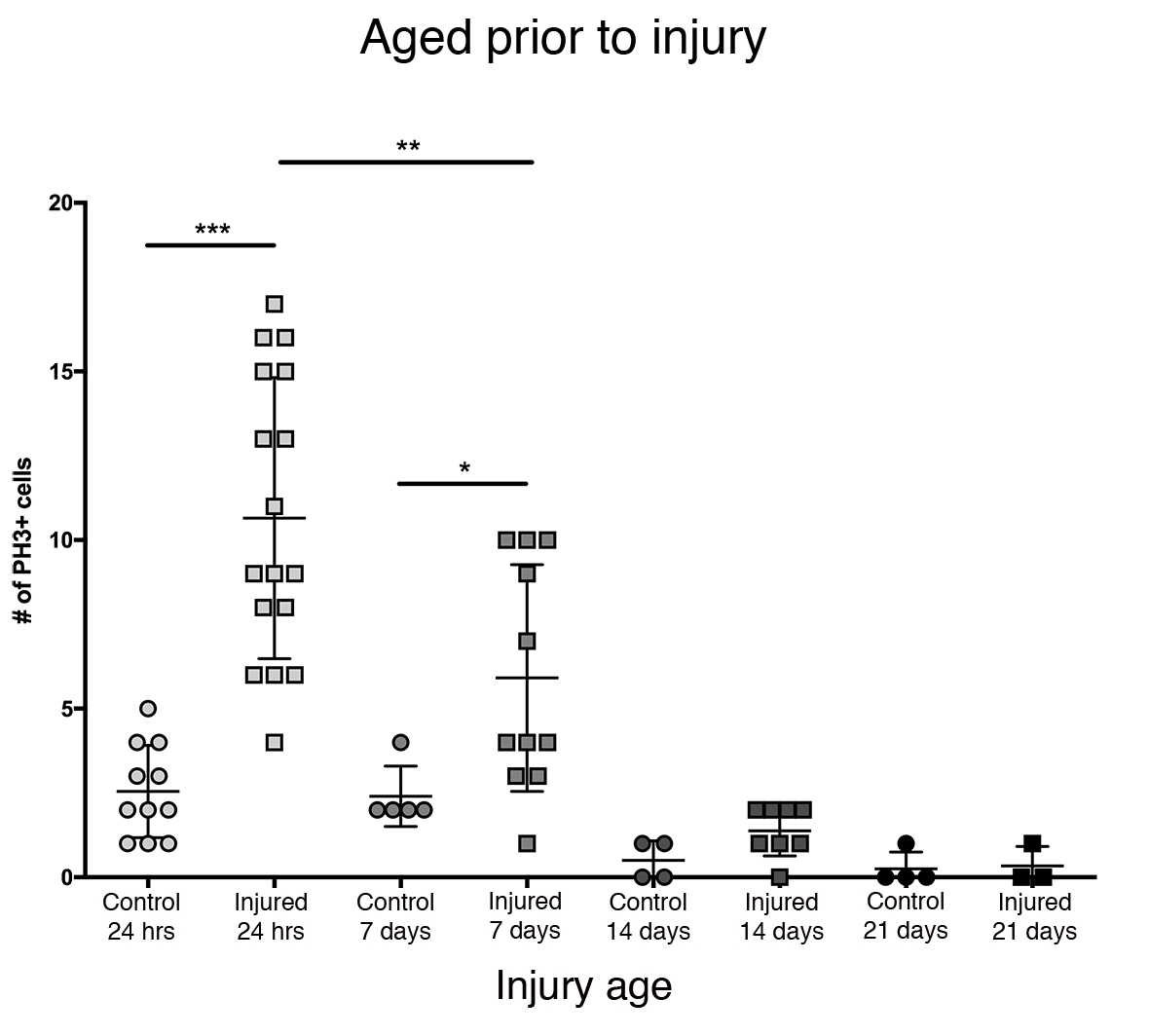
Die größte proliferative Reaktion im Zentralhirn wurde innerhalb der ersten 24 Stunden nach der Eklosion beobachtet (Abbildung 3). 7 Tage nach der Eklosion verursacht eine penetrierende Verletzung immer noch einen signifikanten Anstieg der Proliferation mit durchschnittlich 6 PH3 + -Zellen pro zentralem Gehirn. Dennoch nimmt die Fähigkeit der Zellen, sich nach ptBI zu teilen, 14 Tage nach der Eklosion signifikant auf 1 sich teilende Zelle ab, ähnlich wie bei Kontrollgehirnen (Abbildung 3). Somit ist das Potenzial für die Zellproliferation nach PTBI altersabhängig.
Neu erstellte Neuronen können projizieren, um Zielbereiche zu korrigieren
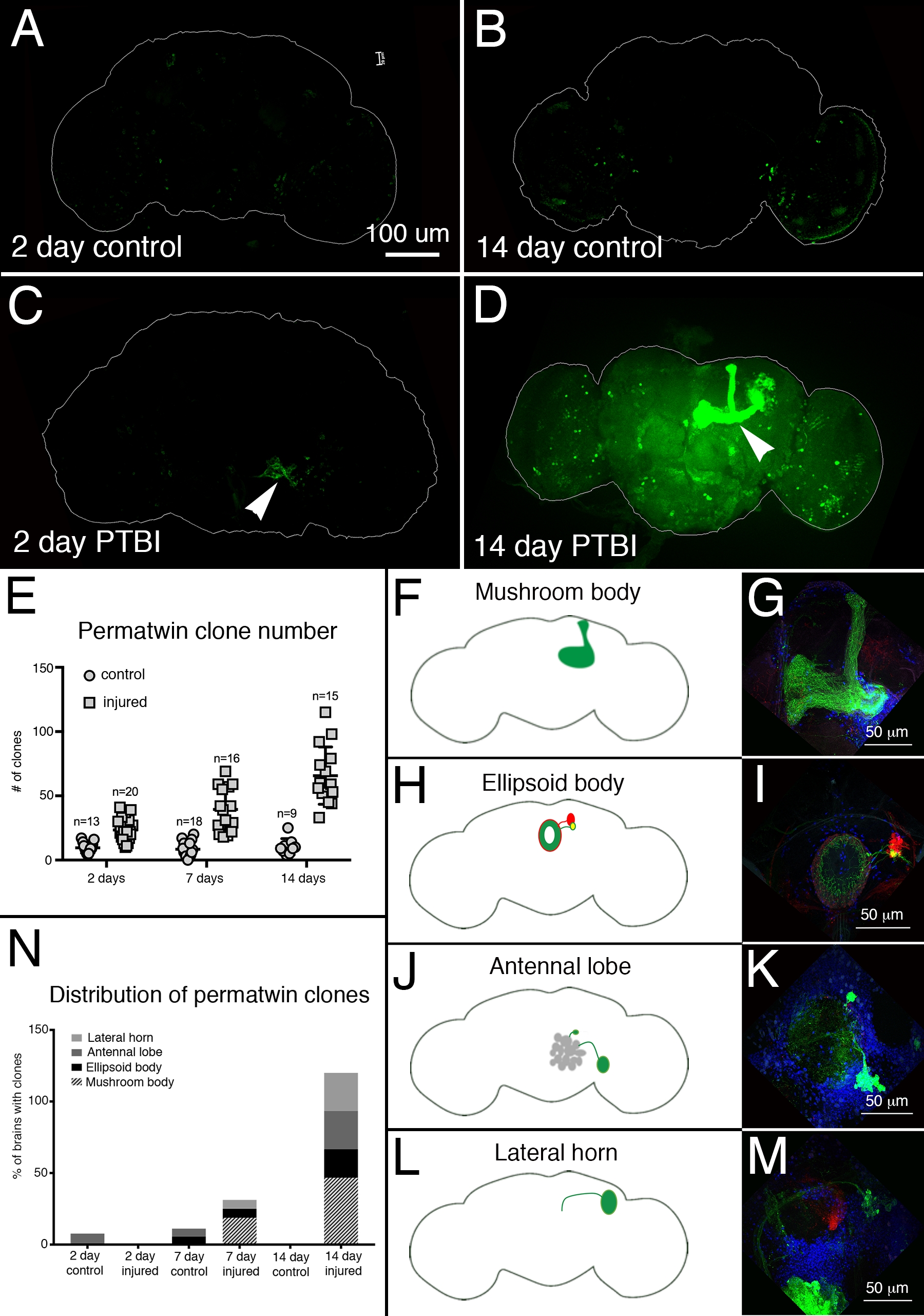
Zur Bewertung der neuronalen Regeneration nach PTBI wurde das Perma-Twin-Markierungssystem15 verwendet. Perma-Twin-Lineage Tracing markiert sich teilende Zellen und ihre Nachkommen dauerhaft mit einem grün fluoreszierenden Protein (GFP) oder rot fluoreszierenden Protein (RFP)15. In verletzten Proben wurden nach 2 Tagen und 2 Wochen mehr Perma-Zwillingsklone nachgewiesen als in Kontrollen (Abbildung 4A-E). Bemerkenswert ist, dass es 2 Wochen nach der Verletzung in ~ 50% der PTBI-Gehirne neue Pilzkörperneuronen gab (Abbildung 4N). Diese neuen Neuronen projizierten ihre Dendriten entsprechend auf den Pilzkörperkelch und ihre Axone entsprechend auf die Pilzkörperlappen (Abbildung 4D,F,G). Dies deutet darauf hin, dass die neu geschaffenen Zellen funktionelle Neuronen sein können, die an der Reparatur der beschädigten Pilzkörper beteiligt sind. Andere Bereiche des Gehirns, die sich zu regenerieren schienen, sind der Ellipsoidkörper (EB) (Abbildung 4H, I), die Antennenlappen (AL) (Abbildung 4J, K) und das Lateralhorn (LH) (Abbildung 4L, M), die etwa 26%, 26% bzw. 20% der Zeit große Klone besaßen (Abbildung 4N). Diese Ergebnisse unterstreichen den Nutzen dieses Systems für die Untersuchung der adulten Neurogenese. Ein vorgeschlagenes Modell für die Abfolge von Ereignissen nach PTBI, die zur Erzeugung neuer Neuronen führen, ist in Abbildung 5 dargestellt.

Abbildung 1: Penetrating Traumatic Brain Injury (PTBI) am erwachsenen Drosophila-Zentralhirn . (A) Schematische Darstellung des Äußeren eines erwachsenen Fliegenkopfes. Dies ist eine Frontalansicht. Somit befindet sich die rechte Seite des Tieres links vom Betrachter. (B) Schematische Darstellung des Inneren eines erwachsenen Drosophila-Kopfes mit der grau markierten Verletzungsbahn. Dies ist eine nachträgliche Ansicht. So befindet sich in diesem Bild und den nachfolgenden Abbildungen die rechte Seite des Gehirns rechts. Das zentrale Gehirn PTBI beeinflusst mehrere Gehirnstrukturen, einschließlich des Pilzkörpers (grün) und Gewebe außerhalb des Gehirns, einschließlich des Fettkörpers (blau) und der Hämozyten (rot). CB = zentrale Hirnregion. OL= optische Lappenregion. (C) Dorsale Ansicht eines lebenden erwachsenen Kopfes, in dem Pilzkörper (Pfeilspitzen) mit einem grün fluoreszierenden Protein (GFP) markiert sind. Dies ist der "Standard-Genotyp" (siehe Text für Details). Das PTBI-Protokoll führt reproduzierbar zu Verletzungen der Pilzkörper. Diese Abbildung wurde von Reference16 übernommen. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 2: PTBI stimuliert die Zellproliferation. Schaltpläne für die unbeletzte Kontrolle (A) und PTBI (B). Die blauen Kästchen in den oberen rechten Ecken zeigen die Gehirnregionen an, die bei höherer Vergrößerung in den Panels (C) und (D) gezeigt werden. (C,D) PH3-Antikörper (rot) wurde verwendet, um die Zellproliferation 24 h nach der Verletzung zu untersuchen. In Kontrollgehirnen (C) gibt es nur wenige PH3 + -Zellen und keine in der Nähe des MB. In PTBI-Gehirnen (D) gibt es jedoch PH3 + -Zellen in der Nähe des MB. (E) Quantifizierung proliferierender Zellen. Die Zahlen spiegeln wuchernde Zellen im gesamten Gehirn wider, nicht nur in der Nähe des Pilzkörpers. Bei 24 h hatten unverletzte Kontrollgehirne durchschnittlich 3 PH3 + -Zellen / Gehirn (n = 11 Gehirne, 28 Zellen), während 24 h nach PTBI Gehirne durchschnittlich 11 PH3 + -Zellen / Gehirn hatten (n = 17 Gehirne, 181 Zellen). Nach 7 Tagen haben unverletzte Kontrollen nur wenige EdU + -Zellen mit durchschnittlich 2 EdU + -Zellen / Gehirn (n = 15 Gehirne, 24 Zellen), während 7-Tage-Post-PTBI-Gehirne durchschnittlich 11 EdU + -Zellen / Gehirn hatten (n = 22 Gehirne, 238 Zellen). Nach 14 Tagen haben unverletzte Kontrollen durchschnittlich 1 EdU + -Zelle / Gehirn (n = 8 Gehirne, 11 Zellen), während 14-Tage-Post-PTBI-Gehirne durchschnittlich 29 EdU + -Zellen / Gehirn haben (n = 14 Gehirne, 400 Zellen). Für diese Reihe von Experimenten wurden junge erwachsene Männchen innerhalb von 6 h eklosion verwendet. Ungepaarten t-Tests von Kontroll- und PTBI-Proben an den 3-Zeitpunkten ergeben Werte von p<0,0001, p<0,0001 bzw<0,0002. Fehlerindikatoren spiegeln die Standardabweichung (SD) wider. Diese Abbildung wurde von Reference16 übernommen. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 3: Die proliferative Reaktion auf PTBI nimmt mit zunehmendem Alter ab. Um zu untersuchen, ob das Alter die Menge der Zellproliferation nach der Verletzung beeinflusst, wurden neu ausgeschiedene erwachsene Männer mit Tieren im Alter von 7 Tagen, 14 Tagen und 28 Tagen vor PTBI verglichen, wobei Anti-PH3 verwendet wurde, um die Zellproliferation 24 h nach der Verletzung zu untersuchen. Fliegen, die innerhalb von 6 h nach der Eklosion verletzt wurden, hatten durchschnittlich 11 PH3 + -Zellen / Gehirn (n = 17 Gehirne, 182 Zellen) im Vergleich zu durchschnittlich 3 PH3 + -Zellen / Gehirn in altersangepassten Kontrollen (n = 11 Gehirne, 28 Zellen). Fliegen im Alter von bis zu 7 Tagen, dann PTBI unterzogen, hatten durchschnittlich 6 PH3 + -Zellen / Gehirn (n = 11 Gehirne, 65 Zellen) im Vergleich zu altersgerechten Kontrollen mit durchschnittlich 2 PH3 + -Zellen / Gehirn (n = 5 Gehirne, 12 Zellen). Wenn Fliegen auf 14 Tage vor PTBI gealtert und 24 h später untersucht wurden, gab es einen Durchschnitt von 1 PH3 + -Zelle / Gehirn (n = 8 Gehirne, 11 Zellen), ähnlich wie altersangepasste Kontrollen, die auch durchschnittlich 1 PH3 + -Zelle / Gehirn (n = 4 Gehirne, 2 Zellen) betrugen. 28-tägige unverletzte Kontrolle (n = 4, 1 Zelle) und PTBI (n = 3, 1 Zelle) Fliegen hatten beide durchschnittlich 0 PH3 + Zellen / Gehirn. Ungepaarte t-Tests für PTBI zur Kontrolle von Vergleichen zu diesen 4-Zeitpunkten sind p<0,0001, p<0,04, p<0,07 bzw<0,84. Diese Abbildung wurde von Reference16 übernommen. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 4: Die Perma-Zwillings-Linienverfolgung zeigt die Regeneration des Gehirns und eine angemessene Ausrichtung von Axonen nach PTBI. Das Perma-Twin-Lineage-Tracing-System15 wurde verwendet, um die Neurogenese nach PTBI zu analysieren. Dieses System markiert sich teilende Zellen und Nachkommen dauerhaft mit einem grün fluoreszierenden Protein (GFP) oder einem rot fluoreszierenden Protein (RFP). Fliegen wurden bei 17 °C aufgezogen, um das System während der Entwicklung fernzuhalten. F1-Männchen, die Perma-Zwillings-Transgene trugen, wurden bei der Eklosion gesammelt, dann verletzt und bei 30 ° C platziert, um sich für 2 oder 14 Tage zu erholen. (A) Bei 2-tägigen unverletzten Kontrollen gibt es einige GFP+-Zellen, die im gesamten Gehirn verstreut sind. (B) Nach 14 Tagen sind relativ wenige GFP+-Zellen im zentralen Kontrollhirn vorhanden. (C) Im Vergleich dazu haben 2-Tage-verletzte Gehirne mehr GFP+-Zellen, die dazu neigen, sich in der Nähe der Verletzung zu gruppieren (Pfeilspitze). (D) 14 Tage nach der Verletzung befinden sich große Klone in der Nähe der Verletzungsstelle. Einige dieser Klone haben Axone, die entlang der Pilzkörperbahnen (Pfeilspitze) ragen. Hier wird nur der GFP-Kanal gezeigt; es gab ähnliche RFP+-Klone in den PTBI-Proben. (E) Die Anzahl der Klone nimmt im Laufe der Zeit nach PTBI zu.Kontroll-unverletzte Gehirne (n = 13) haben durchschnittlich 10 Klone nach 2 Tagen, während 2-Tage-PTBI-Gehirne (n = 20) durchschnittlich 23 Klone (p<0,00002) haben. Nach 7 Tagen hatten Kontrollgehirne durchschnittlich 9 Klone pro Gehirn (n = 18), während 7-Tage-PTBI-Gehirne durchschnittlich 39 Klone pro Gehirn (n = 16) hatten (p-Wert<0,00000002). Dies ist deutlich mehr als die Anzahl der Klone, die 2 Tage nach der Verletzung beobachtet wurden (p-Wert<0,0009). In 14-Tage-Kontrollgehirnen gibt es durchschnittlich 10 Klone pro Gehirn, was sich nicht wesentlich von den 2-Tage- und 7-Tage-Kontrollen unterscheidet. 14 Tage nach PTBI gibt es jedoch durchschnittlich 66 GFP+-Klone, was deutlich mehr ist als entweder altersangepasste Kontrollen (p<0,0000003) oder 2-Tage-Post-PTBI-Gehirne (p-Wert<0,0001). (F-M) PTBI stimuliert die Klonbildung in mehreren Regionen des Gehirns. Panels auf der linken Seite sind Schemata von Gehirnregionen, in denen große Klone 14 Tage nach PTBI gefunden wurden (A, H, J, L). Tafeln auf der rechten Seite zeigen hohe Vergrößerungen repräsentativer Gehirne (G, I, K, M). Viele 14-Tage-Gehirne hatten Klone, die auf bestimmte Zielbereiche projiziert wurden. Dazu gehörten der Pilzkörper (MB) (F,G), der Ellipsoidkörper (EB) (H,I), der Antennenlappen (AL) (J,K) und das Seitenhorn (LH) (L,M). (N) Sowohl die Klonzahl als auch die Klongröße nehmen mit der Zeit nach der PTBI zu.Die Anteile der Hirnregionen mit großen Klonen wurden nach 2, 7 und 14 Tagen in Kontrollen und verletzten Gehirnen berechnet. Nach 2 Tagen zeigten ~ 8% der Kontrollgehirne (n = 13) AL-Klone, während es keine AL-Klone in 2-Tage-verletzten Gehirnen gab (n = 20). In 7-Tage-Kontrollgehirnen (n = 18) hatten 6% AL und 6% EB-Klone. 7 Tage nach PTBI (n = 16) hatten 6% der Gehirne auch AL-Klone, 6% hatten EB-Klone und 19% hatten große MB-Klone. Nach 14 Tagen zeigten Kontrollgehirne (n = 9) keine spezifischen Bereiche mit Klonen, während 47% der PTBI-Gehirne (n = 15) MB-Klone, 20% der PTBI-Gehirne AL-Klone und 27% der PTBI-Gehirne EB-Klone und 27% LH-Klone hatten. Diese Abbildung wurde von Reference16 übernommen. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

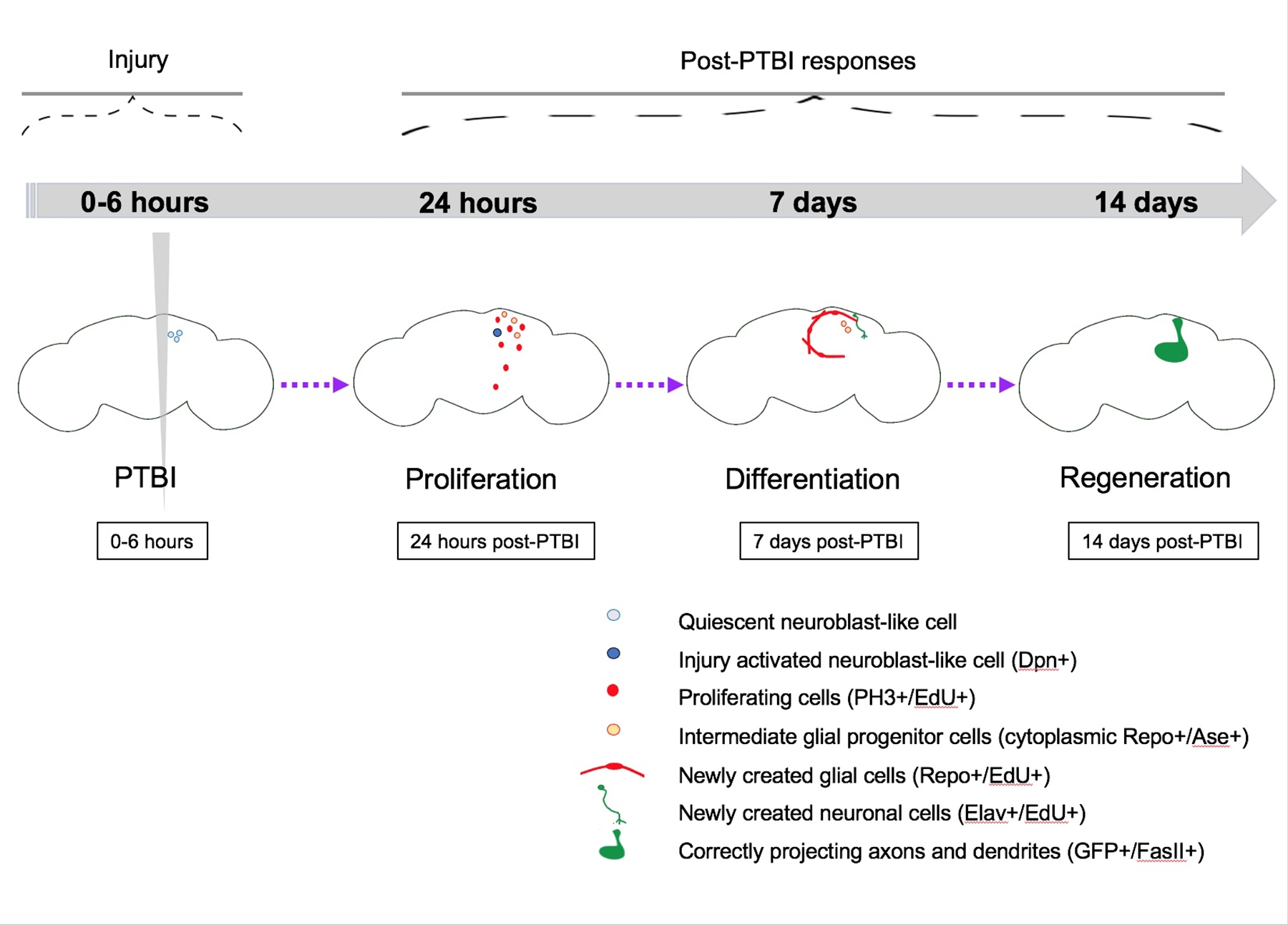
Abbildung 5: Zusammenfassendes Modell für die Regeneration nach penetrierender traumatischer Hirnverletzung (PTBI). Bei jungen erwachsenen Drosophila gibt es ruhende neuroblastenähnliche Zellen im zentralen Gehirn, denen die Expression kanonischer Neuroblastengene fehlt. Bis 24 h nach PTBI werden die ruhenden neuroblastenähnlichen Zellen aktiviert, exprimieren Neuroblastengene und beginnen sich zu vermehren. Sowohl 4 h als auch 24 h nach PTBI kommt es zu einer Welle des Zelltods16. Nach 7 Tagen ist die Proliferationsrate immer noch hoch, und viele der neuen Zellen haben reife Zellidentitäten angenommen und sind zu Neuronen oder Gliazellen geworden. 14 Tage nach der PTBI projizieren große Klone neuer Neuronen mit Axonen und Dendriten korrekt auf ihre jeweiligen Zielbereiche. Bewegungsmangel wird ebenfalls um 14 Tage wiederhergestellt, was darauf hindeutet, dass sich erwachsene Drosophila funktionell und strukturell regenerieren kann. Diese Abbildung wurde von Reference16 übernommen. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
Diskussion
Obwohl durchdringende Verletzungen des erwachsenen Drosophila-Gehirns bereits beschrieben wurden15,17,18, konzentrierten sich diese Verletzungen auf die Sehlappen und nicht auf das zentrale Gehirn. Weitere, detaillierte Anweisungen zur Durchführung der Verletzungen fehlen bisher. Dieses Protokoll beschreibt ein Modell für die penetrationsbedingte Verletzung des erwachsenen Drosophila-Zentralhirns, das statistisch signifikante Beweise für die adulte Neurogenese nach PTBI reproduziert.
Die Reproduzierbarkeit dieses PTBI-Protokolls ist zum Teil auf den Pilzkörper als Verletzungszielregion zurückzuführen. Der Pilzkörper ist groß und besteht aus ~2200 Neuronen mit komplexen Dendriten- und Axonlauben in großen und stark stereotypen Arrays18. Die Zellkörper von Pilzkörperneuronen liegen in der Nähe der Gehirnoberfläche und können durch die Kopfkutikula durch die Expression von grün fluoreszierendem Protein (GFP) visualisiert werden (Abbildung 1C). Pilz-Körpervorläufer sind die letzten neuralen Stammzellen, die während der Entwicklung apoptosiert werden13,12,19. Daher sind viele Pilzkörperneuronen zum Zeitpunkt der Eklosion ziemlich jung. Dies führte zu der Hypothese, dass der Pilzkörper mehr mitotisches Potenzial haben könnte als andere Hirnregionen16. Darüber hinaus ist der Pilzkörper entscheidend für das Lernen und das Gedächtnis18. Auf diese Weise kann man fragen, ob ptBI-ausgelöste Neurogenese zu einer funktionellen Erholung führt.
Andere Faktoren, die zur Reproduzierbarkeit der Ergebnisse beitragen, sind die Verwendung von ausgekreuzten Fliegen konsistenter Genotypen, die Durchführung von Kreuzungen in die gleiche Richtung jedes Mal, die genaue Kontrolle der Aufzucht- und Alterungstemperaturen und die separate Analyse von Männchen und Weibchen. Die Verwendung von F1-Fliegen aus einem Outcross reduziert die Wahrscheinlichkeit, homozygote Gehirne auf spontane Mutationen zu analysieren. Das Standardkreuz von y[1] w[1]; UAS-mCD8-GFP;; OK107-GAL4 adulte Weibchen zu y[1] w[1] adulten männlichen Fliegen führt zu F1 Nachkommen des Genotyps y[1] w[1]; UAS-mCD8-GFP/+;; OK107-GAL4/+. OK107-GAL4 wird in allen intrinsischen Neuronen des Pilzkörpers exprimiert und treibt die Expression des membrangebundenen Reporters UAS-mCD8-GFP an, der die Visualisierung von Pilzkörpern und deren Projektionen ermöglicht. Bei den Perma-Zwillingskreuzen müssen die Kreuze jederzeit bei 17 °C bleiben, um das Linienverfolgungssystem ausgeschaltet zu halten. Dies stellt sicher, dass während der Entwicklung keine sich teilenden Zellen markiert werden und dass nur adulte Neuronen und Glia markiert werden. Dazu kann auch der Fliegenraum bei 17 °C gehalten werden. Obwohl in der erstbeschreibung des Perma-Twin-Systems15 empfohlen wurde, Fliegen bei 18 °C aufzuziehen, kann dies zu einer signifikanten Hintergrundkennzeichnung führen.
Für die Konsistenz wird auch empfohlen, die Kontrolle unverletzte Fliegen auf dem CO2-Pad zu halten, während man die PTBI durchführt. Dies stellt sicher, dass beide Fliegengruppen eine identische anästhetische Exposition haben. Darüber hinaus ist es wünschenswert, dass die Reproduzierbarkeit den Kopf vollständig durchdringt. Es muss jedoch darauf geachtet werden, dass die Spitze des Stiftes nicht gegen das Pad gebogen wird, wodurch es für zukünftige Verletzungen unbrauchbar wird. Für erfahrene Praktiker gibt es wenig unbeabsichtigten Schaden für PTBI-Fliegen. Dennoch kann es tödlich sein, zu stark auf den Brustkorb zu drücken, um die Fliege während einer Verletzung zu stabilisieren. Eine Möglichkeit, das Ausmaß der unbeabsichtigten Verletzung zu beurteilen, besteht darin, die Mortalität von PTBI-Fliegen 24 h nach der Verletzung zu quantifizieren. Für einseitig verletzte Fliegen kann dies für Anfänger 50% oder höher sein. Um sicherzustellen, dass die beobachteten Ergebnisse auf PTBI und nicht auf unbeabsichtigte Verletzungen zurückzuführen sind, wird daher empfohlen, dass Anfänger die Verabreichung von PTBI an ~ 20 Fliegen täglich über mehrere Wochen üben und die resultierenden Gehirne nicht analysieren, bis die 24-Stunden-Mortalität konstant <10% beträgt.
Um die Menge der Proliferation zu quantifizieren, die durch PTBI im Zentralen Gehirn stimuliert wird, können sowohl Antiphosphohton H3 (PH3) Immunfärbung als auch 5-Ethynyl-2'-desoxyuridin (EdU) Einbau verwendet werden. Anti-PH3 markiert Zellen vor und während der Metaphase und beschränkt den Nachweis auf nur einen Bruchteil der sich aktiv teilenden Zellen. Somit bietet die Anti-PH3-Färbung nur einen teilweisen Einblick in die Proliferation. EdU ist ein Thymidin-Analogon, das in neu synthetisierte DNA eingebaut werden kann. Durch die Fütterung von Fliegen EdU vor und nach der Verletzung ist es möglich, ein vollständigeres Bild der Zellen zu erhalten, die sich nach der Verletzung entweder teilen oder geteilt haben. Die Tatsache, dass alle Zellen, die sich teilen, dauerhaft markiert sind, ist sowohl für die Identifizierung von langsam kreisenden Zellen als auch für den Test des Überlebens von Zellen nach der anfänglichen Proliferation hilfreich. Aus unklaren Gründen, aber vielleicht aufgrund der begrenzten Durchlässigkeit der Blut-Hirn-Schranke, ist die EdU-Markierung ineffizient und berichtet nicht über die Zellproliferation im erwachsenen Gehirn. Dies wird durch die ähnliche Anzahl von PH3+ und EdU+-Zellen sowohl im Kontroll- als auch im Experimentalgehirn 24 h nach PTBI belegt und durch die Beobachtung, dass nur eine Untergruppe neuer Zellen in Perma-Zwillings-Klonen EdU16 enthält. Für eine maximale Kennzeichnung ist es wichtig, die Fliegen mit EdU vorzufüttern, da verletzte Fliegen die Fütterung mehrere Stunden nach der PTBI nicht wieder aufnehmen. Die Fütterung wurde durch Zugabe von Lebensmittelfarbe zur EdU-Lösung und Überwachung der Farbstoffmenge im Darm durch die Bauchkutikula beurteilt16.
Es ist zu beachten, dass, während wir in Schritt 4 ein Gehirndissektionsprotokoll zur Verfügung gestellt haben, alternative Techniken verwendet werden können. Einige davon sind in zuvor veröffentlichten Protokollen verfügbar20,21,22. Drosophila melanogaster bietet ein kostengünstiges Modell mit leistungsstarken genetischen und molekularen Werkzeugen, mit denen die Mechanismen untersucht werden können, die der Regeneration mehrerer Gewebe, einschließlich des Darms und der Komponenten des Nervensystems, zugrunde liegen. Ein neuartiges und reproduzierbares Verletzungsmodell, mit dem die Reaktion auf Hirnverletzungen untersucht werden kann, wird hier beschrieben. Daten, die mit diesen Protokollen gewonnen werden, unterstützen die Idee, dass das erwachsene Drosophila-Zentralgehirn die proliferative Fähigkeit beibehält und als Reaktion auf Verletzungen neue Neuronen erzeugt. Diese Beobachtungen rechtfertigen eine weitere Untersuchung sowohl des Ausmaßes der adulten Neurogenese als auch der zugrunde liegenden molekularen Mechanismen. Sobald die Komponenten, die an der neuronalen Regeneration beteiligt sind, in diesem System identifiziert sind, können wir unser Wissen über die adulte Drosophila-Neurogenese auf den Menschen übertragen.
Offenlegungen
Die Autoren haben keine Interessenkonflikte.
Danksagungen
Wir danken Stacey Rimkus und Becky Katzenberger für die technische Unterstützung und Eduardo Moreno für die gemeinsame Nutzung der Perma-Twin-Bestände. Wir möchten Barry Ganetzky und David Wassarman für lebhafte Diskussionen danken, die zweifellos die Wissenschaft verbessert haben, und Kent Mok, Cayla Guerra und Bailey Spiegelberg für ihre Beiträge zum Labor. Die FasII-Antikörper wurden von Corey Goodman entwickelt und von der Developmental Studies Hybridoma Bank gewonnen, die vom NICHD des NIH erstellt und an der University of Iowa, Department of Biology, Iowa City, IA 52242, gepflegt wird. Die meisten der in dieser Studie verwendeten Drosophila-Stämme wurden vom Bloomington Drosophila Stock Center (BDSC; NIH P40OD018537). Diese Arbeit wurde von NIH T32 GM007133 (KLC) unterstützt; NIH NS090190 (GBF); NIH NS102698 (GBF); die University of Wisconsin Graduate School (GBF); und das UW-Madison Women in Science and Engineering Leadership Institute (WISELI) (GBF).
Materialien
| Name | Company | Catalog Number | Comments |
| #11 disposable scalpels | Santa Cruz Biotechnology | sc-395923 | used for separating Drosophila heads from trunks prior to brain dissection |
| 150 mm diameter black Sylgard dishes | Dow | 1696157 | made in the laboratory with reagents from Dow; used for brain dissection |
| 18 mm coverslips | any | for mounting brains on microscope slides | |
| 4',6-Diamidino-2-Phenylindole, Dihydrochloride (DAPI) | ThermoFisher | D1306 | for immunohistochemistry |
| 70% Ethanol | made from 95% ethanol sourced variously | ||
| anti-mouse Cy5 | Jackson ImmunoResearch | 715-175-151 | for immunohistochemistry |
| anti-rabbit 568 | ThermoFisher | A11036 | for immunohistochemistry |
| bovine serum albumin (BSA) | SIgma Aldrich | A7030 | for immunohistochemisty |
| Clear nail polish | any | for sealing coverslips | |
| Click-It EdU labeling kit | InVitrogen | C10640 | to detect newly synthesized DNA |
| CO2 bubbler | Genesee Scientific | 59-181 | for anesthesia |
| CO2 pad | Genesee Scientific | 59-114 | for anesthesia |
| CO2 regulator and supply | any | for anesthesia | |
| Confocal microscope | any | for imaging fixed, stained and mounted brains | |
| cotton plugs | Genesee Scientific | 51-101 | for EdU labeling |
| Drosophila vials | Genesee Scientific | 32-109 | for EdU labeling |
| Fix buffer (Pipes, EGTA, Magnesium; PEM) | components sourced from various companies | for fixing adult brains; 100 mM piperazine-N,N’-bis(2-ethanesulfonic acid) [PIPES], 1 mM EGTA, 1 mM MgSO4, pH 7.0 | |
| Formaldehyde | Sigma Aldrich | 252549 | for fixing adult brains, added to PEM |
| Grade 3 round Whatman filters, 23 mm round | Tisch Scientific | 1003-323 | for EdU labeling |
| Microfuge tubes | any | for fixing and staining reactions and for storing Minutien pins | |
| Microscope slides | any | for mounting brains | |
| Minutien pins | Fine Science Tools | 26002-10 | for brain injury; 12.5 μm diameter tip and 100 μm diameter rod |
| mouse anti-Fasiclin II | Developmental Studies Hybridoma Bank | 1D4-s | for immunohistochemistry |
| NIGHTSEA stereo microscope fluorescence adaptor | Electron Microscopy Sciences | SFA-GR | fluorescence setup for dissecting microscope |
| P20, P200 and P1000 pipettors and tips | any | for measuring solutions | |
| phosphate buffered saliine (PBS) | components sourced from various companies | for dissecting brains and making immunohistochemistry blocking and washing solutions; 100 mM of K2HPO4, 140 mM of NaCl, pH 7.0 | |
| phosphate buffered saline with 0.1% Triton X-100 (PT) | components sourced from various companies | for washing dissected brains | |
| phosphate buffered saline with 0.1% Triton X-100 + 2% bovine serum albumin (PBT) | components sourced from various companies | blocking solution for immunohistochemistry and for diluting antibodies | |
| rabbit anti-PH3 | Santa Cruz Biotechnology, Inc | sc-8656-R | for immunohistochemistry |
| Reinforcement labels | Avery | 5721 | to maintain space between the microscope slide and the coverslip |
| Size 0 paintbrushes | any | to manipulate and stabilize adult Drosophila during injury | |
| Triton X-100 | Sigma Aldrich | 93443 | |
| Two pair of #5 watchmakers forceps | Fine Science Tools | 11255-20 | used to hold the Minutien pins and for brain dissections |
| Vectashield | Vector Laboratories | H-1000 | mounting medium for microscope slides |
Referenzen
- Thurman, D., et al. Report to Congress: Traumatic brain injury in the United States. , Available from: https://www.cdc.gov/traumaticbraininjury/pubs/tbi_report_to_congress.html (1999).
- NIEHS. , Available from: https://www.nih.gov/research/supported/health/neurodegenerative/index.cfm (2021).
- Morton, N. V., Wehman, P. Psychosocial and emotional sequelae of individuals with traumatic brain injury: A literature review and recommendations. Brain Injury. 9 (1), 81-92 (2017).
- Bonini, N. M., Berger, S. L. The sustained impact of model organisms-in genetics and epigenetics. Genetics. 205, 1-4 (2017).
- Brace, E. J., DiAntonio, A. Models of axon regeneration in Drosophila. Experimental Neurology. 287, Pt 3 310-317 (2017).
- Hao, Y., Collins, C. Intrinsic mechanisms for axon regeneration: insights from injured axons in Drosophila. Current Opinion in Genetics & Development. 44, 84-91 (2017).
- Chiang, A. S., et al. Three-dimensional reconstruction of brain-wide wiring networks in Drosophila at single-cell resolution. Current Biology. 21 (1), 1-11 (2011).
- Meinertzhagen, I. A. The organisation of invertebrate brains: cells, synapses and circuits. Acta Zoologica. 91 (1), 64-71 (2010).
- Bellen, H. J., Tong, C., Tsuda, H. 100 years of Drosophila research and its impact on vertebrate neuroscience: A history lesson for the future. Nature Reviews Neuroscience. 11 (7), 514-522 (2010).
- Lessing, D., Bonini, N. M. Maintaining the brain: insight into human neurodegeneration from Drosophila melanogaster mutants. Nature Reviews Genetics. 10 (6), 359-370 (2009).
- Boone, J. Q., Doe, C. Q. Identification of Drosophila type II neuroblast lineages containing transit amplifying ganglion mother cells. Developmental Neurobiology. 68 (9), 1185-1195 (2008).
- Ito, K., Hotta, Y. Proliferation pattern of postembryonic neuroblasts in the brain of Drosophila melanogaster. Developmental Biology. 149 (1), 134-148 (1992).
- Siegrist, S. E., Haque, N. S., Chen, C. H., Hay, B. A., Hariharan, I. K. Inactivation of both Foxo and reaper promotes long-term adult neurogenesis in Drosophila. Current Biology. 20 (7), 643-648 (2010).
- von Trotha, J. W., Egger, B., Brand, A. H. Cell proliferation in the Drosophila adult brain revealed by clonal analysis and bromodeoxyuridine labelling. Neural Development. 4, 9(2009).
- Fernandez-Hernandez, I., Rhiner, C., Moreno, E. Adult neurogenesis in Drosophila. Cell Reports. 3 (6), 1857-1865 (2013).
- Crocker, K. L., et al. Neurogenesis in the adult Drosophila brain. Genetics. , (2021).
- Plavicki, J., Mader, S., Pueschel, E., Peebles, P., Boekhoff-Falk, G. Homeobox gene distal-less is required for neuronal differentiation and neurite outgrowth in the Drosophila olfactory system. Proceedings of the National Academy of Sciences of the United States of America. 109 (5), 1578-1583 (2012).
- Aso, Y. The neuronal architecture of the mushroom body provides a logic for associative learning. Elife. 3, 04577(2014).
- Ito, K., Awano, W., Suzuki, K., Hiromi, Y., Yamamoto, D. The Drosophila mushroom body is a quadruple structure of clonal units each of which contains a virtually identical set of neurones and glial cells. Development. 124 (4), 761-771 (1997).
- Tito, A. J., Cheema, S., Jiang, M., Zhang, S. A. Simple one-step dissection protocol for whole-mount preparation of adult Drosophila brains. Journal of Visualized Experiments. (118), e55128(2016).
- Kelly, S. M., Elchert, A., Kahl, M. Dissection and immunofluorescent staining of mushroom body and photoreceptor neurons in adult Drosophila melanogaster brains. Journal of Visualized Experiments. (129), e56174(2017).
- Arain, U., Valentino, P., Islam, I. M., Erclik, T. Dissection, immunohistochemistry and mounting of larval and adult Drosophila brains for optic lobe visualization. Journal of Visualized Experiments. (170), e61273(2021).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenWeitere Artikel entdecken
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten