
Method Article
Нарезка слухового ствола мозга эмбриональной курицы для оценки тонотопических градиентов и микросхем
В этой статье
Резюме
Здесь мы представляем протокол получения некорональных слуховых срезов ствола мозга куриного эмбриона для исследования тонотопических свойств и траекторий развития в пределах одного среза ствола мозга. Эти срезы включают сагиттальные, горизонтальные и горизонтальные/поперечные участки, охватывающие более крупные тонотопные области в пределах отдельной плоскости среза, чем традиционные корональные сечения.
Аннотация
Куриный эмбрион является широко принятой животной моделью для изучения слухового ствола мозга, состоящей из узкоспециализированных микросхем и нейронной топологии, дифференциально ориентированной вдоль тонотопической (т.е. частотной) оси. Тонотопическая ось позволяет сегрегированное кодирование высокочастотных звуков в рострально-медиальной плоскости и низкочастотное кодирование в каудо-боковых областях. Традиционно корональные срезы ствола мозга эмбриональной ткани позволяют изучать относительную индивидуальную изочастотную пластинку. Хотя этого достаточно для исследования анатомических и физиологических вопросов, относящихся к отдельным изочастотным областям, изучение тонотопных вариаций и их развития в более крупных слуховых областях ствола мозга несколько ограничено. Этот протокол сообщает о методах нарезки ствола мозга из куриных эмбрионов, которые охватывают более крупные градиенты частотных областей в нижнем слуховом стволе мозга. Использование различных методов нарезки слуховой ткани ствола мозга курицы позволяет проводить электрофизиологические и анатомические эксперименты в пределах одного среза ствола мозга, где более крупные градиенты тонотопических свойств и траекторий развития лучше сохраняются, чем корональные срезы. Несколько методов нарезки позволяют улучшить исследование разнообразных анатомических, биофизических и тонотопических свойств слуховых микросхем ствола мозга.
Введение
Куриный эмбрион является ценной исследовательской моделью для изучения основных биологических вопросов в многочисленных и разнообразных научных областях, включая клеточную биологию, иммунологию, патологию и нейробиологию развития. Микросхема слухового ствола мозга курицы является отличным примером узкоспециализированной схемы, которую можно понять с точки зрения слуховой морфологии и физиологии. Например, Рубель и Паркс (1975) впервые описали тонотопическую ориентацию (т.е. градиент частоты) куриного ядра magnocellularis (NM) и ядра laminaris (NL) как линейную функцию по оси ядер, ориентированную ~ 30° по отношению к сагиттальной плоскости. Отдельные нейроны в NM и NL кодируют свою лучшую звуковую частоту, известную как их характеристическая частота (CF), вдоль рострально-медиальной плоскости до каудо-латеральной области. Высокочастотно-чувствительные нейроны находятся в рострально-медиальной области, а низкочастотно-чувствительные нейроны расположены каудо-латерально. Таким образом, традиционные методы рассечения слуховой ткани ствола мозга для изучения тонотопических свойств использовали последовательные корональные срезы. Действительно, слуховые микросхемы развивающихся куриных эмбрионов были созданы в качестве модельной системы для изучения обработки сигналов тонотопических слуховых функций через последовательные каудально-ростральные корональные плоские срезы ствола мозга в течение десятилетий 1,2,3,4,5,6.
Однако тонотопическая организация НМ и НЛ топологически и морфологически запутанна. Входы слухового нерва распределены таким образом, что входы с высоким cf заканчиваются в эндбульбоподобных структурах, которые покрывают, по меньшей мере, одну четверть соматической окружности аденритной NM-клетки. И наоборот, входы с низким cf организованы не с концевыми лампоподобными терминалами, а с несколькими бутон-синапсами на дендритах нейронов NM. Средние входы CF заканчиваются как концевой колбой, так и бутоноподобными синапсами 4,7,8,9,10,11,12. В NL очень стереотипный дендритный градиент проявляется не только в дендритной длине, но и в дендритной ширине. Этот уникальный дендритный градиент тесно связан с тонотопической осью. Дендриты подвергаются 11-кратному увеличению длины и пятикратному увеличению ширины от нейронов с высоким и низким CF соответственно6. Чтобы преодолеть такие запутанные распределения этих ядер в корональных срезах, этот протокол описывает подходы к рассечению в сагиттальной, горизонтальной и горизонтальной/поперечной плоскостях. Эти методы нарезки предоставляют примеры слуховой ткани ствола мозга, которые проявляют максимальные тонотопические свойства в отдельной плоскости среза.
протокол
Все процедуры были одобрены Институциональными комитетами по уходу и использованию животных Северо-Западного университета (IACUC) и были выполнены в соответствии с Руководящими принципами Национальных институтов здравоохранения по уходу и использованию лабораторных животных. Протоколы вскрытия и подготовки ткани ствола мозга соответствуют предыдущим протоколам 5,13.
1. Обработка яиц
- Приобретите оплодотворенные яйцеклетки (Gallus gallus domesticus) у местного поставщика животных, одобренного IACUC.
- Храните яйца сразу по прибытии в холодильнике при 14 °C и инкубируйте в течение 5 дней.
ПРИМЕЧАНИЕ: Жизнеспособность эмбриона значительно снижается через 1 неделю. - Стерилизуйте яйца 70% этанолом перед инкубацией при 38 ± 1 °C и влажности ~50%.
2. Состав и препарат искусственной спинномозговой жидкости (ОКСДП)
- Смешайте следующие химические вещества в 1 л 18,2 МОмсм dH2O для создания 10x ACSF запасного раствора: NaCl (хлорид натрия) 130 мМ, NaHCO3 (бикарбонат натрия) 26 мМ, KCl (хлорид калия) 2,5 мМ,2PO4 (дигидрофосфат натрия) 1,25 мМ, декстроза (D-(+)-глюкоза) 10 мМ. Храните раствор в холодильнике.
- Готовят MgCl2 (магния хлорид) 1 M и CaCl2 (кальция хлорид) 1 M растворы отдельно в 18,2 МОмсм dH2O и хранят в холодильнике.
- Непосредственно перед применением разводят 10x ACSF до 1x и пузырьки непрерывно с 95% O2/5% CO2 в течение 15-20 мин и добавляют MgCl2 и CaCl2. Для получения ACSF и dACSF (рассечение ACSF) подстройте к конечной концентрации Mg2+ 1 мМ, Ca2 + 3 мМ и Mg2 + 3 мМ, Ca2 + 1 мМ соответственно.
- Установите скорость пузырьков для ACSF таким образом, чтобы рН составлял 7,2-7,4 с осмоляльностью от 300 до 310 мОсм/л.
ПРИМЕЧАНИЕ: Помещение ACSF в ледяную ванну во время пузырьков полезно для поддержания низкой температуры раствора, которая будет поддерживать структурную целостность тканей при рассечении.
3. Агароза (5%) блочный препарат
- Смешать 5 г агарозы в 100 мл ДАКСС. Используйте водяную баню при температуре 100 °C или микроволновую печь в течение 2-3 минут, помешивая каждые 30 с, чтобы предотвратить комочки, пока агароза полностью не растворится и не начнет бурлить.
- Перелить расплавленную агарозу в пустую чашку Петри толщиной до 5 мм и держать при комнатной температуре до застывания. После схватывания запечатайте чашку Петри парапленкой и храните при температуре 4 °C.
- Нарежьте агарозу на кубические блоки острым лезвием и используйте их в момент рассечения.
4. Протокол рассечения и изоляция слухового ствола мозга
- Очистите область рассечения с помощью 70% раствора этилового спирта.
- Приклейте опорный или угловой блок агарозы на лоток вибратома.
- Выбирайте яйца нужного возраста (Е20 и Е21 в настоящем протоколе). Обрабатывайте и инкубируйте яйца в соответствии с протоколами, перечисленными выше, как на шаге 1.
- Найдите заполненное воздухом пространство, поместив яйцо под яркий свет и найдя это пространство на большей или круглой стороне яйца.
- Акклиматизируйте яйца до комнатной температуры, расколойте скорлупу над заполненным воздухом пространством и обнажите мембранный мешок.
- Сделайте аккуратный разрез в мешочке, чтобы обнажить клюв.
- Скальпелем аккуратно вытяните шею и голову из яйца.
- Быстро обезглавить голову острыми ножницами.
- После обезглавливания очистите голову охлажденным льдом dACSF, чтобы удалить лишнюю кровь из прокладки для рассечения.
- Держите голову устойчиво в охлажденном льдом dACSF и сделайте ростро-каудальный разрез. Начните разрез сзади и между глазами и следите за длиной собранной шеи.
ПРИМЕЧАНИЕ: Молодым эмбрионам может потребоваться меньшее давление при выполнении разреза. - Отделите кожу, чтобы обнажить череп.
- Вырежьте череп за глазом в среднем и боковом направлении. Сделайте это для обоих полушарий.
ПРИМЕЧАНИЕ: Этот шаг помогает отделить ростральную часть черепа от прикрепленного мозга, сохраняя при этом ткань мозга нетронутой5. - Нарежьте ростральную часть черепа. Поместите лезвие за глаза и сделайте быстрый разрез.
ПРИМЕЧАНИЕ: Может потребоваться усилие, чтобы аккуратно разрезать прикрепленный череп. - Погрузите голову в блюдо с холодным dACSF.
- Используя небольшую пару ножниц, сделайте средние и боковые разрезы в каудальной области черепа, чтобы попытаться отделить мозг от черепа, не вызывая повреждения тканей.
- Осторожно обнажите ствол мозга и мозжечок. Втяните спинную область всего черепа, аккуратно удалите ствол мозга и обнажите его с помощью тонкой кисти с мягкими санками. Используйте изогнутые щипцы, чтобы очистить ствол мозга от соединительной ткани и кровеносных сосудов. Уделите особое внимание8-й области черепного нерва и обязательно оставьте небольшую длину неповрежденных нервных волокон с обеих сторон.
- Отделите ствол мозга от мозжечка, разрезав цветоносы и аккуратно удалив кровеносные сосуды. Обрежьте ствол мозга дополнительных кровеносных сосудов.
ПРИМЕЧАНИЕ: Убедитесь, что вся процедура выполняется в охлажденном льдом dACSF, непрерывно пузырящемся с карбоксигеном (95% O2/5% CO2).
5. Нарезка вибратома
ПРИМЕЧАНИЕ: На следующих этапах задняя часть ткани должна быть поддержана кубическим куском агарозы.
- Поместите лезвие вибратома вдоль горизонтальной оси и приклейте ствол мозга на лоток для нарезки. Приклеиваем ростральную сторону, сохраняя рострально-каудальную ось вертикальной для корональных срезов.
- Держите латерально-медиальную ось вертикальной для сагиттальных срезов.
- Приклеиваем вентральную сторону, сохраняя дорсально-вентральную ось вертикальной для горизонтальных срезов.
- Чтобы достичь острой угловой сагиттально-горизонтальной плоскости, склеивают вентральную сторону ствола мозга, сохраняя вентрально-дорсальную ось вертикальной на гипотенузной поверхности блока агарозы, которая разрезается под углом 45°. Приклейте противоположную поверхность блока агарозы лицом к нарезному лотку и держите рострально-каудальную ось параллельно кромке лезвия.
6. Работа с хрупкими или крупными кусками ткани ствола мозга
- В альтернативном подходе к шагу 5 поместите изолированный ствол мозга в агарозу с низкой температурой плавления (LMP) с 4% при ~ 40 ° C в чашку Петри размером 35 мм х 10 мм.
- После выливания агарозы на погруженный ствол мозга поместите чашку Петри на лед, чтобы она затвердела. Вырежьте кубический блок агарозы встроенным стволом мозга с помощью острого лезвия бритвы.
- Приклейте блок агарозы LMP на его ростральную сторону, сохраняя рострально-каудальную ось ствола мозга вертикальной.
- Принимайте корональные срезы до тех пор, пока область NM не будет визуализирована.
- Удалите блок агарозы из клея острым лезвием. Чтобы обнаружить ядра, аккуратно поместите 0,5 мкл красителя (толуидиновый синий или оранжевый G) на НМ тонкой иглой.
- Повторно установите этот блок на лоток для нарезки для сагиттальных или горизонтальных срезов и определите ядра относительно окрашенной области.
- Для достижения наилучшей производительности установите скорость нарезки вибратома на 4 - 5 (~ 30 ± 4 мм / мин), частоту вибрации на 85-87 Гц и амплитуду нарезки на 4-6 (~ 1 ± 0,2 мм).
- После секционирования ствола мозга поместите последовательно собранные ломтики размером 200-300 мкм в коммерчески доступную камеру срезов для уравновешивания в течение 1 ч при комнатной температуре в ACSF, непрерывно пузырящуюся смесью 95% O2/5% CO2 (рН 7,2-7,4, осмолярность 300-310 мОсм/л). В этих условиях ломтики сохраняют жизнеспособность до 5-6 ч.
7. Электрофизиология: процедура зажима пластыря
- Перенесите срез ствола мозга в камеру записи с непрерывной перфузией карбоксигенированного ACSF ~ 1,5 ± 0,5 мл / мин.
- Вытягивайте патч-пипетки с помощью съемника микропипетки диаметром наконечника 1-2 мкм и сопротивлением в диапазоне 3-6 МОм.
- Наполните пипетки внутренним раствором на основе K-глюконата (для регистрации тока).
- Чтобы проверить свойства нейронов в различных тонотопных областях внутри среза, найдите нейроны на обоих концах плоскости среза и подойдите к записывающему электроду.
- Поддерживайте положительное давление воздуха на кончике пипетки при приближении к нейрону.
- Двигайтесь в сторону сомы до тех пор, пока на нейроне не будет визуализирован отступ. Выполните следующие два шага быстро.
- Сделайте гигаомное (1 ГОм) уплотнение, выпустив положительное давление воздуха.
- Держите настройку усилителя в режиме зажима напряжения и корректируйте смещение пипетки как ноль пА. Проведите испытание уплотнения (испытательный импульс 10 мВ при 100 Гц). Примените отрицательное давление воздуха, чтобы разорвать небольшой участок мембраны нейрона.
- Чтобы проверить активные внутренние свойства слуховых нейронов, применяют гиперполяризующие и деполяризующие инъекции соматического тока.
ПРИМЕЧАНИЕ: Примеры этой процедуры можно визуализировать в дополнительном видео S1, S2. Подробности этой процедуры приведены в легендах видео.
Результаты
Все срезы ствола мозга, показанные здесь, были получены из ткани ствола мозга (~ 200-300 мкм) и визуализированы с использованием 5-кратной объективной и дифференциальной интерференционной контрастной (DIC) оптики. Камера была установлена на рассекающем микроскопе и подключена к компьютеру с программным обеспечением для получения изображений (см. Таблицу материалов). Спутниковые вставки для этих фигур (правые панели) были сфотографированы с использованием 60-кратного увеличения водяного погружного объектива. Были приняты меры для обеспечения того, чтобы все области среза ствола мозга были одинаково увеличены при получении цифровых изображений. Фотографии были сделаны с оптимальной яркостью и фокусировкой. Цифровые изображения срезов ствола мозга были сшиты планарным способом на основе перекрывающейся области и импортированы на настольный компьютер для дальнейшей настройки яркости, контрастности и оттенков серого. Основные микросхемы слухового ствола мозга курицы были идентифицированы в соответствии с предыдущей работой 1,2,5,13. Под микроскопом (объектив 5x) слуховые ядра были идентифицированы как область, прилегающая к сильно миелинизированным нервным волокнам, курсирующим вокруг каждого ядра как ипсилатерально, так и контралатерально вдоль дорсальных областей среза.
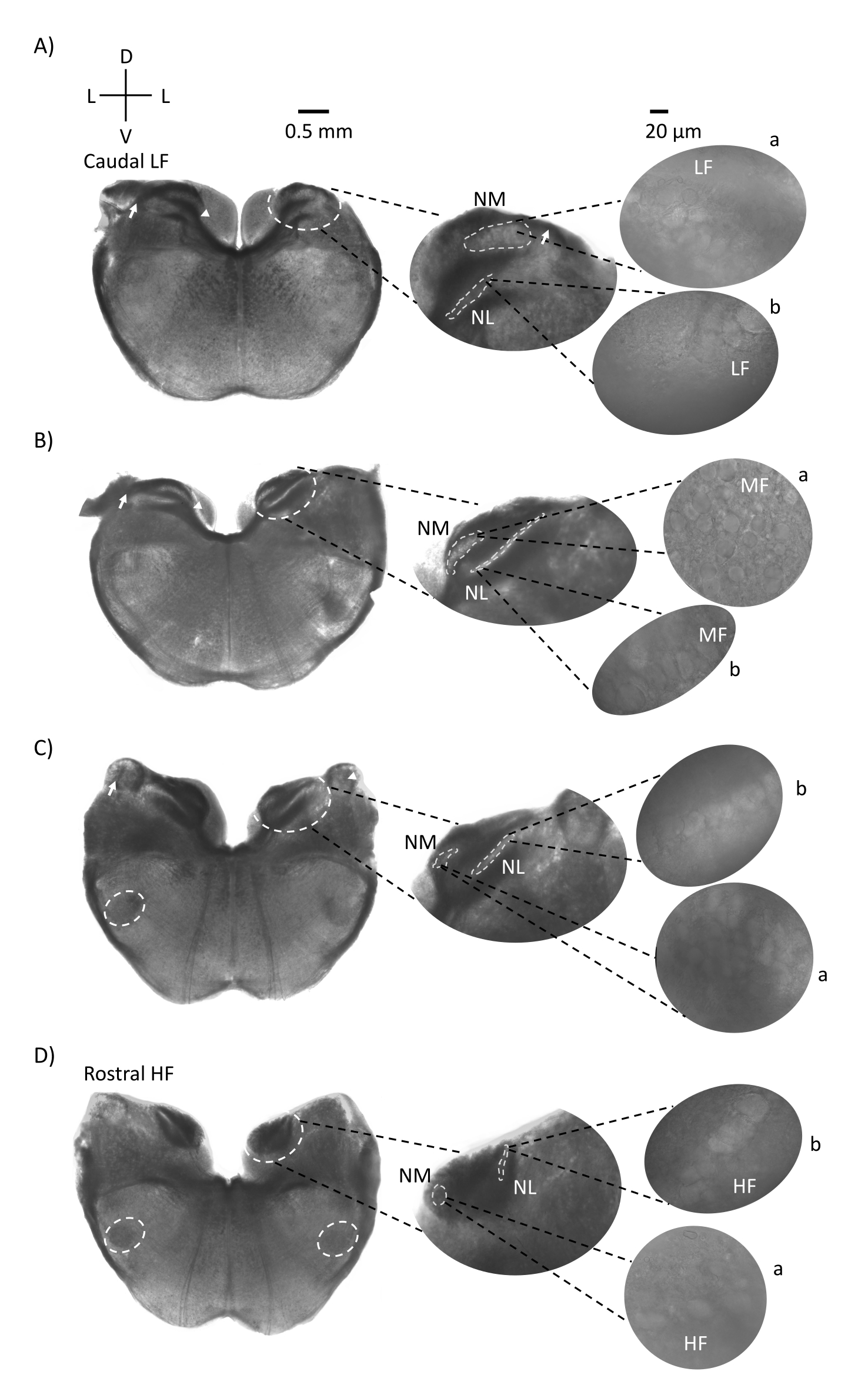
На рисунке 1 показаны традиционные корональные участки ткани ствола мозга (200-300 мкм) куриного эмбриона E21. Четыре корональных среза, показанные здесь, представляют относительные изочастотные области слуховых ядер ствола мозга от самой низкой слуховой области CF (рисунок 1A, каудо-латеральная), прогрессирующей до самой высокой слуховой области CF (рисунок 1D, рострально-медиальный). Для всех четырех корональных срезов на рисунке 1A-D увеличенные области меченых NM и NL показаны в средней колонке и увеличены (объектив 60x) на правом видении панели фигур (a и b, соответственно, в спутниковых вставках). Стрелка на рисунке 1A,B показывает вход слуховых нервных волокон, а наконечник стрелки показывает бифуркацию аксонов NM слева от среза. На рисунке 1C показана другая структура кохлеарного ядра птицы, известная как nucleus angularis (NA, стрелка слева и наконечник стрелки справа). Два самых ростральных корональных среза показывают верхнее оливарное ядро (SON), расположенное вдоль вентрально-латеральной области коронального среза (рисунок 1C, D, белые пунктирные круги).
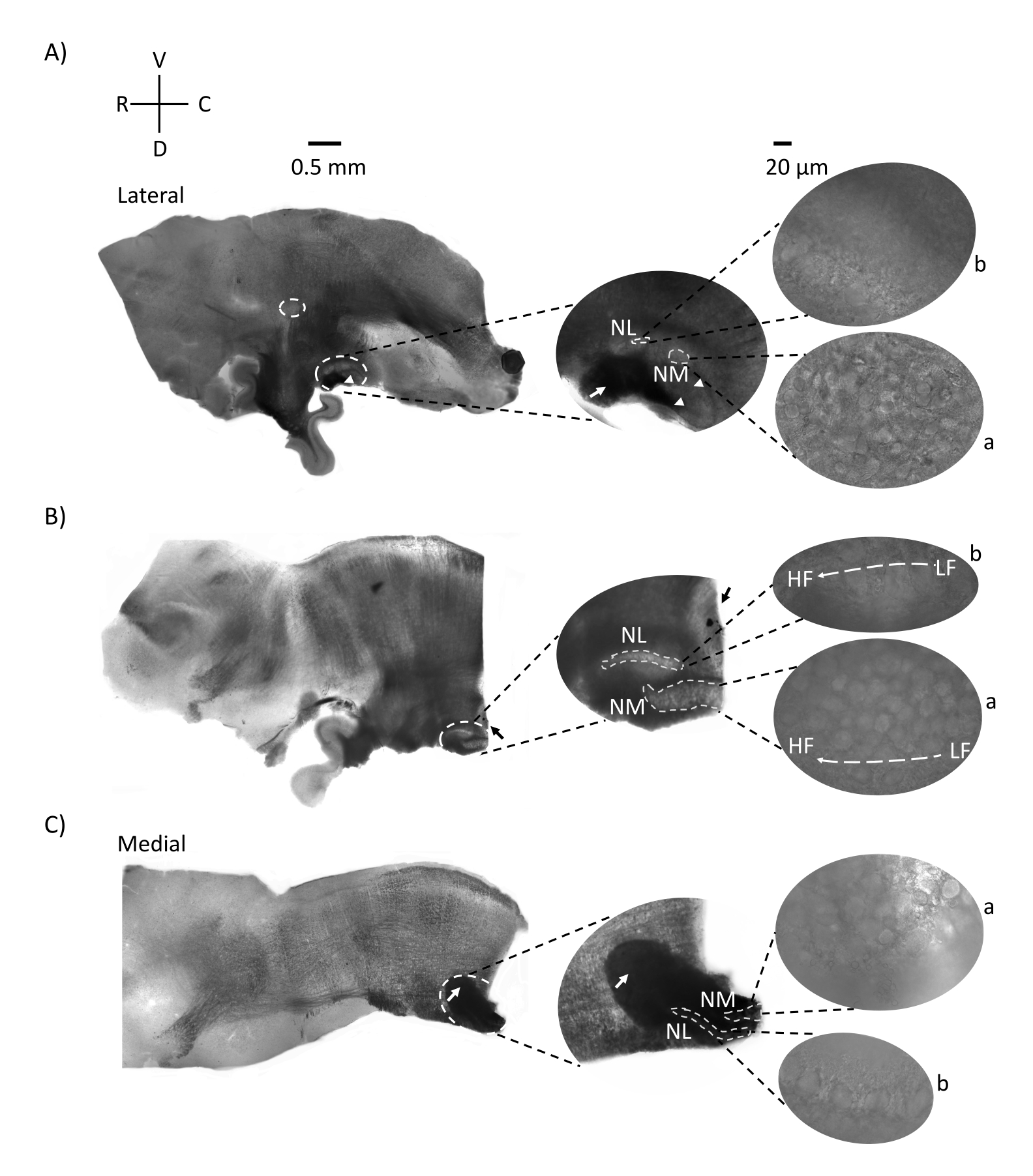
На рисунке 2 показаны сагиттальные участки ткани ствола мозга (200-300 мкм) из куриного эмбриона E21. Для всех трех сагиттальных срезов (рисунок 2A-C) увеличенные области меченых NM и NL показаны в средней колонке и увеличены (объектив в 60 раз) на правом видении фигурной панели (a и b, соответственно на спутниковых снимках). NM и NL были идентифицированы там, где слуховые нервные волокна (рисунок 2A, средняя стрелка) вошли в кластер нейронов, наблюдаемых при более высоком увеличении (рисунок 2A, средний, маленький, белые пунктирные круги и наконечники стрел) и выделяют начальную точку слуховой области (рисунок 2A, левый, большой, белый пунктирный круг и наконечник стрелки). SON был идентифицирован в ростро-латеральной области самого бокового среза (рисунок 2А, маленький, белый, пунктирный круг). На рисунке 2B показаны расширенные тонотопные области, которые содержат как относительно низкие, так и высокие слуховые области CF от NM и NL вдоль рострально-каудальной оси (белые очерченные области, см. также вставку спутника). На рисунке 2C показаны ипсилатеральные и контралатеральные аксональные пучки в самом медиальном срезе и конечной точке слуховой области (стрелки влево и в середине). Ориентация срезов, показанных здесь, контрастирует с традиционной ориентацией срезов, как показано на рисунке 1 (т.е. коронал). Это было выполнено для отображения ориентации, которая наилучшим образом соответствует подходу стеклянной пипетки, необходимой для электрофизиологических записей.
Чтобы подтвердить, что большая область тонотопической оси была представлена на рисунке 2B, электрофизиологические записи с токовым зажимом были выполнены из нейронов NM. На рисунке 3 показаны функциональные сходства и различия зрелых (E21) NM нейронов, записанных из коронального среза (рисунок 3A, B) и сагиттального среза (рисунок 3C, D, дополнительное видео S1, S2). Два нейрона NM были выбраны из медиального и латерального концов коронального среза (аналогично срезу, показанному на рисунке 1B), а два нейрона NM были выбраны из рострального и каудального концов NM в сагиттальном срезе (как на срезе, показанном на рисунке 2B). На рисунке 3А,В показаны свойства электрофизиологического ответа, аналогичные инъекциям соматического тока (от −100 пА до +200 пА, приращения +10 пА, длительность 100 мс). Паттерн возбуждения этих двух нейронов NM демонстрирует тонкие различия в этой плоскости среза, что указывает на относительную изочастотную пластинку для нейронов NM средней частоты. Рисунок 3C,D показывает, что паттерны срабатывания имеют существенные различия по рострально-каудальной оси, что указывает на относительно более высокий тонотопический градиент от низкочастотного nm-нейрона (рисунок 3C) к высокочастотному nm-нейрону (рисунок 3D). Оба нейрона представили свои стереотипные паттерны срабатывания, каксообщалось ранее 14,15.
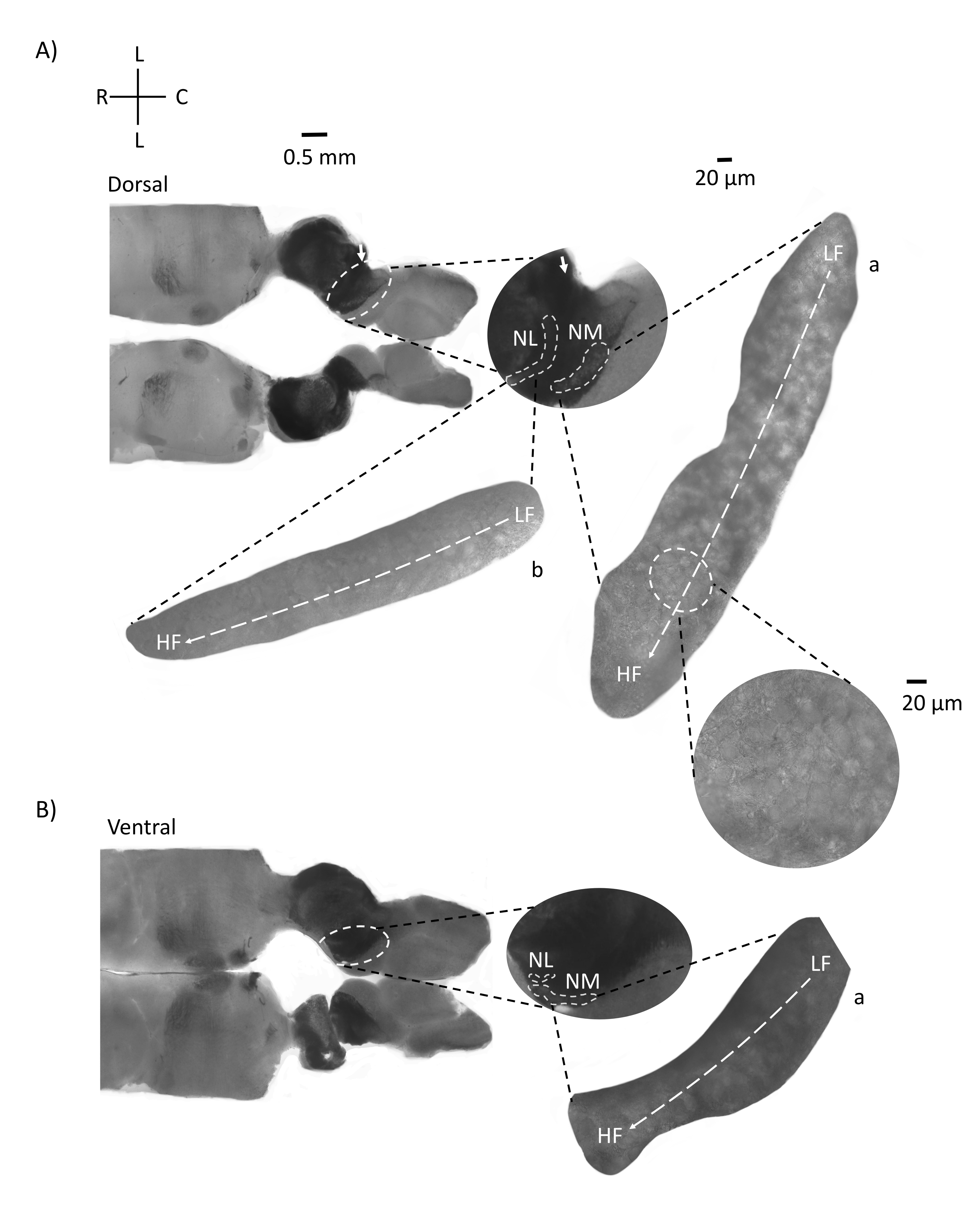
На рисунке 4 показаны горизонтальные участки ткани ствола мозга (200-300 мкм) куриного эмбриона E21. Для обоих горизонтальных срезов (рисунок 4A,B) увеличенные области меченых NM и NL показаны в средней колонке и увеличены (60-кратный объектив) на правом прицеле фигурной панели (a и b, соответственно, в спутниковых вставках). В горизонтальных срезах NM и NL были идентифицированы к средней линии, а нейроны были распределены вдоль латерально-медиальной оси (рисунок 4A, B, средние, белые, пунктирные области контура). Увеличенные изображения показывают большую протяженность тонотопического градиента. Низкочастотные нейроны находятся в каудо-латеральных областях, а высокочастотные нейроны — в рострально-медиальных областях (рисунок 4A,B, справа, спутники). Волокна, проходящие через среднюю линию вдоль рострально-каудальной оси, показывают контралатеральные связи слуховых ядер, но организация этих волокон не находится в простой плоскости. Однако острые угловатые срезы из горизонтального/поперечного сечения могут следовать за этими аксональными волокнами к сагиттальной плоскости. Срезы ткани ствола мозга толщиной 200-300 мкМ под острым углом (45°) от горизонтальной плоскости показаны на рисунке 5. Слуховые ядра ствола мозга можно увидеть через большой диагональный разворот, начиная от самого бокового среза и заканчивая самым медиальным срезом (рисунок 5A-C, обозначенные средними панелями, белая очерченная область). Кроме того, угловая ориентация областей NM и NL также может быть визуализирована в последовательных асимметричных срезах (рисунок 5A-C, обозначенные средние панели, белая, пунктирная очерченная область). Увеличенные изображения (объектив 60x) показывают тонотопическую ось слуховых ядер, когда она движется вдоль рострально-медиальной к каудо-боковой оси (рисунок 5A-C, справа, вставка спутника). Ориентация срезов на рисунке 5 аналогична ориентации на рисунке 2. Они контрастируют с традиционным представлением изображений, но больше подходят для электрофизиологических экспериментов.

Рисунок 1: Репрезентативные корональные последовательные участки ствола мозга. (A-D) Слева: срезы от каудальной до ростральной оси, слуховые ядра и соединительные волокна, отмеченные белым пунктирным кругом. Средняя вставка представляет собой больший вид слуховой области, где ядра показаны в белых пунктирных кругах a: NM и b: NL. Стрелки показывают слуховые нервные афферентные волокна, а наконечник стрелы показывает бифуркацию АКСОНА NM в A,B. Стрелка показывает NA в C. Боковой белый пунктирный круг показывает SON в C,D. Справа: спутниковая вставка показывает эти ядра в 60x объективе: a: NM и b: NL. Сокращения: NM = магноцеллюлярное ядро; NL = ядро laminaris; NA = угловатое ядро; SON = верхнее оливковое ядро; LF = относительно низкочастотные нейроны; MF = нейроны средней частоты; HF = высокочастотные нейроны; D = спинной; L = латеральный; V = вентральный. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2: Репрезентативные сагиттальные последовательные участки ствола мозга. (A-C) Слева: срезы от боковой до медиальной оси со слуховыми ядрами, помеченными белым пунктирным кругом. Средняя вставка показывает ту же область слуховых ядер в большем виде, отмеченную в белых пунктирных кругах. (A) Белый пунктирный круг в центре среза выделяет SON; стрелка, показывающая слуховые нервные волокна, и наконечник стрелы, показывающий NA. Темное черное пятно на правом конце среза является артефактом визуализации. Области мозжечка можно увидеть дорсально к слуховой области в обоих срезах A и B в левой панели. (B) Сагиттальный срез, ориентация которого была изменена на корональную плоскость (во время нарезки). Слуховая область была идентифицирована синим красителем (черная стрелка) и снова разрезана в сагиттальной плоскости. (А-С) Средняя вставка области NM и NL, обозначенные пунктирными белыми линиями. Справа: вид со спутника показывает a: NM и b: NL, наблюдаемые в 60-кратном увеличении объектива. Тонотопический градиент LF и HF в слуховых ядрах показан вдоль ростро-каудальной оси. Стрелки, указывающие на темную область в (C), показывают сильно миелинизированные NM-волокна, проходящие через среднюю линию через медиальную ось. Волокна соединяют обе стороны слуховых ядер. Сокращения: NM = магноцеллюлярное ядро; NL = ядро laminaris; NA = угловатое ядро; SON = верхнее оливковое ядро; LF = относительно низкочастотные нейроны; HF = высокочастотные нейроны; D = спинной; V = вентральный; R = рострал; C = каудальный. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3: Электрофизиологические записи реакции нейронов на инъекции соматического тока (от −100 пА до +200 пА, приращение +10 пА, длительность 100 мс) в режиме зажима тока. Нейроны были отобраны для записи в одном срезе, но в крайне противоположных областях НМ. (А,Б) Репрезентативные нейронные реакции в одном корональном срезе, указывающие на относительные изочастотные свойства с тонкими различиями. Свойства ответа представляют собой два различных нейрона MF, записанных из наиболее медиальной (A) и латеральной (B) областей NM в корональном срезе. (С,Г) Репрезентативные нейронные записи из одного сагиттального среза. Записи показывают относительно НЧ НМ отклик (C) и ответ HF NM (D), подчеркивая существенные различия в тонотопическом градиенте вдоль одного сагиттального участка. Сокращения: NM = магноцеллюлярное ядро; LF = относительно низкочастотные нейроны; MF = нейроны средней частоты; HF = высокочастотные нейроны. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 4: Репрезентативные горизонтальные последовательные участки ствола мозга. (А,Б) Слева: срезы вдоль дорсальной до вентральной оси, слуховые ядра отмечены белыми пунктирными кругами. Афферентные волокна8-го черепного нерва соединяют слуховые ядра, отмеченные стрелкой. Средняя вставка представляет собой более крупный вид области слуховых ядер со слуховыми ядрами, отмеченными под белыми пунктирными линиями NM и NL областей. Четкое топологическое движение слуховых ядер можно увидеть в A,B. (А,Б) Справа: большой вид со спутника, показывающий a: NM и b: NL. Правая вставка показывает слуховые ядра, наблюдаемые в 60-кратном объективном увеличении и изогнутой топологической оси от LF до HF вдоль каудо-латеральной до рострально-медиальной оси. Сокращения: NM = магноцеллюлярное ядро; NL = ядро laminaris; LF = относительно низкочастотные нейроны; HF = высокочастотные нейроны; L = латеральный; R = рострал; C = каудальный. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 5: Репрезентативные горизонтальные/поперечные острые угловые (45°) последовательные участки. (A-C) Слева: последовательные участки ствола мозга, слуховые ядра, отмеченные белым пунктирным кругом. Средняя вставка представляет собой больший вид слуховой области. (A) Средняя вставка показывает наибольшее распространение нейронов NM и NL в этих срезах. (В,С) Средняя вставка: слуховые ядра, отмеченные белыми пунктирными линиями, показывают постепенное топологическое изменение по сравнению с (A-C). Справа: спутниковая вставка, показывающая слуховые ядра a: NM и b: NL в 60-кратном объективном увеличении. Тонотопная ось от НЧ до ВЧ областей в NM и NL поворачивается угловато от боковых к медиальным срезам. Сокращения: NM = магноцеллюлярное ядро; NL = ядро laminaris; LF = относительно низкочастотные нейроны; HF = высокочастотные нейроны; V = вентральный; R = рострал; D = спинной; C = каудальный. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Дополнительное видео S1: Гиперполяризующие и деполяризующие инъекции соматического тока. Свойства отклика от низкочастотного и высокочастотного нейрона до 100 мс соматического тока в режиме тока. Нейроны были выбраны из того же сагиттального среза ствола мозга. Инъекции варьируются от -100 до +200 пА с шагом +10 пА, продолжительностью 100 мс. Потенциалы действия рассматриваются в ответ на достаточные деполяризующие текущие шаги. Видео соответствует окончательным следам, показанным на рисунке 3C. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительное видео S2: Гиперполяризующие и деполяризующие инъекции соматического тока. Подобно Дополнительному Видео S1, это видео показывает свойства отклика от низкочастотного и высокочастотного нейрона до 100 мс соматических впрысков тока в режиме текущего зажима. Нейроны были выбраны из того же сагиттального среза ствола мозга. Инъекции варьируются от -100 до +200 пА с шагом +10 пА, продолжительностью 100 мс. Потенциалы действия рассматриваются в ответ на достаточные деполяризующие текущие шаги. Видео соответствует окончательным следам, показанным на рисунке 3D. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Обсуждение
Корональные срезы эмбриональной ткани ствола мозга курицы позволили изучать относительную индивидуальную изочастотную пластинку на протяжениидесятилетий 1,2,5. Однако тонотопическая (т.е. частотная) организация слухового ствола мозга курицы топологически запутана и может быть более доступной в других анатомических осях в зависимости от конкретного вопроса исследования. Хотя этого достаточно для исследования анатомических и физиологических вопросов, относящихся к отдельным изочастотным областям, изучение тонотопических вариаций и их развитие в более крупных слуховых областях ствола мозга несколько ограничено корональными срезами. Чтобы преодолеть это ограничение, этот протокол описывает подходы в сагиттальной, горизонтальной и горизонтальной / поперечной плоскостях, чтобы предоставить дополнительные примеры слуховой ткани ствола мозга, которые проявляют максимальные тонотопические свойства и градиенты в отдельном участке ствола мозга.
Сагиттальные участки слуховых областей ствола мозга показывают, что различные тонотопные области распределены по большей области внутри среза по сравнению с корональными отделами (сагиттальная слуховая область = ~ 300-600 мкм, корональная слуховая область = ~ 200-350 мкм). Например, области NM и NL были визуализированы на большей площади вдоль ростро-каудальной оси в сагиттальных срезах (например, рисунок 2B), а функциональный тонотопический градиент, который проходит вдоль этой анатомической оси, в значительной степени содержался в одном сагиттальном срезе. Это было дополнительно подтверждено записями внутренних нейронных различий, которые изменяются вдоль рострально-каудального градиента, как сообщалось ранее14,15 (например, рисунок 3C, D). Будущие эксперименты, которые подчеркивают анатомические и иммуногистохимические свойства вдоль тонотопической оси, могут дополнительно исследовать известные градиенты слуховых свойств в пределах одной плоскости сагиттального среза. Они включают, но не ограничиваются ими, паттерны окрашивания MAP2 и экспрессии калиевых каналов, которые являются известными градиентами дендритной архитектуры и внутренними свойствами NM и NL, которые были ранее показаны в последовательных корональных разделах16.
Горизонтальные участки слуховых областей ствола мозга показывают, что NM и NL расположены к средней линии. Часть слуховых аксональных волокон проходит по диагонали или перпендикулярно горизонтальной плоскости (рисунок 4). За этими волокнами можно следовать, делая острый угловой срез на 45° к сагиттальной плоскости. Полученные горизонтальные/поперечные срезы были больше, чем сагиттальные или горизонтальные срезы, а длинные аксональные волокна проходили через ростро-каудальную ось как для ипсилатеральной, так и для контралатеральной сторон. Как NM, так и NL могут быть визуализированы в большей диагональной области (~ 400-700 мкм), так что контралатеральные связи могут быть визуализированы вдоль латерально-медиальной оси. Кроме того, горизонтальная/поперечная плоскость среза также показывает, как слуховые области и результирующий тонотопический градиент совершают угловой поворот (рисунок 5). Угловое воздействие контралатеральных соединений на большей площади делает эти срезы более подходящими для электрофизиологической стимуляции и исследований микросхем, чем традиционные корональные срезы.
Дополнительные преимущества
Формирование слуховых микросхем требует пространственно-временной координации сигналов, которые способствуют выживанию нейронов, синаптогенезу, аксональной дифференцировке, дендритной архитектуре и созреванию. Таким образом, альтернативные участки ствола мозга слуховой микросхемы куриного эмбриона могут быть использованы по следующим темам исследований: морфологическая организация нейронов в топографически различных измерениях; организация и картирование коннектомов всех слуховых и вестибулярных ядер; идентификация и характеристика паттернов активности составляющих контура в изочастотной и тонотопной плоскостях; топографическая организация возбуждающих и тормозных микросхем и связи со специализированными популяциями нейронов (ядрами); пространственное расположение нейронов слуховых ядер и его прогностический CF17; систематическое нацеливание на специфические тонотопные типы нейронов; отслеживание клеток-предшественников и их развитие в законсервированные ядра; генетическая линия клеток к эволюции нейронных цепей18; сравнительная анатомия ствола мозга между видами; исследование вестибулярных цепей, таких как вестибулярный комплекс Дейтера (DC)19; и синхронность и перекрестный разговор между вестибулярными ядрами.
Многогранный подход с использованием различных плоскостей срезов может помочь ответить на фундаментальные вопросы о неизвестных анатомических и биофизических свойствах микросхем ствола мозга. Хорошим примером является связь между основными слуховыми ядрами (NM, NA, NL и SON) и вестибулярными ядрами, включая дорсальное ядро латерального лемниска (LLDp), полулунное ядро (SLu)20 и тангенциальное ядро (TN)3. Тем не менее, этот протокол и эти исследования на основе срезов имеют некоторые ограничения.
Меры предосторожности и ограничения
В зависимости от учреждения, проводящего эксперименты, этические принципы и обращение с куриными эмбрионами могут отличаться. В то время как Национальные институты здравоохранения Руководящие принципы по уходу и использованию лабораторных животных допускают быстрое обезглавливание, существуют альтернативные методы эвтаназии куриного эмбриона21. Ранняя развивающаяся ткань ствола мозга куриного эмбриона мягкая и деликатная по сравнению с более старыми эмбрионами. Он имеет несколько соединений и кровеносных сосудов на поверхности, которые нуждаются в дополнительной осторожности при их удалении. Ткань следует хранить в ледяном dACSF и перфузить 95% O2/5% CO2 для повышения жизнеспособности.
Метод сагиттальной нарезки полезен только при ипсилатеральной тонотопии. Этот метод нарезки обеспечивает более крупные срезы, чем корональные срезы, обработка которых может быть ненадежной. Тем не менее, можно обрезать срезы с помощью методов поперечных игл, подробно описанных в другом месте22. Использование 4% LMP агарозного блока встроенного ствола мозга может спасти тонкие структуры в срезах, но необходимо соблюдать осторожность, чтобы не налить чрезмерно горячую агарозу. Быстрая установка его путем помещения поврежденного агарозой ствола мозга в охлажденную среду в течение ~ 1 мин делает срезы более жизнеспособными для электрофизиологических записей.
Применение суперклея в избыточных количествах может быть токсичным. Он должен применяться минимально, а избыточные количества должны быть немедленно вымыты путем замены dACSF. Для острых угловых (45°) срезов критическим является разрезание угла блока агарозы; можно использовать зеркало, чтобы увидеть передний угол при разрезании блока агарозы острым лезвием. Коммерчески доступные лезвия могут иметь восковое покрытие, которое следует протереть спиртом и высушить перед использованием. Оптимизация требуется для скорости и частоты резки вибратома, поскольку пучки аксонального волокна тверже, чем кортикальная или матричная ткань. Поддержание высокой амплитуды и использование охлажденного раствора для рассечения может предотвратить повреждение тканей.
Все растворы должны быть приготовлены свежими, а Ca2+ и Mg2+ следует добавлять в ACSF после пузырьков 95% O2/5% CO2 . В противном случае могут выпасть осадки Ca2+. Кисть должна использоваться для осторожной обработки срезов внутри вибратома. Сохраняйте общее время нарезки менее 15 минут, если это возможно. Стеклянная пипетка Пастера может быть использована для маневрирования срезами ствола мозга.
Не используйте моющие или коррозионные моющие средства для стеклянной посуды и оборудования, которые контактируют с срезами, используемыми в электрофизиологии. Полученные снимки представляют собой появление ткани толщиной 200-300 мкМ при дифференциальном интерференционном контрасте (ДВС-синдроме). Визуальное качество будет хуже, чем иммуногистохимия или электронная микроскопия, но оно точно отражает то, что увидит экспериментатор при выполнении электрофизиологических записей.
Исследования, относящиеся к раннему развитию микросхем вдоль альтернативной анатомической оси, будь то дорсально-вентральные, рострально-каудальные или ипсилатерально-контралатеральные, ограничены в слуховом стволе мозга курицы. Одна из причин этого заключается в том, что роль транскрипционных кодов и регуляции тонотопического развития в стволе мозга до сих пор не полностью понята. Функциональные явления, такие как нисходящая модуляция и спонтанная активность, часто теряются при наблюдении активности in vitro. Тем не менее, исследование in vivo дополняется специфическими и прямыми записями одиночных нейронов, возможными только в этих условиях среза. Уточнение получения ткани ствола мозга вдоль различных ориентаций может дать проницательную информацию о развитии и сложности тонотопических градиентов в слуховой микросхеме ствола мозга курицы.
Раскрытие информации
Все авторы заявляют, что исследование было проведено без какого-либо коммерческого или финансового интереса и что у них нет никаких конфликтов интересов.
Благодарности
Эта работа поддерживается грантом NIH/NIDCD R01 DC017167. Мы благодарим Кристин Маклеллан за редакционные комментарии к более ранней версии рукописи.
Материалы
| Name | Company | Catalog Number | Comments |
| Adobe photoshop 2021 | Adobe | ||
| Anti-vibration table 30"x 36" - OTMC - 63533 | TMC | ||
| Cell sens standard software | OLYMPUS | ||
| Digidata 1440A | MOLECULAR DEVICES | ||
| Digital amplifier multiclamp 700B | MOLECULAR DEVICES | ||
| DSK line-up linearslicer pro7 | TED PELLA, INC | ||
| Micromanipulator MPC-385 / OSI-MPC-385-2 | OLYMPUS AMERICA INC | ||
| Micropipette puller P-97 | SUTTER INSTRUMENTS | ||
| Microscope BX51W1 | OLYMPUS AMERICA INC | ||
| MS ICE software | Microsoft Corporation | ||
| Ohaus balance model AV212 | Ohaus Adventurer | ||
| Olympus DPSI0 /DPS80 camera | OLYMPUS | ||
| pClamp and Axoclamp data Acquisition Softwares | MOLECULAR DEVICES | ||
| pH meter lab 850 benchtop | SCHOTT INSTRUMENTS | ||
| Sharp stainless blade | Dorco/Personna | ||
| Vapor pressure osmometer model 5600 | WESCOR INC | ||
| Water purification systems Smart2pure 6UV/UF | Thermo Scientific | ||
| Chemicals- list | |||
| Agrose Low melt IB70051 | IBI SCIENTIFIC | ||
| CaCl2 (Calcium Chloride) | ACROS organics | ||
| Cynergy instant adhesive CA6001 | Resinlab | ||
| Dextrose (D-(+)-glucose) | VWR Life Science | ||
| Ethyl alcohol | IBI SCIENTIFIC | ||
| KCl (Potassium Chloride) | Amresco.Inc | ||
| MgCl2 (Magnesium Chloride) | Sigma-Aldrich | ||
| NaCl (Sodium Chloride) | Amresco.Inc | ||
| NaH2PO4 (Sodium Dihydrogen Phosphate) | Amresco.Inc | ||
| NaHCO3 (Sodium Bicarbonate) | Amresco.Inc |
Ссылки
- Rubel, E. W., Parks, T. N. Organization and development of brain stem auditory nuclei of the chicken: tonotopic organization of n. magnocellularis and n. laminaris. Journal of Comparative Neurology. 164 (4), 411-433 (1975).
- Rubel, E. W., et al. Organization and development of brain stem auditory nuclei of the chicken: ontogeny of n. magnocellularis and n. laminaris. Journal of Comparative Neurology. 166 (4), 469-489 (1976).
- Shao, M., et al. Spontaneous synaptic activity in chick vestibular nucleus neurons during the perinatal period. Neuroscience. 127 (1), 81-90 (2004).
- Fukui, I., Ohmori, H. Tonotopic gradients of membrane and synaptic properties for neurons of the chicken nucleus magnocellularis. Journal of Neuroscience. 24 (34), 7514-7523 (2004).
- Sanchez, J. T., Seidl, A. H., Rubel, E. W., Barria, A. Preparation and culture of chicken auditory brainstem slices. Journal of Visualized Experiments. (49), e2527 (2011).
- Sanchez, J. T., Lu, Y., Fay, R. R., Popper, A. N., Cramer, K., Coffin, A. Glutamate signaling in the auditory brainstem. Auditory Development and Plasticity: Springer Handbook of Auditory Research. 64 (4), 75-108 (2017).
- Parks, T. N. Morphology of axosomatic endings in an avian cochlear nucleus: nucleus magnocellularis of the chicken. Journal of Comparative Neurology. 203 (3), 425-440 (1981).
- Jhaveri, S., Morest, D. K. Sequential alterations of neuronal architecture in nucleus magnocellularis of the developing chicken: a Golgi study. Neuroscience. 7 (4), 837-853 (1982).
- Carr, C. E., Boudreau, R. E. Central projections of auditory nerve fibers in the barn owl. Journal of Comparative Neurology. 314 (2), 306-318 (1991).
- Köppl, C. Auditory nerve terminals in the cochlear nucleus magnocellularis: differences between low and high frequencies. Journal of Comparative Neurology. 339 (3), 438-446 (1994).
- Fukui, I., et al. Improvement of phase information at low sound frequency in nucleus magnocellularis of the chicken. Journal of Neurophysiology. 96 (2), 633-641 (2006).
- Wang, X., et al. Postsynaptic FMRP regulates synaptogenesis in vivo in the developing cochlear nucleus. Journal of Neuroscience. 38 (29), 6445-6460 (2018).
- Lu, T., Cohen, A. L., Sanchez, J. T. In ovo electroporation in the chicken auditory brainstem. Journal of Visualized Experiments. (124), e55628 (2017).
- Hong, H., Sanchez, J. T. Need for speed and precision: structural and functional specialization in the cochlear nucleus of the avian auditory system. Journal of Experimental Neuroscience. (12), 1-16 (2018).
- Hong, H., et al. Diverse intrinsic properties shape functional phenotype of low-frequency neurons in the auditory brainstem. Frontiers in Cellular Neuroscience. 12, 1-24 (2018).
- Wang, X., Hong, H., Brown, D. H., Sanchez, J. T., Wang, Y. Distinct neural properties in the low-frequency region of the chicken cochlear nucleus magnocellularis. eNeuro. 4 (2), 1-26 (2017).
- Tabor, K. M., et al. Tonotopic organization of the superior olivary nucleus in the chicken auditory brainstem. Journal of Comparative Neurology. 520 (7), 1493-1508 (2012).
- Lipovsek, M., Wingate, R. J. Conserved and divergent development of brainstem vestibular and auditory nuclei. Elife. 7, 40232 (2018).
- Passetto, M. F., et al. Morphometric analysis of the AMPA-type neurons in the Deiter's vestibular complex of the chick brain. Journal of Chemical Neuroanatomy. 35 (4), 334-345 (2008).
- Curry, R. J., Lu, Y. Intrinsic properties of avian interaural level difference sound localizing neurons. Brain Research. 1752, 147258 (2021).
- Aleksandrowicz, E., Herr, I. Ethical euthanasia and short-term anesthesia of the chick embryo. ALTEX - Alternatives to Animal Experimentation. 32 (2), 143-147 (2015).
- Palkovits, M. Isolated removal of hypothalamic or other brain nuclei of the rat. Brain Research. 14 (59), 449-450 (1973).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены