
Method Article
Tonotopik gradyanları ve mikrodevreleri değerlendirmek için embriyonik tavuk işitsel beyin sapının dilimlenmesi
Bu Makalede
Özet
Burada, bir beyin sapı dilimi içinde tonotopik özelliklerin ve gelişimsel yörüngelerin araştırılması için tavuk embriyosunun koronal olmayan işitsel beyin sapı dilimlerinin elde edilmesine yönelik bir protokol sunuyoruz. Bu dilimler, geleneksel koronal kesitlerin tek bir dilim düzlemi içindeki daha büyük tonotopik bölgeleri kapsayan sagital, yatay ve yatay / enine bölümleri içerir.
Özet
Tavuk embriyosu, tonotopik (yani frekans) eksen boyunca farklı şekilde yönlendirilmiş son derece uzmanlaşmış mikrodevre ve nöronal topolojiden oluşan işitsel beyin sapını incelemek için yaygın olarak kabul edilen bir hayvan modelidir. Tonotopik eksen, rostral-medial düzlemde yüksek frekanslı seslerin ayrılmış kodlamasına ve kaudo-lateral bölgelerde düşük frekanslı kodlamaya izin verir. Geleneksel olarak, embriyonik dokunun koronal beyin sapı dilimleri, göreceli bireysel izo-frekanslı laminanın çalışmasına izin verir. Bireysel izo-frekans bölgelerine ait anatomik ve fizyolojik soruları araştırmak için yeterli olsa da, tonotopik varyasyonun incelenmesi ve daha büyük işitsel beyin sapı alanlarında gelişimi biraz sınırlıdır. Bu protokol, alt işitsel beyin sapındaki frekans bölgelerinin daha büyük gradyanlarını kapsayan tavuk embriyolarından beyin sapı dilimleme tekniklerini bildirmektedir. Tavuk işitsel beyin sapı dokusu için farklı dilimleme yöntemlerinin kullanılması, tonotopik özelliklerin ve gelişimsel yörüngelerin daha büyük gradyanlarının koronal kesitlerden daha iyi korunduğu bir beyin sapı dilimi içinde elektrofizyolojik ve anatomik deneylere izin verir. Çoklu dilimleme teknikleri, işitsel beyin sapı mikrodevrelerinin çeşitli anatomik, biyofiziksel ve tonotik özelliklerinin daha iyi araştırılmasına izin verir.
Giriş
Tavuk embriyosu, hücre biyolojisi, immünoloji, patoloji ve gelişimsel nörobiyoloji dahil olmak üzere çok sayıda ve çeşitli bilimsel alanlarda temel biyolojik soruları incelemek için değerli bir araştırma modelidir. Tavuk işitsel beyin sapının mikro devresi, işitsel morfoloji ve fizyoloji açısından anlaşılabilecek son derece uzmanlaşmış bir devrenin mükemmel bir örneğidir. Örneğin, Rubel and Parks (1975) ilk olarak tavuk çekirdeği magnocellularis (NM) ve nucleus laminaris'in (NL) tonotopik yönelimini (yani frekans gradyanı), sagital düzleme göre ~ 30 ° yönlendirilmiş çekirdeklerin ekseni boyunca doğrusal bir fonksiyon olarak tanımlamıştır. NM ve NL'deki bireysel nöronlar, karakteristik frekansları (CF) olarak bilinen en iyi ses frekanslarını, rostral-medial düzlem boyunca kaudo-lateral bölgeye kodlarlar. Yüksek frekansa duyarlı nöronlar rostral-medial bölgededir ve düşük frekansa duyarlı nöronlar kaudo-lateral olarak bulunur. Bu nedenle, tonotopik özellikleri incelemek için işitsel beyin sapı dokusunun geleneksel diseksiyon yöntemleri, ardışık koronal dilimleri kullanmıştır. Gerçekten de, gelişmekte olan tavuk embriyolarının işitsel mikro devreleri, on yıllardır 1,2,3,4,5,6 için ardışık kaudal-rostral koronal düzlem beyin sapı dilimleri aracılığıyla tonotopik işitsel fonksiyonların sinyal işlemesini incelemek için bir model sistem olarak kurulmuştur.
Bununla birlikte, NM ve NL'nin tonotopik organizasyonu topolojik ve morfolojik olarak kıvrımlıdır. İşitme siniri girdileri, yüksek CF girişlerinin, bir adendritik NM hücresinin somatik çevresinin en az dörtte birini kaplayan ampul benzeri yapılarda sona ereceği şekilde dağıtılır. Tersine, düşük CF girişleri uç ampul benzeri terminallerle değil, NM nöronlarının dendritleri üzerinde çoklu bukon sinapslarıyla düzenlenir. Orta CF girişleri hem uç ampul hem de buton benzeri sinapslar 4,7,8,9,10,11,12 olarak sona erer. NL'de, oldukça klişeleşmiş dendritik gradyan sadece dendritik uzunlukta değil, aynı zamanda dendritik genişlikte de belirgindir. Bu eşsiz dendritik gradyan, tonotopik eksene yakından uyar. Dendritler, sırasıyla yüksek ila düşük CF nöronlarından uzunlukta 11 kat ve genişlikte beş kat artışa uğrar,6. Bu çekirdeklerin koronal dilimlerdeki bu tür kıvrımlı dağılımlarının üstesinden gelmek için, bu protokol sagital, yatay ve yatay / enine düzlemlerde diseksiyon yaklaşımlarını tanımlar. Bu dilimleme teknikleri, bireysel bir dilim düzleminde maksimum tonotopik özellikler sergileyen işitsel beyin sapı dokusu örnekleri sağlar.
Protokol
Tüm prosedürler Northwestern Üniversitesi Kurumsal Hayvan Bakım ve Kullanım Komiteleri (IACUC) tarafından onaylandı ve Laboratuvar Hayvanlarının Bakımı ve Kullanımı için Ulusal Sağlık Enstitüleri Kılavuzlarına uygun olarak gerçekleştirildi. Beyin sapı dokusunun diseksiyonu ve hazırlanmasına yönelik protokoller önceki protokoller 5,13 ile uyumludur.
1. Yumurta işleme
- IACUC onaylı yerel bir hayvan tedarikçisinden döllenmiş yumurta (Gallus gallus domesticus) satın alın.
- Yumurtaları varışta hemen 14 ° C'de bir buzdolabında saklayın ve 5 gün içinde kuluçkaya yatırın.
NOT: Embriyo canlılığı 1 hafta sonra önemli ölçüde azalır. - Kuluçkadan önce yumurtaları% 70 etanol ile 38 ± 1 ° C ve ~% 50 nemde sterilize edin.
2. Yapay beyin-omurilik sıvısı (ACSF) bileşimi ve hazırlanması
- 10x ACSF stok çözeltisi oluşturmak için aşağıdaki kimyasalları 1 L'lik 18,2 MΩcm dH 2 O'da karıştırın: NaCl (sodyum klorür) 130 mM, NaHCO3 (sodyum bikarbonat) 26 mM, KCl (potasyum klorür) 2,5 mM, NaH2PO4 (sodyum dihidrojen fosfat) 1,25 mM, dekstroz (D-(+)-glikoz) 10 mM. Stok çözeltisini buzdolabında saklayın.
- MgCl 2 (magnezyum klorür) 1 M ve CaCl 2 (kalsiyum klorür) 1 M çözeltilerini ayrı ayrı 18,2 MΩcm dH2 O'da hazırlayın ve buzdolabında saklayın.
- Kullanmadan hemen önce, 10x ACSF'yi 1x'e seyreltin ve 15-20 dakika boyunca% 95 O 2/5% CO 2 ile sürekli kabarcık yapın ve MgCl2 ve CaCl2 ekleyin. ACSF ve dACSF'yi (ACSF'yi diseke ederek) hazırlamak için, sırasıyla Mg 2 + 1 mM, Ca 2 + 3 mM ve Mg 2 + 3 mM, Ca 2 + 1 mM'lik bir son konsantrasyona ayarlayın.
- ACSF için köpürme hızını, pH 300 ila 310 mOsm / L arasında bir ozmolalite ile 7.2-7.4 olacak şekilde ayarlayın.
NOT: ACSF'nin köpürürken bir buz banyosuna yerleştirilmesi, diseksiyon sırasında doku yapısal bütünlüğünü destekleyecek düşük bir çözelti sıcaklığını korumak için faydalıdır.
3. Agarose (% 5) blok hazırlığı
- 100 mL dACSF içinde 5 g agarozu karıştırın. 2-3 dakika boyunca 100 ° C'lik bir su banyosu veya mikrodalga kullanın, agaroz tamamen çözünene ve köpürmeye başlayana kadar topakları önlemek için her 30 saniyede bir karıştırın.
- Eritilmiş agarozu 5 mm kalınlığa kadar boş bir Petri kabına dökün ve ayarlamak için oda sıcaklığında tutun. Ayarladıktan sonra, Petri kabını parafilm kullanarak kapatın ve 4 ° C'de saklayın.
- Agarozu keskin bir bıçakla kübik bloklar halinde kesin ve diseksiyon sırasında kullanın.
4. Diseksiyon protokolü ve işitsel beyin sapının izole edilmesi
- Diseksiyon alanını% 70 etil alkol çözeltisi spreyi kullanarak temizleyin.
- Destek veya açılı agaroz bloğunu vibratom tepsisine yapıştırın.
- İstediğiniz yaştaki yumurtaları seçin (mevcut protokolde E20 ve E21). Yumurtaları yukarıda listelenen protokolleri izleyerek adım 1'de olduğu gibi kullanın ve kuluçkaya yatırın.
- Yumurtayı parlak bir ışığın altına yerleştirerek (konserve) hava dolu boşluğu bulun ve bu alanı yumurtanın daha büyük veya daha yuvarlak tarafında arayın.
- Yumurtaları oda sıcaklığına alıştırın, kabuğu hava dolu alanın üzerinde çatlatın ve membran kesesini açığa çıkarın.
- Gagayı açığa çıkarmak için kese içinde nazik bir kesi yapın.
- Bir neşter ile, boynu ve başını yavaşça yumurtadan çekin.
- Hızlı bir şekilde keskin makas kullanarak başın kafasını kesin.
- Kafa kesmeden sonra, diseksiyon pedinden fazla kanı çıkarmak için kafayı buz soğutmalı dACSF ile temizleyin.
- Kafayı buzla soğutulmuş dACSF'de sabit tutun ve rostro-kaudal bir kesi yapın. Gözlerin arkasında ve arasında kesiyi başlatın ve hasat edilen boynun uzunluğunu takip edin.
NOT: Genç embriyolar insizyon yaparken daha az basınç gerektirebilir. - Kafatasını ortaya çıkarmak için cildi ayırın.
- Kafatasını gözün arkasında orta çizgiden yanal yöne doğru kesin. Bunu her iki yarım küre için de yapın.
NOT: Bu adım, beyin dokusunu sağlam tutarken kafatasının rostral kısmını bağlı beyinden ayırmaya yardımcı olur5. - Kafatasının rostral kısmını dilimleyin. Bıçağı gözlerin arkasına yerleştirin ve hızlı bir kesim yapın.
NOT: Takılı kafatasını temiz bir şekilde kesmek için çaba sarf edilmesi gerekebilir. - Kafayı soğuk bir dACSF kabına batırın.
- Küçük bir makas çifti kullanarak, doku hasarına neden olmadan beyni kafatasından ayırmaya çalışmak için kafatasının kaudal bölgesinde orta çizgiden yanal kesiklere kadar yapın.
- Beyin sapını ve beyinciği nazikçe ortaya çıkarın. Tüm kafatasının sırt bölgesini geri çekin, beyin sapını dikkatlice çıkarın ve hafif kızakla ince bir boya fırçası yardımıyla ortaya çıkarın. Beyin sapını doku ve kan damarlarını birbirine bağlayan dokudan temizlemek için kavisli forseps kullanın. 8. kranial sinir bölgesine daha fazla dikkat edin ve her iki tarafta kısa bir süre sağlam sinir lifi bıraktığınızdan emin olun.
- Pedinkülleri keserek ve kan damarlarını dikkatlice çıkararak beyin sapını beyincikten ayırın. Ek kan damarlarının beyin sapını kesin.
NOT: Tüm prosedürün sürekli olarak karbonhidrat ile kabarcıklanmış buz soğutmalı dACSF'de yapıldığından emin olun (%95 O 2/%5 CO2).
5. Vibratom dilimleme
NOT: Aşağıdaki adımlarda, dokunun arkası kübik bir agaroz parçası ile desteklenmelidir.
- Vibratom bıçağını yatay eksen boyunca yerleştirin ve beyin sapını bir dilimleme tepsisine yapıştırın. Rostral tarafı yapıştırın, rostral-kaudal ekseni koronal dilimler için dikey tutun.
- Sagital dilimler için lateral-medial ekseni dikey tutun.
- Yatay dilimler için dorsal-ventral ekseni dikey tutarak ventral tarafı yapıştırın.
- Akut açısal sagital-yatay düzlemi elde etmek için, beyin sapının ventral tarafını yapıştırın ve ventral-dorsal ekseni agaroz bloğunun hipotenüs yüzeyinde dikey tutarak 45 ° 'lik bir açıyla kesin. Dilimleme tepsisine bakan agaroz bloğunun karşı yüzeyini yapıştırın ve rostral-kaudal ekseni bıçak kenarına paralel tutun.
6. Kırılgan veya büyük beyin sapı dokusu parçalarının taşınması
- Adım 5'e alternatif bir yaklaşımda, izole edilmiş beyin sapını 35 mm x 10 mm'lik bir Petri kabında ~ 40 ° C'de% 4 düşük erime noktası (LMP) agarozuna batırın.
- Daldırılmış beyin sapına agaroz döktükten sonra, katılaşması için Petri kabını buzun üzerine yerleştirin. Kübik agaroz bloğunu gömülü beyin sapı ile keskin bir tıraş bıçağı kullanarak kesin.
- LMP agaroz bloğunu rostral tarafına yapıştırın ve beyin sapının rostral-kaudal eksenini dikey tutun.
- NM bölgesi görselleştirilene kadar koronal dilimler alın.
- Agaroz bloğunu tutkaldan keskin bir bıçakla çıkarın. Çekirdekleri tespit etmek için, ince bir iğne ile NM'ye yavaşça 0,5 μL boya (toluidin mavisi veya turuncu G) yerleştirin.
- Bu bloğu sagital veya yatay dilimler için dilimleme tepsisine tekrar monte edin ve çekirdekleri lekeli bölgeye göre tanımlayın.
- En iyi performans için, titreşimatom dilimleme hızını 4 - 5 (~ 30 ± 4 mm / dak), titreşim frekansını 85-87 Hz'de ve dilimleme genliğini 4-6'da (~1 ± 0,2 mm) ayarlayın.
- Beyin sapı kesitinden sonra, 200-300 μm sıralı olarak toplanan dilimleri, ACSF'deki oda sıcaklığında 1 saat boyunca dengelemek için, sürekli olarak% 95 O 2/5% CO2 (pH 7.2-7.4, ozmolarite 300-310 mOsm / L) karışımı ile kabarcıklandırılmış, ticari olarak temin edilebilen bir dilim odasına yerleştirin. Bu koşullarda, dilimler 5-6 saate kadar canlı kalır.
7. Elektrofizyoloji: yama kelepçesi prosedürü
- Karboksijenli ACSF ~ 1.5 ± 0.5 mL / dak'nın sürekli perfüzyonu ile bir beyin sapı dilimini kayıt odasına aktarın.
- Yamalı pipetleri, uç çapı 1-2 μm olan ve direnci 3-6 MΩ aralığında olan bir mikropipet çektirme makinesi ile çekin.
- Pipetleri K-Glukonat bazlı dahili çözelti ile doldurun (akım kelepçesi kaydı için).
- Bir dilim içindeki farklı tonotopik bölgeler boyunca nöronal özellikleri test etmek için, dilim düzleminin her iki ucundaki nöronları bulun ve kayıt elektroduna yaklaşın.
- Bir nörona yaklaşırken pipet ucunda pozitif hava basıncını koruyun.
- Nöron üzerinde bir girinti görselleşene kadar soma'ya doğru ilerleyin. Sonraki iki adımı hızlı bir şekilde gerçekleştirin.
- Pozitif hava basıncını serbest bırakarak gigaohm (1 GΩ) sızdırmazlık yapın.
- Amplifikatör ayarını voltaj kelepçesi modunda tutun ve pipet ofsetini sıfır pA olarak düzeltin. Bir sızdırmazlık testi yapın (100 Hz'de 10 mV test darbesi). Nöronal membranın küçük bir parçasını yırtmak için negatif hava basıncı uygulayın.
- İşitsel nöronların aktif intrinsik özelliklerini test etmek için, hiperpolarize edici ve depolarizan somatik akım enjeksiyonları uygulayın.
NOT: Bu prosedürün örnekleri Ek Video S1, S2'de görselleştirilebilir. Bu prosedürün ayrıntıları video açıklamalarında verilmiştir.
Sonuçlar
Burada gösterilen tüm beyin sapı dilimleri beyin sapı dokusundan (~ 200-300 μm) elde edildi ve 5x objektif ve diferansiyel girişim kontrastı (DIC) optikleri kullanılarak görüntülendi. Kamera diseksiyon mikroskobuna monte edildi ve görüntü yakalama yazılımı ile bir bilgisayara bağlandı (bkz. Bu rakamlar için uydu girişi (sağ paneller) 60x büyütme suyu daldırma hedefi kullanılarak görüntülendi. Dijital görüntüler elde edilirken beyin sapı diliminin tüm alanlarının eşit şekilde büyütülmesini sağlamak için özen gösterildi. Fotoğraflar optimum parlaklık ve odakta çekildi. Beyin sapı dilimlerinin dijital görüntüleri, üst üste binen alana dayalı düzlemsel bir şekilde dikildi ve parlaklık, kontrast ve gri tonlamanın daha fazla ayarlanması için bir masaüstü bilgisayara aktarıldı. Tavuk işitsel beyin sapının temel mikro devreleri önceki çalışmalara göre tanımlanmıştır 1,2,5,13. Mikroskop altında (5x objektif), işitsel çekirdekler, dilimin dorsal bölgeleri boyunca her çekirdeğin etrafında hem ipsilateral hem de kontralateral olarak akan ağır miyelinli sinir liflerine bitişik alan olarak tanımlandı.
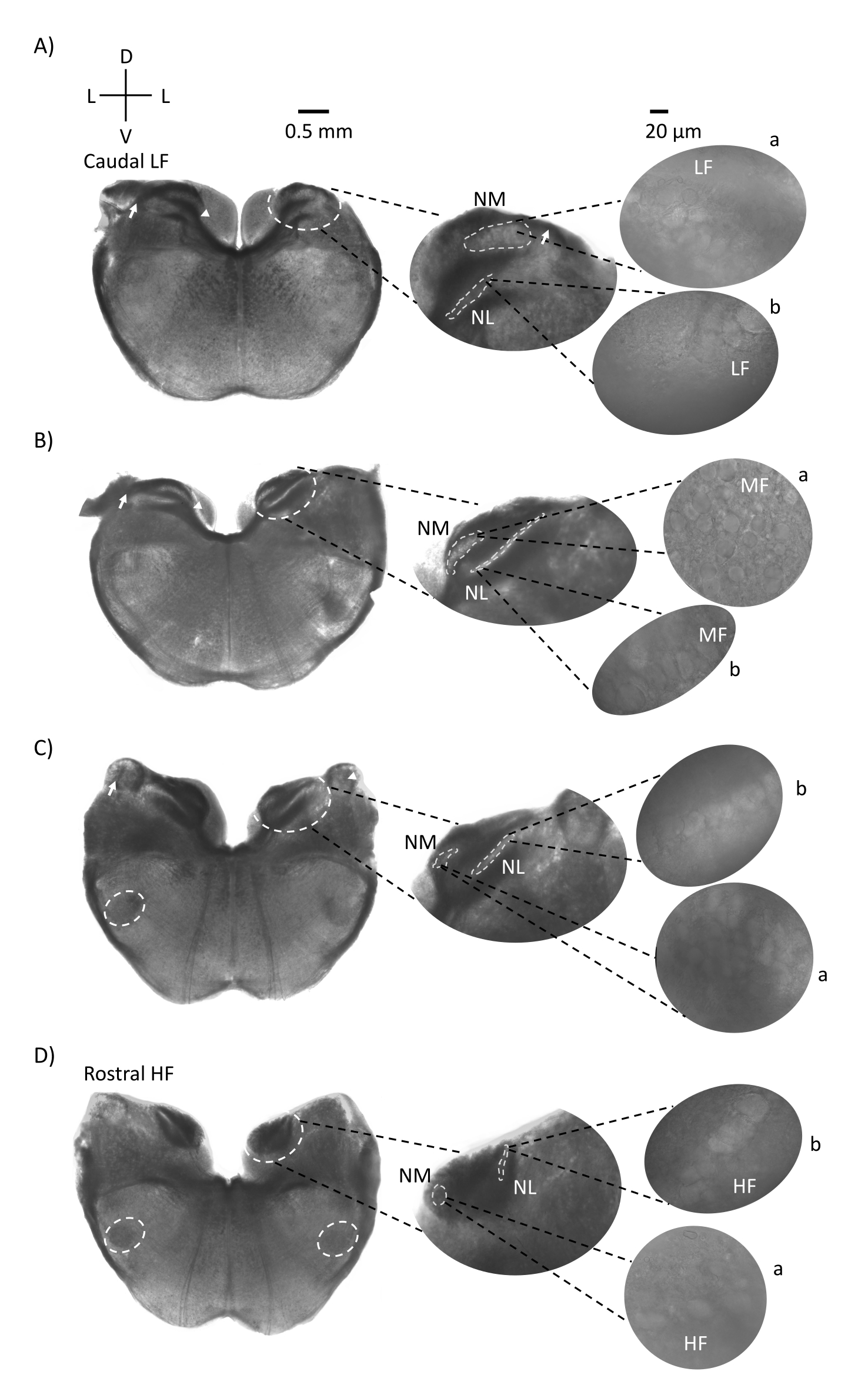
Şekil 1, bir E21 tavuk embriyosundan beyin sapı dokusunun (200-300 μm) geleneksel koronal bölümlerini göstermektedir. Burada gösterilen dört koronal dilim, işitsel beyin sapı çekirdeklerinin en düşük CF işitsel bölgesinden (Şekil 1A, kaudo-lateral) en yüksek CF işitsel bölgesine (Şekil 1D, rostral-medial) ilerleyen göreceli izo-frekans bölgelerini temsil eder. Şekil 1A-D'deki dört koronal dilimin tümü için, etiketli NM ve NL'nin büyütülmüş bölgeleri orta sütunda gösterilir ve şekil panelinin sağ görüşünde büyütülür (60x hedef) (uydu insets'te sırasıyla a ve b). Şekil 1A,B'deki ok, işitme siniri liflerinin girişini gösterir ve ok ucu, dilimin solundaki NM aksonlarının çatallanmasını gösterir. Şekil 1C, çekirdek angularis olarak bilinen başka bir kuş koklear çekirdek yapısını göstermektedir (NA, solda ok ve sağda ok ucu). En rostral iki koronal dilim, koronal dilimin ventral-lateral bölgesi boyunca yer alan superior olivary çekirdeğini (SON) göstermektedir (Şekil 1C, D, beyaz kesikli daireler).
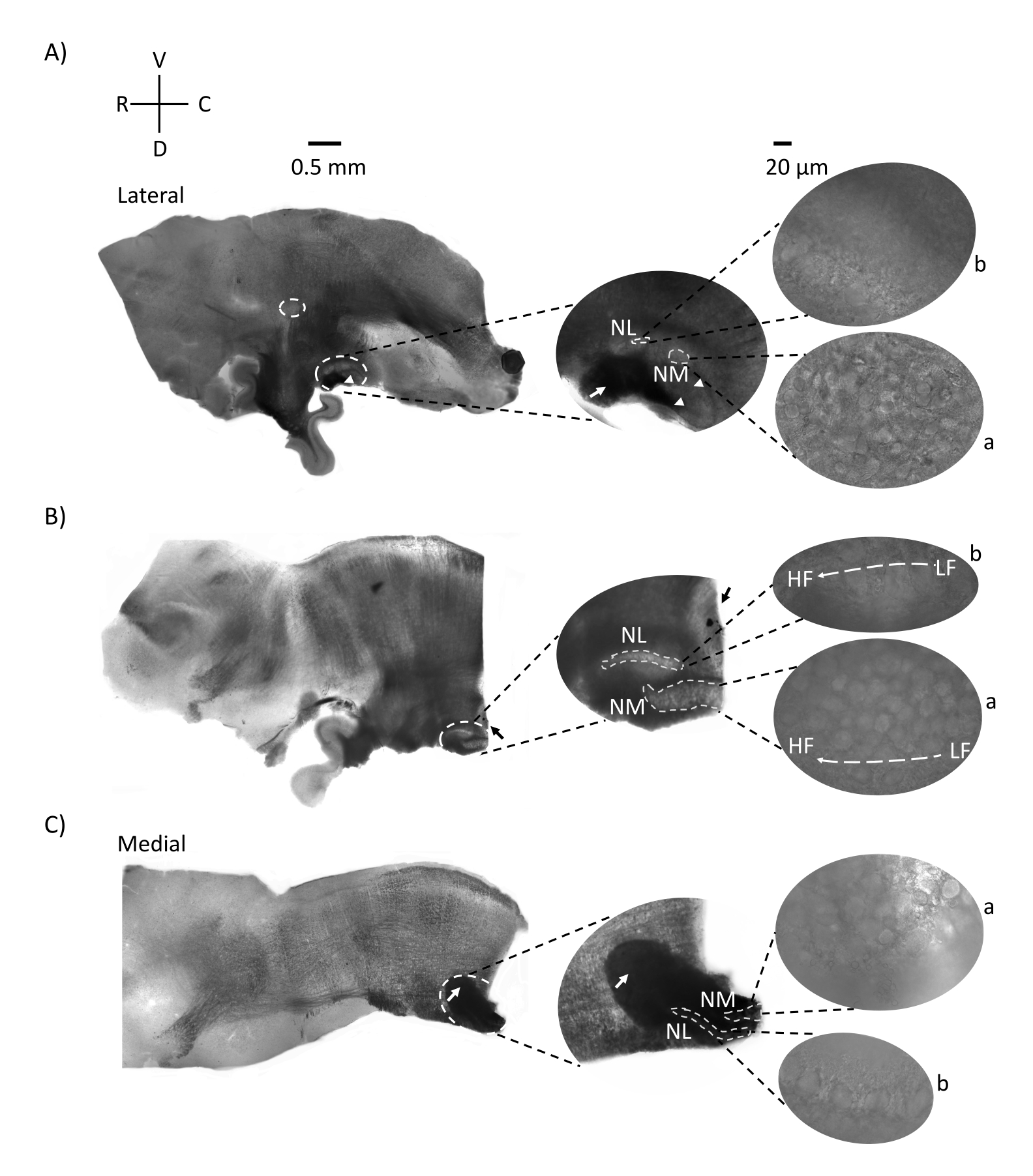
Şekil 2, bir E21 tavuk embriyosundan beyin sapı dokusunun (200-300 μm) sagital bölümlerini göstermektedir. Her üç sagital dilim için de (Şekil 2A-C), etiketli NM ve NL'nin büyütülmüş bölgeleri orta sütunda gösterilir ve şekil panelinin sağ görüşünde büyütülmüş (60x nesnel) (uydu görüntülerinde sırasıyla a ve b). İşitme siniri liflerinin (Şekil 2A, orta ok) daha yüksek büyütmede gözlenen nöron kümesine girdiği (Şekil 2A, orta, küçük, beyaz kesikli daireler ve ok uçları) ve işitsel bölgenin başlangıç noktasını vurguladığı (Şekil 2A, sol, büyük, beyaz kesikli daire ve ok ucu) NM ve NL tanımlanmıştır. SON, en lateral dilimin rostro-lateral bölgesinde tanımlandı (Şekil 2A, küçük, beyaz, kesikli daire). Şekil 2B, rostral-kaudal eksen boyunca NM ve NL'den hem nispeten düşük hem de yüksek CF işitsel bölgeleri içeren genişletilmiş tonotopik bölgeleri göstermektedir (beyaz ana hatlı bölgeler, ayrıca uydu girişine bakınız). Şekil 2C, en medial dilimdeki ipsilateral ve kontralateral aksonal tutamları ve işitsel bölgenin son noktasını (sol ve orta oklar) göstermektedir. Burada gösterilen dilimlerin oryantasyonu, Şekil 1'de görüldüğü gibi dilimlerin geleneksel yönelimi ile çelişmektedir (yani, koronal). Bu, elektrofizyolojik kayıtlar için gerekli olan bir cam pipetin yaklaşımını en iyi şekilde karşılayan yönü göstermek için gerçekleştirildi.
Tonotopik eksenin geniş bir bölgesinin Şekil 2B'de temsil edildiğini doğrulamak için, NM nöronlarından akım-kelepçeli elektrofizyoloji kayıtları yapıldı. Şekil 3, bir koronal dilimden (Şekil 3A, B) ve bir sagital dilimden (Şekil 3C, D, Ek Video S1, S2) kaydedilen olgun (E21) NM nöronlarının fonksiyonel benzerliklerini ve farklılıklarını göstermektedir. Bir koronal dilimin medial ve lateral uçlarından iki NM nöronu seçildi (Şekil 1B'de gösterilen dilime benzer) ve iki NM nöronu, bir sagital dilimde NM'nin rostral ve kaudal uçlarından seçildi (Şekil 2B'de gösterilen dilimde olduğu gibi). Şekil 3A,B, somatik akım enjeksiyonlarına benzer elektrofizyolojik yanıt özelliklerini göstermektedir (-100 pA ila +200 pA, +10 pA artışlar, 100 ms süre). Bu iki NM nöronun ateşleme paterni, bu dilim düzleminde ince farklılıklar gösterir ve orta frekanslı NM nöronları için göreceli izo-frekanslı laminayı gösterir. Şekil 3C, D, ateşleme modellerinin rostral-kaudal eksen boyunca önemli farklılıklara sahip olduğunu ve düşük frekanslı bir NM nörondan (Şekil 3C) yüksek frekanslı bir NM nöronuna (Şekil 3D) nispeten daha yüksek bir tonotopik gradyanı gösterdiğini göstermektedir. Her iki nöron da daha önce bildirildiği gibi basmakalıp ateşleme modelleriyle birlikte sunuldu14,15.
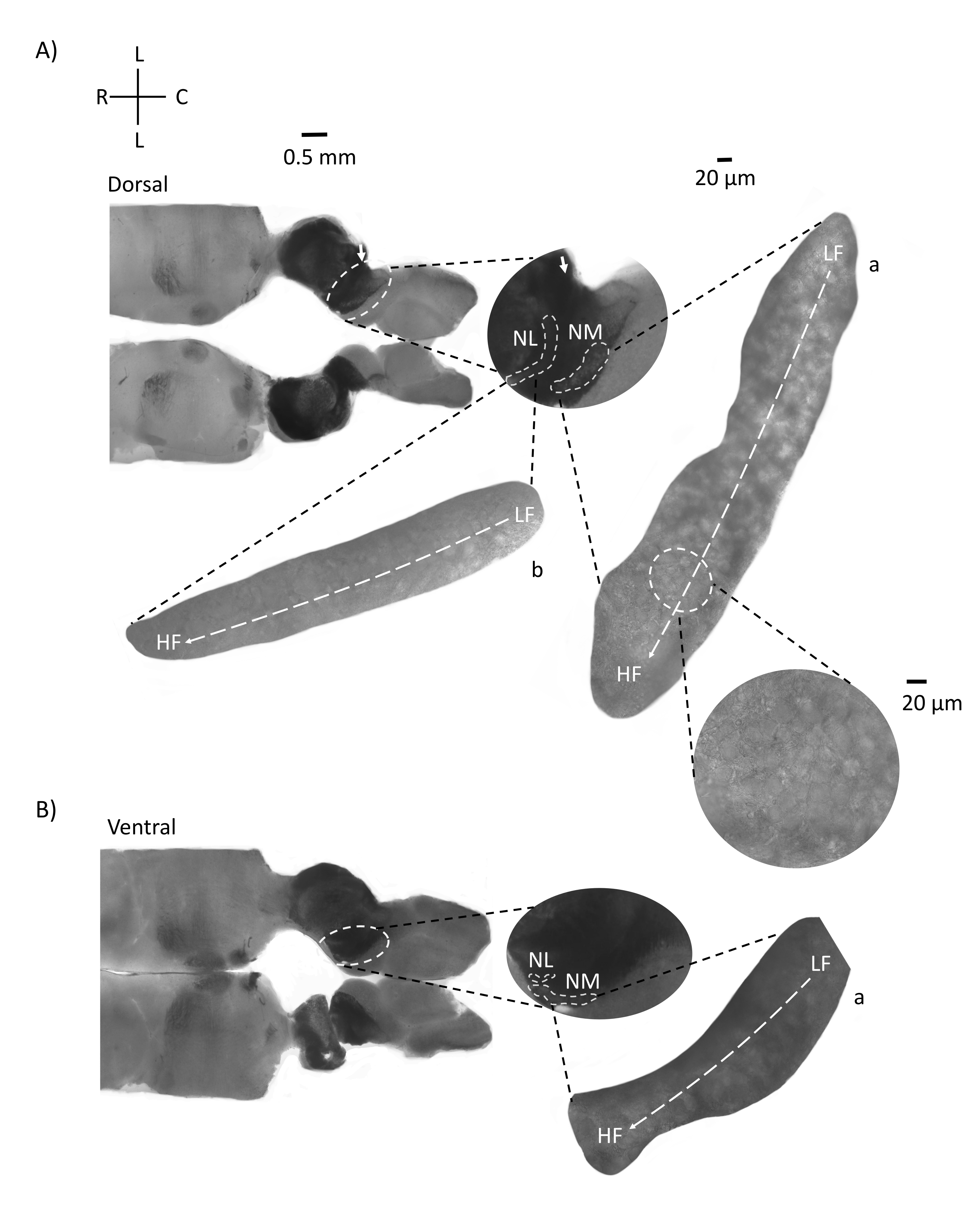
Şekil 4, bir E21 tavuk embriyosunun beyin sapı dokusunun (200-300 μm) yatay bölümlerini göstermektedir. Her iki yatay dilim için de (Şekil 4A,B), etiketli NM ve NL'nin büyütülmüş bölgeleri orta sütunda gösterilir ve şekil panelinin sağ görüşünde büyütülmüş (60x hedef) (uydu insets'te sırasıyla a ve b). Yatay dilimlerde orta hatta doğru NM ve NL saptanmış ve nöronlar lateral-medial eksen boyunca yayılmıştır (Şekil 4A,B, orta, beyaz, kesikli anahat bölgeleri). Büyütülmüş görüntüler tonotopik gradyanın büyük boyutunu göstermektedir. Düşük frekanslı nöronlar kaudo-lateral bölgelerde, yüksek frekanslı nöronlar ise rostral-medial bölgelerdedir (Şekil 4A,B, sağda, uydular). Rostral-kaudal eksen boyunca orta hattan geçen lifler, işitsel çekirdeklerin kontralateral bağlantılarını gösterir, ancak bu liflerin organizasyonu basit bir düzlemde değildir. Bununla birlikte, yatay / enine bir kesitten akut açısal dilimler, bu aksonal lifleri sagital düzleme doğru takip edebilir. Yatay düzlemden akut açılı (45°) 200-300 μM kalınlığında beyin sapı dokusu dilimleri Şekil 5'te gösterilmiştir. İşitsel beyin sapı çekirdekleri, en lateral dilimden başlayıp en medial dilimde biten büyük bir diyagonal yayılım boyunca görülebilir (Şekil 5A-C, etiketli orta paneller, beyaz ana hatlı alan). Ayrıca, NM ve NL bölgelerinin açısal yönelimi ardışık asimetrik dilimlerde de görselleştirilebilir (Şekil 5A-C, etiketli orta paneller, beyaz, kesikli ana hatlı alan). Büyütülmüş görüntüler (60x nesnel), işitsel çekirdeklerin tonotopik eksenini, rostral-medial ila kaudo-lateral eksen boyunca ilerlerken göstermektedir (Şekil 5A-C, sağ, uydu girişi). Şekil 5'teki dilimlerin oryantasyonu Şekil 2'dekine benzer. Görüntülerin geleneksel sunumunu karşılaştırırlar, ancak elektrofizyolojik deneyler için daha uygundurlar.

Şekil 1: Beyin sapının temsili koronal seri bölümleri. (A-D) Sol: kaudalden rostral eksene dilimler, işitsel çekirdekler ve beyaz kesikli bir daire ile işaretlenmiş bağlantı lifleri. Orta kesici uç, çekirdeklerin beyaz kesikli daireler a: NM ve b: NL içinde gösterildiği işitsel bölgenin daha büyük bir görünümüdür. Oklar işitsel sinir afferent liflerini gösterir ve ok ucu A, B'de NM akson bifurkasyonunu gösterir. Ok, C'de NA'yı gösterir. Yanal beyaz kesikli daire C,D'de SON'u gösterir. Sağda: uydu eki bu çekirdekleri 60x hedefte gösterir: a: NM ve b: NL. Kısaltmalar: NM = nucleus magnocellularis; NL = çekirdek laminaris; NA = çekirdek angularis; SON = superior olivary çekirdeği; LF = nispeten düşük frekanslı nöronlar; MF = orta frekanslı nöronlar; HF = yüksek frekanslı nöronlar; D = sırt; L = yanal; V = ventral. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: Beyin sapının temsili sagital seri bölümleri. (A-C) Sol: İşitsel çekirdekleri beyaz kesikli bir daire içinde etiketlenmiş yanal eksenden medial eksene dilimler. Orta kesici uç, beyaz kesikli daireler içinde işaretlenmiş aynı işitsel çekirdek bölgesini daha büyük bir görünümde gösterir. (A) Dilimin ortasındaki beyaz kesikli daire SON'u vurgular; işitsel sinir liflerini gösteren ok ve NA'yı gösteren ok ucu. Dilimin sağ ucundaki koyu siyah nokta bir görüntüleme eseridir. Beyincik bölgeleri sol paneldeki A ve B dilimlerinin her ikisinde de işitsel bölgeye dorsal olarak görülebilir. (B) Oryantasyonu koronal düzleme (dilimleme sırasında) değiştirilen bir sagital dilim. İşitsel bölge mavi boya (siyah ok) ile tanımlandı ve tekrar sagital düzlemde dilimlendi. (A-C) Kesikli beyaz çizgiler altında işaretlenmiş orta uç NM ve NL bölgesi. Sağda: uydu görüntüsü a: NM ve b: NL'yi 60x nesnel büyütmede gözlemlenen gösterir. İşitsel çekirdeklerde LF ve HF tonotopik gradyanı rostro-kaudal eksen boyunca gösterilmiştir. (C) deki karanlık alana işaret eden oklar, medial eksen boyunca orta hat boyunca uzanan ağır miyelinli NM liflerini göstermektedir. Lifler işitsel çekirdeklerin her iki tarafını birbirine bağlar. Kısaltmalar: NM = nucleus magnocellularis; NL = çekirdek laminaris; NA = çekirdek angularis; SON = superior olivary çekirdeği; LF = nispeten düşük frekanslı nöronlar; HF = yüksek frekanslı nöronlar; D = sırt; V = ventral; R = rostral; C = kaudal. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: Mevcut kelepçe modunda somatik akım enjeksiyonlarına (-100 pA ila +200 pA, +10 pA artışlar, 100 ms süre) nöronal yanıtın elektrofizyolojik kayıtları. Nöronlar aynı dilimde, ancak NM'nin aşırı zıt bölgelerinde kayıtlar için seçildi. (A,B) İnce farklılıklarla göreceli izo-frekans özelliklerini gösteren tek bir koronal dilimdeki temsili nöronal yanıtlar. Yanıt özellikleri, bir koronal dilimde NM'nin en medial (A) ve lateral (B) bölgelerinden kaydedilen iki farklı MF nöronunu temsil eder. (C,D) Tek bir sagital dilimden temsili nöronal kayıtlar. Kayıtlar nispeten LF NM yanıtı (C) ve HF NM yanıtı (D) göstererek, tek bir sagital kesit içinde tonotopik gradyandaki önemli farklılıkları vurgulamaktadır. Kısaltmalar: NM = nucleus magnocellularis; LF = nispeten düşük frekanslı nöronlar; MF = orta frekanslı nöronlar; HF = yüksek frekanslı nöronlar. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 4: Beyin sapının temsili yatay seri bölümleri. (A,B) Solda: dorsal boyunca ventral eksene kadar olan dilimler, işitsel çekirdekler beyaz kesikli dairelerle işaretlenmiştir. 8. kranial sinir afferent lifleri, okla işaretlenmiş işitsel çekirdekleri birbirine bağlar. Orta uç, işitsel çekirdek bölgesinin daha geniş bir görünümüdür ve beyaz kesikli çizgiler altında işaretlenmiş işitsel çekirdekler NM ve NL bölgeleri gösterilmiştir. İşitsel çekirdeklerin açık bir topolojik hareketi A,B'de görülebilir. (A,B) Sağda: a: NM ve b: NL'yi gösteren büyük uydu görünümü. Sağ ek, 60x nesnel büyütmede gözlenen işitsel çekirdekleri ve bir kaudo-lateral ila rostral-medial eksen boyunca LF'den HF'ye kavisli topolojik ekseni gösterir. Kısaltmalar: NM = nucleus magnocellularis; NL = çekirdek laminaris; LF = nispeten düşük frekanslı nöronlar; HF = yüksek frekanslı nöronlar; L = yanal; R = rostral; C = kaudal. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 5: Temsili yatay/enine akut açısal (45°) seri kesitler. (A-C) Sol: beyin sapının seri bölümleri, beyaz kesikli daire ile işaretlenmiş işitsel çekirdekler. Orta kesici uç, işitsel bölgenin daha büyük bir görünümüdür. (A) Orta uç, bu dilimlerdeki NM ve NL nöronlarının en büyük yayılımını gösterir. (B,C) Orta ek: Beyaz kesikli çizgilerle işaretlenmiş işitsel çekirdekler, (A-C) ile karşılaştırıldığında kademeli topolojik değişim gösterir. Sağda: 60x objektif büyütmede işitsel çekirdekler a: NM ve b: NL'yi gösteren uydu girişi. NM ve NL'de LF'den HF bölgelerine tonotopik eksen, lateral dilimlerden medial dilimlere açısal olarak döner. Kısaltmalar: NM = nucleus magnocellularis; NL = çekirdek laminaris; LF = nispeten düşük frekanslı nöronlar; HF = yüksek frekanslı nöronlar; V = ventral; R = rostral; D = sırt; C = kaudal. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}
Ek Video S1: Hiperpolarize edici ve depolarize edici somatik akım enjeksiyonları. Düşük frekanslı ve yüksek frekanslı bir nörondan akım kelepçesi modunda 100 ms somatik akım enjeksiyonlarına yanıt özellikleri. Nöronlar aynı sagital beyin sapı diliminden seçildi. Enjeksiyonlar +10 pA'lık artışlarla, 100 ms zaman süresiyle -100 ila +200 pA arasında değişir. Aksiyon potansiyelleri, yeterli depolarize edici mevcut adımlara yanıt olarak görülür. Video, Şekil 3C'de gösterilen son izlere karşılık gelir. Bu Dosyayı indirmek için lütfen tıklayınız.
Ek Video S2: Hiperpolarize edici ve depolarize edici somatik akım enjeksiyonları. Ek Video S1'e benzer şekilde, bu video da düşük frekanslı ve yüksek frekanslı bir nörondan mevcut kelepçe modunda 100 ms somatik akım enjeksiyonlarına tepki özelliklerini gösterir. Nöronlar aynı sagital beyin sapı diliminden seçildi. Enjeksiyonlar +10 pA'lık artışlarla, 100 ms zaman süresiyle -100 ila +200 pA arasında değişir. Aksiyon potansiyelleri, yeterli depolarize edici mevcut adımlara yanıt olarak görülür. Video, Şekil 3B'de gösterilen son izlere karşılık gelir. Bu Dosyayı indirmek için lütfen tıklayınız.
Tartışmalar
Tavuk embriyonik beyin sapı dokusunun koronal bölümleri, on yıllardır göreceli bireysel izo-frekanslı laminanın incelenmesine izin vermiştir 1,2,5. Bununla birlikte, tavuk işitsel beyin sapının tonotopik (yani frekans) organizasyonu topolojik olarak kıvrımlıdır ve spesifik araştırma sorusuna bağlı olarak diğer anatomik eksenlerde daha erişilebilir olabilir. Bireysel izo-frekans bölgelerine ait anatomik ve fizyolojik soruları araştırmak için yeterli olmasına rağmen, tonotopik varyasyonların incelenmesi ve daha büyük işitsel beyin sapı alanlarında gelişimi koronal kesitlerle sınırlıdır. Bu sınırlamanın üstesinden gelmek için, bu protokol, bireysel bir beyin sapı bölümünde maksimum tonotopik özellikler ve gradyanlar sergileyen işitsel beyin sapı dokusunun ek örneklerini sağlamak için sagital, yatay ve yatay / enine düzlemlerdeki yaklaşımları açıklar.
İşitsel beyin sapı bölgelerinin Sagital bölümleri, farklı tonotopik alanların koronal kesitlere kıyasla dilim içinde daha geniş bir bölgeye dağıldığını göstermektedir (sagital işitsel alan = ~ 300-600 μm, koronal işitsel alan = ~ 200-350 μm). Örneğin, NM ve NL bölgeleri, sagital kesitlerde rostro-kaudal eksen boyunca daha geniş bir alan üzerinde görselleştirildi (örneğin, Şekil 2B) ve bu anatomik eksen boyunca uzanan fonksiyonel tonotopik gradyan büyük ölçüde tek bir sagital dilim içinde yer aldı. Bu, daha önce14,15 olarak bildirilen rostral-kaudal gradyan boyunca değişen intrinsik nöronal farklılıkların akım-kelepçe kayıtları ile daha da doğrulanmıştır (örneğin, Şekil 3C, D). Tonotopik eksen boyunca anatomik ve immünohistokimyasal özellikleri vurgulayan gelecekteki deneyler, tek bir sagital dilim düzlemi içindeki işitsel özelliklerin bilinen gradyanlarını daha fazla araştırabilir. Bunlar, bunlarla sınırlı olmamak üzere, dendritik mimarinin bilinen gradyanları ve daha önce ardışık koronal bölüm16'da gösterilen NM ve NL'nin içsel özellikleri olan MAP2 boyama ve potasyum kanal ekspresyon paternlerini içerir.
İşitsel beyin sapı bölgelerinin yatay bölümleri, NM ve NL'nin orta hatta doğru yerleştirildiğini göstermektedir. İşitsel aksonal liflerin bir kısmı yatay düzleme çapraz veya dik olarak uzanır (Şekil 4). Bu lifleri, sagital düzleme 45 ° akut açısal bir dilim yapılarak takip edilebilir. Ortaya çıkan yatay / enine dilimler sagital veya yatay dilimlerden daha büyüktü ve uzun aksonal lifler hem ipsilateral hem de kontralateral taraflar için rostro-kaudal eksen boyunca ilerledi. Hem NM hem de NL, daha büyük bir diyagonal bölgede (~ 400-700 μm) görselleştirilebilir, böylece kontralateral bağlantılar lateral-medial bir eksen boyunca görselleştirilebilir. Ek olarak, yatay/enine dilim düzlemi, işitsel bölgelerin ve ortaya çıkan tonotopik gradyanın nasıl açısal bir dönüş yaptığını da gösterir (Şekil 5). Kontralateral bağlantıların daha geniş bir alanda açısal maruziyeti, bu dilimleri elektrofizyolojik stimülasyon ve mikrodevre çalışmaları için geleneksel koronal dilimlerden daha uygun hale getirir.
Ek avantajlar
İşitsel mikrodevrelerin oluşumu, nöronal sağkalımı, sinaptogenezi, aksonal farklılaşmayı, dendritik mimariyi ve olgunlaşmayı teşvik eden ipuçlarının mekansal zamansal koordinasyonunu gerektirir. Bu nedenle, tavuk embriyosu işitsel mikrodevresinin alternatif bir beyin sapı bölümleri aşağıdaki araştırma konuları için kullanılabilir: topografik olarak farklı boyutlarda nöronların morfolojik organizasyonu; tüm işitsel ve vestibüler çekirdeklerin bağlantılarını organize etmek ve haritalamak; izo-frekans ve tonotopik düzlemlerde devre bileşenlerinin aktivite paternlerinin tanımlanması ve karakterizasyonu; uyarıcı ve inhibitör mikrodevrelerin topografik organizasyonu ve uzmanlaşmış nöron popülasyonları (çekirdekler) ile ilişkileri; işitsel çekirdek nöronların uzamsal konumu ve prediktif CF17; spesifik tonotopik nöronal tiplerin sistematik olarak hedeflenmesi; progenitör hücrelerin izlenmesi ve korunmuş çekirdeklere gelişimleri; nöronal devrelerin evrimine hücrelerin genetik soyu18; türler arasında karşılaştırmalı beyin sapı anatomisi; Deiter'in vestibüler kompleksi (DC)19 gibi vestibüler devrelerin araştırılması; ve vestibüler çekirdekler arasında senkron ve çapraz konuşma.
Farklı dilim düzlemleri kullanan çok yönlü bir yaklaşım, beyin sapı mikrodevrelerinin bilinmeyen anatomik ve biyofiziksel özellikleri hakkındaki temel soruları cevaplamaya yardımcı olabilir. İyi bir örnek, majör işitsel çekirdekler (NM, NA, NL ve SON) ile lateral lemniscusun dorsal çekirdeği (LLDp), semilunar çekirdeği (SLu)20 ve teğetsel çekirdek (TN)3 dahil olmak üzere vestibüler çekirdekler arasındaki ilişkidir. Bununla birlikte, bu protokolün ve bu dilim tabanlı çalışmaların bazı sınırlamaları vardır.
Önlemler ve sınırlamalar
Deneyleri yapan kuruma bağlı olarak, etik kurallar ve tavuk embriyolarının işlenmesi farklılık gösterebilir. Ulusal Sağlık Enstitüleri Laboratuvar Hayvanlarının Bakımı ve Kullanımı için Kılavuzlar hızlı kafa kesmeye izin verirken, tavuk embriyosu ötenazisi21 için alternatif yöntemler vardır. Erken gelişen tavuk embriyosu beyin sapı dokusu, yaşlı embriyolara kıyasla yumuşak ve hassastır. Yüzeyde, bunları çıkarırken ekstra dikkat gerektiren birkaç bağlantıya ve kan damarına sahiptir. Doku buz gibi soğuk dACSF'de tutulmalı ve canlılığı arttırmak için %95 O 2/%5 CO2 ile perfüze edilmelidir.
Sagital dilimleme yöntemi sadece ipsilateral tonotopi için yararlıdır. Bu dilimleme yöntemi, kullanımı güvencesiz olabilecek koronal dilimlerden daha büyük dilimler sağlar. Bununla birlikte, başka bir yerde ayrıntılı olarak açıklanan çapraz iğne yöntemlerini kullanarak dilimleri kesebilirsiniz22. % 4 LMP agaroz bloğu gömülü beyin sapı kullanmak, hassas yapıları dilimler halinde koruyabilir, ancak aşırı sıcak agaroz dökülmemesine dikkat edilmelidir. Agaroz bloke edilmiş beyin sapını ~ 1 dakika boyunca soğutulmuş bir ortama yerleştirerek hızlı bir şekilde ayarlamak, dilimleri elektrofizyolojik kayıtlar için daha uygun hale getirir.
Süper yapıştırıcının aşırı miktarlarda uygulanması toksik olabilir. Minimum düzeyde uygulanmalı ve fazla miktarlar dACSF'yi değiştirerek derhal yıkanmalıdır. Akut açısal (45 °) dilimler için, agaroz bloğunun açısını kesmek kritik öneme sahiptir; agaroz bloğunu keskin bir bıçakla keserken ön açıyı görmek için bir ayna kullanılabilir. Ticari olarak temin edilebilen bıçaklar, alkolle silinmesi ve kullanımdan önce kurutulması gereken bir balmumu kaplamasına sahip olabilir. Aksonal fiber tutamları kortikal veya matris dokudan daha sert olduğu için vibratom kesme hızı ve frekansı için optimizasyon gereklidir. Yüksek genliğin korunması ve soğutulmuş diseksiyon solüsyonunun kullanılması doku hasarını önleyebilir.
Tüm çözeltiler taze hazırlanmalı ve % 95 O 2/5% CO 2 köpürtüldükten sonra ACSF'ye Ca2 + ve Mg2 + eklenmelidir. Aksi takdirde, Ca 2+ yağışolabilir. Dilimleri vibratom içinde nazikçe işlemek için bir boya fırçası kullanılmalıdır. Mümkünse toplam dilimleme süresini 15 dakikanın altında tutun. Beyin sapı dilimlerini manevra yapmak için cam Pasteur pipet kullanılabilir.
Elektrofizyolojide kullanılan dilimlerle temas eden cam eşya ve ekipmanlar için deterjan veya aşındırıcı yıkama maddeleri kullanmayın. Çekilen görüntüler, diferansiyel girişim kontrastı (DIC) optikleri altında 200-300 μM kalınlığında dokunun görünümünü temsil etmektedir. Görsel kalite, immünohistokimya veya elektron mikroskobundan daha düşük olacaktır, ancak bir deneycinin elektrofizyolojik kayıtlar yaparken ne göreceğini doğru bir şekilde yansıtır.
Alternatif bir anatomik eksen boyunca mikrodevrelerin erken gelişimi ile ilgili çalışmalar, ister dorsal-ventral, rostral-kaudal veya ipsilateral-kontralateral olsun, tavuk işitsel beyin sapında sınırlıdır. Bunun bir nedeni, beyin sapındaki transkripsiyonel kodların ve tonotopik gelişimin düzenlenmesinin rolünün hala tam olarak anlaşılamamasıdır. Yukarıdan aşağıya modülasyon ve spontan aktivite gibi fonksiyonel fenomenler, in vitro aktiviteyi gözlemlerken sıklıkla kaybolur. Bununla birlikte, in vivo araştırma, yalnızca bu dilim koşullarında mümkün olan spesifik ve doğrudan tek nöron kayıtları ile tamamlanmaktadır. Farklı yönelimler boyunca beyin sapı dokusu elde etmenin iyileştirilmesi, tavuk işitsel beyin sapı mikrodevresindeki tonotopik gradyanların gelişimi ve karmaşıklığı hakkında anlayışlı bilgiler sağlayabilir.
Açıklamalar
Tüm yazarlar araştırmanın herhangi bir ticari veya mali çıkar olmaksızın yürütüldüğünü ve herhangi bir çıkar çatışmasının bulunmadığını beyan ederler.
Teşekkürler
Bu çalışma NIH/NIDCD R01 DC017167 hibesi tarafından desteklenmektedir. Kristine McLellan'a makalenin önceki bir versiyonu hakkında editoryal yorumlar sağladığı için teşekkür ederiz.
Malzemeler
| Name | Company | Catalog Number | Comments |
| Adobe photoshop 2021 | Adobe | ||
| Anti-vibration table 30"x 36" - OTMC - 63533 | TMC | ||
| Cell sens standard software | OLYMPUS | ||
| Digidata 1440A | MOLECULAR DEVICES | ||
| Digital amplifier multiclamp 700B | MOLECULAR DEVICES | ||
| DSK line-up linearslicer pro7 | TED PELLA, INC | ||
| Micromanipulator MPC-385 / OSI-MPC-385-2 | OLYMPUS AMERICA INC | ||
| Micropipette puller P-97 | SUTTER INSTRUMENTS | ||
| Microscope BX51W1 | OLYMPUS AMERICA INC | ||
| MS ICE software | Microsoft Corporation | ||
| Ohaus balance model AV212 | Ohaus Adventurer | ||
| Olympus DPSI0 /DPS80 camera | OLYMPUS | ||
| pClamp and Axoclamp data Acquisition Softwares | MOLECULAR DEVICES | ||
| pH meter lab 850 benchtop | SCHOTT INSTRUMENTS | ||
| Sharp stainless blade | Dorco/Personna | ||
| Vapor pressure osmometer model 5600 | WESCOR INC | ||
| Water purification systems Smart2pure 6UV/UF | Thermo Scientific | ||
| Chemicals- list | |||
| Agrose Low melt IB70051 | IBI SCIENTIFIC | ||
| CaCl2 (Calcium Chloride) | ACROS organics | ||
| Cynergy instant adhesive CA6001 | Resinlab | ||
| Dextrose (D-(+)-glucose) | VWR Life Science | ||
| Ethyl alcohol | IBI SCIENTIFIC | ||
| KCl (Potassium Chloride) | Amresco.Inc | ||
| MgCl2 (Magnesium Chloride) | Sigma-Aldrich | ||
| NaCl (Sodium Chloride) | Amresco.Inc | ||
| NaH2PO4 (Sodium Dihydrogen Phosphate) | Amresco.Inc | ||
| NaHCO3 (Sodium Bicarbonate) | Amresco.Inc |
Referanslar
- Rubel, E. W., Parks, T. N. Organization and development of brain stem auditory nuclei of the chicken: tonotopic organization of n. magnocellularis and n. laminaris. Journal of Comparative Neurology. 164 (4), 411-433 (1975).
- Rubel, E. W., et al. Organization and development of brain stem auditory nuclei of the chicken: ontogeny of n. magnocellularis and n. laminaris. Journal of Comparative Neurology. 166 (4), 469-489 (1976).
- Shao, M., et al. Spontaneous synaptic activity in chick vestibular nucleus neurons during the perinatal period. Neuroscience. 127 (1), 81-90 (2004).
- Fukui, I., Ohmori, H. Tonotopic gradients of membrane and synaptic properties for neurons of the chicken nucleus magnocellularis. Journal of Neuroscience. 24 (34), 7514-7523 (2004).
- Sanchez, J. T., Seidl, A. H., Rubel, E. W., Barria, A. Preparation and culture of chicken auditory brainstem slices. Journal of Visualized Experiments. (49), e2527 (2011).
- Sanchez, J. T., Lu, Y., Fay, R. R., Popper, A. N., Cramer, K., Coffin, A. Glutamate signaling in the auditory brainstem. Auditory Development and Plasticity: Springer Handbook of Auditory Research. 64 (4), 75-108 (2017).
- Parks, T. N. Morphology of axosomatic endings in an avian cochlear nucleus: nucleus magnocellularis of the chicken. Journal of Comparative Neurology. 203 (3), 425-440 (1981).
- Jhaveri, S., Morest, D. K. Sequential alterations of neuronal architecture in nucleus magnocellularis of the developing chicken: a Golgi study. Neuroscience. 7 (4), 837-853 (1982).
- Carr, C. E., Boudreau, R. E. Central projections of auditory nerve fibers in the barn owl. Journal of Comparative Neurology. 314 (2), 306-318 (1991).
- Köppl, C. Auditory nerve terminals in the cochlear nucleus magnocellularis: differences between low and high frequencies. Journal of Comparative Neurology. 339 (3), 438-446 (1994).
- Fukui, I., et al. Improvement of phase information at low sound frequency in nucleus magnocellularis of the chicken. Journal of Neurophysiology. 96 (2), 633-641 (2006).
- Wang, X., et al. Postsynaptic FMRP regulates synaptogenesis in vivo in the developing cochlear nucleus. Journal of Neuroscience. 38 (29), 6445-6460 (2018).
- Lu, T., Cohen, A. L., Sanchez, J. T. In ovo electroporation in the chicken auditory brainstem. Journal of Visualized Experiments. (124), e55628 (2017).
- Hong, H., Sanchez, J. T. Need for speed and precision: structural and functional specialization in the cochlear nucleus of the avian auditory system. Journal of Experimental Neuroscience. (12), 1-16 (2018).
- Hong, H., et al. Diverse intrinsic properties shape functional phenotype of low-frequency neurons in the auditory brainstem. Frontiers in Cellular Neuroscience. 12, 1-24 (2018).
- Wang, X., Hong, H., Brown, D. H., Sanchez, J. T., Wang, Y. Distinct neural properties in the low-frequency region of the chicken cochlear nucleus magnocellularis. eNeuro. 4 (2), 1-26 (2017).
- Tabor, K. M., et al. Tonotopic organization of the superior olivary nucleus in the chicken auditory brainstem. Journal of Comparative Neurology. 520 (7), 1493-1508 (2012).
- Lipovsek, M., Wingate, R. J. Conserved and divergent development of brainstem vestibular and auditory nuclei. Elife. 7, 40232 (2018).
- Passetto, M. F., et al. Morphometric analysis of the AMPA-type neurons in the Deiter's vestibular complex of the chick brain. Journal of Chemical Neuroanatomy. 35 (4), 334-345 (2008).
- Curry, R. J., Lu, Y. Intrinsic properties of avian interaural level difference sound localizing neurons. Brain Research. 1752, 147258 (2021).
- Aleksandrowicz, E., Herr, I. Ethical euthanasia and short-term anesthesia of the chick embryo. ALTEX - Alternatives to Animal Experimentation. 32 (2), 143-147 (2015).
- Palkovits, M. Isolated removal of hypothalamic or other brain nuclei of the rat. Brain Research. 14 (59), 449-450 (1973).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır