
Method Article
Cortar el tronco cerebral auditivo embrionario del pollo para evaluar gradientes tonotópicos y microcircuitos
En este artículo
Resumen
Aquí presentamos un protocolo para obtener cortes auditivos no coronales del tronco encefálico del embrión de pollo para la investigación de propiedades tonotópicas y trayectorias de desarrollo dentro de un corte del tronco encefálico. Estos cortes incluyen secciones sagitales, horizontales y horizontales / transversales que abarcan regiones tonotópicas más grandes dentro de un plano de corte individual que las secciones coronales tradicionales.
Resumen
El embrión de pollo es un modelo animal ampliamente aceptado para estudiar el tronco cerebral auditivo, compuesto por microcircuitos altamente especializados y topología neuronal orientada diferencialmente a lo largo de un eje tonotópico (es decir, frecuencia). El eje tonotópico permite la codificación segregada de sonidos de alta frecuencia en el plano rostral-medial y la codificación de baja frecuencia en regiones caudo-laterales. Tradicionalmente, las cortes coronales del tronco encefálico de tejido embrionario permiten el estudio de láminas de isofrecuencia individuales relativas. Aunque es suficiente para investigar cuestiones anatómicas y fisiológicas relacionadas con regiones individuales de isofrecuencia, el estudio de la variación tonotópica y su desarrollo en áreas auditivas más grandes del tronco encefálico es algo limitado. Este protocolo informa técnicas de corte del tronco encefálico de embriones de pollo que abarcan gradientes más grandes de regiones de frecuencia en el tronco cerebral auditivo inferior. La utilización de diferentes métodos de corte para el tejido auditivo del tronco encefálico del pollo permite experimentos electrofisiológicos y anatómicos dentro de un corte del tronco encefálico, donde los gradientes más grandes de propiedades tonotópicas y trayectorias de desarrollo se conservan mejor que las secciones coronales. Múltiples técnicas de corte permiten una mejor investigación de las diversas propiedades anatómicas, biofísicas y tonotópicas de los microcircuitos auditivos del tronco encefálico.
Introducción
El embrión de pollo es un valioso modelo de investigación para estudiar cuestiones biológicas básicas en numerosas y diversas áreas científicas, incluyendo biología celular, inmunología, patología y neurobiología del desarrollo. El microcircuito del tronco encefálico auditivo del pollo es un excelente ejemplo de un circuito altamente especializado que puede entenderse en términos de morfología y fisiología auditiva. Por ejemplo, Rubel y Parks (1975) describieron por primera vez la orientación tonotópica (es decir, gradiente de frecuencia) del núcleo magnocelular del pollo (NM) y el núcleo laminar (NL) como una función lineal a través del eje de los núcleos, orientada ~ 30 ° con respecto al plano sagital. Las neuronas individuales en NM y NL codifican su mejor frecuencia de sonido, conocida como su frecuencia característica (CF), a lo largo del plano rostral-medial hasta la región caudo-lateral. Las neuronas sensibles a la alta frecuencia se encuentran en la región rostral-medial y las neuronas sensibles a la baja frecuencia se encuentran caudolateralmente. Como tal, los métodos tradicionales de disección del tejido auditivo del tronco encefálico para estudiar las propiedades tonotópicas han utilizado cortes coronales sucesivos. De hecho, los microcircuitos auditivos de embriones de pollo en desarrollo se han establecido como un sistema modelo para estudiar el procesamiento de señales de las funciones auditivas tonotópicas a través de sucesivas cortes del tronco encefálico del plano coronal caudal a rostral durante décadas 1,2,3,4,5,6.
Sin embargo, la organización tonotópica de NM y NL es topológica y morfológicamente complicada. Las entradas nerviosas auditivas se distribuyen de tal manera que las altas entradas de CF terminan en estructuras similares a bulbos terminales que cubren al menos una cuarta parte de la circunferencia somática de una célula NM adendrítica. Por el contrario, las entradas bajas de CF no están organizadas con terminales finales similares a bulbos, sino con múltiples sinapsis de bouton en dendritas de neuronas NM. Las entradas de CF medio terminan como sinapsisde bulbo final y bouton 4,7,8,9,10,11,12. En NL, el gradiente dendrítico altamente estereotipado es evidente no solo en la longitud dendrítica sino también en el ancho dendrítico. Este gradiente dendrítico único se ajusta estrechamente al eje tonotópico. Las dendritas experimentan un aumento de 11 veces en longitud y cinco veces en anchura de neuronas de alta a baja CF, respectivamente6. Para superar tales distribuciones complicadas de estos núcleos en cortes coronales, este protocolo describe enfoques de disección en los planos sagital, horizontal y horizontal/transversal. Estas técnicas de corte proporcionan ejemplos de tejido auditivo del tronco encefálico que exhiben propiedades tonotópicas máximas en un plano de corte individual.
Protocolo
Todos los procedimientos fueron aprobados por los Comités Institucionales de Cuidado y Uso de Animales de la Universidad Northwestern (IACUC) y se llevaron a cabo de acuerdo con las Pautas de los Institutos Nacionales de Salud para el Cuidado y Uso de Animales de Laboratorio. Los protocolos para la disección y preparación del tejido del tronco encefálico están en adhesión a los protocolos anteriores 5,13.
1. Manipulación de huevos
- Compre huevos fertilizados (Gallus gallus domesticus) de un proveedor local de animales aprobado por IACUC.
- Conservar los huevos inmediatamente después de su llegada en nevera a 14 °C e incubar en un plazo de 5 días.
NOTA: La viabilidad embrionaria disminuye considerablemente después de 1 semana. - Esterilizar los huevos con etanol al 70% antes de la incubación a 38 ± 1 °C y ~50% de humedad.
2. Composición y preparación del líquido cefalorraquídeo artificial (ACSF)
- Mezclar los siguientes productos químicos en 1 L de 18,2 MΩcmdH2Opara crear una solución madre de ACSF 10x: NaCl (cloruro de sodio) 130 mM, NaHCO3 (bicarbonato de sodio) 26 mM, KCl (cloruro de potasio) 2,5 mM, NaH2PO4 (dihidrogenofosfato de sodio) 1,25 mM, dextrosa (D-(+)-glucosa) 10 mM. Mantenga la solución madre en el refrigerador.
- Preparar las soluciones de MgCl 2 (cloruro de magnesio) 1 M y CaCl 2 (cloruro de calcio) 1 M por separado en 18,2 MΩcm dH2Oy conservar en la nevera.
- Inmediatamente antes de usar, diluir 10x ACSF a 1x y burbujear continuamente con 95% O 2/5% CO 2 durante 15-20 min y agregar MgCl 2 y CaCl 2. Para preparar ACSF y dACSF (disecante de ACSF), ajustar a una concentración final de Mg 2+ 1 mM, Ca 2+ 3 mM y Mg 2+ 3 mM, Ca 2+ 1 mM, respectivamente.
- Establezca la velocidad de burbujeo para el ACSF de modo que el pH sea 7.2-7.4 con una osmolalidad entre 300 y 310 mOsm / L.
NOTA: Tener el ACSF colocado en un baño de hielo mientras burbujea es beneficioso para mantener una temperatura baja de la solución, lo que apoyará la integridad estructural del tejido durante la disección.
3. Preparación del bloque de agarosa (5%)
- Mezclar 5 g de agarosa en 100 ml de dACSF. Use un baño de agua a 100 ° C o un microondas durante 2-3 minutos, revolviendo cada 30 s para evitar grumos hasta que la agarosa se disuelva completamente y comience a burbujear.
- Vierta la agarosa derretida en una placa de Petri vacía de hasta 5 mm de espesor y manténgala a temperatura ambiente para que se fije. Después del cuajado, sellar la placa de Petri con parafilm y conservar a 4 °C.
- Corte la agarosa en bloques cúbicos con una cuchilla afilada y úselos en el momento de la disección.
4. Protocolo de disección y aislamiento auditivo del tronco encefálico
- Limpie el área de disección con un aerosol de solución de alcohol etílico al 70%.
- Pegue el bloque de agarosa de soporte o en ángulo en la bandeja de vibratomo.
- Elija huevos de la edad deseada (E20 y E21 en el presente protocolo). Manipule e incube los huevos siguiendo los protocolos enumerados anteriormente como en el paso 1.
- Localice el espacio lleno de aire colocando el huevo bajo una luz brillante (vela de araña) y buscando este espacio en el lado más grande o más redondo del huevo.
- Aclimate los huevos a temperatura ambiente, agriete la cáscara sobre el espacio lleno de aire y exponga el saco de membrana.
- Haga una incisión suave en el saco para exponer el pico.
- Con un bisturí, tire suavemente del cuello y la cabeza fuera del huevo.
- Decapitar rápidamente la cabeza con unas tijeras afiladas.
- Después de la decapitación, limpie la cabeza con dACSF helada para eliminar el exceso de sangre de la almohadilla de disección.
- Mantenga la cabeza firme en dACSF helada y haga una incisión rostro-caudal. Comience la incisión detrás y entre los ojos y siga la longitud del cuello cosechado.
NOTA: Los embriones más jóvenes pueden requerir menos presión al hacer la incisión. - Separe la piel para exponer el cráneo.
- Corte el cráneo detrás del ojo en dirección media a lateral. Haz esto para ambos hemisferios.
NOTA: Este paso ayuda a separar la porción rostral del cráneo del cerebro adjunto mientras mantiene intacto el tejido cerebral5. - Corte la porción rostral del cráneo. Coloque la cuchilla detrás de los ojos y haga un corte rápido.
NOTA: Es posible que se requiera un esfuerzo para cortar el cráneo adjunto limpiamente. - Sumerja la cabeza en un plato de dACSF frío.
- Utilizando un pequeño par de tijeras, haga incisiones de línea media a lateral en la región caudal del cráneo para tratar de separar el cerebro del cráneo sin causar daño tisular.
- Exponga suavemente el tronco encefálico y el cerebelo. Retraiga el área dorsal de todo el cráneo, retire el tronco encefálico con cuidado y expóngalo con la ayuda de un pincel fino con trineo suave. Use fórceps curvos para limpiar el tronco encefálico de la conexión de tejidos y vasos sanguíneos. Preste especial atención a la8ª área del nervio craneal y asegúrese de dejar una longitud corta de fibras nerviosas intactas en ambos lados.
- Separe el tronco encefálico del cerebelo cortando los pedúnculos y retirando los vasos sanguíneos con cuidado. Recorte el tronco encefálico de vasos sanguíneos adicionales.
NOTA: Asegúrese de que todo el procedimiento se realiza en dACSF refrigerado con hielo burbujeado continuamente con carboxi (95% O 2/5% CO2).
5. Rebanado de vibratorio
NOTA: En los siguientes pasos, la parte posterior del tejido debe apoyarse con una pieza cúbica de agarosa.
- Coloque la cuchilla de vibratomo a lo largo del eje horizontal y pegue el tronco encefálico en una bandeja de corte. Pegue el lado rostral, manteniendo el eje rostral-caudal vertical para cortes coronales.
- Mantenga el eje lateral-medial vertical para cortes sagitales.
- Pegue el lado ventral, manteniendo el eje dorsal-ventral vertical para rodajas horizontales.
- Para lograr el plano sagital-horizontal angular agudo, pegue el lado ventral del tronco encefálico, manteniendo el eje ventral-dorsal vertical en la superficie de hipotenusa del bloque de agarosa, que se corta en un ángulo de 45 °. Pegue la superficie opuesta del bloque de agarosa frente a la bandeja de corte y mantenga el eje rostral-caudal paralelo al borde de la cuchilla.
6. Manipulación de trozos frágiles o grandes de tejido del tronco encefálico
- En un enfoque alternativo al paso 5, sumerja el tronco encefálico aislado en agarosa de bajo punto de fusión (LMP) al 4% a ~ 40 ° C en una placa de Petri de 35 mm x 10 mm.
- Después de verter agarosa en el tronco encefálico sumergido, coloque la placa de Petri en hielo para solidificar. Corte el bloque de agarosa cúbico con tronco encefálico incrustado con una cuchilla de afeitar afilada.
- Pegue el bloque de agarosa LMP en su lado rostral, manteniendo vertical el eje rostral-caudal del tronco encefálico.
- Tome cortes coronales hasta que se pueda visualizar la región NM.
- Retire el bloque de agarosa del pegamento con una cuchilla afilada. Para detectar los núcleos, coloque suavemente 0.5 μL de colorante (azul de toluidina o naranja G) en el NM con una aguja fina.
- Vuelva a montar este bloque en la bandeja de corte para rodajas sagitales u horizontales e identifique los núcleos con respecto a la región teñida.
- Para obtener el mejor rendimiento, ajuste la velocidad de corte del vibratomo en 4 - 5 (~30 ± 4 mm/min), la frecuencia de vibración a 85-87 Hz y la amplitud de corte a 4-6 (~1 ± 0,2 mm).
- Después de la sección del tronco encefálico, coloque las rodajas recolectadas secuencialmente de 200-300 μm en una cámara de corte disponible comercialmente para equilibrar durante 1 h a temperatura ambiente en ACSF, burbujeando continuamente con una mezcla de 95% O 2/5% CO 2 (pH 7.2-7.4, osmolaridad 300-310 mOsm / L). En estas condiciones, las rodajas permanecen viables hasta 5-6 h.
7. Electrofisiología: procedimiento de pinza de parche
- Transfiera un corte del tronco encefálico a la cámara de grabación con perfusión continua de ACSF carboxigenado ~ 1.5 ± 0.5 ml / min.
- Tire de pipetas de parche con un extractor de micropipeta de diámetro de punta 1-2 μm y resistencia en el rango de 3-6 MΩ.
- Llene las pipetas con una solución interna a base de K-gluconato (para el registro de la abrazadera de corriente).
- Para probar las propiedades neuronales en las diferentes regiones tonotópicas dentro de una rebanada, localice las neuronas en cada extremo del plano de la rebanada y acérquese con el electrodo de registro.
- Mantenga la presión de aire positiva en la punta de la pipeta mientras se acerca a una neurona.
- Muévase hacia el soma hasta que se visualice una hendidura en la neurona. Realice los siguientes dos pasos rápidamente.
- Haga un sello de gigaohmios (1 GΩ) liberando presión de aire positiva.
- Mantenga el ajuste del amplificador en modo de abrazadera de voltaje y corrija el desplazamiento de la pipeta como cero pA. Ejecute una prueba de sellado (pulso de prueba de 10 mV a 100 Hz). Aplique presión de aire negativa para romper un pequeño parche de la membrana neuronal.
- Para probar las propiedades intrínsecas activas de las neuronas auditivas, aplique inyecciones de corriente somática hiperpolarizantes y despolarizantes.
NOTA: Ejemplos de este procedimiento se pueden visualizar en el vídeo complementario S1, S2. Los detalles de este procedimiento se proporcionan en las leyendas de vídeo.
Resultados
Todos los cortes del tronco encefálico que se muestran aquí se adquirieron del tejido del tronco encefálico (~ 200-300 μm) y se obtuvieron imágenes utilizando un objetivo 5x y una óptica de contraste de interferencia diferencial (DIC). La cámara se montó en el microscopio de disección y se conectó a una computadora con software de adquisición de imágenes (consulte la Tabla de materiales). El recuadro satelital para estas figuras (paneles derechos) se visualizó utilizando un objetivo de inmersión en agua de aumento de 60x. Se tuvo cuidado de garantizar que todas las áreas del corte del tronco encefálico se magnificaran por igual mientras se obtenían imágenes digitales. Las fotografías se tomaron con un brillo y enfoque óptimos. Las imágenes digitales de los cortes del tronco encefálico se cosieron de manera plana en función del área superpuesta y se importaron a una computadora de escritorio para realizar ajustes adicionales de brillo, contraste y escala de grises. Los microcircuitos básicos del tronco encefálico auditivo del pollo fueron identificados de acuerdo con trabajos previos 1,2,5,13. Bajo el microscopio (objetivo 5x), se identificaron núcleos auditivos como el área adyacente a las fibras nerviosas fuertemente mielinizadas que corren alrededor de cada núcleo, tanto ipsilateralmente como contralateralmente a lo largo de las regiones dorsales del corte.
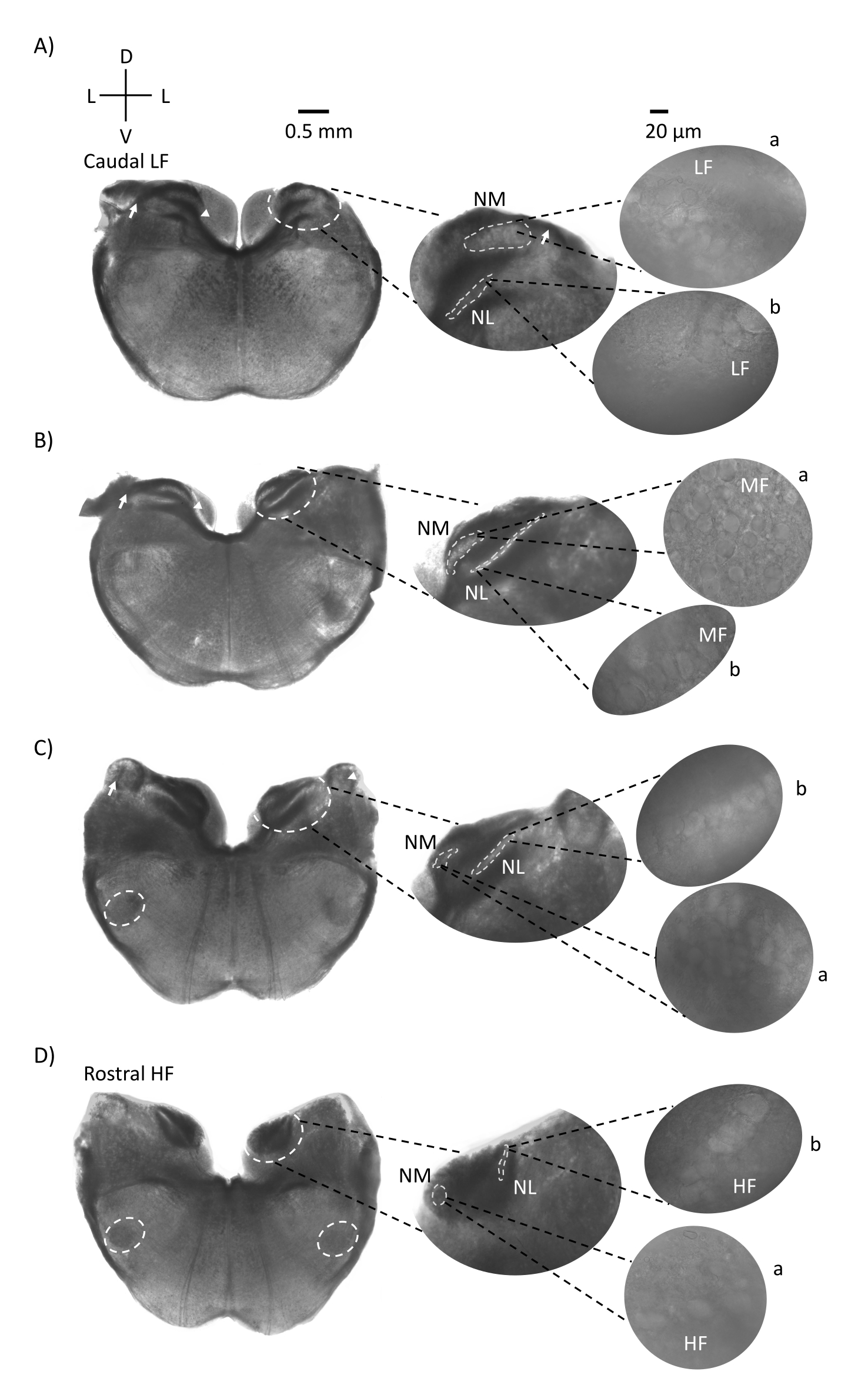
La Figura 1 muestra las secciones coronales tradicionales de tejido del tronco encefálico (200-300 μm) de un embrión de pollo E21. Los cuatro cortes coronales que se muestran aquí representan las regiones de isofrecuencia relativa de los núcleos auditivos del tronco encefálico, desde la región auditiva de CF más baja (Figura 1A, caudo-lateral) progresando hasta la región auditiva de FQ más alta (Figura 1D, rostral-medial). Para los cuatro cortes coronales de la Figura 1A-D, las regiones ampliadas de NM y NL marcadas se muestran en la columna central y se magnifican (objetivo 60x) en la vista derecha del panel de figuras (a y b, respectivamente, en recuadros de satélite). La flecha en la Figura 1A,B muestra la entrada de fibras nerviosas auditivas, y la punta de flecha muestra la bifurcación de los axones NM a la izquierda del corte. La Figura 1C muestra otra estructura del núcleo coclear aviar conocida como núcleo angular (NA, flecha a la izquierda y punta de flecha a la derecha). Los dos cortes coronales más rostrales muestran el núcleo ovárico superior (SON) ubicado a lo largo de la región ventral-lateral del corte coronal (Figura 1C, D, círculos discontinuos blancos).
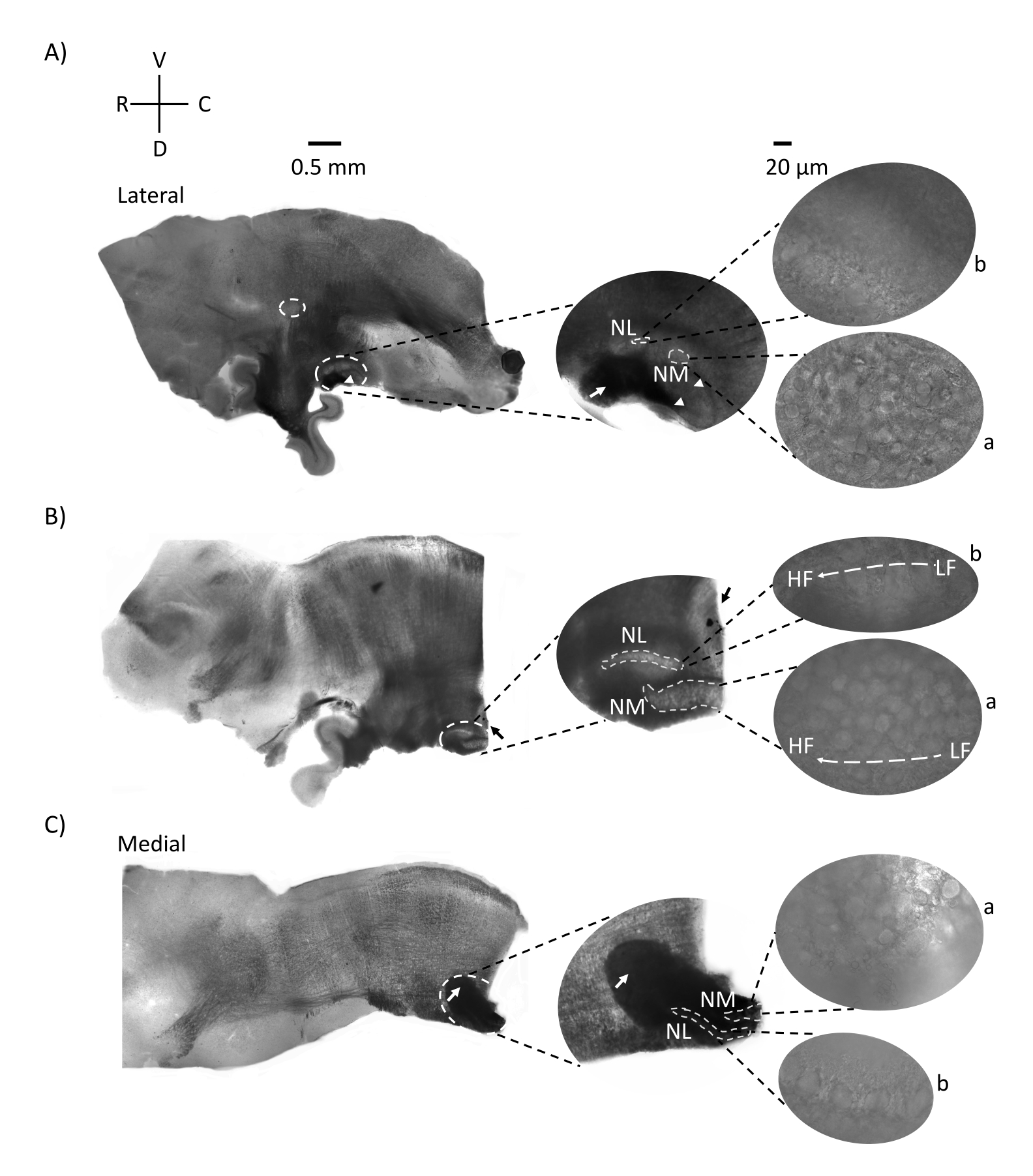
La Figura 2 muestra secciones sagitales de tejido del tronco encefálico (200-300 μm) de un embrión de pollo E21. Para los tres cortes sagitales (Figura 2A-C), las regiones ampliadas de NM y NL marcadas se muestran en la columna central y ampliadas (objetivo 60x) en la vista derecha del panel de figuras (a y b, respectivamente en imágenes de satélite). Se identificaron NM y NL donde las fibras nerviosas auditivas (Figura 2A, flecha central) entraron en el grupo de neuronas observadas con mayor aumento (Figura 2A, círculos y puntas de flecha puntas de flecha blancos, medios, pequeños) y resalta el punto de partida de la región auditiva (Figura 2A, izquierda, grande, círculo discontinuo blanco y punta de flecha). SON se identificó en la región rostro-lateral del corte más lateral (Figura 2A, círculo pequeño, blanco, discontinuo). La Figura 2B muestra regiones tonotópicas extendidas que contienen regiones auditivas relativamente bajas y altas de CF de NM y NL a lo largo del eje rostral-caudal (regiones delineadas en blanco, ver también recuadro de satélite). La Figura 2C muestra los mechones axonales ipsilaterales y contralaterales en el corte más medial y el punto final de la región auditiva (flechas izquierda y media). La orientación de las rebanadas que se muestra aquí contrasta con la orientación tradicional de las rodajas como se ve en la Figura 1 (es decir, coronal). Esto se realizó para mostrar la orientación que mejor se adapta al enfoque de una pipeta de vidrio requerida para los registros electrofisiológicos.
Para confirmar que una gran región del eje tonotópico estaba representada en la Figura 2B, se realizaron registros electrofisiológicos de pinza de corriente a partir de neuronas NM. La Figura 3 muestra similitudes funcionales y diferencias de neuronas NM maduras (E21) registradas a partir de un corte coronal (Figura 3A, B) y un corte sagital (Figura 3C, D, Video Suplementario S1, S2). Se seleccionaron dos neuronas NM de los extremos medial y lateral de un corte coronal (similar al corte que se muestra en la Figura 1B), y dos neuronas NM se seleccionaron de los extremos rostral y caudal de NM en un corte sagital (como en el corte que se muestra en la Figura 2B). La Figura 3A,B muestra propiedades de respuesta electrofisiológica similares a las inyecciones de corriente somática (−100 pA a +200 pA, incrementos de +10 pA, duración de 100 ms). El patrón de disparo de estas dos neuronas NM exhibe diferencias sutiles en este plano de corte, lo que indica una lámina de isofrecuencia relativa para las neuronas NM de frecuencia media. La Figura 3C,D muestra que los patrones de disparo tienen diferencias sustanciales a través del eje rostral-caudal, lo que indica un gradiente tonotópico relativamente más alto de una neurona NM de baja frecuencia (Figura 3C) a una neurona NM de alta frecuencia (Figura 3D). Ambas neuronas presentaron sus patrones de disparo estereotipados como se informó anteriormente14,15.
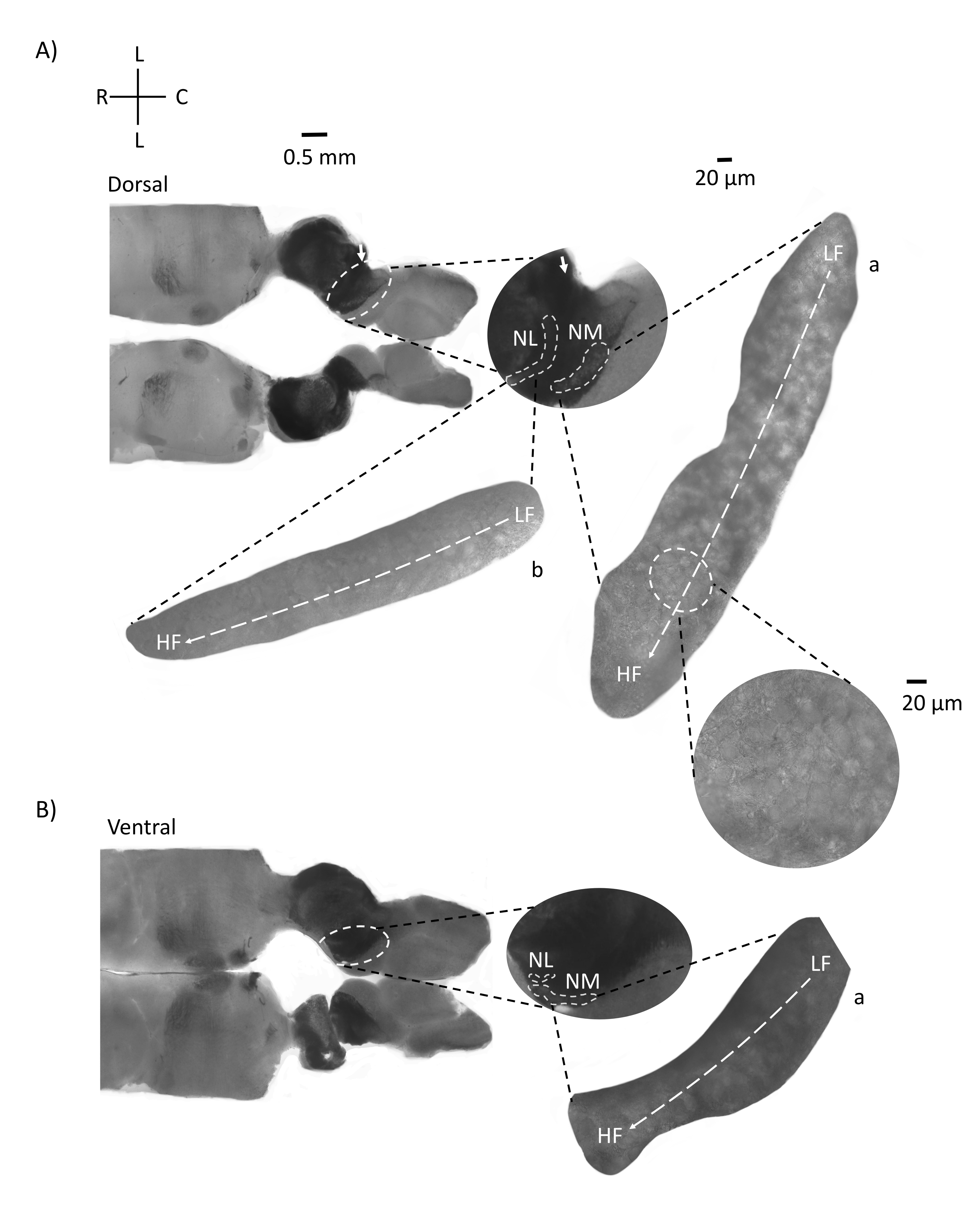
La Figura 4 muestra secciones horizontales de tejido del tronco encefálico (200-300 μm) de un embrión de pollo E21. Para ambos sectores horizontales (Figura 4A, B), las regiones ampliadas de NM y NL marcadas se muestran en la columna central y ampliadas (objetivo 60x) en la vista derecha del panel de figuras (a y b, respectivamente, en recuadros de satélite). En los cortes horizontales, se identificaron NM y NL hacia la línea media y las neuronas se extendieron a lo largo del eje lateral-medial (Figura 4A, B, regiones medias, blancas, de contorno discontinuo). Las imágenes ampliadas muestran la gran extensión del gradiente tonotópico. Las neuronas de baja frecuencia están en las regiones caudo-laterales y las neuronas de alta frecuencia están en las áreas rostral-medial (Figura 4A, B, derecha, satélites). Las fibras que atraviesan la línea media a lo largo del eje rostral-caudal muestran las conexiones contralaterales de los núcleos auditivos, pero la organización de estas fibras no está en un plano simple. Sin embargo, los cortes angulares agudos de una sección horizontal / transversal pueden seguir estas fibras axonales hacia el plano sagital. En la Figura 5 se muestran cortes de tejido del tronco encefálico de 200-300 μM de espesor en un ángulo agudo (45°) desde un plano horizontal. Los núcleos auditivos del tronco encefálico se pueden ver a través de una gran extensión diagonal que comienza desde el corte más lateral y termina en el corte más medial (Figura 5A-C, paneles centrales etiquetados, área delineada en blanco). Además, la orientación angular de las regiones NM y NL también se puede visualizar en cortes asimétricos sucesivos (Figura 5A-C, paneles centrales etiquetados, blanco, área delineada discontinua). Las imágenes ampliadas (objetivo 60x) muestran el eje tonotópico de los núcleos auditivos a medida que avanza a lo largo del eje rostral-medial a caudo-lateral (Figura 5A-C, derecha, recuadro satélite). La orientación de los sectores de la figura 5 es similar a la de la figura 2. Contrastan la presentación tradicional de imágenes, pero son más adecuados para experimentos electrofisiológicos.

Figura 1: Secciones seriadas coronales representativas del tronco encefálico. (A-D) Izquierda: cortes del eje caudal al rostral, los núcleos auditivos y fibras de conexión marcadas con un círculo discontinuo blanco. El inserto central es una vista más grande de la región auditiva, donde los núcleos se muestran dentro de los círculos discontinuos blancos a: NM y b: NL. Las flechas muestran fibras aferentes del nervio auditivo, y la punta de flecha muestra la bifurcación del axón NM en A, B. La flecha muestra NA en C. El círculo lateral blanco discontinuo muestra SON en C,D. Derecha: el inserto satelital muestra estos núcleos a 60x objetivo: a: NM y b: NL. Abreviaturas: NM = núcleo magnocellularis; NL = núcleo laminaris; NA = núcleo angular; SON = núcleo ovárico superior; LF = neuronas de frecuencia relativamente baja; MF = neuronas de frecuencia media; HF = neuronas de alta frecuencia; D = dorsal; L = lateral; V = ventral. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: Secciones seriadas sagitales representativas del tronco encefálico. (A-C) Izquierda: cortes del eje lateral al medial con los núcleos auditivos marcados en un círculo discontinuo blanco. El inserto central muestra la misma región de núcleos auditivos en una vista más grande, marcada dentro de círculos guionados blancos. (A) El círculo discontinuo blanco en el centro de la rebanada resalta el SON; flecha que muestra las fibras nerviosas auditivas y punta de flecha que muestra NA. Una mancha negra oscura en la punta derecha de la rebanada es un artefacto de imagen. Las regiones del cerebelo se pueden ver dorsal a la región auditiva en ambos cortes A y B en el panel izquierdo. (B) Un corte sagital cuya orientación se cambió al plano coronal (durante el corte). La región auditiva se identificó con tinte azul (flecha negra) y nuevamente se cortó en el plano sagital. (A-C) Inserte en el centro la región NM y NL marcada bajo líneas blancas discontinuas. Derecha: la vista satelital muestra a: NM y b: NL observados con un aumento objetivo de 60x. El gradiente tonotópico de LF y HF en los núcleos auditivos se muestra a lo largo del eje rostro-caudal. Las flechas que apuntan al área oscura en (C) muestran fibras NM fuertemente mielinizadas que atraviesan la línea media a través del eje medial. Las fibras conectan ambos lados de los núcleos auditivos. Abreviaturas: NM = núcleo magnocellularis; NL = núcleo laminaris; NA = núcleo angular; SON = núcleo ovárico superior; LF = neuronas de frecuencia relativamente baja; HF = neuronas de alta frecuencia; D = dorsal; V = ventral; R = rostral; C = caudal. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: Registros electrofisiológicos de la respuesta neuronal a inyecciones de corriente somática (−100 pA a +200 pA, incrementos de +10 pA, duración de 100 ms) en modo clamp actual. Las neuronas fueron seleccionadas para grabaciones en el mismo segmento pero en regiones opuestas extremas de NM. (A,B) Respuestas neuronales representativas en un solo corte coronal que indican propiedades relativas de isofrecuencia con diferencias sutiles. Las propiedades de respuesta representan dos neuronas MF diferentes registradas de las regiones más medial (A) y lateral (B) de NM en un corte coronal. (C,D) Grabaciones neuronales representativas de un solo corte sagital. Los registros muestran una respuesta NM (C) relativamente LF y una respuesta NM HF (D), destacando las diferencias sustantivas en el gradiente tonotópico dentro de una sola sección sagital. Abreviaturas: NM = núcleo magnocellularis; LF = neuronas de frecuencia relativamente baja; MF = neuronas de frecuencia media; HF = neuronas de alta frecuencia. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: Secciones seriadas horizontales representativas del tronco encefálico. (A,B) Izquierda: cortes a lo largo del eje dorsal a ventral, los núcleos auditivos están marcados con círculos discontinuos blancos. Las fibras aferentes del8º nervio craneal conectan núcleos auditivos marcados con flecha. El inserto central es una vista más grande de la región de los núcleos auditivos con núcleos auditivos marcados bajo líneas discontinuas blancas Se muestran las regiones NM y NL. Un claro movimiento topológico de los núcleos auditivos se puede ver en A,B. (A,B) Derecha: gran vista de satélite que muestra a: NM y b: NL. El inserto derecho muestra los núcleos auditivos observados en un aumento objetivo de 60x y el eje topológico curvo de LF a HF a lo largo de un eje caudo-lateral a rostral-medial. Abreviaturas: NM = núcleo magnocellularis; NL = núcleo laminaris; LF = neuronas de frecuencia relativamente baja; HF = neuronas de alta frecuencia; L = lateral; R = rostral; C = caudal. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 5: Secciones seriadas horizontales representativas/transversales agudas (45°). (A-C) Izquierda: secciones seriadas del tronco encefálico, núcleos auditivos marcados en un círculo discontinuo blanco. El inserto central es una vista más grande de la región auditiva. (A) El inserto central muestra la mayor propagación de neuronas NM y NL en estos cortes. (B,C) Inserción media: los núcleos auditivos marcados con líneas discontinuas blancas muestran un cambio topológico gradual en comparación con (A-C). Derecha: inserto satelital que muestra los núcleos auditivos a: NM y b: NL en un aumento objetivo de 60x. El eje tonotópico de las regiones LF a HF en NM y NL gira angularmente de cortes laterales a mediales. Abreviaturas: NM = núcleo magnocellularis; NL = núcleo laminaris; LF = neuronas de frecuencia relativamente baja; HF = neuronas de alta frecuencia; V = ventral; R = rostral; D = dorsal; C = caudal. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Video complementario S1: Inyecciones de corriente somática hiperpolarizantes y despolarizantes. Propiedades de respuesta de una neurona de baja y alta frecuencia a inyecciones de corriente somática de 100 ms en modo de pinza actual. Las neuronas fueron seleccionadas del mismo corte del tronco encefálico sagital. Las inyecciones varían de -100 a +200 pA en pasos de incrementos de +10 pA, duración de tiempo de 100 ms. Los potenciales de acción se observan en respuesta a suficientes pasos actuales de despolarización. El video corresponde a las trazas finales que se muestran en la Figura 3C. Haga clic aquí para descargar este archivo.
Video complementario S2: Inyecciones de corriente somática hiperpolarizantes y despolarizantes. Similar al video suplementario S1, este video muestra las propiedades de respuesta de una neurona de baja y alta frecuencia a inyecciones de corriente somática de 100 ms en modo de pinza actual. Las neuronas fueron seleccionadas del mismo corte del tronco encefálico sagital. Las inyecciones varían de -100 a +200 pA en pasos de incrementos de +10 pA, duración de tiempo de 100 ms. Los potenciales de acción se observan en respuesta a suficientes pasos actuales de despolarización. El vídeo corresponde a las trazas finales que se muestran en la Figura 3D. Haga clic aquí para descargar este archivo.
Discusión
Las secciones coronales del tejido embrionario del tronco encefálico del pollo han permitido el estudio de la lámina de isofrecuencia individual relativa durante décadas 1,2,5. Sin embargo, la organización tonotópica (es decir, la frecuencia) del tronco encefálico auditivo del pollo es topológicamente complicada y puede ser más accesible en otros ejes anatómicos dependiendo de la pregunta de investigación específica. Aunque es suficiente para investigar cuestiones anatómicas y fisiológicas relacionadas con regiones individuales de isofrecuencia, el estudio de las variaciones tonotópicas y su desarrollo en áreas auditivas más grandes del tronco encefálico están algo limitados por las secciones coronales. Para superar esta limitación, este protocolo describe enfoques en los planos sagital, horizontal y horizontal/transversal para proporcionar ejemplos adicionales de tejido auditivo del tronco encefálico que exhiben propiedades tonotópicas máximas y gradientes en una sección individual del tronco encefálico.
Las secciones sagitales de las regiones auditivas del tronco encefálico muestran que diferentes áreas tonotópicas se distribuyen en una región más grande dentro del corte en comparación con las secciones coronales (área auditiva sagital = ~ 300-600 μm, área auditiva coronal = ~ 200-350 μm). Por ejemplo, las regiones NM y NL se visualizaron en un área más grande a lo largo del eje rostro-caudal en secciones sagitales (por ejemplo, Figura 2B), y el gradiente tonotópico funcional que corre a lo largo de este eje anatómico estaba contenido en gran medida dentro de un solo corte sagital. Esto se confirmó aún más con registros de pinza de corriente de diferencias neuronales intrínsecas que varían a lo largo del gradiente rostral-caudal como se informó anteriormente14,15 (por ejemplo, Figura 3C, D). Los experimentos futuros que resalten las propiedades anatómicas e inmunohistoquímicas a lo largo del eje tonotópico podrían investigar más a fondo los gradientes conocidos de las propiedades auditivas dentro de un solo plano de corte sagital. Estos incluyen, pero no se limitan a, tinción MAP2 y patrones de expresión de canales de potasio, que son gradientes conocidos de arquitectura dendrítica y propiedades intrínsecas de NM y NL que se han demostrado previamente en secciones coronales sucesivas16.
Las secciones horizontales de las regiones auditivas del tronco encefálico muestran que el NM y el NL están ubicados hacia la línea media. Una porción de fibras axonales auditivas corre diagonal o perpendicular al plano horizontal (Figura 4). Estas fibras pueden ser seguidas haciendo un corte angular agudo de 45° al plano sagital. Los cortes horizontales/transversales resultantes eran más grandes que los cortes sagitales u horizontales, y las fibras axonales largas corrían a través del eje rostro-caudal tanto para el lado ipsilateral como para el contralateral. Tanto NM como NL se pueden visualizar en una región diagonal más grande (~ 400-700 μm) de modo que las conexiones contralaterales se pueden visualizar a lo largo de un eje lateral-medial. Además, el plano de corte horizontal/transversal también muestra cómo las regiones auditivas y el gradiente tonotópico resultante hacen un giro angular (Figura 5). La exposición angular de las conexiones contralaterales en un área más grande hace que estos cortes sean más adecuados para la estimulación electrofisiológica y los estudios de microcircuitos que los cortes coronales tradicionales.
Ventajas adicionales
La formación de microcircuitos auditivos requiere la coordinación espaciotemporal de señales que promueven la supervivencia neuronal, la sinaptogénesis, la diferenciación axonal, la arquitectura dendrítica y la maduración. Por lo tanto, se pueden utilizar secciones alternativas del microcircuito auditivo del tronco encefálico del embrión de pollo para los siguientes temas de investigación: organización morfológica de neuronas en dimensiones topográficamente diferentes; organizar y mapear los conectomas de todos los núcleos auditivos y vestibulares; identificación y caracterización de los patrones de actividad de los constituyentes de circuitos en planos de isofrecuencia y tonotópicos; la organización topográfica de microcircuitos excitatorios versus inhibitorios y relaciones con poblaciones de neuronas especializadas (núcleos); localización espacial de las neuronas de los núcleos auditivos y su CFpredictiva 17; focalización sistemática de tipos neuronales tonotópicos específicos; seguimiento de las células progenitoras y su desarrollo en núcleos conservados; linaje genético de las células a la evolución de los circuitos neuronales18; anatomía comparativa del tronco encefálico entre especies; investigación de circuitos vestibulares como el complejo vestibular de Deiter (DC)19; y sincronía y diafonía entre núcleos vestibulares.
Un enfoque multifacético que utiliza diferentes planos de corte puede ayudar a responder preguntas fundamentales sobre propiedades anatómicas y biofísicas desconocidas de los microcircuitos del tronco encefálico. Un buen ejemplo es la relación entre los principales núcleos auditivos (NM, NA, NL y SON) y los núcleos vestibulares, incluyendo el núcleo dorsal del lemniscus lateral (LLDp), el núcleo semilunar (SLu)20 y el núcleo tangencial (TN)3. Sin embargo, este protocolo y estos estudios basados en cortes tienen algunas limitaciones.
Precauciones y limitaciones
Dependiendo de la institución que realiza los experimentos, las pautas éticas y el manejo de embriones de pollo pueden diferir. Mientras que las Directrices de los Institutos Nacionales de Salud para el Cuidado y Uso de Animales de Laboratorio permiten la decapitación rápida, existen métodos alternativos para la eutanasia de embriones de pollo21. El tejido del tronco encefálico del embrión de pollo en desarrollo temprano es suave y delicado en comparación con los embriones más viejos. Tiene varias conexiones y vasos sanguíneos en la superficie que necesitan precaución adicional al eliminarlos. El tejido debe mantenerse en dACSF helado y perfundirse con 95% de O2/5% deCO2 para aumentar la viabilidad.
El método de corte sagital solo es útil para la tonotopía ipsilateral. Este método de corte proporciona cortes más grandes que los cortes coronales, cuyo manejo podría ser precario. Sin embargo, se pueden recortar las rodajas utilizando métodos de agujas cruzadas descritos en detalle en otra parte22. El uso de un 4% de tronco encefálico incrustado en el bloque de agarosa LMP puede salvar estructuras delicadas en rodajas, pero se debe tener cuidado de no verter agarosa excesivamente caliente. Configurarlo rápidamente colocando el tronco encefálico bloqueado con agarosa en un ambiente frío durante ~ 1 minuto hace que las rebanadas sean más viables para las grabaciones electrofisiológicas.
La aplicación de superglue en cantidades excesivas puede ser tóxica. Debe aplicarse mínimamente, y las cantidades excedentes deben lavarse inmediatamente cambiando el dACSF. Para cortes angulares agudos (45°), cortar el ángulo del bloque de agarosa es crítico; Uno puede usar un espejo para ver el ángulo frontal mientras corta el bloque de agarosa con una cuchilla afilada. Las cuchillas disponibles comercialmente pueden tener un recubrimiento de cera que debe limpiarse con alcohol y secarse antes de su uso. Se requiere optimización para la velocidad y frecuencia de corte del vibratomo, ya que los mechones de fibra axonal son más duros que el tejido cortical o matriz. Mantener una amplitud alta y usar una solución de disección refrigerada puede prevenir el daño tisular.
Todas las soluciones deben prepararse frescas, y Ca 2+ y Mg2+ deben añadirse al ACSF después de burbujear 95% O 2/5% CO2 . De lo contrario, puede haber precipitación de Ca2+. Se debe usar un pincel para manejar las rodajas suavemente dentro del vibratomo. Mantenga el tiempo total de corte por debajo de 15 minutos si es posible. Se puede utilizar una pipeta de vidrio Pasteur para maniobrar cortes del tronco encefálico.
No utilice detergente ni agentes de lavado corrosivos para cristalería y equipos que entren en contacto con las rodajas utilizadas en electrofisiología. Las imágenes tomadas representan la apariencia de tejido de 200-300 μM de espesor bajo óptica de contraste de interferencia diferencial (DIC). La calidad visual será más pobre que la inmunohistoquímica o la microscopía electrónica, pero refleja con precisión lo que un experimentador verá al realizar registros electrofisiológicos.
Los estudios relacionados con el desarrollo temprano de microcircuitos a lo largo de un eje anatómico alternativo, ya sean dorsal-ventral, rostral-caudal o ipsilateral-contralateral, están limitados en el tronco cerebral auditivo del pollo. Una razón para esto es porque el papel de los códigos transcripcionales y la regulación del desarrollo tonotópico en el tronco encefálico aún no se comprende completamente. Los fenómenos funcionales como la modulación de arriba hacia abajo y la actividad espontánea a menudo se pierden cuando se observa la actividad in vitro. Sin embargo, la investigación in vivo se complementa con registros específicos y directos de neuronas individuales que solo son posibles en estas condiciones de corte. El refinamiento de la obtención de tejido del tronco encefálico a lo largo de diferentes orientaciones podría proporcionar información perspicaz sobre el desarrollo y la complejidad de los gradientes tonotópicos en los microcircuitos auditivos del tronco encefálico del pollo.
Divulgaciones
Todos los autores declaran que la investigación se llevó a cabo sin ningún interés comercial o financiero y que no tienen ningún conflicto de intereses.
Agradecimientos
Este trabajo es apoyado por la subvención NIH/NIDCD R01 DC017167. Agradecemos a Kristine McLellan por proporcionar comentarios editoriales sobre una versión anterior del manuscrito.
Materiales
| Name | Company | Catalog Number | Comments |
| Adobe photoshop 2021 | Adobe | ||
| Anti-vibration table 30"x 36" - OTMC - 63533 | TMC | ||
| Cell sens standard software | OLYMPUS | ||
| Digidata 1440A | MOLECULAR DEVICES | ||
| Digital amplifier multiclamp 700B | MOLECULAR DEVICES | ||
| DSK line-up linearslicer pro7 | TED PELLA, INC | ||
| Micromanipulator MPC-385 / OSI-MPC-385-2 | OLYMPUS AMERICA INC | ||
| Micropipette puller P-97 | SUTTER INSTRUMENTS | ||
| Microscope BX51W1 | OLYMPUS AMERICA INC | ||
| MS ICE software | Microsoft Corporation | ||
| Ohaus balance model AV212 | Ohaus Adventurer | ||
| Olympus DPSI0 /DPS80 camera | OLYMPUS | ||
| pClamp and Axoclamp data Acquisition Softwares | MOLECULAR DEVICES | ||
| pH meter lab 850 benchtop | SCHOTT INSTRUMENTS | ||
| Sharp stainless blade | Dorco/Personna | ||
| Vapor pressure osmometer model 5600 | WESCOR INC | ||
| Water purification systems Smart2pure 6UV/UF | Thermo Scientific | ||
| Chemicals- list | |||
| Agrose Low melt IB70051 | IBI SCIENTIFIC | ||
| CaCl2 (Calcium Chloride) | ACROS organics | ||
| Cynergy instant adhesive CA6001 | Resinlab | ||
| Dextrose (D-(+)-glucose) | VWR Life Science | ||
| Ethyl alcohol | IBI SCIENTIFIC | ||
| KCl (Potassium Chloride) | Amresco.Inc | ||
| MgCl2 (Magnesium Chloride) | Sigma-Aldrich | ||
| NaCl (Sodium Chloride) | Amresco.Inc | ||
| NaH2PO4 (Sodium Dihydrogen Phosphate) | Amresco.Inc | ||
| NaHCO3 (Sodium Bicarbonate) | Amresco.Inc |
Referencias
- Rubel, E. W., Parks, T. N. Organization and development of brain stem auditory nuclei of the chicken: tonotopic organization of n. magnocellularis and n. laminaris. Journal of Comparative Neurology. 164 (4), 411-433 (1975).
- Rubel, E. W., et al. Organization and development of brain stem auditory nuclei of the chicken: ontogeny of n. magnocellularis and n. laminaris. Journal of Comparative Neurology. 166 (4), 469-489 (1976).
- Shao, M., et al. Spontaneous synaptic activity in chick vestibular nucleus neurons during the perinatal period. Neuroscience. 127 (1), 81-90 (2004).
- Fukui, I., Ohmori, H. Tonotopic gradients of membrane and synaptic properties for neurons of the chicken nucleus magnocellularis. Journal of Neuroscience. 24 (34), 7514-7523 (2004).
- Sanchez, J. T., Seidl, A. H., Rubel, E. W., Barria, A. Preparation and culture of chicken auditory brainstem slices. Journal of Visualized Experiments. (49), e2527 (2011).
- Sanchez, J. T., Lu, Y., Fay, R. R., Popper, A. N., Cramer, K., Coffin, A. Glutamate signaling in the auditory brainstem. Auditory Development and Plasticity: Springer Handbook of Auditory Research. 64 (4), 75-108 (2017).
- Parks, T. N. Morphology of axosomatic endings in an avian cochlear nucleus: nucleus magnocellularis of the chicken. Journal of Comparative Neurology. 203 (3), 425-440 (1981).
- Jhaveri, S., Morest, D. K. Sequential alterations of neuronal architecture in nucleus magnocellularis of the developing chicken: a Golgi study. Neuroscience. 7 (4), 837-853 (1982).
- Carr, C. E., Boudreau, R. E. Central projections of auditory nerve fibers in the barn owl. Journal of Comparative Neurology. 314 (2), 306-318 (1991).
- Köppl, C. Auditory nerve terminals in the cochlear nucleus magnocellularis: differences between low and high frequencies. Journal of Comparative Neurology. 339 (3), 438-446 (1994).
- Fukui, I., et al. Improvement of phase information at low sound frequency in nucleus magnocellularis of the chicken. Journal of Neurophysiology. 96 (2), 633-641 (2006).
- Wang, X., et al. Postsynaptic FMRP regulates synaptogenesis in vivo in the developing cochlear nucleus. Journal of Neuroscience. 38 (29), 6445-6460 (2018).
- Lu, T., Cohen, A. L., Sanchez, J. T. In ovo electroporation in the chicken auditory brainstem. Journal of Visualized Experiments. (124), e55628 (2017).
- Hong, H., Sanchez, J. T. Need for speed and precision: structural and functional specialization in the cochlear nucleus of the avian auditory system. Journal of Experimental Neuroscience. (12), 1-16 (2018).
- Hong, H., et al. Diverse intrinsic properties shape functional phenotype of low-frequency neurons in the auditory brainstem. Frontiers in Cellular Neuroscience. 12, 1-24 (2018).
- Wang, X., Hong, H., Brown, D. H., Sanchez, J. T., Wang, Y. Distinct neural properties in the low-frequency region of the chicken cochlear nucleus magnocellularis. eNeuro. 4 (2), 1-26 (2017).
- Tabor, K. M., et al. Tonotopic organization of the superior olivary nucleus in the chicken auditory brainstem. Journal of Comparative Neurology. 520 (7), 1493-1508 (2012).
- Lipovsek, M., Wingate, R. J. Conserved and divergent development of brainstem vestibular and auditory nuclei. Elife. 7, 40232 (2018).
- Passetto, M. F., et al. Morphometric analysis of the AMPA-type neurons in the Deiter's vestibular complex of the chick brain. Journal of Chemical Neuroanatomy. 35 (4), 334-345 (2008).
- Curry, R. J., Lu, Y. Intrinsic properties of avian interaural level difference sound localizing neurons. Brain Research. 1752, 147258 (2021).
- Aleksandrowicz, E., Herr, I. Ethical euthanasia and short-term anesthesia of the chick embryo. ALTEX - Alternatives to Animal Experimentation. 32 (2), 143-147 (2015).
- Palkovits, M. Isolated removal of hypothalamic or other brain nuclei of the rat. Brain Research. 14 (59), 449-450 (1973).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados