Method Article
Análisis de la formación de complejos proteicos a concentraciones micromolares mediante el acoplamiento de microfluídica con fotometría de masas
En este artículo
Resumen
Este protocolo combina la fotometría de masas con un novedoso sistema de microfluídica para investigar las interacciones proteína-proteína de baja afinidad. Este enfoque se basa en la dilución rápida de complejos altamente concentrados en solución, lo que permite mediciones de baja afinidad y amplía la aplicabilidad de la fotometría masiva.
Resumen
La fotometría de masas es una tecnología versátil de medición de masas que permite el estudio de las interacciones biomoleculares y la formación de complejos en solución sin etiquetas. La fotometría de masas es generalmente adecuada para analizar muestras en el rango de concentración de 100 pM-100 nM. Sin embargo, en muchos sistemas biológicos, es necesario medir muestras más concentradas para estudiar interacciones transitorias o de baja afinidad. Aquí, demostramos un método que expande de manera efectiva el rango de concentraciones de muestras que se pueden analizar por fotometría de masas desde nanomolares hasta decenas de micromolares.
En este protocolo, la fotometría de masas se combina con un novedoso sistema de microfluídica para investigar la formación de complejos proteicos en solución en el rango de concentración micromolar. Con el sistema de microfluídica, los usuarios pueden mantener una muestra a una concentración deseada más alta, seguida de una dilución hasta el rango nanomolar, varios milisegundos antes de la medición de la fotometría de masas. Debido a la velocidad de la dilución, los datos se obtienen antes de que el equilibrio de la muestra haya cambiado (es decir, disociación del complejo).
La técnica se aplica para medir las interacciones entre un anticuerpo de inmunoglobulina G (IgG) y el receptor Fc neonatal, mostrando la formación de complejos de alto orden que no fueron cuantificables con mediciones de fotometría de masa estática.
En conclusión, la combinación de fotometría de masas y microfluídica permite caracterizar muestras en el rango de concentración micromolar y es competente en la medición de interacciones biomoleculares con afinidades más débiles. Estas capacidades se pueden aplicar en una variedad de contextos, incluido el desarrollo y el diseño de bioterapéuticos, lo que permite una caracterización exhaustiva de diversas interacciones proteína-proteína.
Introducción
Las interacciones proteína-proteína subrayan la mayoría de las funciones celulares, desde la regulación inmunitaria hasta la replicación y traducción del ADN. Como resultado, existe una necesidad fundamental en todas las ciencias de la vida para investigar una amplia gama de interacciones a través de diversos complejos heterogéneos que se forman comúnmente. Sin embargo, su detección, caracterización y cuantificación suelen ser un reto, especialmente en el caso de las interacciones de baja afinidad1.
Los ensayos de inmunoprecipitación se utilizan a menudo para detectar interacciones de alta afinidad, pero para interacciones transitorias y de baja afinidad, la detección es en gran medida inviable2. También se pueden utilizar técnicas de fluorescencia, pero requieren la adición potencialmente disruptiva de etiquetasfluorescentes 2. Cryo-EM puede proporcionar una instantánea estructural y una lectura de conjunto de los complejos de proteínas formados con alta resolución espacial, pero también suele requerir trabajar a concentraciones que son demasiado bajas para obtener imágenes de interacciones de baja afinidad. Cryo-EM también presenta desafíos relacionados con el costo, la accesibilidad, la preparación de muestras y el tiempo de análisis3.
Además, la resonancia de plasmón de superficie (SPR) se ha convertido en una forma popular de cuantificar las interacciones proteína-proteína, aunque requiere la inmovilización de proteínas, lo que puede afectar el equilibrio de unión y dar lugar a tasas de activación variables, reduciendo así la precisión de la medición 4,5. También implica varios pasos de ensayo antes de la recopilación y el análisis de datos6.
La fotometría masiva es una técnica de una sola molécula que se ha utilizado para analizar las interacciones proteína-proteína 5,6,7. Funciona midiendo la masa de moléculas individuales o complejos en función de la luz que dispersan cuando aterrizan en la superficie de un cubreobjetos de vidrio8. Las mediciones de fotometría masiva se han utilizado para cuantificar las afinidades de unión a partir de la abundancia relativa de los socios de unión y los complejos que forman5. Sin embargo, al igual que otras técnicas de una sola molécula, la concentración de la muestra que se va a medir suele ser inferior a 100 nM. Si la concentración es mayor, las moléculas que aterrizan en la superficie del vidrio se superpondrán espacialmente, lo que dará como resultado una mala calidad de los datos7. En consecuencia, las interacciones más débiles (KD ~ micromolares), que se disocian a estas concentraciones más bajas, no pueden medirse de manera confiable ya que no es posible observar la mezcla necesaria de especies unidas y no unidas5.
Aquí, describimos un enfoque que supera esta limitación basado en un nuevo dispositivo de fotometría masiva de microfluídica acoplada. Específicamente, un sistema de microfluídica se utiliza en combinación con el fotómetro de masas para expandir de manera efectiva la gama de interacciones que se pueden cuantificar mediante fotometría de masas. Se ha demostrado que la microfluídica ofrece una gama de posibilidades para investigar las interacciones proteína-proteína, incluida la dilución rápida para detectar interacciones débiles 1,9. El sistema descrito en este documento funciona diluyendo rápidamente la muestra hasta 10.000 veces en un chip microfluídico y fluyendo inmediatamente a través del área de observación del chip, lo que permite que la medición de la fotometría de masas comience dentro de los 50 ms desde que las moléculas comenzaron el proceso de dilución10. La dilución se produce cuando la muestra y el tampón se combinan en un mezclador de válvula Tesla inversa en el chip, y los caudales relativos de las dos soluciones determinan la cantidad de dilución que se produce (consulte el paso 8 del protocolo). El caudal es controlable con el software de control microfluídico. La alteración del caudal puede cambiar la población relativa de la especie, ya que puede afectar el número de eventos de aterrizaje en la superficie del vidrio, que es lo que mide el fotómetro de masa.
La velocidad del proceso es lo suficientemente rápida como para que la medición se complete antes de que se interrumpa la integridad de la interacción (para más detalles, consulte también la Discusión). Esto se puede entender a través de una breve mirada a la teoría de las reacciones de primer orden, donde  . La constante de velocidad hacia adelante (asociación) es kf, la constante de tasa hacia atrás (disociación) es kb y la constante de disociación de equilibrio (KD) se define como
. La constante de velocidad hacia adelante (asociación) es kf, la constante de tasa hacia atrás (disociación) es kb y la constante de disociación de equilibrio (KD) se define como
KD= kb/ kf
Para la unión a proteínas, kfestá generalmente limitada por la difusión de los reactivos11 y, por lo tanto, está restringida al rango de 10 6-107 M-1·s-1. Debido a que el rango de es limitado, una reacción de baja afinidad (KD ~ micromolares) tendrá kb≈ 1 s-1. Es decir, kb= kf · KD= (106 M-1·s-1) (10-6 M) = 1 s-1, con una vida media del complejo de alrededor de 0,7 s11,12.
Nuestro sistema de ejemplo es la unión del anticuerpo monoclonal IgG trastuzumab al dominio soluble del receptor Fc neonatal de IgG (FcRn), que se sabe que interactúan13. Los datos publicados anteriormente obtenidos utilizando solo fotometría masiva convencional (es decir, con dilución manual de muestras) mostraron que las proteínas forman múltiples especies. Los monómeros de FcRn, los dímeros de FcRn y la IgG no unida fueron claramente visibles, mientras que los complejos IgG-FcRn (en proporciones 1:1 y 1:2) también se detectaron (a pH 5,0) pero solo con una abundancia muy baja5. Esta observación plantea la cuestión de si la formación del complejo IgG-FcRn podría detectarse más claramente si se mide a una concentración más alta. De hecho, la combinación de la fotometría masiva con un enfoque de dilución rápida acoplada descrito aquí proporcionó pruebas más sólidas de la formación de complejos mediante un aumento de las partículas medidas.
El protocolo de fotometría de masas y microfluídica aquí descrito permite caracterizar la formación de complejos con un KD hasta el rango micromolar. Una determinación empírica del KD requerirá mejoras adicionales en la precisión del sensor de flujo, la estabilidad de la bomba, las variaciones de viruta a viruta y la ubicación de la medición dentro de la ventana de observación, ya que todos estos factores influirían en el tiempo desde que la muestra se diluye hasta que se mide.
El mismo enfoque podría aplicarse para investigar la unión entre cualquier proteína soluble, siempre que tenga pesos moleculares distintos (separados por al menos 25 kDa) que se encuentren en el rango adecuado para el análisis con un fotómetro de masa (30 kDa a 6 MDa). Los conocimientos obtenidos podrían ser útiles para estudios en una variedad de contextos, desde la obtención de una comprensión mecanicista de las funciones celulares hasta el diseño de nuevos medicamentos bioterapéuticos.
Protocolo
1. Preparación de los instrumentos y puesta en marcha del software
- Encienda el fotómetro de masas y déjelo encendido durante al menos 1 hora antes de comenzar cualquier medición.
NOTA: Esto se debe a que el instrumento necesita alcanzar una temperatura constante. No dejar el fotómetro de masa encendido durante 1 h podría provocar un cambio de masa y, por lo tanto, resultados inexactos. - Encienda la mesa antivibratoria encendiendo la alimentación y presionando el botón de aislamiento (debajo del fotómetro de masas). Las vibraciones limitan el rendimiento del instrumento de fotometría de masas, por lo que la mesa antivibratoria es importante para garantizar la máxima sensibilidad del instrumento. Encienda la caja de microfluídica.
- Inicie el software de adquisición de datos y el software de control de microfluídica. Encienda el compresor de aire.
2. Preparación de muestras de proteínas, tampón y soluciones de limpieza
- Agregue 200 mL de tampón PBS (pH 7.4) a una botella limpia de 200 mL y 200 mL de tampón PBS (pH 5.0) a una segunda botella de 200 mL.
NOTA: Las botellas de 200 ml deberán conectarse al sensor de la unidad de flujo "L" en los siguientes pasos. Utilice tampón PBS de pH 7,4 para la medición de calibración y tampón PBS de pH 5,0 para la medición de la muestra. - En un tubo de centrífuga de 0,5 mL, mezcle IgG (2 μM) y FcRn (20 μM) en una proporción de 1:10 para un volumen total de 60 μL.
- Las soluciones madre para FcRn e IgG son de 91,9 μM y 13,5 μM, respectivamente. Prepare la reacción mezclando 9 μL de IgG + 13 μL de FcRn + 38 μL de PBS (pH 5,0, temperatura ambiente [RT]) en un tubo de centrífuga para un volumen total de 60 μL. Las concentraciones finales de los dos reactivos fueron de 2 μM para IgG y 20 μM para FcRn. Mantenga todas las reservas de proteínas en hielo durante todo el proceso.
- En otro tubo de centrífuga de 0,5 mL, dispense 20 μL del calibrante de β-amilasa a una concentración de 20 μM.
- A modo de ejemplo, para preparar el calibrante de β-amilasa utilizado aquí, siga el procedimiento descrito en 2.3.1.1-2.3.1.2.
- Resuspender 20 mg de polvo de β-amilasa en el vial en 3,57 mL de PBS (5% de glicerol), correspondientes a 5,6 mg/mL y una concentración molar de 100 μM (ya que el peso molecular es de 56 kDa).
- Para diluir a 20 μM, combine 200 μL de la culata de 100 μM con 800 μL de PBS (pH 7,4, RT) en un tubo de centrífuga de 1 mL.
- A modo de ejemplo, para preparar el calibrante de β-amilasa utilizado aquí, siga el procedimiento descrito en 2.3.1.1-2.3.1.2.
- Incubar las muestras en RT durante al menos 30 min.
NOTA: Comience a preparar la muestra y las diluciones del calibrante después de encender el fotómetro de masas. Para este experimento, las muestras se incubaron durante 120 min. - En tres tubos de centrífuga de 50 mL, alícuota: 50 mL de PBS (pH 7,4) - solución de limpieza 1 (CS1), 50 mL de NaOH 0,5 M - solución de limpieza 2 (CS2) y 50 mL de IPA 100% - solución de limpieza 3 (CS3).
3. Montaje experimental
- Para cargar la muestra y el calibrador, coloque el tubo de centrífuga calibrante en la posición 4 y el tubo de centrífuga de muestra en la posición 5 de la caja de microfluídica (Figura 1).
- Para cargar las soluciones de limpieza, coloque CS1, CS2 y CS3 en las posiciones 1, 2 y 3 de la caja de microfluídica (Figura 1).
NOTA: La muestra, el calibrador y las soluciones de limpieza están conectadas al interruptor múltiple (m-switch). El m-switch está conectado al sensor de la unidad de flujo "S". - Enrosque el tapón que se conecta a la línea de tampón a la botella tampón de 200 ml (pH 7,4).
NOTA: La línea de amortiguación está conectada a una válvula de cierre, que está conectada al sensor de la unidad de flujo "L". La válvula de cierre evita cualquier efecto de sifón que pueda ocurrir cuando se detiene el flujo de amortiguación. El sifón puede provocar la contaminación del tampón por el fluido residual diluido que regresa al depósito tampón. - Coloque el chip de microfluídica en una placa de preparación.
Empuje el otro extremo del tubo del sensor de la unidad de flujo "S" en la "entrada de muestra" del primer canal en el chip microfluídico (Figura 1). La línea de muestra está completa. - Empuje el otro extremo del tubo del sensor de la unidad de flujo "L" en la "entrada de búfer" del primer canal en el chip microfluídico (Figura 1). La línea de búfer está completa.
- Para recoger el flujo de salida, empuje un tubo a la "Salida" del primer canal en el chip de microfluídica; coloque el otro extremo de modo que se drene en un matraz o vaso de precipitados receptible de residuos (Figura 1).
4. Cebado de la muestra y las líneas de tampón con tampón
- Abra la válvula de cierre manual en la línea de amortiguación.
- En el software de control de microfluídica, ajuste el caudal de la línea de tampón a 1000 μL/min y asegúrese de que la presión de la línea de tampón no supere los 110 mbar (Figura 2). El flujo se iniciará automáticamente en la línea de amortiguación (Figura 1).
NOTA: Si la presión de la línea de amortiguación supera los 110 mbar, podría deberse al paso del aire a través del sensor de flujo. Si la presión no vuelve a la normalidad en unos pocos segundos, es posible que haya una obstrucción (generalmente debido a un tubo apretado en las conexiones). En ese caso, detenga el flujo y verifique las conexiones en la línea de amortiguación, comenzando por la válvula de cierre. - En el software de control de microfluídica, seleccione la posición 1 del m-switch (correspondiente al tampón PBS), luego ajuste el caudal de la línea de muestreo a 8 μL/min y asegúrese de que la presión de la línea de muestreo no supere los 350 mbar. El flujo se iniciará automáticamente en la línea de muestreo (Figura 1).
- Utilice una punta de pipeta (u otro componente de plástico blando) para aplicar una presión suave desde arriba cerca de las burbujas atrapadas en el chip para desalojar las burbujas de aire y asegurarse de que se eliminen a través de la salida.
NOTA: Asegúrese de eliminar todas las burbujas en las secciones 'mezclador' y 'área de observación' (Figura 1) del canal en uso. Tenga cuidado de no empujar demasiado fuerte, ya que esto podría dañar el chip.
5. Colocar el chip de microfluídica en el fotómetro de masas y encontrar el foco
- Aplique una gota de aceite de inmersión para microscopio sobre el objetivo del fotómetro de masas.
- Coloque el chip microfluídico en el soporte del fotómetro de masas con la "entrada de muestra" hacia arriba y manténgalo unido a las abrazaderas de la etapa (Figura 1). Asegúrese de que todas las conexiones de los tubos permanezcan conectadas.
- Utilizando el software de adquisición de datos, mueva la plataforma para asegurarse de que el "Área de observación" del canal 1 esté alineado con el objetivo (Figura 1).
- Cierre la tapa del fotómetro de masa y presione la opción Droplet-Dilution Find Focus en el software de adquisición de datos.
- Verifique el anillo de enfoque blanco en la esquina inferior izquierda del software de adquisición de datos (Figura 3). Los espacios en el anillo indican la presencia de una burbuja de aire en el aceite de inmersión; Elimine esto aumentando la velocidad del escenario al máximo y moviendo suavemente el escenario lateralmente.
- Una vez completado el enfoque, espere 2-3 minutos antes de realizar la primera grabación. A continuación, pulse Grabar para registrar una medición de 1 minuto y asegurarse (observando la medición) de que no aparezcan impurezas.
NOTA: Las impurezas pueden estar en la superficie del vidrio o en el tampón. El valor de nitidez en el software de adquisición de datos debe estar por encima del 4,5%.
6. Calibración de fotometría masiva
- En el software de control de microfluídica, cambie el interruptor m a la posición 4 (correspondiente al calibrante) y asegúrese de que el caudal de la línea de muestreo esté ajustado a 8 μL/min y que la presión de la línea de muestreo no supere los 350 mbar (Figura 2).
- El calibrador comenzará el flujo a través del chip. Espere aproximadamente 1,5-2,5 minutos (o hasta que el calibrador se vea de manera consistente en el software de adquisición de datos). El tiempo puede variar según la longitud del tubo de la línea de muestreo.
- Una vez que el calibrante se ve consistentemente (es decir, el número de eventos es suficiente para una medición precisa de la fotometría de masas), en el software de control de microfluídica, reduzca el caudal a 0,5 μL/min, el nivel de dilución objetivo para este experimento.
NOTA: Comenzar con un caudal más alto simplemente reduce el tiempo necesario para que el calibrante llegue al chip. - Asegúrese de que no haya "muy pocas" o "demasiadas" moléculas aterrizando en la superficie de medición (Figura 4).
- Si la densidad del evento de aterrizaje no es "ideal" (Figura 4), cambie el caudal en el software de control de microfluídica. Si la densidad del evento es demasiado baja, aumente el caudal de la muestra hasta que se observen eventos de aterrizaje bien separados (pero no supere los 8 μL/min). Si es demasiado alto, reduzca el caudal de la muestra (pero no baje de 0,1 μL/min).
- Si el volumen del calibrador se está agotando en el tubo de muestra, cambie la posición del interruptor m a la línea PBS pH 7.4 (posición 1) para evitar inyectar aire.
- Presione Grabar y tome una medida de 60 seg. Guarde el archivo en la carpeta elegida.
NOTA: El software de adquisición de datos produce archivos con .mp como extensión.
7. Limpieza de la línea de muestreo y detención del flujo
- En el software de control de microfluídica, cambie a la posición 2 del interruptor m y cambie la presión de la muestra a 800 mbar (Figura 2). Enjuague el sistema con CS2 (NaOH) durante 4 min.
- Cambie a la posición 3 del m-switch y enjuague el sistema con CS3 (IPA) durante 4 min.
- Cambie a la posición 1 del m-switch y enjuague el sistema con CS1 (PBS) durante 4 min.
- En el software de control de microfluídica, detenga todo el flujo ajustando las presiones de la línea de muestra y de la línea de tampón a 0 y cierre la válvula tampón.
- Desconecte el chip de la plataforma y vuelva a colocarlo en la placa de preparación. Desconecte todos los tubos y coloque el extremo del tubo de muestra en una botella de desecho. Limpie el objetivo con isopropanol y toallitas.
8. Medición de la muestra de fotometría masiva
- Desenrosque el tapón conectado al frasco tampón de 200 ml (pH 7,4) y enrósquelo al frasco tampón de 200 ml (pH 5,0).
- Repita los pasos 3.4-5.6, pero utilice el segundo canal del chip en lugar del primero.
- En el software de control de microfluídica, cambie el interruptor m a la posición 5 (correspondiente a la muestra), ajuste el caudal de la línea de muestreo a 8 μL/min y asegúrese de que la presión de la línea de muestra no supere los 350 mbar (Figura 2).
- Repita los pasos 6.2-6.4 para medir la muestra.
- Para calcular los caudales a utilizar, asegúrese de que la diferencia de pliegue en el caudal coincida con el factor de dilución de muestra deseado. Por ejemplo, para lograr la dilución de 2000x aquí, los caudales difieren en un factor de 2000; el caudal del tampón es de 1000 μL/min y el caudal de la muestra es de 0,5 μL/min. Al final de un experimento, deje siempre las líneas limpias siguiendo el protocolo de limpieza.
9. Análisis de datos
- Una vez finalizada la adquisición de datos, inicie el software de análisis de datos (Figura 5).
- Haga clic en el icono más (+) en la parte superior izquierda y seleccione el archivo calibrador .mp. El software comenzará a analizar el archivo cargado. Dependiendo del tamaño y el número de medidas, esto puede tardar unos minutos
- No analice los archivos .mp en el software de análisis de datos mientras adquiere datos con el software de adquisición de datos, ya que esto puede reducir la calidad de los datos que se adquieren.
- Presione el botón Crear calibración de masa (abajo a la derecha). Se abrirá un cuadro de diálogo que muestra una tabla rellenada con los valores de contraste de los picos ajustados.
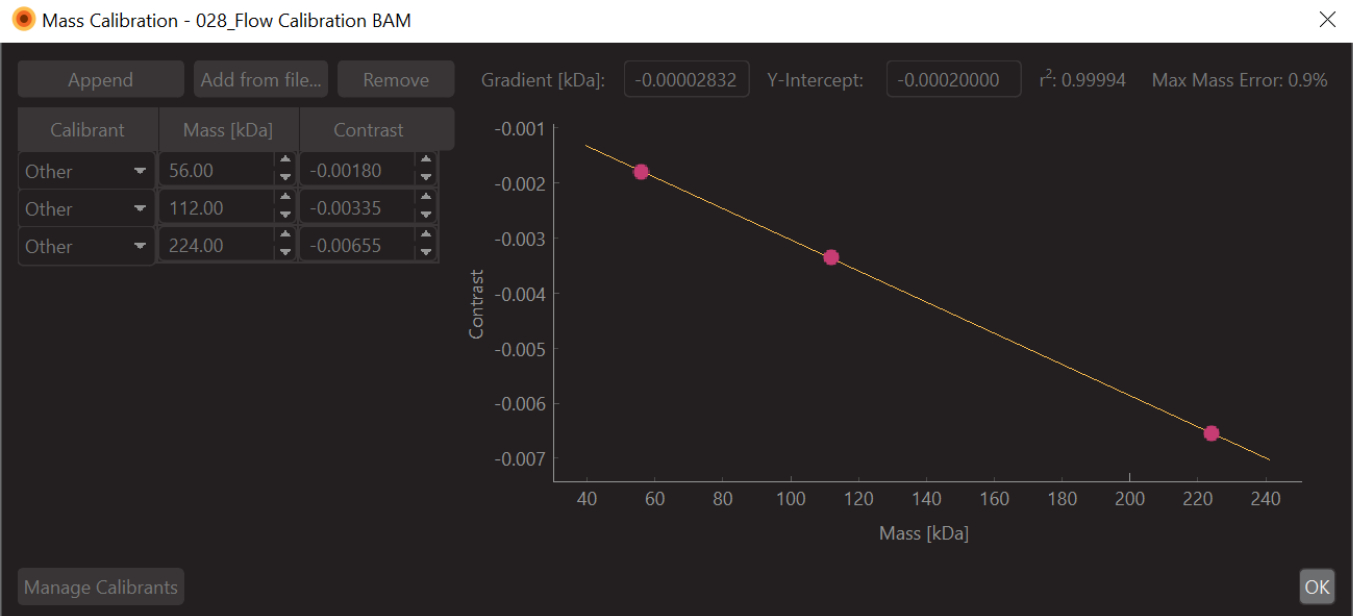
- Cambie los valores a los valores de masa conocidos para los picos ajustados (para la β-amilasa, estos valores son 56 kDa, 112 kDa y 224 kDa) y pulse Guardar. El archivo de calibración recién creado (extensión .mc) aparecerá en el panel Calibración de masa (abajo a la izquierda del software de análisis de datos) (Figura 6).
- Haga clic en el icono más (+) en la parte superior izquierda y seleccione el archivo de muestra .mp.
- Para crear un histograma de masa, como se muestra en la Figura 5, vaya a la pestaña Análisis , seleccione el modo Histograma y la opción Gráfico de masa . Ajuste el ancho de la bandeja, los límites de masa y otros parámetros según sea necesario.
- Personalice aún más el gráfico en la pestaña Figuras si lo desea antes de exportar figuras y/o guardar todo el espacio de trabajo como un archivo .dmp.
Resultados
Se utilizó fotometría masiva para medir la interacción entre el anticuerpo monoclonal IgG trastuzumab y el dominio soluble del receptor Fc neonatal de IgG (FcRn). Una mezcla 1:10 de las dos proteínas (a 2 μM para IgG y 20 μM para FcRn) se diluyó manualmente a 10 nM y 20 nM en PBS, respectivamente.
El paso de dilución es necesario porque la fotometría de masa, una técnica de medición de masa de una sola molécula, solo puede analizar muestras en el rango de 100 pM-100 nM. Intentar medir muestras con concentraciones fuera de este rango puede poner en peligro la precisión de los resultados (Figura 4). Este experimento no está incluido en este Protocolo, ya que ha sido descrito anteriormente 5,6.
En los histogramas de masas producidos en un experimento de fotometría de masas, la intensidad de la señal de dispersión (o "contraste") para cada evento de aterrizaje se traza en el eje x y se convierte en masa molecular a través del paso de calibración de contraste de masa. Mientras tanto, el eje Y indica el número de moléculas contadas con una masa (o contraste) dada. Por lo tanto, un pico indica la presencia de una población de moléculas dentro del rango de pesos moleculares que se muestra en el eje x. El número de eventos (recuentos) que componen el pico refleja el tamaño de esa población.
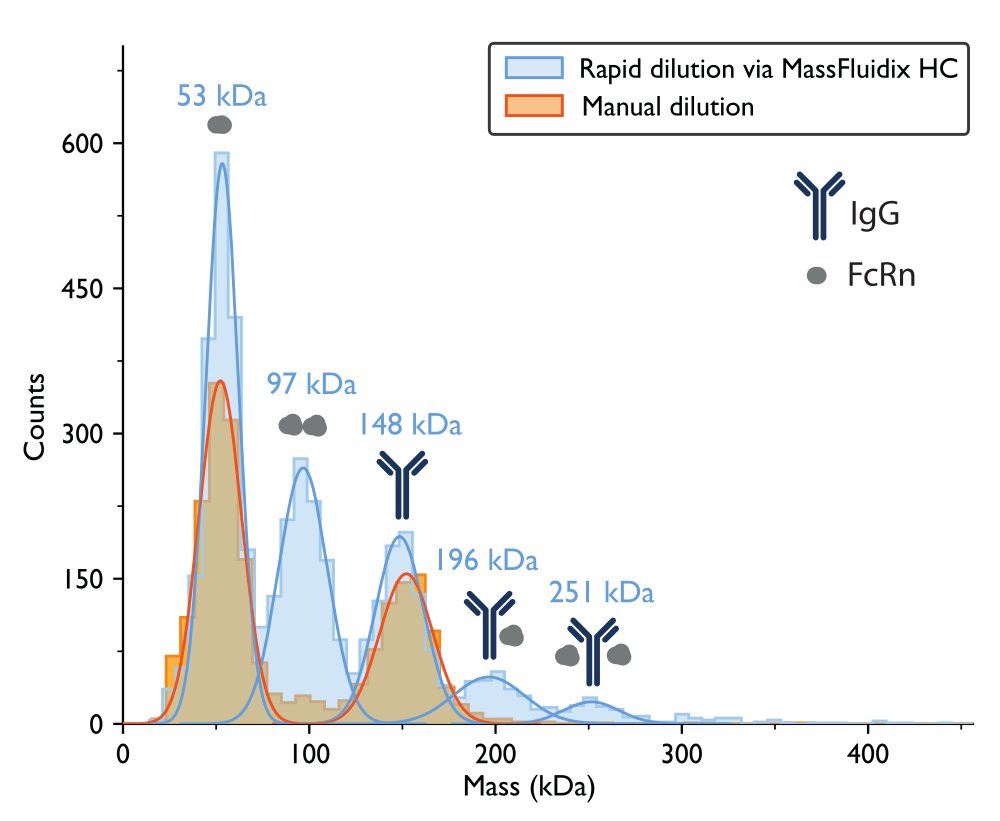
La medición de la fotometría de masas con dilución manual dio como resultado un histograma de masas en el que los dos picos más grandes, basados en los pesos moleculares esperados de las proteínas, correspondieron a monómeros de FcRn no unidos (~50 kDa) y monómeros de anticuerpos IgG (~150 kDa) (Figura 7). Al igual que los datos de fotometría masiva publicados anteriormente5, las especies no unidas eran prominentes, mientras que los picos en los rangos de masa que corresponderían a los complejos eran mucho menos evidentes.
El experimento se repitió utilizando un sistema de microfluídica de dilución rápida para diluir la mezcla a la concentración necesaria inmediatamente antes de la medición de la fotometría de masas. Este enfoque permitió la detección de complejos adicionales de baja afinidad, que pueden haberse disociado durante la etapa de dilución manual. Para lograr el mismo factor de dilución 2000x utilizado durante el experimento de dilución manual, la mezcla de muestra 1:10 (a una concentración de 2 μM para IgG y 20 μM para FcRn) se fluyó sobre el chip microfluídico a una velocidad de 0,5 μL/min, junto con tampón PBS (pH 5,0) a un caudal de 1 mL/min. Para garantizar que las proteínas no se degradaran en el momento de la medición, se midieron IgG (2 μM) y FcRn (20 μM) bajo flujo con el sistema microfluídico después de 120 min de incubación de las muestras. Las mediciones de control individuales no mostraron degradación de proteínas (Figura suplementaria 1)
Para la muestra que se sometió a una dilución rápida, se pudieron observar nuevamente los picos correspondientes a monómeros de FcRn (53 kDa), dímeros de FcRn (97 kDa) y monómeros de IgG (148 kDa). Además, se observaron claramente dos picos adicionales a 196 kDa y 251 kDa, correspondientes a complejos IgG-FcRn con estequiometrías 1:1 y 1:2 (Figura 7). Las masas esperadas para estos dos complejos eran de 200 kDa y 250 kDa, respectivamente. La variabilidad en la medición de masa está dentro de que el error de medición para el fotómetro de masa utilizado en este estudio es de ±5%14 (2% para el complejo 1:1 y 0,4% para el complejo 1:2).
La presencia de estos complejos solo en la muestra rápidamente diluida es consistente con la idea de que tienden a disociarse a concentraciones más bajas, a una velocidad que es rápida en relación con un proceso de dilución manual, pero lenta en relación con el proceso de dilución rápida logrado con el sistema de microfluídica15.

Figura 1: Combinación de microfluídica con fotometría de masas. (A,B) La caja de microfluídica utilizada en este protocolo, vista desde el frente (A) y la parte superior (B). Las soluciones de limpieza se colocan en las posiciones 1-3 y las muestras y calibrantes en las posiciones 4-6. Todas las soluciones están conectadas al m-switch, que a su vez está conectado al sensor de la unidad de caudal "Small". (C) Visión general de todo el sistema. La computadora (arriba a la izquierda) está conectada a la caja de microfluídica (que se muestra junto a los tubos tampón y de muestra) y al fotómetro de masas (abajo a la derecha), donde se encuentra el chip de microfluídica. El monitor de caudal pequeño (FRMS) monitorea el flujo a través de la línea de muestreo, mientras que el monitor de caudal grande (FRML) monitorea el flujo del búfer. La muestra y el tubo de la línea tampón se conectan a un canal en el chip de microfluídica, ubicado dentro del fotómetro de masas. (D) Cada chip tiene tres canales. El FRMS está conectado a la entrada de muestra y el FRML a la entrada del búfer. El área del mezclador es donde se lleva a cabo la dilución rápida de la muestra, mientras que el área de observación es donde se realizan las mediciones de fotometría de masas. La salida es monitoreada por un sensor de flujo adicional para garantizar que no haya fugas en el chip. Haga clic aquí para ver una versión más grande de esta figura.
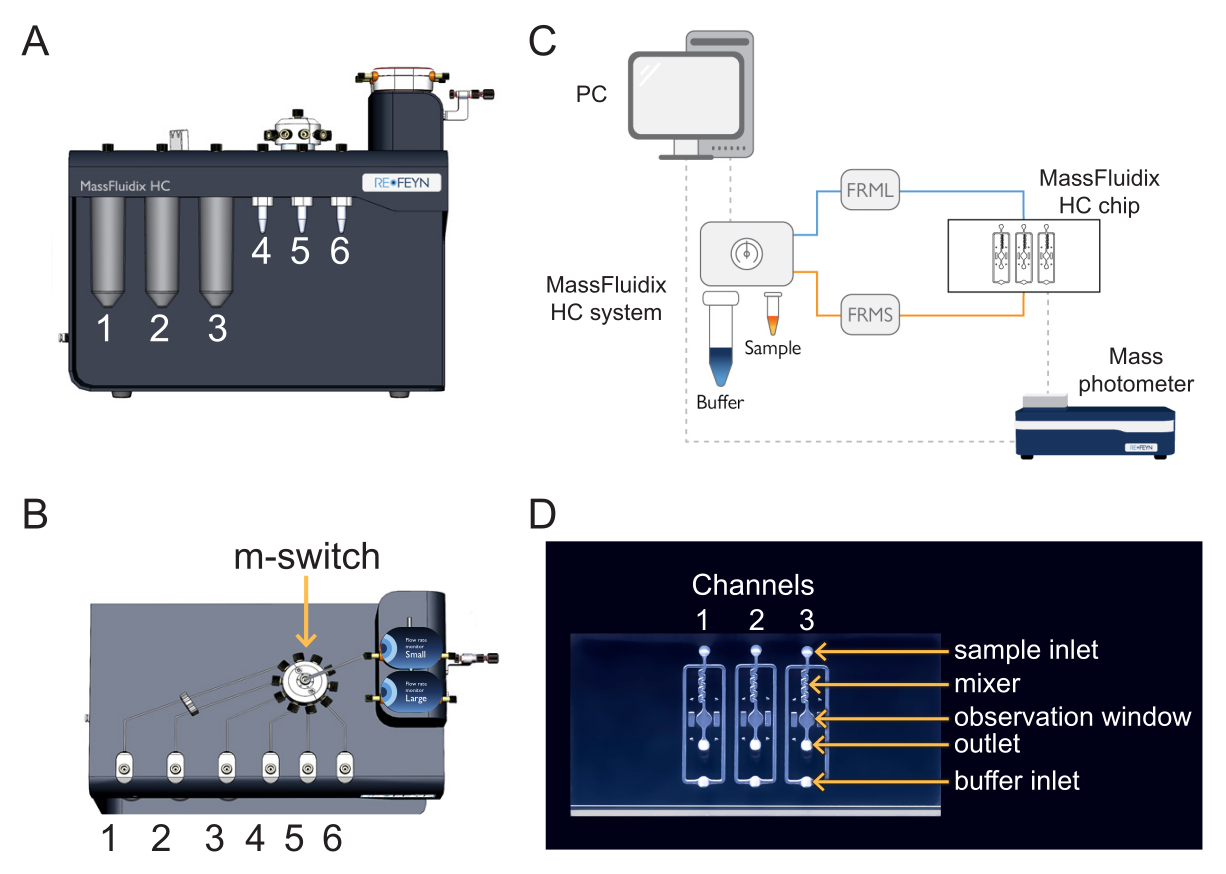{kind=link}

Figura 2: El software de control de microfluídica. Con este software, configure y controle los caudales, las líneas de presión del sistema microfluídico y la posición del interruptor múltiple (m-switch). Haga clic aquí para ver una versión más grande de esta figura.
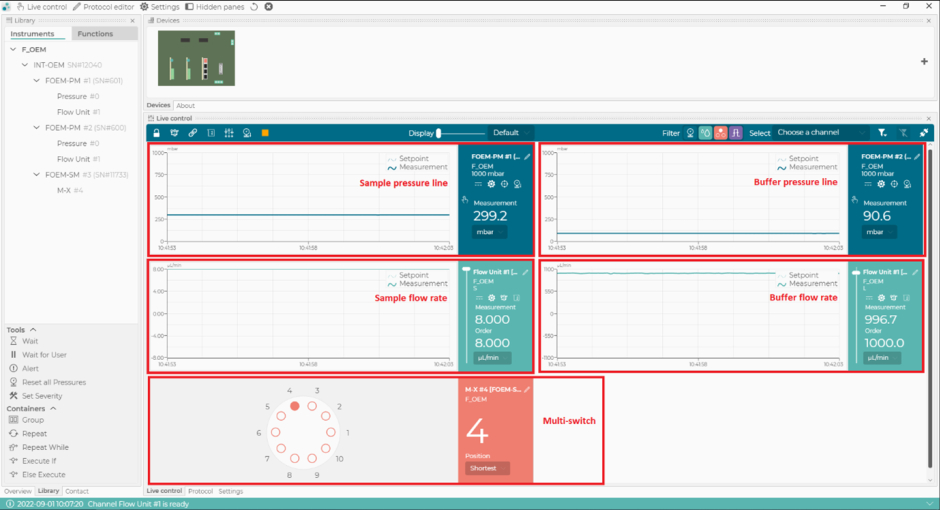{kind=link}

Figura 3: Identificación de la presencia de burbujas en el aceite. Los ejemplos del software de adquisición de datos muestran un anillo de enfoque claro e ininterrumpido (izquierda) y un anillo "roto", lo que indica la presencia de burbujas de aire en el aceite de inmersión (derecha). Haga clic aquí para ver una versión más grande de esta figura.
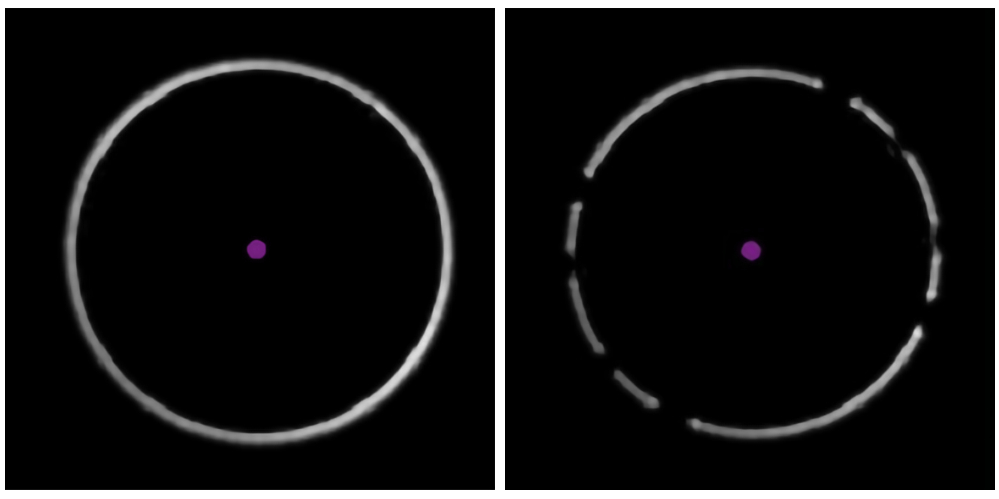{kind=link}

Figura 4: Ejemplos representativos de concentraciones de muestras en las que el número de eventos de aterrizaje de moléculas es demasiado poco, ideal o demasiado. Las moléculas que aterrizan en la superficie del área de observación en el chip de microfluídica aparecerán como manchas oscuras en la vista radiométrica de la imagen de fotometría de masas. De manera óptima, la concentración deseada debe permitir que se produzca una densidad ideal de eventos de aterrizaje durante la adquisición de datos (medio). Si la densidad del evento de aterrizaje es demasiado baja (arriba, 'Demasiado pocos'), no se puede completar un análisis estadístico preciso de los datos de fotometría masiva. Si es demasiado alto (abajo, 'Demasiadas'), las moléculas de aterrizaje se superpondrán espacialmente, lo que dará lugar a una mala calidad de los datos. Estas imágenes se capturaron con el software de adquisición de datos utilizando un fotómetro de masas. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 5: Captura de pantalla del software de análisis de datos para fotometría masiva. Aquí, los archivos .mp exportados desde el software de adquisición de datos se pueden cargar para analizar los datos. Se pueden generar figuras a partir de los datos procesados. Haga clic aquí para ver una versión más grande de esta figura.
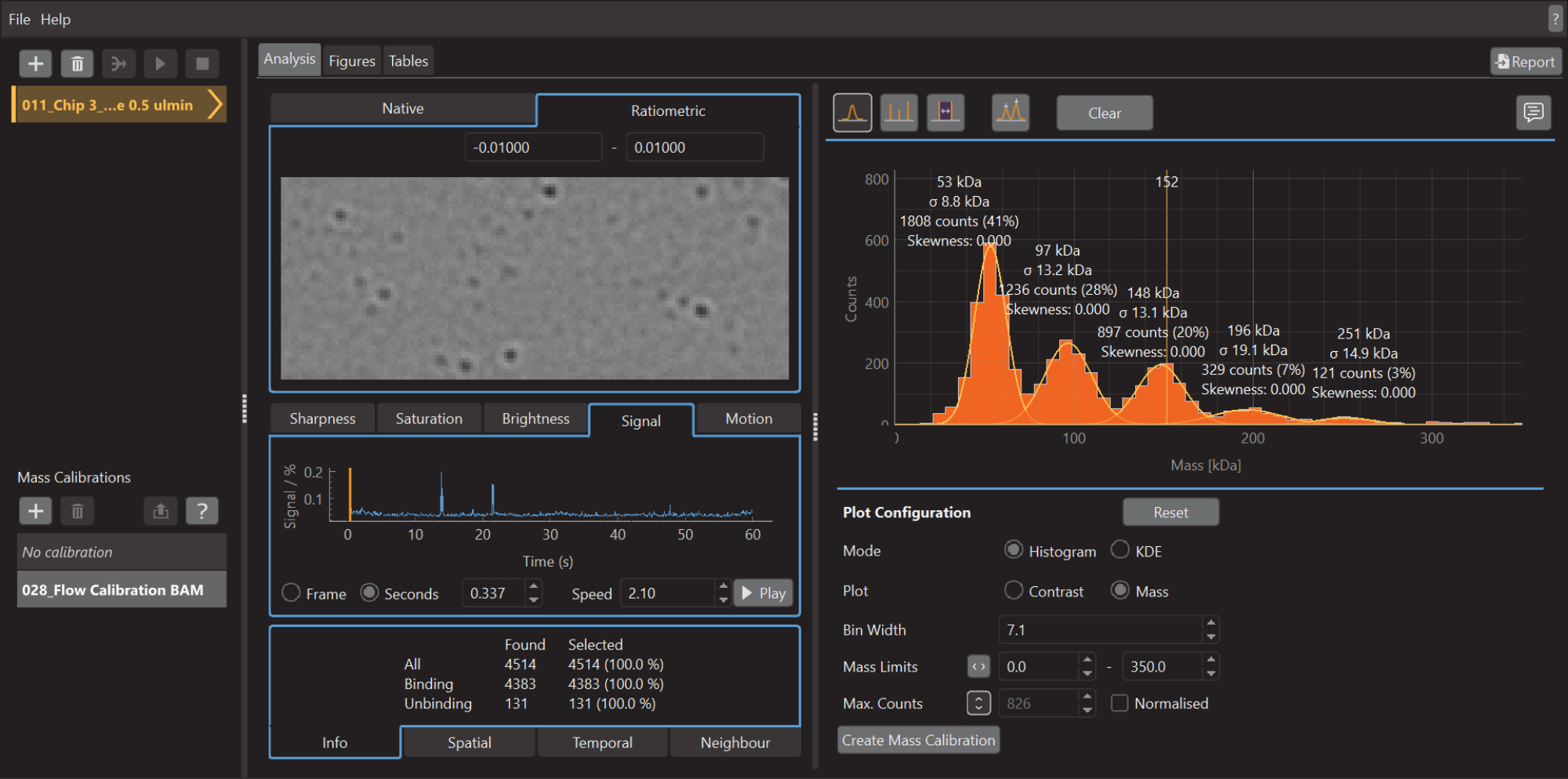{kind=link}

Figura 6: Una captura de pantalla de la calibración de contraste de masa para este experimento. El calibrante utilizado fue la β-amilasa, que se sabe que forma tres especies: monómero (56 kDa), dímero (112 kDa) y tetrámero (224 kDa). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 7: Los histogramas de masa revelan la formación de complejos solo después de una dilución rápida de la muestra. Se muestran los histogramas de masa y las correspondientes distribuciones gaussianas de mejor ajuste para las mediciones de muestras que contienen IgG y FcRn después de una dilución manual (naranja) o una dilución rápida a través del sistema microfluídicoHC (azul). Las etiquetas de masa indican que los valores medios de las curvas gaussianas se ajustan a los histogramas de masa medidos después de una dilución rápida. Después de la dilución manual, se observaron picos correspondientes a monómeros de FcRn (52 kDa, medidos con dilución manual) y monómeros de IgG (152 kDa). Después de una dilución rápida, además de los monómeros FcRn e IgG, también se observaron claramente picos correspondientes a dímeros de FcRn y complejos IgG-FcRn con estequiometría 1:1 y 1:2. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Datos complementarios Figura 1: Las mediciones de fotometría masiva de las muestras de control después de una dilución rápida no muestran degradación de proteínas. (A) Histograma de masa y distribución gaussiana de mejor ajuste para la muestra solo de FcRn. La concentración inicial de la muestra antes de la dilución rápida fue de 20 μM y el caudal se estableció en 0,5 μL/min. Se observaron picos de masa correspondientes a monómeros de FcRn (53 kDa) y dímeros de FcRn (97 kDa). (B) Histograma de masa y distribución gaussiana de mejor ajuste para la muestra de IgG (Herceptin) únicamente. La concentración inicial de la muestra antes de la dilución rápida fue de 2 μM y el caudal se estableció en 1 μL/min. Se pudo observar un único pico de masa correspondiente al monómero de IgG (155 kDa). Haga clic aquí para descargar este archivo.
Discusión
El protocolo descrito aquí proporciona un método para detectar y cuantificar las interacciones proteína-proteína de baja afinidad. Utiliza un fotómetro de masas acoplado a un sistema de microfluídica de dilución rápida. La fotometría de masas es una herramienta bioanalítica sin marcadores que puede medir de forma fiable la masa molecular en solución para biomoléculas16, para aquellas dentro del rango de 30 kDa a 6 MDa. Como la fotometría de masas es una técnica de una sola molécula que analiza las muestras una por una, generalmente se limita a muestras en el rango de concentración de 100 pM-100 nM. Por encima de este rango, las moléculas que aterrizan en la superficie del vidrio se superpondrán espacialmente, lo que dará lugar a una mala calidad de los datos; Por debajo de este rango, se obtienen muy pocos datos para realizar un análisis robusto7. Una consecuencia importante es que puede limitar la investigación de las interacciones de las proteínas a aquellas que forman una mezcla de especies unidas y no unidas dentro de ese rango.
Aquí, detallamos un protocolo paso a paso para usar un sistema de microfluídica de dilución rápida para expandir de manera efectiva el rango de concentraciones de muestras que son susceptibles de fotometría masiva. Al diluir la muestra en el chip microfluídico y luego hacerla fluir a través de la ventana de observación del detector dentro de los 50 ms, el sistema captura los complejos presentes en la muestra sin diluir antes de que cambie el equilibrio de interacción. La muestra se suministra continuamente al detector durante las mediciones individuales. En estas condiciones, el 95% del complejo permanecerá intacto cuando se mida la muestra, incluso para interacciones de baja afinidad, con un KD del orden de los micromolares y tasas de disociación tan rápidas como 1 s-1.
Esto se puede calcular de la siguiente manera: Para una reacción con una velocidad hacia adelante kfy una velocidad hacia atrás kb,

En equilibrio, las concentraciones de las tres especies (A, B y el complejo AB) permanecen constantes, por lo que  y
y  . Bajo la suposición conservadora de que la perturbación (dilución, en este caso) puede hacer que el complejo se disocie, pero la reacción directa (asociación) no procede, el término kf [A] [B] puede tratarse como despreciable, y se puede hacer la siguiente simplificación:
. Bajo la suposición conservadora de que la perturbación (dilución, en este caso) puede hacer que el complejo se disocie, pero la reacción directa (asociación) no procede, el término kf [A] [B] puede tratarse como despreciable, y se puede hacer la siguiente simplificación:

La integración da la siguiente expresión para la concentración de complejo en el tiempo después de la perturbación del equilibrio:

La fracción del complejo que permanece ligada en el tiempo t después de la perturbación del equilibrio es así:

A = 50 ms, para una reacción con kb≈ 1 s-1, el enlace de la fracción es 0,95, o 95%11,12.
La fotometría masiva se utilizó aquí y anteriormente5 para investigar la unión del anticuerpo monoclonal IgG trastuzumab al dominio soluble del FcRn. Se ha informado que los dos socios de unión se unen con afinidad nanomolar a pHácido 17. Se utilizó fotometría masiva para evaluar cualitativamente la abundancia de los complejos formados mientras los socios de unión estaban a pH 5.0, y las muestras se diluyeron rápidamente a través de un sistema microfluídico adicional. El procedimiento fue optimizado para la interacción proteína-proteína particular con base en los resultados reportados previamente5. El mismo procedimiento se puede utilizar para estudiar otras interacciones, siempre que los usuarios tengan conocimientos previos u optimicen las condiciones experimentales para el sistema en cuestión, como qué tampones utilizar, la concentración inicial de proteínas, la estequiometría esperada y la cantidad de incubación necesaria para permitir que la interacción alcance un equilibrio.
Cuando la mezcla IgG-FcRn se diluyó manualmente, fue difícil detectar la presencia de complejos IgG-FcRn, a pesar de que se sabe que estas proteínas interactúan5. Este trabajo muestra que el enfoque de dilución rápida da como resultado una cantidad notablemente mayor de estos complejos. Para la misma muestra, cuando se utilizó una dilución rápida, se observaron claramente complejos FcRn-IgG 1:1 y 2:1 FcRn-IgG. Estas diferencias en la formación de complejos demuestran la importancia de estudiar los sistemas de interacción biomolecular en una amplia gama de concentraciones.
Además, estos resultados también demuestran que es sencillo utilizar la microfluídica con el análisis de una sola molécula para capturar interacciones débiles, llenando un vacío significativo en el método. La combinación de la microfluídica de dilución rápida con la fotometría de masas ofrece ventajas atractivas debido a las ventajas de la fotometría de masas como técnica analítica. Es decir, la fotometría masiva no requiere etiquetas, implica una preparación mínima de la muestra y las mediciones se realizan en solución. Para este protocolo, otra ventaja clave de la fotometría masiva es su capacidad para distinguir y cuantificar todas las especies formadas (siempre que tengan una masa distinta de >30 kDa). Esto contrasta con la SPR, por ejemplo, que puede medir las tasas de unión y desunión, pero no puede proporcionar fácilmente información estequiométrica8.
Para este protocolo, así como para los experimentos de fotometría masiva en general, son útiles varias consideraciones. En primer lugar, la concentración final de proteínas debe estar dentro del límite de lo que la fotometría de masas puede medir (100 pM-100 nM). La concentración de incubación inicial también debe estar dentro del rango del sistema microfluídico (hasta 90 μM) y se teoriza que está por encima del KD real de la interacción10. El punto de partida recomendado es una proporción de mezcla de concentración de 1:1 entre las especies que interactúan a una concentración de μM. La relación podría entonces variarse a 1:2, 1:5 o, como en el caso de esta interacción, 1:10. Si no hay información previa sobre las interacciones de las proteínas, el usuario tendría que optimizar el experimento, comenzando con una concentración alta (recomendada 20 μM) para cada socio para determinar si la afinidad de los componentes está dentro del rango de concentración sostenido por el método presentado (es decir, se forman complejos). La optimización también puede implicar la elección de otras condiciones de búfer para promover las interacciones o la valoración de uno de los componentes de interacción para determinar la proporción de mezcla correcta. Una vez determinados, es posible optimizar las concentraciones y los caudales para permitir condiciones óptimas para el estudio y el método, por ejemplo, disminuyendo las concentraciones para permitir una mejor resolución de los picos.
En segundo lugar, para replicar con éxito este experimento, se deben minimizar las impurezas. Las fuentes comunes de impurezas que se sabe que afectan negativamente a las mediciones de fotometría masiva incluyen otras proteínas o desechos celulares que permanecen después de la purificación, tampones sin filtrar, detergentes formadores de micelas (si están presentes en una concentración demasiado alta) y tampones que contienen altas concentraciones de sal, glicerol u otros componentes. Como se discutió en el Protocolo anterior, las burbujas en el sistema microfluídico deben eliminarse. Se pueden formar burbujas en el sistema de tubos o si las muestras tienen una alta tensión superficial y son propensas a la formación de espuma. También se pueden formar burbujas en el aceite de inmersión, que se pueden detectar desde el anillo de enfoque (Figura 3). Si las burbujas no se pueden eliminar siguiendo los pasos descritos en el protocolo, otra solución es desgasificar la muestra con un desecador y una bomba de vacío, dejando la muestra a presión reducida durante unos minutos. No se recomienda agitar o agitar soluciones de proteínas altamente concentradas, ya que estas acciones pueden promover la formación de burbujas.
Si bien aquí se demuestra la medición de una interacción proteína-proteína específica, el mismo protocolo se puede aplicar a otros sistemas de interacción proteína-proteína sin modificaciones significativas. Otra dirección futura de este protocolo sería utilizar las mediciones para calcular los valores deK D para los complejos identificados, como se ha descrito en otro lugar en el contexto de la fotometría de masas 5,7. Si bien los estudios anteriores utilizaron datos de experimentos que involucraban dilución manual e interacciones más fuertes, el principio de análisis podría aplicarse fácilmente en este contexto, siempre que se implementen mejoras adicionales en el dispositivo microfluídico (como una mayor precisión del sensor de flujo y estabilidad de la bomba).
Más allá de las interacciones proteína-proteína, es probable que haya aplicaciones más amplias para el enfoque combinado de fotometría de masas y microfluídica de dilución rápida. La fotometría masiva se puede utilizar para evaluar la pureza, agregación y homogeneidad de la muestra 18,19; estudiar la oligomerizaciónde proteínas 20, el ensamblaje macromolecular21 o la polimerización22; y en otros ámbitos. El análisis de fotometría masiva también se extiende más allá de las proteínas; Se ha utilizado para investigar las interacciones entre los ácidos nucleicos y las proteínas23, las partículas virales24 y las nanopartículas25. Por lo tanto, este protocolo describe una aplicación importante de un sistema combinado de microfluídica de fotometría de masas: permite la medición directa de interacciones débiles proteína-proteína a nivel de moléculas y complejos individuales. El valor de la presente aplicación es alto, ya que abre la posibilidad de caracterizar directamente interacciones que generalmente han sido difíciles de estudiar, con relevancia en áreas terapéuticas críticas. Este enfoque combinado también podría servir como base para una gama más amplia de investigaciones para muestras con concentraciones de hasta decenas de micromolares.
Divulgaciones
Myndert Claasen y Zornitsa Kofinova son empleados de Refeyn Ltd, que produce el fotómetro de masas y el sistema de microfluídica utilizados en este artículo. Weston Struwe es accionista y consultor de Refeyn Ltd.
Agradecimientos
W.S. cuenta con el apoyo de una beca de Futuros Líderes de UKRI [MR/V02213X/1]. El texto y los gráficos del manuscrito se prepararon con el apoyo de los miembros del equipo de comunicación científica de Refeyn (Panagiota Paganopoulou, Neus Torres Tamarit y Catherine Lichten). También agradecemos los valiosos comentarios de Camille Hetez, Sofia Ferreira y Matthias Langhorst.
Materiales
| Name | Company | Catalog Number | Comments |
| 2-Propanol (Isopropanol) | VWR International LLC | 20880.320 | |
| Data acquisition software | Refeyn | AcquireMP (v2022 R1) | |
| Data analysis software | Refeyn | DiscoverMP (v2022 R1) | |
| FCRN, His-Tag | Sigma | SRP0624 | |
| Herceptin (IgG) | Cambridge Bioscience | HY-P9907-1mg | |
| Mass photometer | Refeyn | TwoMP | |
| Microfluidics box | Refeyn | MassFluidix HC system | |
| Microfluidics chip | Refeyn | MassFluidix HC chip | |
| Microfluidics control software | Fluigent | OxyGEN | |
| Phosphate Buffered Saline (PBS), 1x Ultra Pure | VWR International LLC | K812 | |
| Sodium Hydroxide (NaOH) | Sigma | S2770 | |
| β-Amylase, from sweet potato | Sigma | A8781 |
Referencias
- Arter, W. E., Levin, A., Krainer, G., Knowles, T. P. J. Microfluidic approaches for the analysis of protein-protein interactions in solution. Biophysical Reviews. 12 (2), 575-585 (2020).
- Hellenkamp, B., Thurn, J., Stadlmeier, M., Hugel, T. Kinetics of transient protein complexes determined via diffusion-independent microfluidic mixing and fluorescence stoichiometry. The Journal of Physical Chemistry. B. 122 (49), 11554-11560 (2018).
- Li, Z. Editorial: Methods in structural biology: Cryo-electron microscopy. Frontiers in Molecular Biosciences. 9, 1041386 (2022).
- Herling, T. W., et al. A microfluidic platform for real-time detection and quantification of protein-ligand interactions. Biophysical Journal. 110 (9), 1957-1966 (2016).
- Soltermann, F., et al. Quantifying protein-protein interactions by molecular counting with mass photometry. Angewandte Chemie International Edition. 59 (27), 10774-10779 (2020).
- Wu, D., Piszczek, G. Rapid determination of antibody-antigen affinity by mass photometry. Journal of Visualized Experiments: JoVE. 168, 61784 (2021).
- Wu, D., Piszczek, G. Measuring the affinity of protein-protein interactions on a single-molecule level by mass photometry. Analytical Biochemistry. 592, 113575 (2020).
- Young, G., et al. Quantitative mass imaging of single biological macromolecules. Science. 360 (6387), 423-427 (2018).
- Zijlstra, N., et al. Rapid microfluidic dilution for single-molecule spectroscopy of low-affinity biomolecular complexes. Angewandte Chemie International Edition. 56 (25), 7126-7129 (2017).
- MassFluidix® HC system for rapid dilution via microfluidics. Available from: https://www.refeyn.com/massfluidix-hc-system (2023)
- Pollard, T. D. A guide to simple and informative binding assays. Molecular Biology of the Cell. 21 (23), 4061-4067 (2010).
- Jarmoskaite, I., AlSadhan, I., Vaidyanathan, P. P., Herschlag, D. How to measure and evaluate binding affinities. eLife. 9, e57264 (2020).
- Monnet, C., et al. Selection of IgG variants with increased FcRn binding using random and directed mutagenesis: Impact on effector functions. Frontiers in Immunology. 6, 39 (2015).
- . Refeyn TwoMP: Transforming biomolecular characterisation Available from: https://www.refeyn.com/twomp-mass-photometer (2022)
- Lai, S. -. H., Tamara, S., Heck, A. J. R. Single-particle mass analysis of intact ribosomes by mass photometry and Orbitrap-based charge detection mass spectrometry. iScience. 24 (11), 103211 (2021).
- Wu, D., Piszczek, G. Standard protocol for mass photometry experiments. European Biophysics Journal. 50 (3-4), 403-409 (2021).
- Vaughn, D. E., Bjorkman, P. J. Structural basis of pH-dependent antibody binding by the neonatal Fc receptor. Structure. 6 (1), 63-73 (1998).
- Niebling, S., et al. Biophysical screening pipeline for Cryo-EM grid preparation of membrane proteins. Frontiers in Molecular Biosciences. 9, 882288 (2022).
- Paul, S. S., Lyons, A., Kirchner, R., Woodside, M. T. Quantifying oligomer populations in real time during protein aggregation using single-molecule mass photometry. ACS Nano. 16 (10), 16462-16470 (2022).
- Schulz, L., et al. Evolution of increased complexity and specificity at the dawn of form I Rubiscos. Science. 378 (6616), 155-160 (2022).
- Malay, A. D., et al. An ultra-stable gold-coordinated protein cage displaying reversible assembly. Nature. 569 (7756), 438-442 (2019).
- Hundt, N., Cole, D., Hantke, M. F., Miller, J. J., Struwe, W. B., Kukura, P. Direct observation of the molecular mechanism underlying protein polymerization. Science Advances. 8 (35), eabm7935 (2022).
- Acharya, A., et al. Distinct RPA domains promote recruitment and the helicase-nuclease activities of Dna2. Nature Communications. 12, 6521 (2021).
- Ebberink, E. H. T. M., Ruisinger, A., Nuebel, M., Thomann, M., Heck, A. J. R. Assessing production variability in empty and filled adeno-associated viruses by single molecule mass analyses. Molecular Therapy - Methods & Clinical Development. 27, 491-501 (2022).
- Melo, L., et al. Size distributions of gold nanoparticles in solution measured by single-particle mass photometry. The Journal of Physical Chemistry B. 125 (45), 12466-12475 (2021).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados