
Un abonnement à JoVE est nécessaire pour voir ce contenu. Connectez-vous ou commencez votre essai gratuit.
Method Article
Enregistrements de potentiel d’Action optique sous-type spécifique dans l’homme induit pluripotentes cellules souches dérivées des Cardiomyocytes ventriculaires
* Ces auteurs ont contribué à parts égales
Dans cet article
Résumé
Nous présentons une méthode pour image optiquement les potentiels d’action, plus précisément dans les cardiomyocytes de cellules souches dérivées ventriculaire comme pluripotentes induites. La méthode est basée sur l’expression pilotée par le promoteur d’une protéine fluorescente sensibles au voltage.
Résumé
Cardiomyocytes générés à partir des cellules souches humaines pluripotentes induites (iPSC-CMs) sont des outils émergents en recherche cardiovasculaire. Plutôt que d’être une population homogène de cellules, l’iPSC-CMs générés par des protocoles de différenciation actuels représentent un mélange de cellules avec ventriculaire-, auriculaire-et phénotypes nodal-like, qui complique les analyses phénotypiques. Ici, une méthode aux potentiels d’action optiquement Records spécifiquement des ventriculaire type iPSC-CMs est présentée. Ceci est réalisé par transduction des gènes avec une construction dans laquelle un détecteur de tension codé génétiquement est sous le contrôle d’un élément promoteur ventriculaire spécifique. Lorsque iPSC-CMs sont transduites avec cette construction, le capteur de tension est exprimé exclusivement dans les cellules ventriculaires-like, ce qui permet des enregistrements de potentiel membranaire optique sous-type spécifique à l’aide de la microscopie en fluorescence Time-lapse.
Introduction
Cardiomyocytes (CMs) dérivés des cellules souches pluripotentes induites (CISP) sont un outil émergent à décortiquer les mécanismes moléculaires des maladies cardiaques, d’enquêter sur nouveaux traitements et à l’écran pour cardiaques indésirables effets1,2 ,3. Dès le début, arythmogène maladies tels que les canalopathies ont été un objectif important de ce domaine de recherche4. Par conséquent, les méthodes pour étudier des phénotypes électriques de CMs, comme arythmies ou des changements dans la morphologie du potentiel d’action (PA), sont au cœur de cette technologie.
Une considération importante dans l’application de l’iPSC-CMs est que les protocoles actuels de différenciation cardiaques n’entraînent pas une population homogène de cellules. Au lieu de cela, ils sont plutôt un mélange de cellules ressemblant à n ud sinusal, auriculaire et ventriculaires CMs à différents niveaux de maturation5,6,7,8. Cette hétérogénéité peut être une source pertinente de la variabilité expérimentale, surtout si les paramètres tels que la durée des PA (APD) sont l’objet d’une enquête, qui diffèrent intrinsèquement entre sous-types CM (p. ex., l’APD est plus courte dans auriculaire que dans CMs ventriculaires). L’approche classique pour régler ce problème est d’enquêter sur iPSC-CMs unique à l’aide de la méthode de serrage de patch et de classer chaque cellule sous la forme nodale-, auriculaire-, ou ventriculaire-like, basée sur la morphologie de son AP9. Toute analyse ultérieure peut être restreint alors aux cellules représentant le sous-type de CM d’intérêt. L’inconvénient majeur de cette stratégie est son débit limité et le manque d’évolutivité. En outre, le caractère invasif de l’électrophysiologie de patch clamp ne permet pas de l’imagerie des cellules mêmes séquentiellement sur une période de temps prolongée.
Ici, nous fournissons des détails expérimentaux sur une méthode10 mis au point pour l’image optiquement APs en sous-types spécifiques de l’iPSC-CMs. Cela permet de surmonter le problème de l’hétérogénéité de sous-type et augmente considérablement le débit par rapport aux méthodes conventionnelles, permettant le phénotypage rapide du CISP-CMs transportant des variants génétiques ou d’être des agents exposés à pharmacologique.
Aperçu de l’approche d’imagerie optique du sous-type spécifique
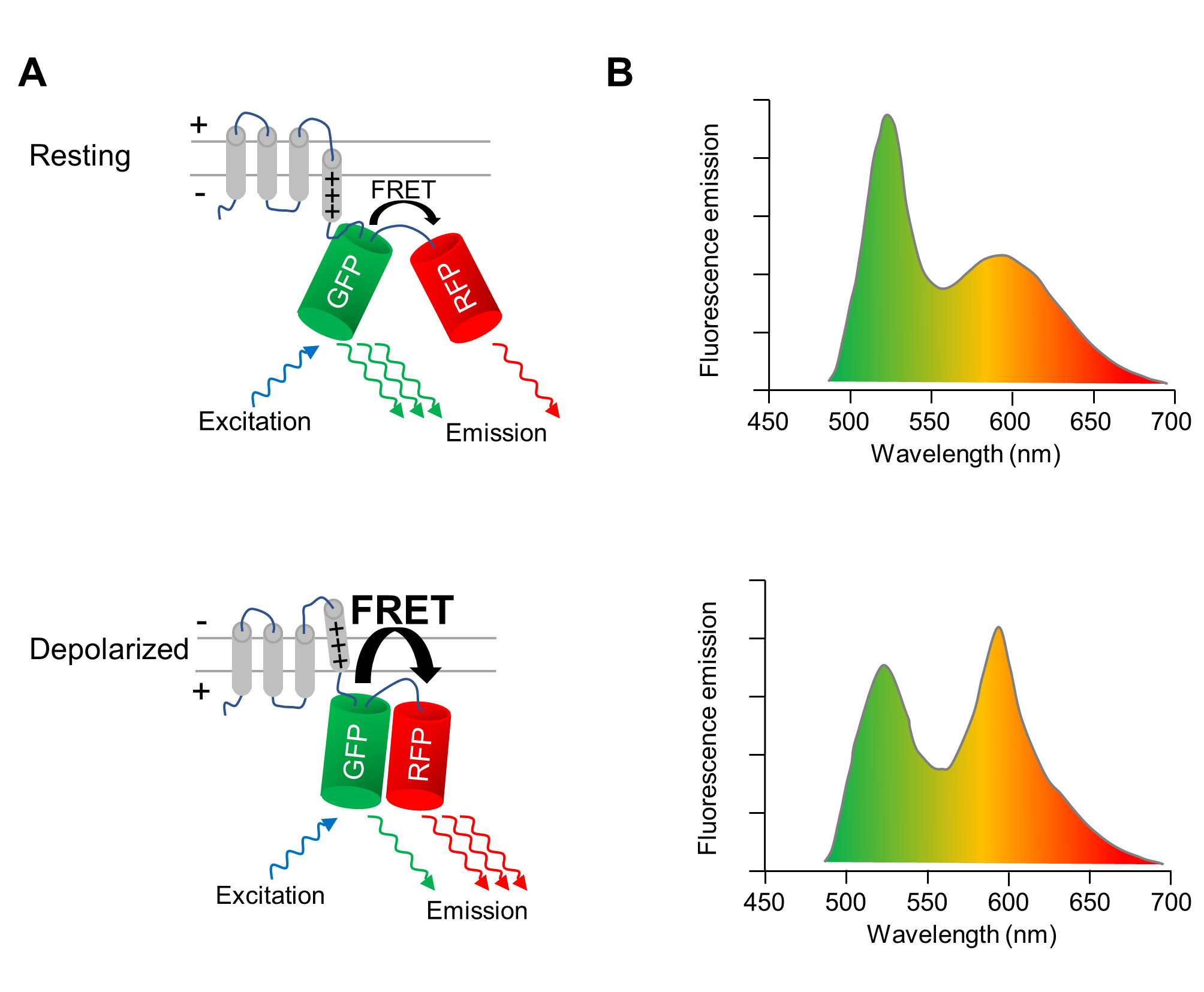
Un indicateur de tension codé génétiquement (GEVI), dont les propriétés fluorescence changent lors de la dépolarisation et la repolarisation de la membrane cellulaire, est utilisé pour image optiquement les changements du potentiel membranaire des CMs. Le GEVI appliqué ici est la protéine fluorescente-détection de tension VSFP-CR11, qui se compose d’un domaine transmembranaire détection tension fusionné à une paire d’un vert (trèfle) et une protéine fluorescente rouge (mRuby2) (Figure 1 a). En raison de la proximité des deux fluorophores, l’excitation de la protéine fluorescente verte résulte en une fraction de l’énergie d’excitation étant transféré à la protéine fluorescente rouge par transfert d’énergie de résonance Förster (FRET). Par conséquent, l’excitation de la protéine fluorescente verte entraîne une émission à la fois le vert et les rouges protéines fluorescentes (Figure 1 a, panneau supérieur). Lorsque la cellule dépolarise, un réarrangement structural de la sonde de tension se produit qui se traduit par une réorientation des deux protéines fluorescentes, augmentant ainsi l’efficacité de la frette. Ainsi, encore plus de l’énergie d’excitation est transféré du vert à la protéine fluorescente rouge (Figure 1 a, panneau inférieur). Ainsi, dans une cellule dépolarisée, l’émission de fluorescence verte est plus faible, et l’émission de fluorescence rouge est plus brillante que dans une cellule au repos le potentiel de membrane (Figure 1 b).

Figure 1 : l’imagerie optique du potentiel avec VSFP-CR. de membrane (A) A schéma illustrant l’action de la protéine fluorescente sensibles à la tension QUE VSFP-CR est montré. Lors de la dépolarisation de la membrane cellulaire, un réarrangement structural du domaine transmembranaire-détection de tension se traduit par une réorientation de la verte (GFP) et rouge protéine fluorescente (DP), augmentant l’efficacité de la Förster intramoléculaire transfert d’énergie par résonance (FRET). Les spectres (B) l’émission d’une VSFP sur l’excitation de la GFP dans les cellules au potentiel de repos membranaire (panneau du haut) et dans les cellules dépolarisées (panneau inférieur) sont représentés. Le changement spectral sur dépolarisation est exagéré pour plus de clarté. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
Les changements dans l’efficacité de la frette reflétant les fluctuations du potentiel membranaire sont imagés à l’aide d’un microscope à fluorescence équipé d’un séparateur de l’image, qui sépare l’émission de fluorescence rouge et vert et les projette sur deux zones adjacentes de la puce d’une caméra sCMOS (Figure 2). Avec ce set-up, l’émission de fluorescence à deux bandes de longueur d’onde différente peut être enregistrée simultanément, ce qui permet le calcul d’un ratio de fluorescence rouge au vert afin de refléter le potentiel de membrane dans chaque image d’une série de Time-lapse.

Figure 2 : Configuration du système d’imagerie. Les principaux composants du système d’imagerie utilisé pour image les modifications spectrales de la protéine fluorescente sensibles au voltage, reflétant les changements potentiels de membrane à une haute résolution temporelle sont représentées. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
L’expression de VSFP-CR dans CMs est obtenue par transduction des gènes. Pour diriger l’expression au sous-type CM d’intérêt, le lentivirus contient un élément de promoteur (le renforceur de MLC2v ) qui anime spécifiquement transcription ventriculaire type iPSC-CMs10. Lorsque l’iPSC-CMs qui représentent un mélange de cellules de type auriculaire, nodal, ventriculaire-type et sont transduites avec ce lentivirus, VSFP-CR est exprimée uniquement dans les cellules ventriculaires. Étant donné que l’imagerie optique potentiel d’action dépend de cette sonde fluorescente, les potentiels d’action enregistrés représentent exclusivement le sous-type de CM d’intérêt (Figure 3).

Figure 3 : expression VSFP pilotée par le promoteur pour l’imagerie de potentiel membranaire sous-type spécifique. (a) ce schéma montre comment enregistrements de potentiel d’action optique sous-type spécifique cardiomyocyte sont atteints. (b) iPSC-CMs infectés par un VSFP sous le contrôle de le MLC2v-amplificateur de ventriculaire spécifiques sont indiqués. L’expression de la sonde de tension est observée seulement dans CMs ventriculaire-comme dans le chenal de la GFP (panneau de gauche). Le contraste de phase (panneau central) et une image de superposition (panneau de droite) sont également fournis. Les pointillés blancs marquent les limites d’une cellule. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
Access restricted. Please log in or start a trial to view this content.
Protocole
1. préparation des Cardiomyocytes dérivés iPSC pour l’imagerie
NOTE : Procédés de différenciation de l’iPSC culture et cardiaques ont été publiés avant12,13,14 et ne sont pas discutés ici en détail. La purification de l’iPSC-CMs par microdissection manuel, la séparation des cellules magnétiques ou sélection de lactate est recommandée, selon le protocole de différenciation utilisé. Pour le protocole suivant, microdissected des explants de battre les aires, générées à l’aide d’une monocouche différenciation protocole13, ont été pris le 15e jour d’une différenciation cardiaque et cultivés jusqu'à 30 jours sur fibronectine enduit les plaques comme décrit avant le10.
- Prendre la cellule plaques de culture hors de l’incubateur et collecter manuellement les explants cardiaques dans des tubes de microcentrifuge à l’aide d’une pipette de 200 µL. Après sédimentation, aspirer le surnageant de culture cellulaire et laver délicatement les explants de symétrique 2 x avec Hank sel solution (HBSS).
- Dissocier l’iPSC-CMs en ajoutant collagénase type II (430 U/mL chez HBSS) et les incuber à 37 ° C pendant 30 min.
- Après que la sédimentation de la cellule de grande touffes, recueillir le surnageant qui contient l’iPSC-CMs unique dissociées et l’ajoute à un milieu CM entretien frais contenant une concentration élevée sérum fœtal (SVF) (DMEM-F12, 20 % FBS, 2 mM L-glutamine, 1/100 MEM non essentiels des acides aminés, 100 U/mL de pénicilline-streptomycine et 0,1 mM β-mercaptoéthanol).
- Répétez la dissociation si les touffes de grandes cellules restent. Frais virés single iPSC-CMs par centrifugation et les remettre en suspension dans un milieu CM contenant une faible concentration de FBS (2 % FBS).
- Réensemencez unique iPSC-CMs dans une densité permettant l’imagerie ultérieure des cellules individuelles sur un 3,5 cm fond en verre cellulaire microcuvette de culture qui a été recouvert de la fibronectine (2 µg/cm2). Optimiser la densité de semis avant de s’engager dans des expériences de haut débit. Habituellement, les explants cardiaques de 5 à 10 par plat de 3,5 cm sont suffisants ; Toutefois, cela dépend fortement la taille de l’explant et la pureté.
- La culture de l’iPSC-CMs unique après leur dissociation dans un milieu d’entretien CM contenant 2 % FBS pendant au moins 48 h afin de garantir une récupération suffisante.
Remarque : Dans les prochaines étapes, l’iPSC-CMs sont transduites avec un lentivirus encodage GEVI VSFP-CR sous le contrôle d’un rehausseur de MLC2v 10 pour obtenir une expression de GEVI ventriculaire spécifique. Constructions des gènes permettant d’exprimer la GEVI spécifiquement dans les cellules de nodal ou auriculaire-type, ou une construction non-spécifique hébergeant le promoteur PGK ubiquitaire exprimés, peut également être utilisé10. Des gènes plasmides sont disponibles sur demande.
ATTENTION : Lorsque vous travaillez avec Particules Lentivirales, veiller à ce que le milieu de travail dispose du niveau de biosécurité adéquat, respecter les consignes de sécurité pour travailler avec des agents potentiellement infectieux et prendre les précautions de sécurité nécessaires. - Préparer le milieu de l’infection en mélangeant les lentivirus10-contenant de culture cellulaire surnageante, qui a été produite dans les cellules HEK293 comme décrit avant15, avec support de maintenance CMs (Voir l’étape 1.3) dans un rapport 1:1.
- Ajouter le bromure de hexadimethrine à une concentration finale de 8 µg/mL pour améliorer l’efficacité de l’infection.
Remarque : En raison de l’infection haute efficacité de concentration préalable iPSC-CMs de lentivirus par ultracentrifugation est habituellement pas nécessaire. - Aspirer le support de maintenance de CM de l’iPSC-CMs simples et remplacez-le par le milieu d’infection établi au cours des étapes 1.7 et 1.8. La culture de l’iPSC-CMs unique infectés dans le milieu de l’infection pendant 12 h à 37 ° C. Ensuite, aspirer le milieu et le remplacer à nouveau avec support de maintenance de CM.
Remarque : Un signal de fluorescence de la GEVI apparaît 48 h après l’infection. Une imagerie des enregistrements potentiel membranaire est recommandé, 72 h après l’infection dans les meilleurs délais, afin d’assurer une force de signal correct. Seul iPSC-CMs infectés peuvent être cultivés pendant au moins 3 semaines et imagés séquentiellement. En cas de vaste photoblanchiment lors d’une session d’imagerie, le signal de fluorescence récupère au fil du temps. - Avant l’imagerie, échanger le milieu de culture cellulaire par une solution de Tyrode additionnée de Ca2 + (135 mM NaCl, KCl 5,4 mM, 1 mM MgCl2, 10 mM de glucose, 1,8 mM CaCl2et 10 mM HEPES ; pH 7,35).
2. optique Membrane éventuels enregistrements
- Placez la chambre d’imagerie contenant l’iPSC-CMs sur la scène d’un microscope à épifluorescence inversé équipé d’un séparateur d’image, les jeux de filtres appropriés et un appareil photo (par exemple, une caméra sCMOS) capable d’imagerie à haute vitesse à un niveau élevé sensibilité.
NOTE : par exemple, utilisez une excitation passe-bande de 480/40 nm filtre combiné avec un miroir dichroïque 500 nm long-pass dans le microscope, et un miroir dichroïque de 568 nm long-col combiné avec 520/28 nm et 630/75 nm passe-bande d’émission filtres dans le séparateur d’image. Pour l’imagerie de cellules individuelles, un objectif à immersion d’huile à fort grossissement, grande ouverture numérique offre le meilleur rapport signal sur bruit. - Si la stimulation électrique est nécessaire, installer des électrodes de stimulation dans la salle d’imagerie et connectez-les à un générateur d’impulsion.
NOTE : Nous avons utilisé une stimulation en médaillon contenant deux électrodes de platine et monté dans les récipients de culture de 3,5 cm fond en verre cellulaire. Les cellules situées entre les électrodes ont été projetés. Paramètres de stimulation typiques sont des impulsions rectangulaires de 5 ms dans la durée, d’une amplitude de distance des électrodes 10 V/cm. - Éventuellement, utiliser une platine du microscope chauffée afin d’assurer une température stable de l’iPSC-CMs lors d’imagerie, ou même un microscope d’incubation chambre Set-up si l’imagerie à long terme est destiné.
- Mettre les cellules à se concentrer et placer une cellule exprimant la GEVI dans le centre de la champ de vision.
Remarque : Plus commodément pour cela à l’aide des oculaires du microscope à l’aide d’une protéine fluorescente verte (GFP) ou un rouge jeu de filtres de protéine fluorescente (DP) pour identifier les cellules exprimant le GEVI. - Définissez le chemin optique afin que la lumière émise est acheminé vers le séparateur d’image. Basculer le cube du filtre au microscope à celui à combiner avec les filtres dans le séparateur d’image (voir la Table des matières).
- Dans le logiciel de commande de la caméra, définissez les paramètres d’acquisition afin que l’imagerie haute vitesse (par exemple, 100 images par seconde, avec un temps de pose de 10 ms par image) est possible.
Remarque : Généralement, cela implique la sélection de pixels appareil photo (par exemple, 8 x 8 pixels de la puce de la caméra sont mis en cellule pour générer un pixel dans le film Time-lapse). - Mettre en place le séparateur d’image selon les instructions du fabricant. Les deux images représentant les deux bandes de longueur d’onde d’émission différentes doivent être adjacents à l’autre, chacune occupant une moitié de l’image.
Remarque : Cette étape doit être exécutée uniquement 1 x au début de chaque séance d’imagerie. - Vérifier et, si nécessaire, régler la netteté de l’image de la caméra.
- Régler la luminosité de l’éclairage afin d’éviter toute saturation des pixels est ; Cela peut être fait plus facilement à l’aide d’une palette de couleurs dans lequel pixels saturés sont représentés par une couleur unique.
Remarque : Au cours de toutes ces préparations avant l’imagerie, limiter l’exposition de l’échantillon à l’excitation lumineuse à la quantité nécessaire (c'est-à-dire, ne pas laisser la lumière sur de longues périodes de temps) pour éviter le photoblanchiment de la GEVI. - Mettre en place l’acquisition de la série temporelle (p. ex., 5 000 images à 100 images par seconde pour une durée de la série de 50 s). La pénombre dans la pièce (y compris les écrans d’ordinateurs) pour éviter la lumière parasite qui influent sur la mesure. Si l’obturateur légère excitation n’est pas contrôlée par le logiciel d’imagerie, l’ouvrir juste avant de commencer l’acquisition.
- Démarrer l’acquisition de la série d’image. Si la stimulation électrique est désirée, lance la séquence de stimulation au point de temps approprié, sauf s’il est lancé automatiquement par le logiciel d’imagerie. Si la demande d’un agent pharmacologique est recherchée, le faire soit manuellement (par exemple, par pipetage 100 µL de l’agent dans une concentration de fin dix fois à la chambre d’enregistrement contenant 900 µL de tampon et mélanger doucement avec la pipette) ou en utilisant un système de perfusion de débit constant.
- Lorsque l’acquisition est terminée, fermer l’obturateur légère excitation si nécessaire. Sauvegarder l’enregistrement sur le disque dur.
- Laissant les paramètres d’enregistrement (p. ex., temps d’exposition, cadence et intensité lumineuse) inchangés, effectuer un enregistrement de la fluorescence de fond (comme au point 2.11) au-dessus d’une région de la lamelle ne contenant ne pas de cellules ou une autre lamelle couvre-objet sur lequel aucun les cellules ont été ensemencées.
Remarque : Si les mesures séquentielles sont effectuées sans modifier les réglages d’acquisition, qu’un seul enregistrement de fluorescence de fond est nécessaire pour toutes ces mesures. - Si vous le souhaitez, réaliser des expériences supplémentaires (mesures 2.4 – 2.13) sur différents iPSC-CMs, situés sur la lamelle couvre-objet même. Sachez que tous les CMs sur la lamelle couvre-objet seront affectées dans le cas où un médicament a été appliqué.
3. analyse
> NOTE : Suivant le logiciel d’imagerie utilisées (ce qui est généralement un paquet de logiciels propriétaires fourni par le fabricant de l’appareil photo ou de la fluorescence de toute système d’imagerie), il peut être possible d’effectuer l’analyse des images acquises en partie ou même entièrement dans ce progiciel. Toutefois, ici un flux de travail d’analyse d’image qui peut être effectuée avec le logiciel open source (c'est-à-dire, la plateforme d’analyse image ImageJ)16, qui peut être commodément installé en utilisant une distribution comme Fidji17et le progiciel R pour calcul statistique18 sont décrites. Brièvement, régions d’intérêt (ROIs) qui représente les cellules ou arrière-plan sont dessinées dans ImageJ et la fluorescence moyenne dans ces ROIs au fil du temps est exportée vers un fichier, puis, encore analyser en R ou, alternativement, avec le logiciel de feuille de calcul.
- Les logiciels d’imagerie, enregistrer ou exporter les séries temporelles d’image acquise (l’enregistrement contenant les cellules et l’enregistrement en arrière-plan correspondant) à un format (par exemple, TIF) qui peut être lu par ImageJ. Vous pouvez également utiliser le plugin de BioFormats (https://imagej.net/Bio-Formats), qui fournit des routines permettant ImageJ lire les formats propriétaires de différentes marques de fabricants de systèmes d’imagerie.
Remarque : Les étapes suivantes supposent que les intervalles d’imagerie sont les mêmes tout au long de l’enregistrement. Si ce n’est pas le cas, il peut être nécessaire d’importer les informations de calendrier depuis le logiciel d’imagerie et l’inclure dans l’analyse. - Ouvrez la pile TIF contenant les cellules dans ImageJ.
- Utilisez l’outil de sélection à main levée pour dessiner un ROI sur un cardiomyocyte fluorescent dans le canal rouge. Veiller à ce que le retour sur investissement est assez grand pour que la cellule ne bouge pas sur le retour sur investissement tout en contractant.
- Ouvrez le plugin manager de ROI ('Analyze | Outils | Gestionnaire de ROI') et appuyez sur le bouton « Ajouter [traduction] » pour ajouter ce ROI comme ROI1.
- Faites glisser le ROI au cours de la même cellule dans le canal vert et ajouter ce ROI comme ROI2.
- Dans le ' analyser | Définir des mesures menu, décochez toutes les options sauf « Signifie la valeur de gris ».
Remarque : Cela doit être fait seulement 1 x dans une session de ImageJ. - Dans le gestionnaire de ROI, appuyez sur la ' plus | Mesure multi' bouton. Sélectionnez les options « Mesurer toutes tranches » et « Une ligne par tranche » et appuyez sur « OK ». Une fenêtre s’ouvre contenant une feuille de calcul comportant trois rangées (le numéro de tranche et la fluorescence moyenne dans la deux ROIs, ce qui représente la cellule dans le rouge et dans le canal vert, respectivement).
- Utilisation ' fichier | Enregistrer sous pour enregistrer la feuille de calcul dans un fichier de valeurs séparées par des virgules (CSV).
- Fermez la fenêtre avec la pile de l’image et la fenêtre de feuille de calcul sans fermer la fenêtre de gestionnaire de ROI. Ouvrez la pile TIF contenant la mesure de l’arrière-plan.
- Dans le gestionnaire de ROI, appuyez sur la ' plus | Mesure multibouton' . Sélectionnez les options « Mesurer toutes tranches » et « Une ligne par tranche » et appuyez sur « OK ».
- Utilisation ' fichier | Enregistrer sous pour enregistrer la feuille de calcul avec les données de base dans un autre fichier CSV.
Remarque : Les calculs suivants peuvent s’effectuer avec le logiciel de feuille de calcul. Nous avons utilisé le logiciel R18. Un exemple simple de script qui lit les données cellulaires et l’arrière-plan des fichiers « cell.csv » et « background.csv » effectue les calculs et trace l’évolution temporelle du signal potentiel membranaire est fourni comme supplémentaire 1 fichier de Code. - À partir de l’enregistrement en arrière-plan, calculer l’intensité moyenne dans le rouge et dans le canal vert. Soustrayez ce signal de fond les signaux qui représente la cellule située dans le canal rouge et vert pour calculer DP corrigées et les signaux GFP, respectivement.
- Comme un substitut pour le potentiel de membrane, calculer le rapport DP/GFP.
NOTE : Ce rapport est sans dimension. Alternativement, il peut être normalisé à sa valeur initiale (par exemple, ΔR/R,0). Ce qui est important, on trouvera des informations utiles dans les changements temporels plutôt que dans les valeurs absolues de ce ratio. En raison de l’inégal photoblanchiment de la GFP et la DP, une dérive de la base de ce rapport peut se produire. Si nécessaire, une telle dérive de base peut être corrigée en construisant une courbe de référence et il en soustrayant le rapport DP/GFP10.
Access restricted. Please log in or start a trial to view this content.
Résultats
Dans la Figure 4 a, un seul représentant iPSC-CM est représenté avec des pointillés blancs marquant le retour sur investissement tirée lors de l’analyse d’imagerie dans la DP (côté gauche) et le canal de la GFP (côté droit). Le signal du canal DP montre une augmentation périodique dans l’intensité de fluorescence au cours de chaque potentiel d’action (Figure 4 b, panneau supérieur). Comme décrit dans l’Intr...
Access restricted. Please log in or start a trial to view this content.
Discussion
La méthode décrite ici permet un enregistrement optique des APs d’un sous-type spécifique (c.-à-d., cellules ventriculaires) de CMs produit du CISP humaine. IPSC-CMs humaines représentent un outil émergent d’aborder une grande variété de problèmes médicaux et biologiques, et la différenciation de différents sous-types de CM est une source importante de la variabilité expérimentale. En utilisant des éléments de promoteurs spécifiques, l’expression d’un GEVI est spécifiquement réalisée ...
Access restricted. Please log in or start a trial to view this content.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Ce travail a été soutenu par des subventions de la Fondation de recherche allemande (Si 1747/1-1), l’autre Kröner-Fresenius-Stiftung et la Deutsche Stiftung für Herzforschung.
Access restricted. Please log in or start a trial to view this content.
matériels
| Name | Company | Catalog Number | Comments |
| ß-Mercaptoethanol | Invitrogen | 21985023 | |
| DMEM-F12 Medium | Invitrogen | 21331046 | |
| FBS (Fetal Bovine Serum) | Invitrogen | 16141079 | |
| MEM Non-Essential Amino Acids | Invitrogen | 11140050 | |
| GlutaMax-I Supplement | Invitrogen | 35050061 | alternative L-Glutamine |
| Penicillin-Streptomycin | Invitrogen | 15140122 | |
| Fibronectin bovine plasma | Sigma-Aldrich | F1141 | |
| Collagenase type II | Worthington Biochem | LS004174 | |
| Hexadimethrine Bromide (Polybrene) | Sigma-Aldrich | H9268 | enhancing lentiviral infection |
| 3.5 cm glass-bottom microdishes | MatTek corporation, Ashland, MA, USA | P35G-1.5-14-C | |
| Microscope stand | Leica Microsystems, Wetzlar, Germany | DMI6000B | |
| Microscope objective | Leica Microsystems, Wetzlar, Germany | HCX PL APO 63X/1.4-0.6 Oil | |
| sCMOS camera | Andor Technology, Belfast, UK | Zyla V | |
| Microscope filter cube: excitation filter | Chroma Technology Corp, Bellows Falls, VT, USA | ET480/40X | bandpass 480/40 |
| Microscope filter cube: dichroic mirror | Chroma Technology Corp, Bellows Falls, VT, USA | T505lpxr | longpass 505 nm |
| Image splitter | Cairn Research, Faversham, UK | OptoSplit II | |
| Image splitter filter cube: dichroic mirror | AHF Analysentechnik GmbH, Tübigen, Germany | 568LPXR | longpass 568 nm |
| Image splitter filter cube: emission filter 1 (GFP emission) | AHF Analysentechnik GmbH, Tübigen, Germany | 520/28 BrightLine HC | bandpass 520/28 nm |
| Image splitter filter cube: emission filter 2 (RFP emission) | AHF Analysentechnik GmbH, Tübigen, Germany | 630/75 ET Bandpass | bandpass 630/75 nm |
| Pacing inset | Warner Instruments, Hamden, CT, USA | RC-37FS |
Références
- Sinnecker, D., Laugwitz, K. L., Moretti, A. Induced pluripotent stem cell-derived cardiomyocytes for drug development and toxicity testing. Pharmacology & Therapeutics. 143 (2), 246-252 (2014).
- Goedel, A., My, I., Sinnecker, D., Moretti, A. Perspectives and Challenges of Pluripotent Stem Cells in Cardiac Arrhythmia Research. Current Cardiology Reports. 19 (3), 23(2017).
- Rocchetti, M., et al. Elucidating arrhythmogenic mechanisms of long-QT syndrome CALM1-F142L mutation in patient-specific induced pluripotent stem cell-derived cardiomyocytes. Cardiovascular Research. 113 (5), 531-541 (2017).
- Sinnecker, D., et al. Modeling long-QT syndromes with iPS cells. Journal of Cardiovascular Translational Research. 6 (1), 31-36 (2013).
- Talkhabi, M., Aghdami, N., Baharvand, H. Human cardiomyocyte generation from pluripotent stem cells: A state-of-art. Life Sciences. , 98-113 (2016).
- Ben-Ari, M., et al. Developmental changes in electrophysiological characteristics of human-induced pluripotent stem cell-derived cardiomyocytes. Heart Rhythm. 13 (12), 2379-2387 (2016).
- Den Hartogh, S. C., Passier, R. Concise Review: Fluorescent Reporters in Human Pluripotent Stem Cells: Contributions to Cardiac Differentiation and Their Applications in Cardiac Disease and Toxicity. Stem Cells. 34 (1), 13-26 (2016).
- Schweizer, P. A., et al. Subtype-specific differentiation of cardiac pacemaker cell clusters from human induced pluripotent stem cells. Stem Cell Research & Therapy. 8 (1), 229(2017).
- Moretti, A., et al. Patient-specific induced pluripotent stem-cell models for long-QT syndrome. The New England Journal of Medicine. 363 (15), 1397-1409 (2010).
- Chen, Z., et al. Subtype-specific promoter-driven action potential imaging for precise disease modelling and drug testing in hiPSC-derived cardiomyocytes. European Heart Journal. 38 (4), 292-301 (2017).
- Lam, A. J., et al. Improving FRET dynamic range with bright green and red fluorescent proteins. Nature Methods. 9 (10), 1005-1012 (2012).
- Chen, G., et al. Chemically defined conditions for human iPSC derivation and culture. Nature Methods. 8 (5), 424-429 (2011).
- Burridge, P. W., et al. Chemically defined generation of human cardiomyocytes. Nature Methods. 11 (8), 855-860 (2014).
- Bhattacharya, S., et al. High efficiency differentiation of human pluripotent stem cells to cardiomyocytes and characterization by flow cytometry. Journal of Visualized Experiments. (91), e52010(2014).
- Wang, X., McManus, M. Lentivirus production. Journal of Visualized Experiments. (32), e1499(2009).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- R Development Core Team. R: A language and environment for statistical computing. , R Foundation for Statistical Computing. Vienna, Austria. Available from: http://www.R-project.org (2008).
- Jung, C. B., et al. Dantrolene rescues arrhythmogenic RYR2 defect in a patient-specific stem cell model of catecholaminergic polymorphic ventricular tachycardia. EMBO Molecular Medicine. 4 (3), 180-191 (2012).
- Lemoine, M. D., et al. Human iPSC-derived cardiomyocytes cultured in 3D engineered heart tissue show physiological upstroke velocity and sodium current density. Scientific Reports. 7, 5464(2017).
- Dorn, T., et al. Direct nkx2-5 transcriptional repression of isl1 controls cardiomyocyte subtype identity. Stem Cells. 33 (4), 1113-1129 (2015).
- Kaestner, L., et al. Genetically Encoded Voltage Indicators in Circulation Research. International Journal of Molecular Sciences. 16 (9), 21626-21642 (2015).
Access restricted. Please log in or start a trial to view this content.
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.