
Method Article
Techniques de reproduction pour la surveillance et le contrôle de l'ovaire chez les amphibiens
Dans cet article
Résumé
L'étude de la biologie des amphibiens fournit des informations précieuses sur les processus reproducteurs, physiologiques, embryologiques et développementaux qui animent les organismes de nombreux groupes taxonomiques. Ici, nous présentons un guide complet sur les différentes méthodologies qui peuvent être utilisées pour étudier le contrôle et la surveillance de l'ovaire chez les amphibiens.
Résumé
Le contrôle et la surveillance ovariennes chez les amphibiens nécessitent une approche à multiples facettes. Il existe plusieurs applications qui peuvent induire avec succès des comportements reproducteurs et l'acquisition de gamètes et d'embryons pour la recherche physiologique ou moléculaire. Les amphibiens contribuent à un quart à un tiers de la recherche sur les vertébrés, et ils contribuent dans ce contexte à la connaissance de la communauté scientifique des processus reproducteurs et du développement embryonnaire. Cependant, la plupart de ces connaissances proviennent d'un petit nombre d'espèces. Ces derniers temps, la décimation des amphibiens à travers le monde a nécessité une intervention croissante de la part des écologistes. Les colonies de rétablissement et d'assurance en captivité qui continuent d'émerger en réponse au risque d'extinction rendent la recherche existante et les applications cliniques inestimables pour la survie et la reproduction des amphibiens détenus sous la protection de l'homme. Le succès de toute population captive est fondé sur sa santé et sa reproduction et sur la capacité de développer des descendants viables qui portent la représentation génétique la plus diversifiée de leur espèce. Pour les chercheurs et les vétérinaires, la capacité de surveiller et de contrôler le développement et la santé de l'ovaire est donc impérative. L'objectif de cet article est de mettre en évidence les différentes techniques de procréation assistée qui peuvent être utilisées pour surveiller et, le cas échéant ou nécessaire, contrôler la fonction ovarienne chez les amphibiens. Idéalement, tous les problèmes de reproduction et de santé devraient être réduits par l'élevage en captivité, mais, comme pour tout animal, les problèmes de santé et de pathologies reproductives sont inévitables. Les techniques non invasives comprennent les évaluations comportementales, l'inspection visuelle et la palpation et les mesures morphométriques pour le calcul des indices de l'état corporel et de l'échographie. Les techniques invasives comprennent les injections hormonales, l'échantillonnage sanguin et la chirurgie. Le contrôle de l'ovaire peut être exercé de plusieurs façons en fonction de l'application requise et des espèces d'intérêt.
Introduction
Les amphibiens sont depuis longtemps reconnus comme d'importants modèles biologiques et médicaux par un large éventail de disciplines de recherche. Les données obtenues en étudiant des espèces particulières telles que Xenopus laevis et X. tropicalis, la grenouille léopard (Lithobates (anciennement Rana) pipiens) et l'axolotl (Ambystoma mexicanum) ont été appliquées à un certain nombre d'autres espèces de vertébrés, y compris les humains. Les techniques vétérinaires, d'élevage et de procréation assistée qui ont émergé de l'étude de ces amphibiens et d'autres fournissent une assistance aux personnes chargées de développer des soins, un entretien et une durabilité réussis des populations plus rares en captivité. 1 Fois , 2 (en) , 3 (en) , 4.
L'intérêt gagne pour l'utilisation simultanée d'approches de conservation in et ex situ pour inverser la tendance de l'extinction pour de nombreuses espèces d'amphibiens en péril1,2. Cet article fournit les méthodologies actuellement disponibles pour surveiller et contrôler la fonction ovarienne d'amphibien dans les espèces modèles des Anurans et des Caudates. En outre, des techniques existantes pour traiter une pathologie reproductrice commune de la conservation d'oeuf est présentée.
Comme dans de nombreux groupes taxonomiques, le contrôle de l'ovaire amphibien implique une série d'interactions étroitement synchronisées entre l'environnement et la physiologie. La température et la photopériode (connues sous le nom de signaux immédiats) sont décodées par l'œil et le cerveau où elles sont rapidement converties en processus génétiques, hormonaux et circadiens (signaux ultimes)3,4. Les méthodes de surveillance et de contrôle de la fonction ovarienne couvertes dans cet article comprennent des techniques invasives et non invasives. Les exigences de recherche et d'enseignement du Comité institutionnel des soins et de l'utilisation des animaux (CCIU) définissent les techniques non invasives comme étant celles qui causeront peu ou pas de douleur physique ou de détresse mentale et qui ne nécessitent aucun médicament analgésique5. Ici, les techniques non invasives incluent l'inspection visuelle et la palpation, les observations comportementales, les évaluations morphométriques et l'échographie. Au contraire, les techniques de collecte de sang, d'administration hormonale et de chirurgie (ovaricectomie et prélèvement d'ovules retenus) sont classées comme invasives, car elles peuvent entraîner une certaine douleur ou inconfort et nécessiter une anesthésie ou une pharmacothérapie post-procédurale.
Les techniques de surveillance ovarienne non invasive peuvent être facilement intégrées à la routine quotidienne des soins pour la plupart des amphibiens captifs. Selon l'espèce, la gravidité ovarienne peut souvent être déterminée par simple inspection visuelle (grenouille de verre). Dans d'autres cas, la palpation peut indiquer si une femelle est gravid. Divers indices de l'état du corps (BCI) tels que le poids, la longueur urostyle museau (SUL), la longueur du museau (SVL) et l'indice de masse standard (SMI) sont disponibles pour prédire la présence ou l'absence d'œufs4,6,7, 8,9. Cependant, il faut faire attention à l'interprétation des résultats, car la plupart ne tiennent pas compte de l'âge, de la forme corporelle ou de la composition (p. ex., l'eau retenue par rapport à la masse ou la graisse ovarienne)6. Les diagnostics reproducteurs définitifs peuvent être obtenus par ultrasons avec des connaissances plus approfondies acquises en ce qui concerne le développement des œufs et la mise en scène du cycle ovarien4,7. L'échographie fournit également un moyen de confirmer et de surveiller les pathologies de reproduction et les conditions physiologiques associées4,8.
En plus de fournir des informations sur l'état de santé, l'échantillonnage de sang peut être utilisé pour mesurer les hormones reproductrices. Si le profilage hormonal est l'objectif final, il est important d'éviter les influences liées au stress qui peuvent confondre les données systémiques stéroïdes. Bien qu'il s'agit d'un outil de surveillance potentiellement puissant, il n'y a pas encore d'étude démontrant des réponses endocrinologiques innées à l'administration exogène d'hormones chez toutes les espèces d'amphibiens. Le sang peut être prélevé en toute sécurité sur plusieurs sites; chez les grenouilles cela comprend la veine abdominale ventrale, plexus lingual, veine fémorale et le cœur9,10. Dans Caudates, le sang est prélevé dans la veine ventrale de la queue. Le degré d'invasivité, la quantité de retenue requise, le besoin d'anesthésique, la délicatesse de l'organe ciblé et la taille de l'animal sont des facteurs à prendre en considération lors du choix d'une technique de collecte pour le patient amphibien. Cet article présentera la technique de collecte de sang de la veine maxillaire ou musculo-cutanée faciale desgrenouilles comme décrit à l'origine par Forzan et autres 9.
Le contrôle de l'ovaire est spécifique à l'espèce et, en tant que tel, les protocoles hormonaux doivent être testés et optimisés. Outre la saisonnalité et le milieu hormonal circulant associé, le contrôle ovarien peut également être étroitement lié à l'âge, au temps passé en captivité et à l'exposition à l'administration répétée d'hormones, pour laquelle il y a peu d'information dans la littérature11 , 12 Ans, états-unis , 13. La mise en œuvre d'hormonothérapies pour susciter des comportements reproducteurs, la production de gamètes, la maturation et l'oviposition est devenue une approche largement rapportée pour résoudre les problèmes de reproduction courants associés à la captivité4, 8,14,15,16. Étant donné que les mécanismes de contrôle de la reproduction chez les vertébrés sont fortement conservés, il existe un certain nombre d'hormones, de neuropeptides et de médicaments disponibles dans le commerce utilisés thérapeutiquement dans d'autres groupes taxonomiques qui peuvent également être utilisés de façon fiable dans un certain nombre de espèces d'amphibiens (tableau 1). L'hormone de libération de gonadotropine (GnRH) et la gonadotropine chorionique humaine (hCG) (ou variations de celui-ci, c.-à-d., PMSG et eCG)17,18, individuellement ou en combinaison, ont été employées intensivement dans les captifs d'amphibiens programmes d'élevage incluant : le boréal des Rocheuses méridionales (Anaxyrus boreas boreas)4,19,20; le crapaud, Grenouille Dusky Gopher, Rana sevosa (Langhorne et al., non publié)7; le chien d'eau de la côte du Golfe, Necturus beyeri20; Wyoming toad, Anaxyrus baxteri18; la grenouille-taureau, Rana catesbiana21; le toad américain, Anaxyrus americanus22; la grenouille d'herbe, Lymnodyaster tasmaniensis23; le Coqui, Eleutherodactylus coqui24; le Xenopus, Xenopus laevis25; le toadlet du Gunther, Pseduophryne guentheri26; la grenouille léopard du Nord, Lithobates pipiens; l'Argentin Horned-frog, Ceratophrys orné; la grenouille à cornes du Cranwell, C. cranwelli; la grenouille de terre américaine, Odontophrynus americanus27; et la salamandre de feu (Salamandra)228. Les hormones stéroïdes, comme la progestérone (P4), sont moins couramment rapportées, mais ont démontré une bonne efficacité dans l'obtention de l'ovulation et l'oviposition chez certaines espèces d'anurans16,18,29. Les prostaglandines (en particulier la prostaglandine 2-alpha (PGF2)sont impliquées dans l'ovulation avec les corticostéroïdes30,31,32,34 et atteignent des niveaux élevés pendant la phase ovulatoire31.
Dans des études in vitro, PGF2 est un puissant inducteur de l'ovulation31, tandis que in vivo, il peut induire l'oviposition d'ovules conservés dans Rana muscosa4,30,32. Les extraits pituitaires sont également des inducteurs efficaces de l'ovulation15,16,34; cependant, les préoccupations entourant la biosécurité et le potentiel de transmission de la maladie sont souvent dissuasives pour les colonies reproductrices captives lorsqu'on considère cette approche35.
La dernière section de cet article détaille les procédures chirurgicales et fournit des approches alternatives pour étendre les études ovariennes ou l'aide à la résolution des pathologies de reproduction. Les ovariectomies sont le plus souvent pratiquées chez les amphibiens pour obtenir des ovocytes pour la recherche embryologique. Cependant, il peut également fournir un remède pour les œufs conservés lorsque d'autres options échouent. Bien que cette procédure soit invasive, nécessitant une anesthésie complète et des incisions pour exposer les masses d'œufs, elle ne nécessite pas d'euthanasie. En outre, après une ovaricectomie partielle, les animaux peuvent faire un rétablissement complet et continuer à être actifs sur la reproduction après la chirurgie8,36.
Les protocoles décrits ci-dessous décrivent les méthodes invasives et non invasives de contrôle et de surveillance ovariennes à Anurans et Caudates. Les espèces spécifiques choisies pour illustrer les techniques à Anurans comprennent R. mucosa et X. laevis. Necturus maculosus, N. beyeri, N. alabamensis, et A. mexicanum constituent les espèces utilisées pour décrire de la même façon les techniques de Caudates.
Protocole
Les procédures de salamandre ont été approuvées par les protocoles 11-106, 13-110, 14-133 et 15-138 du Comité des soins et de l'utilisation institutionnels du zoo et du jardin botanique de Cincinnati (CZBG). Toutes les procédures de grenouille et de couple ont été approuvées par les protocoles du San Diego Zoo Global (SDZG), du Comité d'aide et d'utilisation en établissement (IACUC) : 15-001, 16-005 et 18-003.
Le soin et le traitement des animaux ont été approuvés par le Comité d'éthique du Muséum national d'histoire naturelle (Paris) (Musée National d'Histoire Naturelle-Ménagerie du Jardin des Plantes (MNHN)), conformément aux Principes directeurs institutionnels et nationaux (Commission de Génie Génétique, Direction Départementale des Services Vétérinaires, Directive de l'Union européenne 2010/63/UE, décision d'accord No. C75-05-01-2 pour la Convention européenne pour les animaux vertébrés utilisés à des fins expérimentales et scientifiques. Tous les protocoles utilisés dans cette étude ont été approuvés sous le numéro de référence 68-037.
1. Techniques de surveillance ovarienne non invasive
- Inspection visuelle et palpation
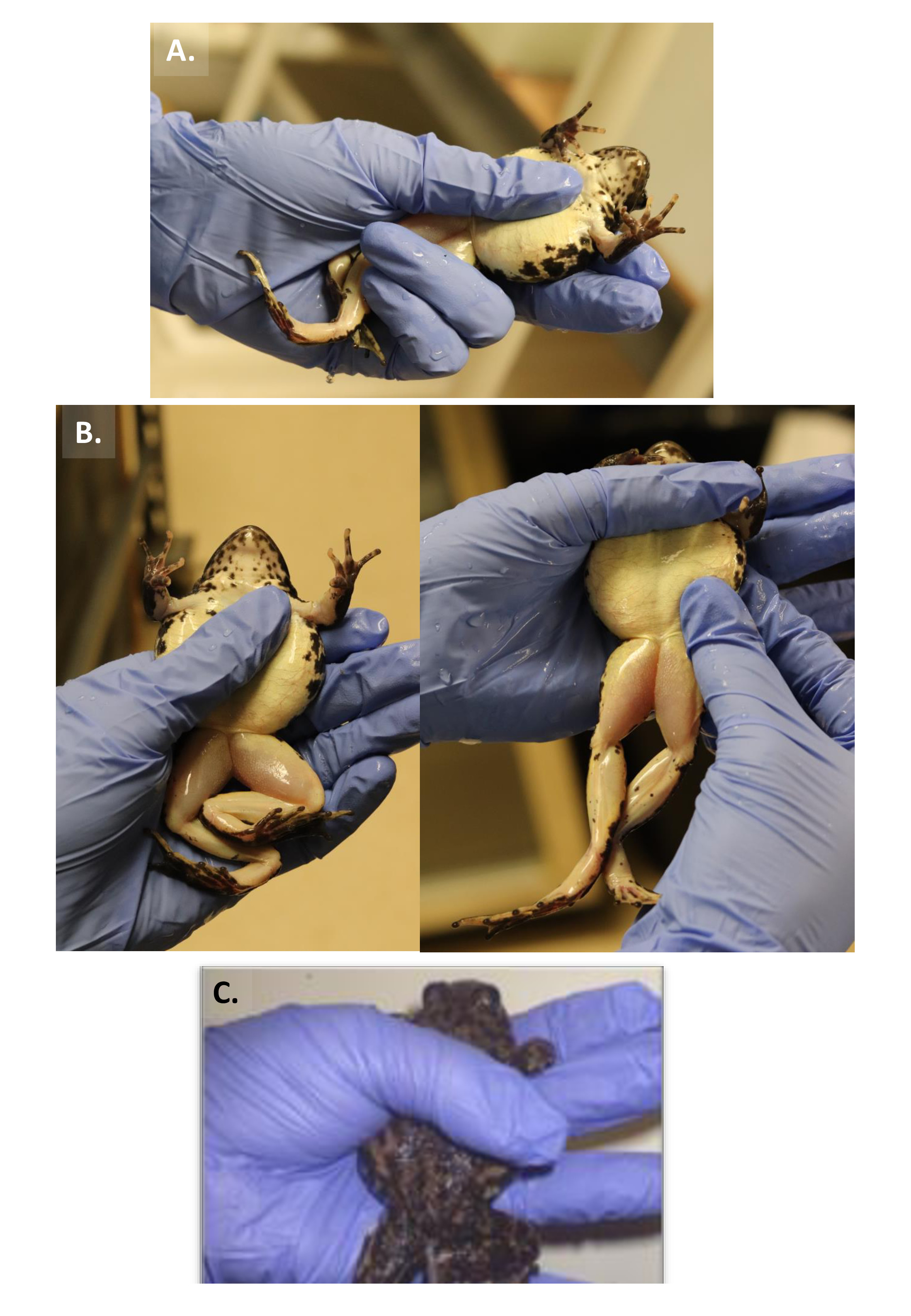
- Tenez la femelle Anuran dans l'une des trois façons décrites ci-dessous.
- Fixer les jambes de la grenouille ou du rebord avec l'anneau et le petit doigt, soutenant le côté dorsal (abdomen) du corps de la grenouille avec l'index et le majeur et le côté ventral avec le pouce (Figure 1A).
- Tenez la grenouille ou le rebord dans la main dominante avec un pouce sur son abdomen et le reste des doigts fixant le dos de l'animal. En utilisant la main non dominante pour palper l'abdomen de l'animal, sentez s'il y a une bosse subdermique (figure 1B).
- Reposez l'abdomen de la grenouille ou du rebord sur la paume de la main, les bras avant drapés sur l'index et un pouce sur le haut du dos.
- Étant donné que les Caudates sont de nature entièrement aquatique, effectuez une inspection visuelle par l'une des deux méthodes décrites ci-dessous.
- Déplacer l'animal dans un récipient séparé de 4 L contenant de l'eau de réservoir. Tenez le contenant (couvercle fixé) et brillez lampe de poche sur le dessous pour visualiser la présence / absence d'œufs.
- Anesthésie dans MS222 (0,5 g/L; Tricaine methanesulfonate, tamponné avec 0,5 M NaHCO3). Après l'induction, faire pivoter l'animal sur son dos et examiner l'abdomen.
- Tenez la femelle Anuran dans l'une des trois façons décrites ci-dessous.
- Évaluations morphométriques
- Anurans (Anurans)
REMARQUE : L'anesthésie n'est pas requise.- À l'aide d'étriers, mesurer l'animal du bout de la bouche, le long du centre du corps jusqu'à la pointe de la queue pour obtenir SUL et SVL (Figure 2A, B).
- Tare un récipient en plastique sur une échelle de précision numérique. Placer l'animal dans un contenant de marécage et peser (Figure 2C).
- Pour les grands animaux, comme les grenouilles-taureaux, ou lors de l'obtention de poids sur le terrain, utilisez une balance suspendue (Figure 2D).
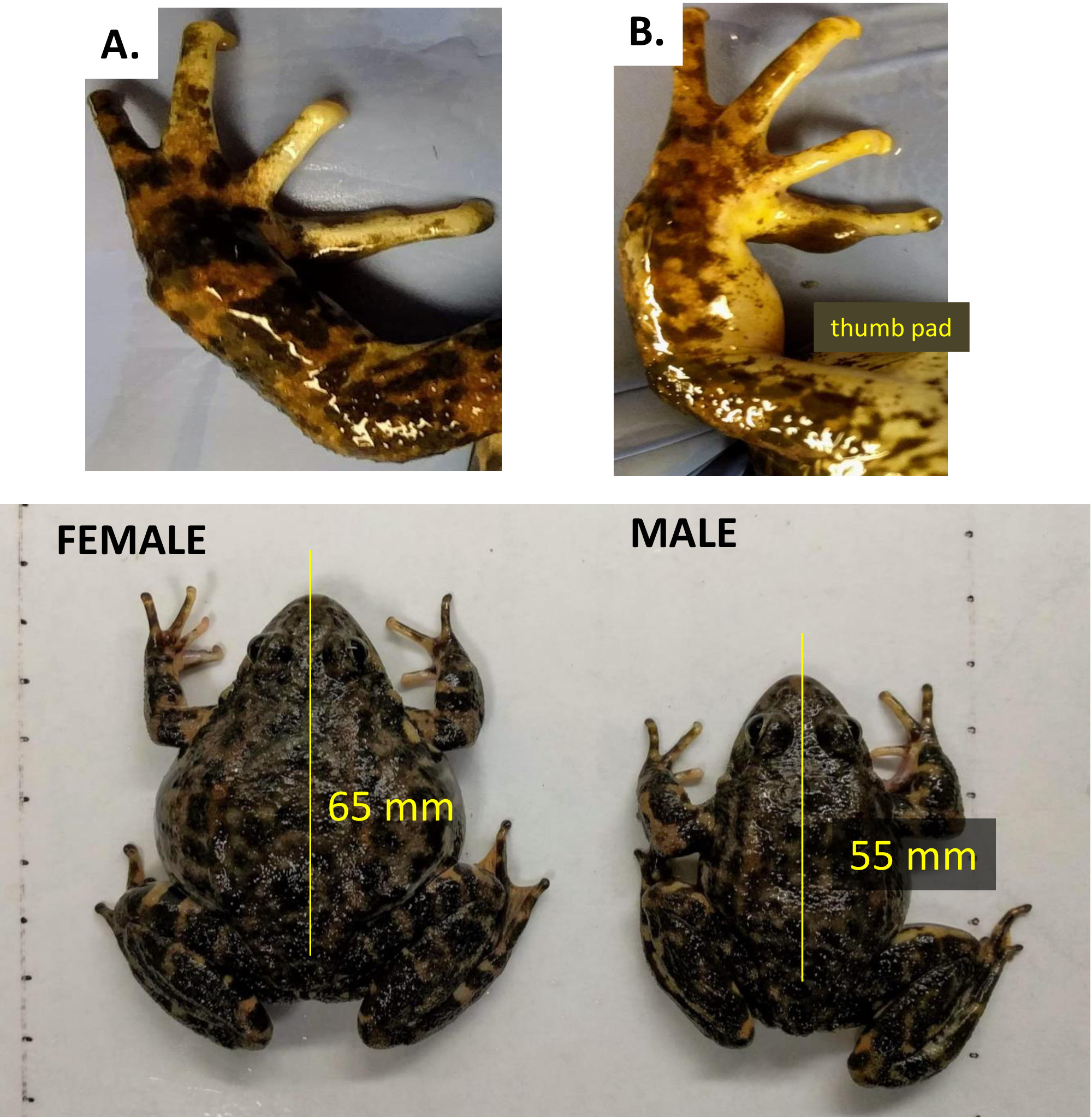
- Comme chez de nombreuses espèces d'anurans, distinguer les femelles adultes R. muscosa des mâles par leur plus grande taille et l'absence de tampons nuptiaux (pouce) sur les mains (Figure 3).
- Calculer l'état du corps, comme une évaluation de base de la santé globale par la formule suivante:
Indice de Fulton : K ' masse ' longueur3
REMARQUE : L'indice de Fulton utilise un équilibre dimensionnel du volume lié à la masse et à la longueur où 3 est l'exposant de mise à l'échelle qui relie la masse et la longueur isométriquement.
- Caudates (Caudates)
- Tare balance avec un sac vide avant de placer l'animal non anesthésié à l'intérieur. Veillez à ne pas introduire l'excès d'eau (figure2D) et agissez rapidement pendant que les animaux sécrètent du mucus comme réponse au stress pour être retenu.
- Obtenir des mesures pour adultes en immobilisant des individus en position droite au fond d'un sac en plastique refermable ou dans un contenant en plastique séparé pouvant accueillir des étriers expansifs.
- Mesurer la longueur du corps avec des étriers (Figure 2E).
- Mesurez le caudate de la pointe du museau à la pointe de la queue (SVL) pour surveiller la croissance.
- Anurans (Anurans)
- Observations comportementales
- Observez physiquement les animaux en temps réel ou utilisez une caméra vidéo pour enregistrer le comportement.
- Enregistrer les observations des animaux catégoriser les comportements et construire un éthogramme (figure 4).
- Classer les comportements reproducteurs
REMARQUE : La figure 4 illustre un type de comportement reproducteur observé chez les anurans.
- ultrason
REMARQUE: Le transducteur d'ultrasons de choix, dans ce cas, 7,5 mHz linéaire ou une multi-fréquence (10-6 mHz) micro-convexe, est recommandé pour Necturus et une sonde de 10 MHz et soluble dans l'eau, gel sans sel pour R. muscosa. L'échographie sur les salamandres peut nécessiter une anesthésie (voir la section 1.5 pour les instructions).- Anurans (Anurans)
- Effectuer l'échographie sur R. muscosa à l'aide de deux personnes (Figure 5A).
- Première personne: Tenir l'animal avec la main dominante et appliquer l'eau soluble, gel sans sel à l'abdomen de l'animal.
- Deuxième personne (ultra-échographe) : Prenez la sonde de 10 MHz dans la main dominante et appliquez-la sur l'abdomen en veillant à établir un bon contact entre la sonde et le gel.
- Glissez vers l'intérieur juste en dessous de la fosse du bras vers le centre de la ligne médiane abdominale de l'animal pour visualiser l'ovaire entier.
- Ultra-échographe : Utilisez la main non dominante pour geler le cadre et capturer les images désirées sur l'échographie.
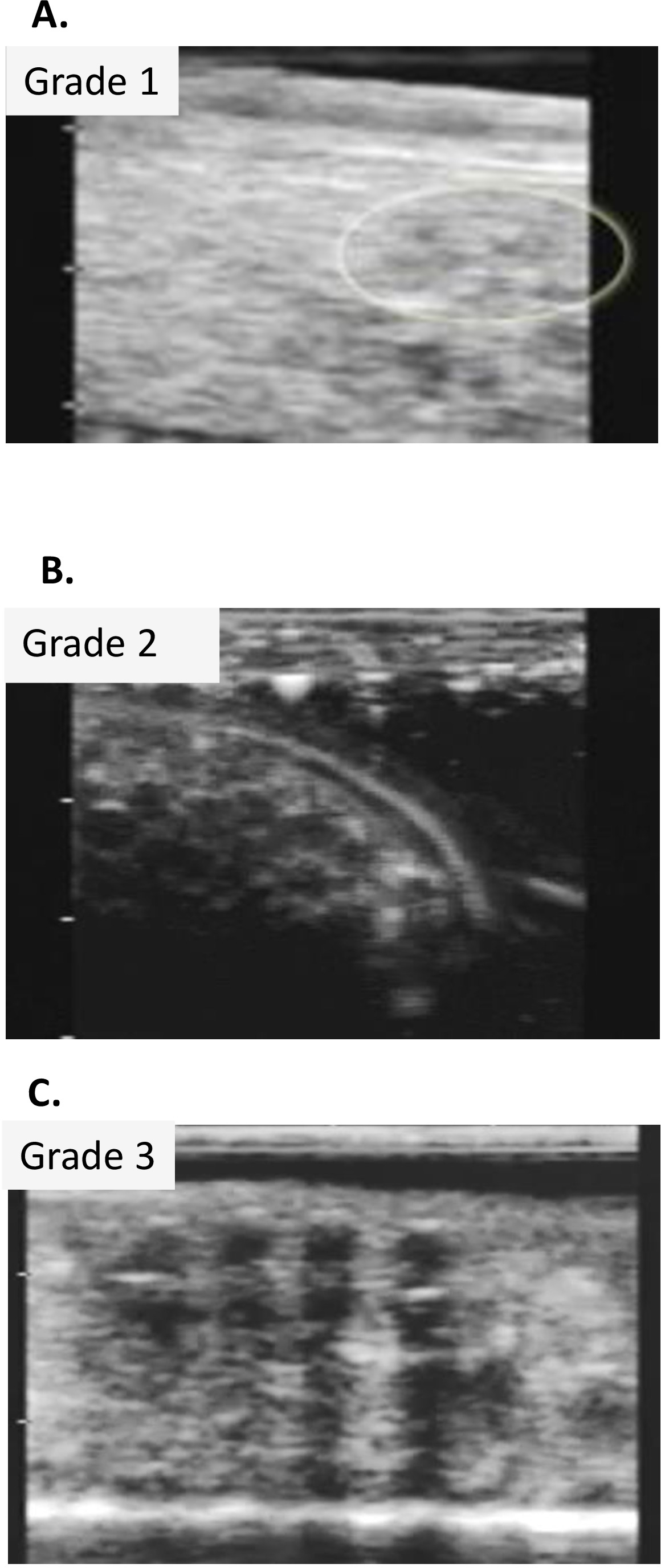
- Catégoriser l'étape du cycle ovarien par le système de classement établi pour le genre4 (tableau2, figure 5B-F).
- Rincer n'importe quel gel de l'animal à la fin de la procédure.
- Caudates (Caudates)
- Transférer le Necturus non anesthésié dans un récipient rectangulaire de 4 L rempli de 2 L d'eau de réservoir.
- Minimisez le mouvement des animaux en éteignant les lumières de la pièce et/ou en tassez une main sur la tête de l'animal.
- Placez le transducteur à une distance de 1-2 cm de la paroi du corps.
- Localiser le cœur au niveau ventral de la ligne médiane jusqu'aux membres antérieurs, puis déplacer le transducteur de loin et examiner le tissu ovarien7.
- Catégoriser les femelles selon le système de classement établi pour le genre4 (Figure 6A, B,C).
- Obtenir des mesures précises des œufs à un stade de mi-gravité tardif en capturant des images lorsque le corps de l'animal est à un angle du transducteur (c.-à-d. non linéaire, mais léger arc; Figure 6B). Sinon, les follicules qui se chevauchent font qu'il est difficile de différencier la taille des œufs individuels.
- Anurans (Anurans)
- Induction et récupération anesthésiques
- Anurans (Anurans)
- Anesthésie dans MS222 (0,5 g/L; Tricaine methanesulfonate tamponné (0,5 M NaHCO3) tel que décrit précédemment.
- Utilisez le réflexe de redressement comme indicateur principal de la mesure dans laquelle l'animal est devenu anesthésié. La perte complète de réflexe démontre un état d'anesthésie profonde.
- Retirez l'animal de l'anesthésie à base de bain d'eau (MS-222) une fois que le réflexe de redressement est perdu.
- Placer l'animal sur une serviette humide (avec de l'eau déchlorée sans anesthésique).
- Assurez-vous de garder l'animal humide pendant toute la procédure chirurgicale.
- Intuber les petits amphibiens avec des cathéters en caoutchouc rouge, des tubes non éraflés ou des tubes endotrachéales menottés classiques sans gonfler le brassard.
- Fournir un faible débit d'oxygène (0,5-0,75 L/min) avec 0,5-1% d'isoflurane.
- Arrêter l'isoflurane après la procédure, mais garder le flux d'oxygène pendant 1 minute.
- Extuber l'animal et rincer l'animal à fond avec de l'eau déchlorée anesthésique pendant 2 minutes.
- Mettre l'animal dans une quantité peu profonde d'eau déchlorée ou sur une serviette humide.
- Évaluer le rétablissement de l'animal en tirant doucement sur un membre postérieur pour s'allonger. Toute contraction de réponse du membre indique le réflexe de retrait.
- Surveillez d'autres indicateurs de récupération tels que les respirations gulaires (mouvement de la gorge) et le réflexe de redressement.
- Considérez l'amphibien récupéré quand tous les réflexes sont revenus, et les taux de coeur et de respiration sont retournés aux valeurs pré-anesthésiques.
- Caudates (Caudates)
- Anesthésiez necturus et Ambystoma dans MS222 (0,5 g/L de méthane sulfonate de Tricaine, tamponné avec 0,5 M NaHCO3, (MS222) dans un réservoir rectangulaire de 4 L.
- Placez une pierre d'air (1 pouce) et une pompe à air dans le réservoir et allumez-la à un flux constant pour fournir une oxygénation adéquate.
- Lorsque la fonction des membres et le réflexe de redressement sont perdus, retirez l'animal de l'anesthésie à base de bain d'eau (MS-222) et mettez l'animal sur une serviette humide (avec de l'eau déchlorée anesthésique).
- Maintenir l'humidité de la peau et des branchies à l'eau d'un réservoir.
- Pour récupérer l'animal, placez-le soigneusement côté ventral vers le bas dans un récipient en plastique de 4 L rempli de 2 L d'eau de réservoir avec une pierre d'air.
REMARQUE : La récupération commence par le clignotement des branchies, suivi de la capacité de déplacer sa queue et de propulser vers l'avant et enfin le mouvement fonctionnel des membres. - Retournez l'animal dans son réservoir d'origine et surveillez de près les 24 prochaines h.
REMARQUE: D'autres méthodes d'anesthésie pour les amphibiens existent et celles-ci sont décrites dans Wright et Whitaker8.
- Anurans (Anurans)
2. Techniques invasives de moniteur et de contrôle ovarien
REMARQUE : Cette procédure a été adaptée à partir de Forzon et coll.10.
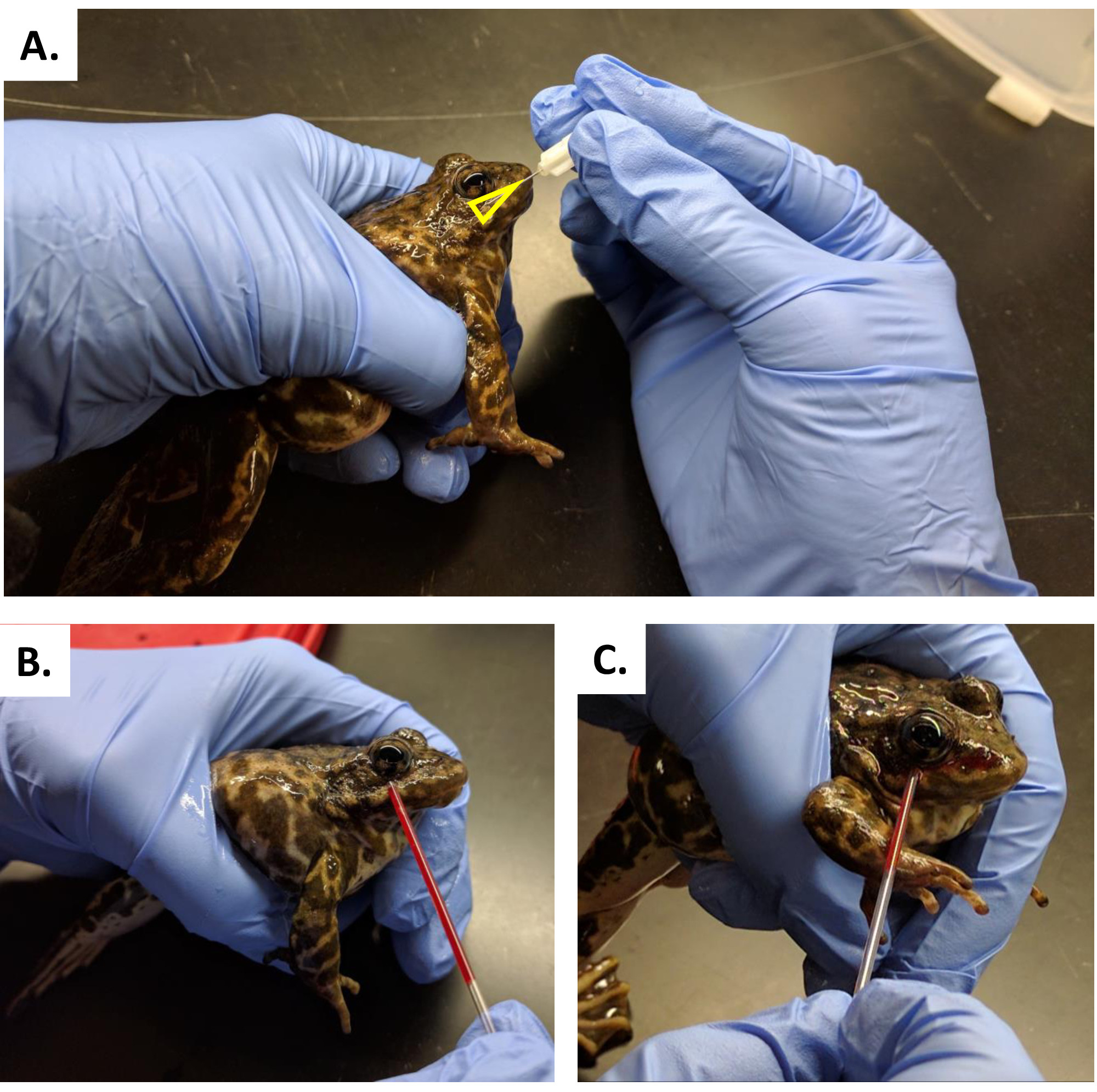
- Tenez la grenouille dans la main dominante, et tamponnez le côté venipuncture du visage de la grenouille avec une lingette stérile ou de la gaze.
- Séchez le visage pour éviter que le sang ne se disperse trop sur la peau.
- Insérer l'aiguille (26 G 1/2" et 27 G 1/2"), avec le bevel vers le haut, à travers la peau où la peau soulevée autour de l'œil et la crête de la mâchoire supérieure se rencontrent pour former le point d'un triangle (contour jaune) (Figure 7A) accédant à la veine facialis près la véna orbitalis postérieure.
- Perforer la veine faciale sous l'œil droit et au-dessus de la crête supérieure de la mâchoire, en commençant entre 1-2 mm en arrière de la ligne médiane de l'œil (Figure 7A).
REMARQUE : Pour les grenouilles plus petites (moins de 20 g), déplacez le point d'insertion plus près d'une position directement en dessous de la ligne médiane de l'œil. - Angle le tube microhematocrit vers le bas pour permettre à la gravité d'aider le sang à circuler dans le tube. Le sang doit couler immédiatement après la ponction (Figure 7B,C).
- Au premier signe de flux sanguin, placez la pointe du tube microhematocrit au site de perforation et de recueillir 1-2 tubes microhématocrit complets de sang et placez les tubes dans des réceptacles appropriés pour la collecte (Figure 7B, C).
- Si le sang ne coule pas facilement, ou si le volume est très faible, déplacez légèrement l'insertion de l'aiguille ou insérez l'aiguille de l'autre côté du visage.
- Arrêtez le saignement en appuyant fermement sur la gaze jusqu'au site de la ponction pendant au moins 20 s.
- Laissez la grenouille hors de l'eau pendant 10 minutes pour confirmer que le site de ponction ne rouvre pas.
- Utilisez une nouvelle aiguille et de nouveaux tubes microhématocrit pour chaque grenouille échantillonnée.
3. Induction d'hormone
- Préparation hormonale
- Préparer les injections d'hormones immédiatement avant l'utilisation pour assurer un effet maximal.
- Sélectionnez une hormone dans la sélection indiquée dans le tableau 1.
- Déterminer la concentration de l'hormone à injecter à l'aide d'un l ou d'un mL/g de poids corporel16.
- Diluer l'hormone dans l'un des éléments suivants : eau, phosphate tamponné salin (PBS), solution salin amphibienringer (SRAS) ou salin.
- Ne dépassez pas un volume d'injection de 200 l pour les grenouilles pesant 30-70 g et 300 l pour les grenouilles pesant 80-110 g (observation personnelle)16.
- Pour la détention correcte d'un animal pendant l'administration hormonale de tout animal allant de 10-100 g, utilisez l'une des méthodes appropriées pour la tenue décrite à la section 1.1.
- Anurans (Anurans)
- Calculez la concentration requise par individu à l'aide d'un gramme par calcul du poids corporel (g/poids corporel).
- Juste avant l'administration, reconstituer dans un diluant stérile de choix.
- Assurez-vous qu'il n'y a pas de bulles dans la seringue avant l'injection.
- Tenez l'animal solidement dans la main non dominante et administrez l'injection avec la main dominante.
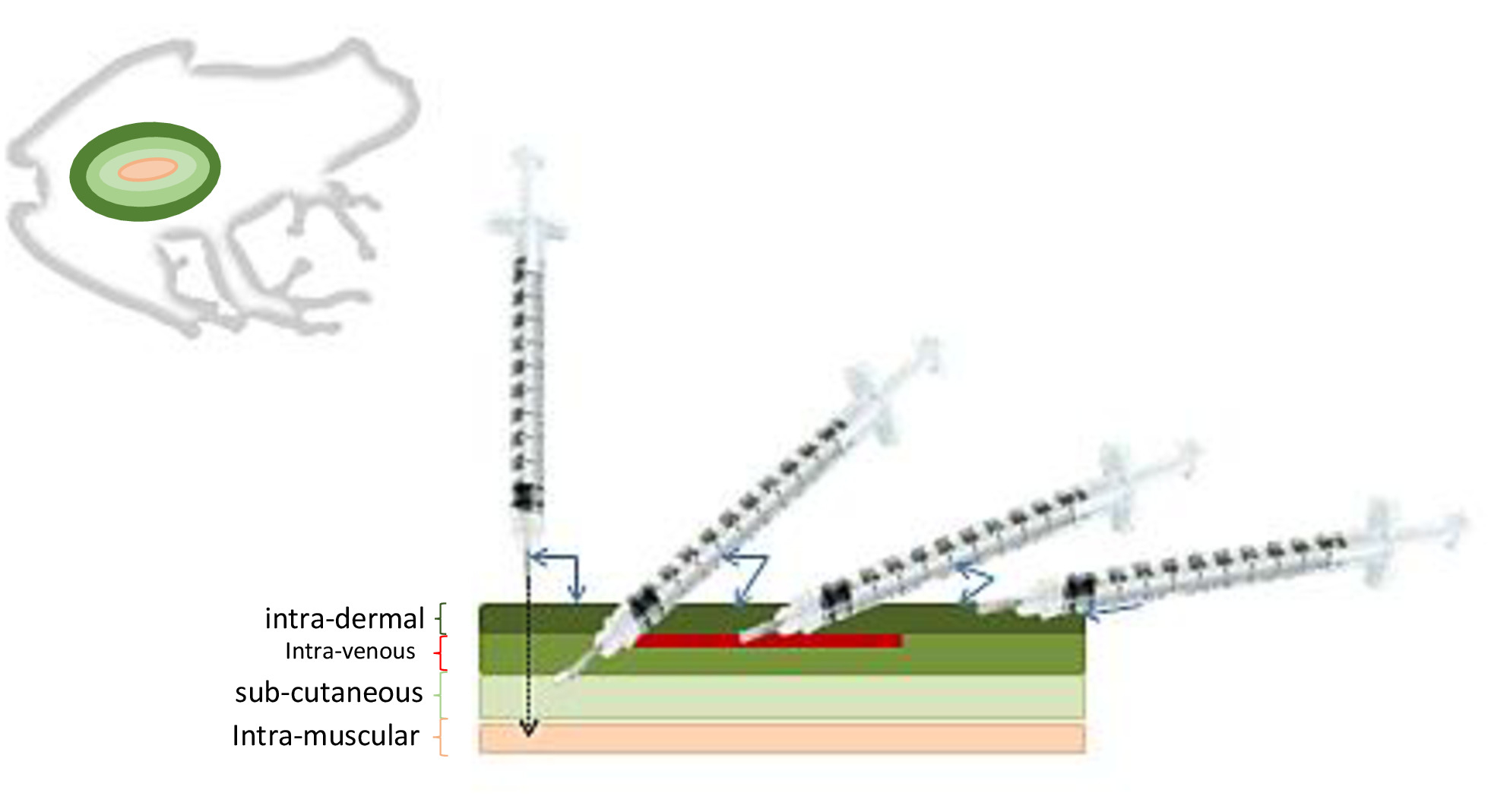
- Administrer l'injection selon les spécifications hormonales. Les injections les plus courantes chez les anuriens sont sous-cutanées, intra péritonéales ou intra musculaires (figure 8).
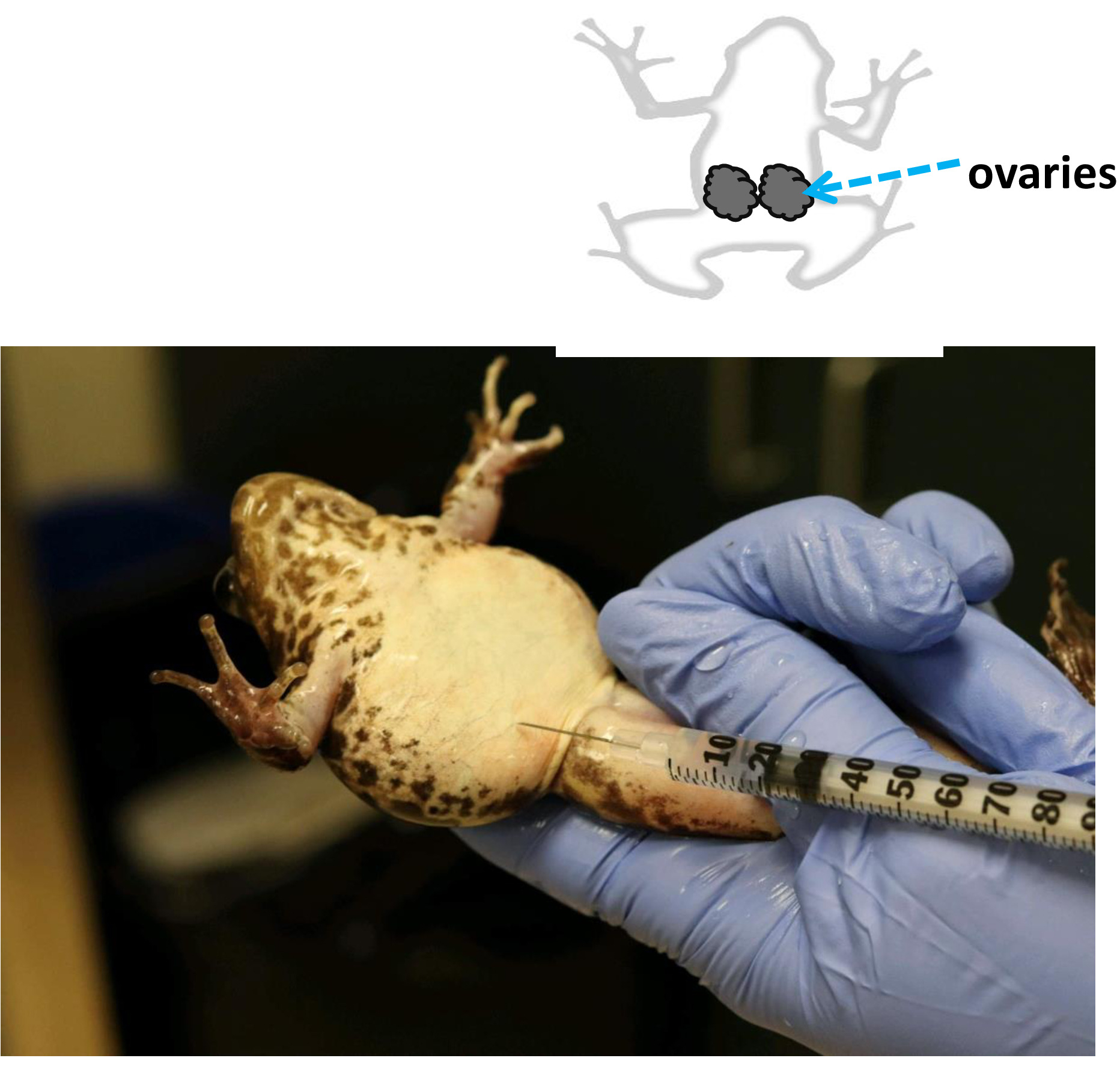
- Administrer des injections de propriété intellectuelle dans la partie inférieure de l'abdomen ou dans la partie inférieure du côté dorsal du corps près de la jambe arrière (figure 9).
- Administrer des injections intramusculaires de préférence dans les pattes postérieures.
- Caudates (Necturus)
- Reconstituer l'hormone de choix dans l'eau stérile selon le gramme par méthode de poids corporel décrit ci-dessus.
- Dans le cas de Necturus, utiliser des doses de 1,7-2,3 g de poids corporel GnRH/g.
- Retirer Necturus de la chambre anesthésique et le placer sur une surface de 45 degrés recouverte d'un drapé chirurgical.
- Placez l'animal avec la tête pointant vers le bas.
- Approchez le quadrant postérieur de l'abdomen (caudal de la jambe arrière) à un angle de 15-20 degrés. Veillez à ne pas introduire d'air dans la seringue.
- Injecter (IP) à l'aide d'une seringue à insuline et d'une aiguille de 27 à 30 G.
- Injectez l'hormone à l'aide d'une seringue à insuline et d'une aiguille de 27 à 30 G.
4. Chirurgie
- Préparation et procédure chirurgicales générales
- Pour maintenir des procédures aseptiques, employer les rideaux stériles clairs de plastique pour isoler le site chirurgical. Réduire l'évaporation en gardant la peau environnante humide.
- Humidifiez tous les matériaux qui entreront en contact avec la peau de l'animal avec de l'eau stérile. Faire l'incision de la peau avec un numéro 15 ou numéro 11 lame de scalpel.
REMARQUE : Combinaison d'acier froid, de radiochirurgie ou de laser à diodes. L'hémostase dans la procédure hémorragique douce peut être réalisée par laser électrocautérisme ou diode. - Utilisez des lances ou des applicateurs à pointe de coton pour permettre l'application d'une pression localisée à de petits vaisseaux qui suivent la perte de sang.
- Utilisez des lances ou des applicateurs à pointe de coton pour gérer de petits espaces confinés au lieu de carrés de gaze standard.
- Utilisez des micro-instruments, comme des instruments ophtalmologiques, avec de petits bouts fins, lors de la chirurgie sur les animaux pesant moins de 1 kg.
- Utilisez des rétracteurs en plastique qui s'auto-retiennent (p. ex., rétracteur lone Star) pour s'adapter à différentes tailles d'incisions.
- Utilisez des rétracteurs de paupière pour rétracter des incisions coelomic.
- Utilisez l'instrumentation de grossissement si nécessaire pour effectuer la chirurgie sur de plus petits patients.
REMARQUE: L'analgésie est nécessaire avec toute intervention chirurgicale chez les amphibiens. Le défaut d'administrer l'analgésie proportionnée pendant la chirurgie a été associé au retour retardé des fonctions normales. De plus, l'analgésie potentialise les effets des médicaments anesthésiques (tableau 3)34.
- Anurans (Anurans)
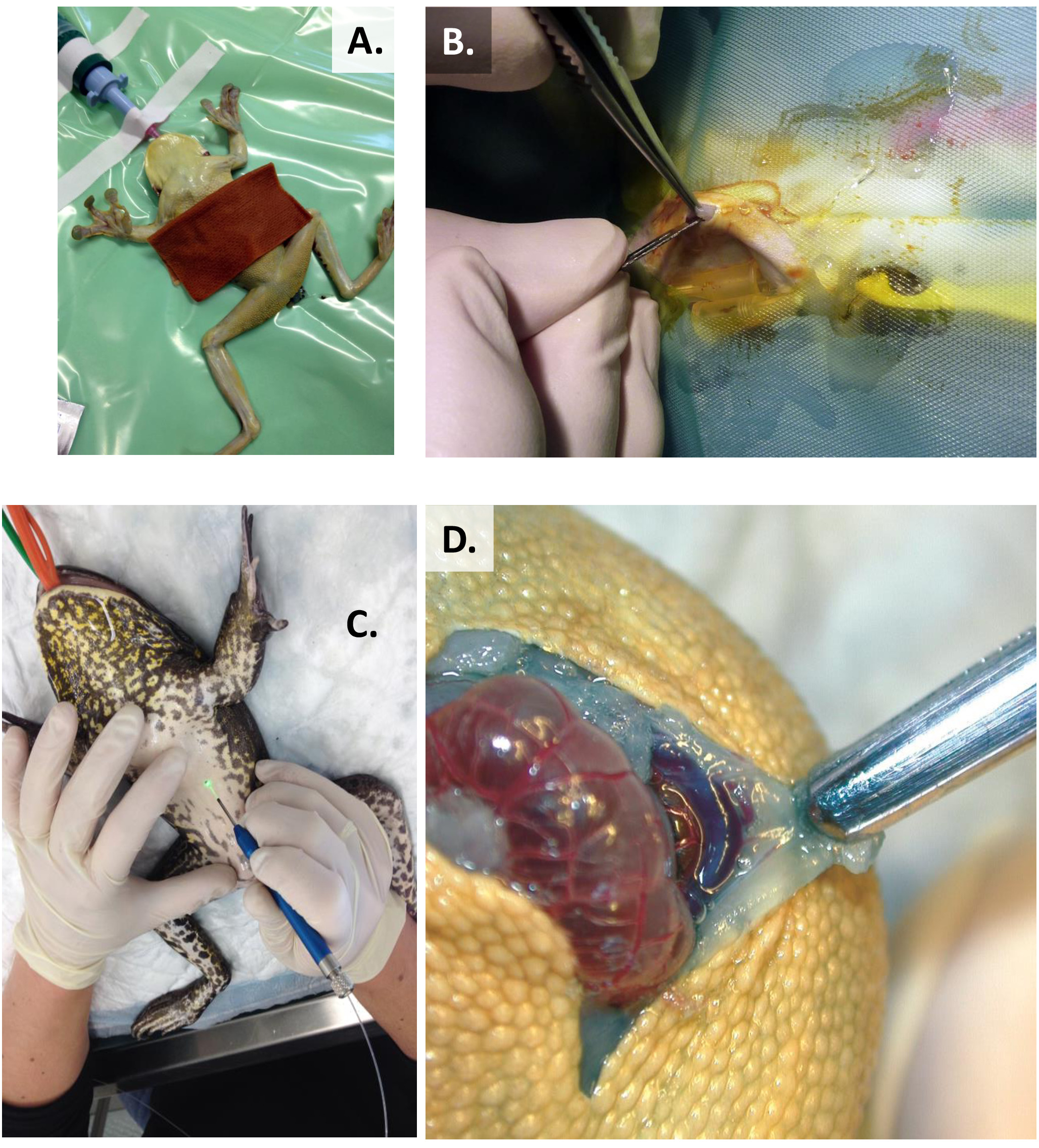
- Une fois que X. laevis a été anesthésié tel que décrit à l'étape 1.5.1, positionner l'animal dans la charge dorsale (Figure 10A,C).
- Préparer le champ chirurgical aseptiquement en essuyant la gaze stérile humide avec la solution povidone-iode diluée (1/10) sur le site pour 10-15 s ou 0.75% solution de chlorhexidine sur le site chirurgical pendant au moins 10 min avant la chirurgie35.
- Faire une incision de peau paramédiande de 3 mm dans le coelom moyen (entre les épaules et le cloaca) avec un coup gras laissant une incision propre utilisant un scalpel no 15 ou no 11.
REMARQUE: On peut utiliser un laser à diodes aussi pour les incisions cutanées. - Élever la membrane abdominale, faire et l'incision et disséquer soigneusement à l'aide d'un scalpel no 15 ou no 11. (Figure 10B,D).
- Retirez les incisions coélomiques avec des rétracteurs de paupières (ou tout équipement approprié).
- Accise une partie de la masse d'oeufs sans ligature retunlant les vaisseaux sanguins.
- Pour une ovarice complète, cautérisez les vaisseaux sanguins environnants par électrocautérisation ou diode laser (figure 11).
- À l'aide de la suture monofilament, fermez l'incision de céliotomie avec un modèle interrompu et de suture d'everting.
- Caudates (Caudates)
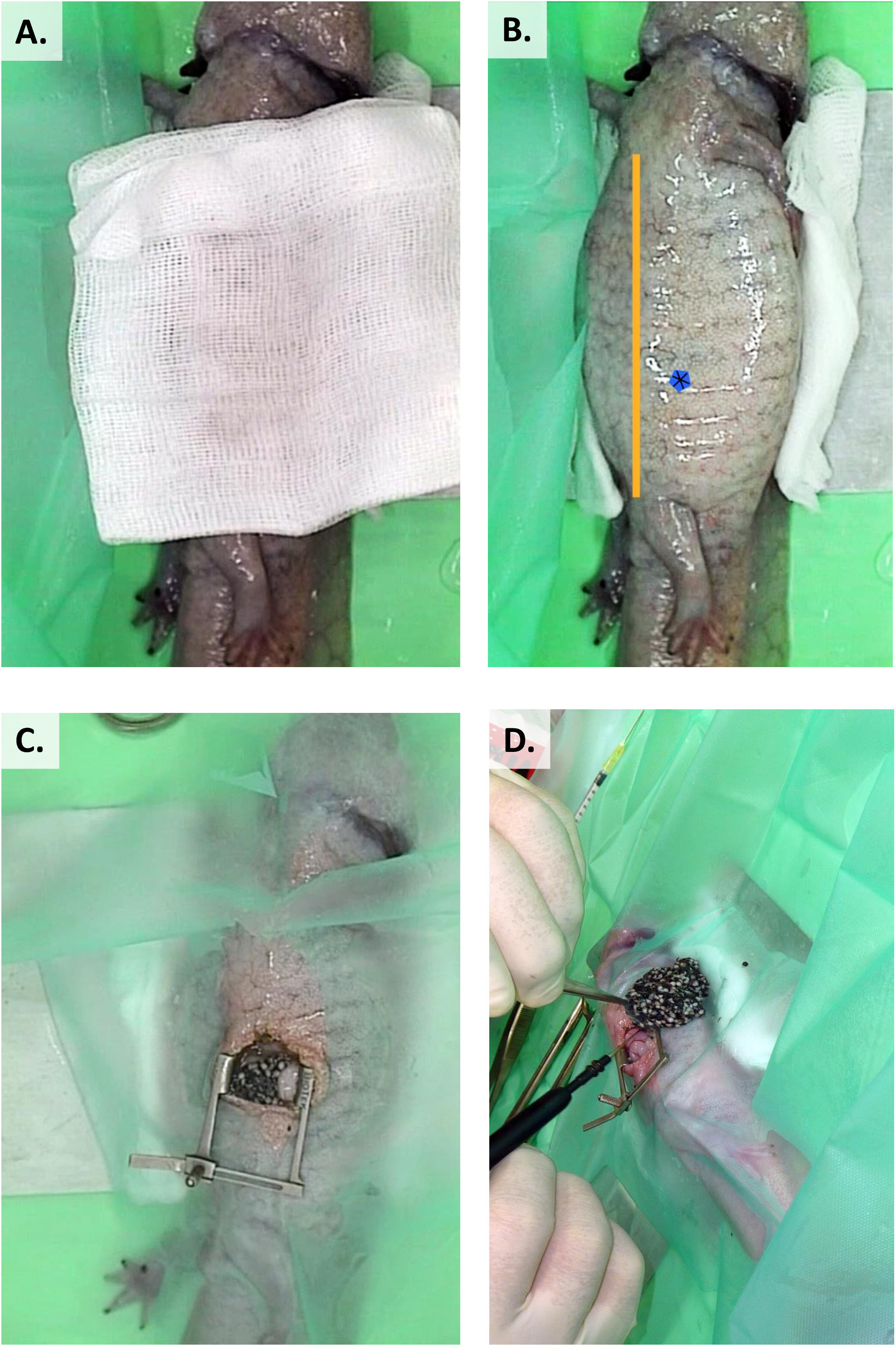
- Une fois A. mexicanum a été anesthésié, placez-le dans le recumbency latéral droit, avec le membre pelvien gauche simplement placé contre la base de la queue.
- Préparer le champ chirurgical aseptiquement en plaçant la gaze stérile humide avec la solution povidone-iode diluée (1:10) sur le site pendant 10-15 s. Alternativement, employer la gaze stérile imbibée dans la solution de chlorhexidine de 0.75% et placer sur le site chirurgical pendant au moins 10 min avant la chirurgie (Figure 12A)36,37 .
- Tracer une ligne entre l'épaule et les membres postérieurs pour diviser le corps en trois parties égales (figure 12B).
- Faire le site d'incision entre les deuxième et troisième parties.
- Saisissez le muscle sous-jacent et éloignez-vous des viscères coelomic.
- Forcer doucement les petits hemostats à travers la musculature coelomic et dans la cavité coelomic.
- Rétracter les incisions coélomiques avec des rétracteurs de paupières (ou tout autre matériau approprié) (figure 12C).
- Pour l'ovaricectomie complète, cautérisez les vaisseaux sanguins environnants par électrocautérisation ou diode laser (figure 12D).
- À l'aide de la suture monofilament, fermez l'incision de céliotomie avec un modèle interrompu et de suture d'everting.
Résultats
Morphométrie et reproduction
La visualisation de l'état reproducteur féminin chez les amphibiens varie selon l'espèce. La méthode la plus efficace est l'échographie; cependant, certaines espèces peuvent faire preuve de divers degrés de transparence de leur peau (figure 13A, B,C). L'inspection visuelle peut souvent illustrer clairement les différences entre une femelle gravid et non-gravid quand la peau est semi-translucide comme observé dans N. alabamensis et N. maculosus (figure 13A, B); ou translucide comme l'illustre la grenouille de verre (Figure 13C). La coloration de la peau tachetée foncée sur l'abdomen de N. beyeri interdit cette évaluation. Dans R. muscosa, la peau n'est pas translucide, mais des différences notables peuvent être détectées entre les femelles qui sont gravid par rapport à ceux qui ont récemment oviposited parce que la peau est flasque, et l'animal semble plus mince (ligne jaune) par rapport à un femelle gravid (ligne bleue) (Figure 13D). Avec l'expérience le gestionnaire peut se familiariser avec la différence entre une grande femelle et une gravid mais la confirmation de l'étape gravid exigera l'ultrason. Les indices de masse corporelle chez les amphibiens peuvent être calculés à l'aide d'un certain nombre de formules, mais leur application comme outil prédictif de reproduction est discutable. Dans le cas de R. muscosa, corrélation entre l'indice de Fulton, la santé et l'état reproducteur reste incertain.
Comportement reproducteur et échographie
Nos résultats montrent comment caractériser les comportements reproducteurs chez R. muscosa pour la prédiction de l'oviposition (Figure 4). Plusieurs étapes de quelques heures à plusieurs semaines comprennent, courtisant où un mâle poursuit activement une femelle (Figure 4A), le mâle monte et s'accroche fermement sur le dos de la femelle, appelé amplexus (Figure 4B). Une fois amplexed, la paire peut rester dans l'amplexus pendant 1 à 5 semaines et la paire affichera d'autres comportements en plus de l'amplexus. Amplexus est un comportement très actif qui comprend le mâle serrant la femelle d'une manière de pompage doux (Figure 4C); la femelle se déplaçant et commençant à afficher des comportements de main-stand par intermittence (figure 4D,E); et plus près du moment de l'oviposition, la femelle, dans un support de main, se penchera vers le haut contre des surfaces qu'elle peut coller des oeufs dessus tandis que le mâle pompe son abdomen vigoureusement (dans ce cas il est possible d'observer également la femelle frottant son abdomen vers le bas de dessous ses fosses de bras vers le cloaque. Il peut s'agit peut-être d'une façon mécanique de pousser les œufs vers le bas des oviductes) (Figure 4F, G).
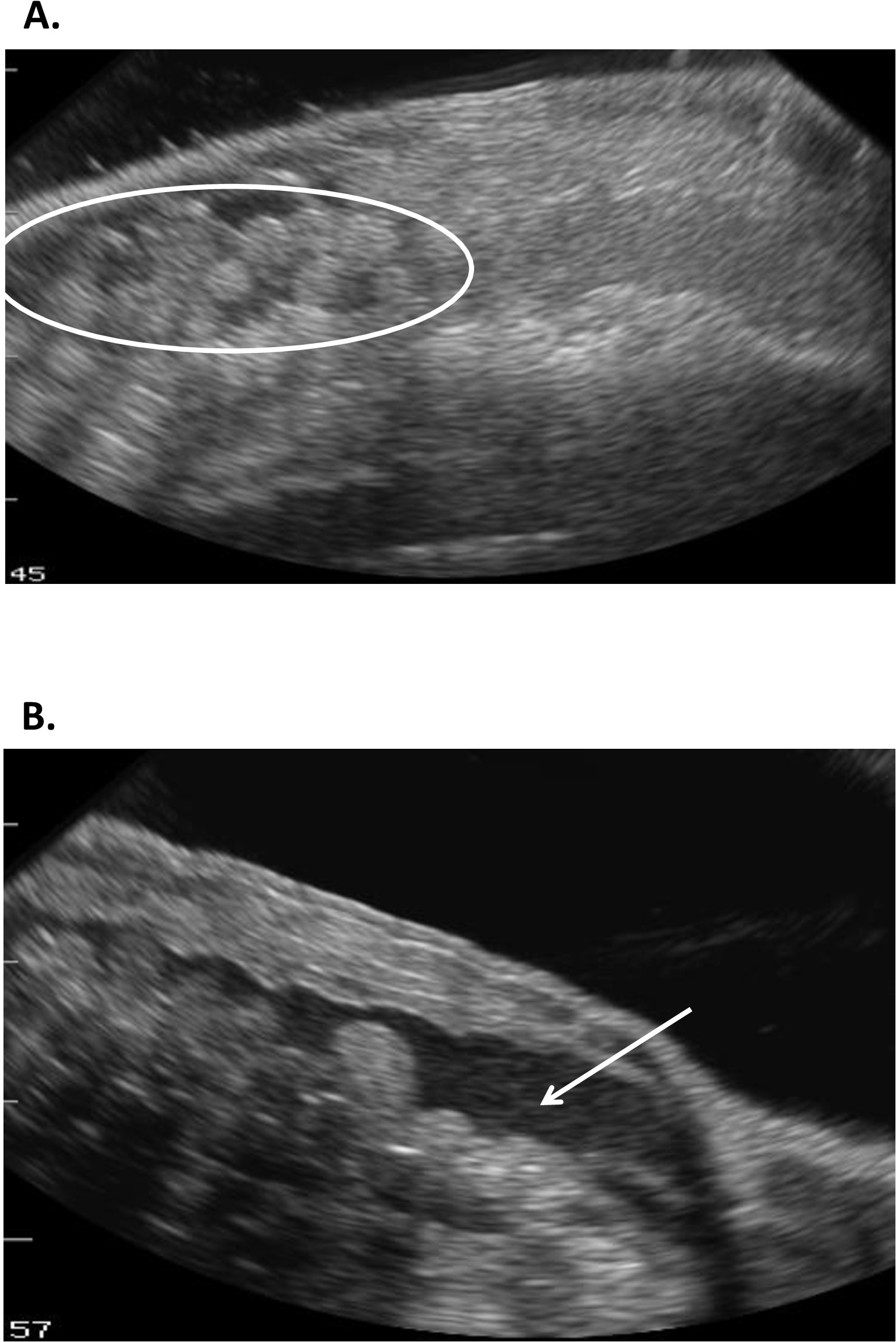
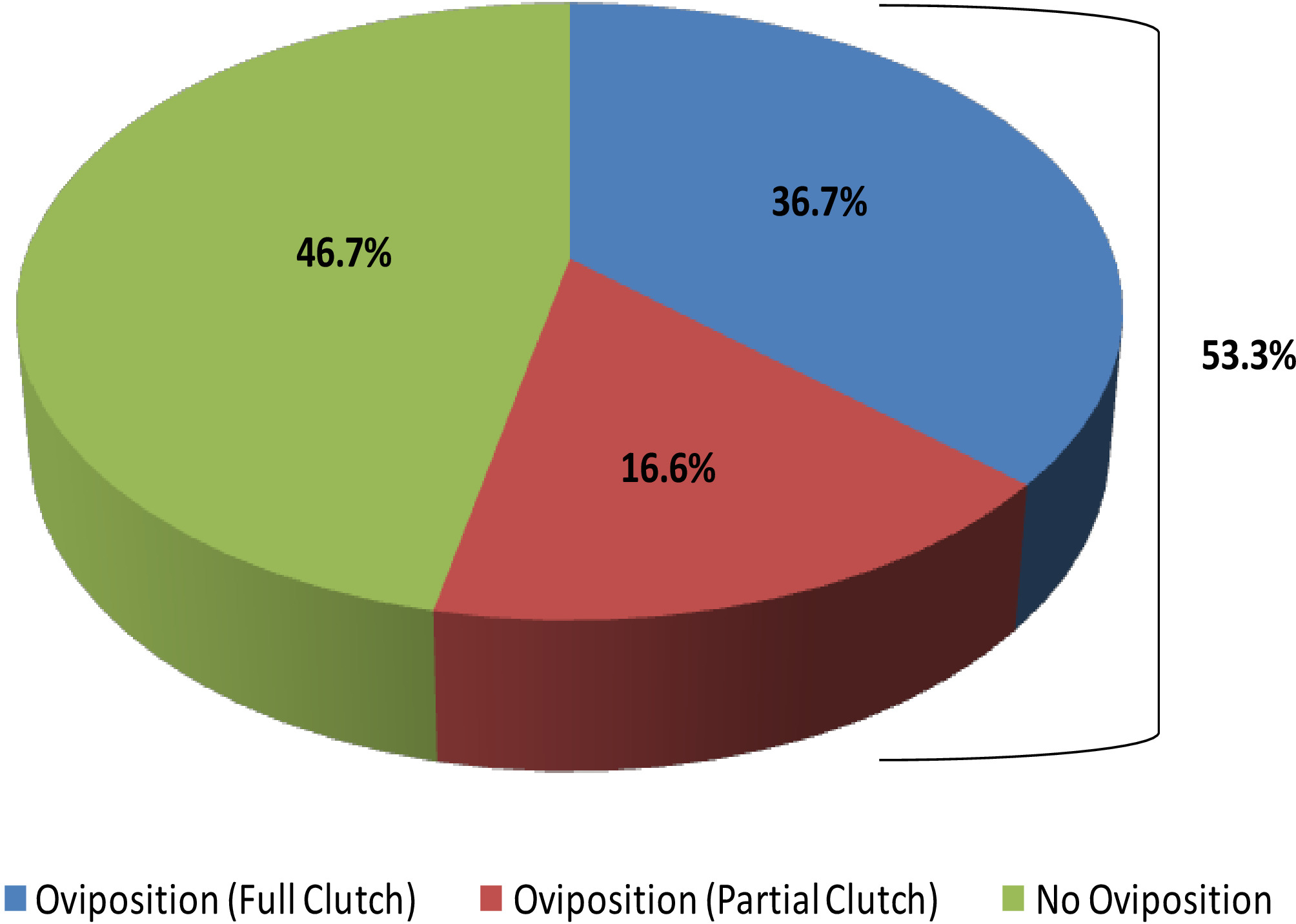
Cette étude illustre comment l'échographie peut fournir des informations avec lesquelles déterminer l'état reproducteur chez les femelles R. muscosa et Necturus. Quatre étapes de développement sont représentées dans R. muscosa (Figure 5C,D,E,F) et sont caractérisées de la même façon dans Necturus4 (Figure 6A,B,C). De plus, les œufs résiduels peuvent ne pas être expulsés, ce qui entraîne la rétention des œufs (Figure 5G, Figure 15A,B). L'étape 1 montre un ovaire directement après l'oviposition où les follicules sont difficiles à visualiser (Figure 5C). L'étape 2 est représentée par l'apparition de points échogéniques (taches blanches) dispersés dans tout l'ovaire (Figure 5D). Les stades 2 et 3 sont représentés par des points échogéniques plus grands et arrondis avec des centres sombres qui représentent des follicules de milieu à grand (figure5E,F). De 2013 à 2017, les necturus femelles captives ont été examinés par échographie sur une base mensuelle. Au cours de chaque examen, les individus ont obtenu une note en fonction des critères de reproduction établis pour le genre (tableau 2). Le pourcentage de femelles qui développent de nouveaux œufs chaque année s'est élevé en moyenne à 88,2 % (tableau5). Bien que le développement des œufs ait été élevé, l'oviposition n'a pas été assurée (figure 16). La majorité des femelles qui ont subi l'oviposition ont déposé le complément complet des oeufs, alors que quelques individus ont déposé seulement une fraction des oeufs qui se sont développés. Ces femelles R. muscosa et Necturus avec des œufs retenus concomitants avec le gain de fluide dans la cavité du corps ont été extérieurement visuellement agrandies avec des taches rouges sur la peau compatibles avec les vaisseaux sanguins éclatés (Figure 14A, B ) . Le degré de rétention d'eau pourrait être évalué davantage par échographie (figure15B). Chez les deux espèces, les œufs retenus ont subi une atrésie ou ont pris une apparence plus échogénique (figure14C, D, figure 15A).
Administration hormonale
Selon la profondeur du type d'injection, l'angle et la profondeur de l'aiguille varient. Pour la plupart des injections, la profondeur de l'aiguille n'a pas besoin d'être supérieure à 1-2 mm de profondeur lorsqu'on travaille avec des espèces comme R. muscosa, mais elle varie dans l'angle de pénétration. Les injections de prostaglandine ont nécessité une insertion intramusculaire (im) d'aiguille s'est inclinée à 90 degrés, dans la patte postérieure de R. muscosa, tandis que des injections intrapéritonéales (ip), avec une profondeur similaire à des injections intramusculaires, ont été administrées dans la région de la cavité coelomic à un 45 ' (Figure 10). L'administration d'Amphiplex n'a eu aucun effet significatif sur l'augmentation du nombre d'œufs déposés par les femelles traitées aux hormones par rapport aux témoins (P - 0,547), et il n'y a eu aucune différence dans le nombre d'embryons qui se sont clivés (P - 0,673) ou qui ont survécu au têtard (P 0.629) (Tableau 4). En général, le pourcentage d'ovipositing féminins est passé de 80 % en 2011 à 28 % en 2014. Le nombre de femelles ovipositing en 2015 a été significativement plus élevé qu'en 2013 (P - 0,0002), 2013 (P - 0,0001) et 2014 (P - 0,0026) mais pas 2011 (P - 0,0885), réaffirmant l'idée que les femelles de cette espèce ne peuvent pas se reproduire chaque année et que les protocoles hormonaux nécessitent nement. Pour les femelles De. muscosa présentant des signes de rétention d'ovules, les injections intramusculaires de PGF2 ont eu un taux de réussite de 60 % pour induire l'expulsion des œufs dégénérés. Cependant, dans 1 des 5 femelles injectées, le PGF2 n'était pas suffisant pour provoquer l'expulsion complète et certains œufs sont restés à l'intérieur de la femelle jusqu'à la saison de reproduction suivante. Dix-sept femelles de Necturus ont reçu LHRH/(GnRH) et 13 ont reçu une injection simulée d'eau stérile pour servir de contrôle (tableau 5). Au total, sept necturus femelles (n 4 alabamensis, n ' 2 beyeri, n ' 1 maculosus) ont continué à oviposit onze embrayages complets qui ont été attribués à la fois GnRH traités (n - 6) et le contrôle (n - 5) individus. Trois femelles (n 2 beyeri, n ' 1 maculosus) ont oviposited cinq embrayages partiels (figure 13). Ce phénomène ne semble pas être associé au traitement hormonal exogène puisque trois femelles témoins ont également déposé des couvées partielles (tableau 5). L'oviposition s'est produite sur une période de 37 jours (3/31-5/7) sur une période de cinq ans (tableau5). Il n'y avait aucune différence (P - 0,194) dans les taux d'oviposition entre les taux d'oviposition entre le traitement LHRH/GnRH (41 - 13,08%, gamme 17-67%) et le contrôle (66,75 à 11,79 %, fourchette de 50 à 100 %) Femelles. Les femelles traitées par LHRH/GnRH ont déposé en moyenne des ovules de 7,44 à 1,41 (gamme 3-13) jours après l'injection. Étant donné la nature entièrement aquatique de l'espèce et l'incapacité de retenir manuellement sans anesthésie, il était nécessaire d'assurer un niveau approprié de sédation avant d'effectuer des injections d'hormones IP (voir la section 3.2 pour les instructions sur l'anesthésie).
Collecte de sang, anesthésie et chirurgie
La technique d'échantillonnage de sang dans cet article a été prise de Forzan et autres 201310 et s'est avérée un moyen efficace de recueillir le sang de R. muscosa avec l'invasivité et le stress minimaux. À l'aide de tubes microhématocrits, environ 35 à 45 degrés de plasma ou de sérum peuvent être recueillis par 70 l de sang entier (figure7). Le volume de collecte maximal en R. muscosa était de 1 tube microhematocrit complet par 10 g de grenouille, jusqu'à 4 tubes par grenouille pour les grenouilles 40 g et plus. Il s'agissait d'un volume de collecte conservateur de 0,7 mL par 100 g, 70 % de la recommandation maximale de 1,0 ml par 100 g (adapté d'Allender et Fry, 2008)13.
L'anesthésie et la chirurgie chez les amphibiens sont rarement signalées, mais il est important de noter que les doses et l'efficacité varient d'une manière spécifique à une espèce. Dans Bombina orientalis par exemple, MS222 a un effet très faible, même avec des doses élevées (1 g/L) tandis que dans les adas boréaux, Anaxyrus boreas boreas, 1 g/L est rapide (matière de minutes) et de longue durée (3 h) (Calatayud, observation personnelle). Dans R. muscosa, l'anesthésie nécessite des doses signalées pour A. boreas boreas et a des effets similaires et des temps de récupération. Le jeûne des amphibiens avant l'anesthésie n'est généralement pas nécessaire car leur larynx reste étroitement fermé même sous anesthésie générale. Cependant, si cela est jugé nécessaire, surtout si la procédure anesthésique doit inclure la chirurgie célomic, les animaux peuvent être jeûnés 24 h avant l'anesthésie.
Pendant la chirurgie, le réflexe de redressement est le principal indicateur que l'animal est devenu anesthésié. Le réflexe de redressement est la capacité et le degré de facilité avec lesquels un animal peut revenir à une position verticale après avoir été placé sur le dos. La perte du réflexe suggère un stade léger de l'anesthésie. Un plan chirurgical est indiqué par la perte du réflexe de sevrage qui comprend tirer légèrement sur le membre pour le redresser et l'animal n'est plus capable de le rétracter7. La chirurgie de reproduction n'a pas d'obstacles écrasants et les patients amphibiens guérissent principalement bien tolérer la perte de sang plus que les vertébrés plus élevés. La chirurgie doit se faire rapidement, d'une durée approximative de 15 minutes du début à la fin. Les étapes doivent être chronométrées approximativement comme suit : 'lt;1 minute pour l'incision initiale et 'lt;2 minute pour l'insertion de céliontomie et de rétracteur, 'lt; 2-3 minutes pour l'isolement par ovaire et 'lt;1 minute pour la suture de navire ou la cautérisation et la suture de peau 'lt; 4 minutes. Le temps total de récupération après la chirurgie avec les protocoles MS222 est d'environ 45 minutes, mais cela peut être spécifique à l'espèce. Dans A. boreas boreas et R. muscosa temps de récupération peut être plus long, jusqu'à 1 à 2 h. Lors de l'exécution de la chirurgie, il faut prendre soin d'éviter de perforer les poumons, le tractus gastro-intestinal ou une vessie distendue, et de ne pas endommager les glandes macroscopiques, les cœurs lymphatiques et les vaisseaux sanguins, en particulier la veine mi-ventrale. Selon la saison, la présence de gros corps gras peut rendre la visualisation d'autres organes difficile. Une fois visiblement éveillé, les réactions d'un animal à la stimulation des membres, telles que la résistance à un étirement doux d'un membre arrière ou clignotant lorsque la zone autour de l'œil est stimulée (observation personnelle), sont classées comme des réponses de sevrage. Le réflexe de redressement ainsi que d'autres indicateurs de récupération, y compris, les réflexes de retrait et les mouvements gulaires, sont des indicateurs importants de la récupération.
| gouvernement | ||||||||||
| Nom commun | espèce | hormone | procédure | Composé d'amorçage | Dose d'amorçage signalée | Nombre de doses d'amorçage | Calendrier (hr avant la dose ovulatoire) | Composé(s) administré pour l'ovulatoire/oviposition finale | Doses | référence |
| Le tondeporto crested | Lémurien peltophryne | GnRH et hCG | Ip | Hcg | 1.5 UI/g | 2 (en) | hCG - 48 | GnRH; hCG; GnRHa et hCG | 0,2 g; 4 UI; 0,5 g à 4 UI | Calatayud et coll. inédits |
| Grenouille jaune-jambes de montagne | Rana muscosa | Amphiplex, Lut | Ip | GnRHa (des-Gly10, D- Ala6, Pro-NHEt9-GnRH) | 0,4 g/g | 1 Fois | 24 Ans, états-unis | GnRH et MET | 1 x 0,4 g/g 10 g/g | Calatayud et coll., 2018 |
| PGF2 | Im | PGF2 | 5 ng/g | 1 Fois | 48 Annonces | PGF2 | 5 ng/g | |||
| Le vaad-toi boréal des Rocheuses du Sud | Anaxyrus boreas boreas | hCG, GnRH | Ip | Hcg | 3.7 UI/g | 2 (en) | 96, 24 ans | hCG et GnRHa | 13,5 UI/g 0,4 g/g | Calatayud et coll., 2015 |
| Grenouille nordique de cricket | Acris crepitan | Amphiplex | ajouté à l'eau (10 ml) | aucun | aucun | 0 (en) | Na | GnRH et MET | 0,17 g et 0,42 g / l | Snyder et coll., 2012 |
| Grenouille de léopard nordique | Lithobates pipiens | Amphiplex | Ip | aucun | aucun | 0 (en) | 24 Ans, états-unis | GnRH et MET | 1 x 0,4 g/g 10 g/g | Trudeau et coll., 2010 |
| Grenouille à cornes argentine | Ceratophyrs ornata | Amphiplex | Ip | aucun | aucun | 0 (en) | 24 Ans, états-unis | GnRH et MET | 1 x 0,4 g/g 10 g/g | |
| Grenouille à cornes de Cranwell | Ceratophrys cranwelli | Amphiplex | Ip | aucun | aucun | 0 (en) | 24 Ans, états-unis | GnRH et MET | 1 x 0,4 g/g 10 g/g | |
| Grenouille de sol américaine | Odontophrynus americanus | Amphiplex | Ip | aucun | aucun | 0 (en) | 24 Ans, états-unis | GnRH et MET | 1 x 0,4 g/g 10 g/g | |
| Grenouille de Gopher de Dusky | Rana sevosa | hCG, GnRH | Ip | Hcg | 3.7 UI/g | 2 (en) | 96 , 24 | GnRH et hCG | 1 x 0,4 g/g 13,5 UI/g | Graham et coll., 2018 |
| Coqui commun | Eleutherodactylus coqui | Poissons, aviaires, mammifères et GnRH (D-Ala, des-Gly, eth LHRH), hCG | Sc | mLHRH; aLHR; fLHRH; GnRHa; Hcg | aucun | 0 (en) | Na | mLHRH; aLHR; fLHRH; GnRHa; Hcg | 7 g, 33 g; 28 g; 7 g, 20 g; 5, 10, 15, 20 g; 165 UI | Michael et coll. |
| Le toadlet de Gunther | Pseudophryne guentheri | Gnrh | GnRHa (En anglais) | 0,4 g/g | 1 Fois | 26 Annonces | GnRHa avec ou sans prime | 0,4 g/g | Silla 2010 | |
| Grenouille de Corroboree | Corroboree pseudophryne | Lucrin Lucrin | Sc | Lucrin Lucrin | 1 g | 1 Fois | 26 Annonces | Lucrin Lucrin | 5 g | Byrne et Silla, 2010 |
| Grenouille de Corroboree nordique | Pseudophryne pengilleyi | GnRHa GnRH (D-Ala, des-Gly, eth LHRH) | merci | aucun | aucun | 0 (en) | Na | GnRHa (En anglais) | 0,5 -2,0 g/g | Silla et coll., 2017 |
| Chien d'eau de côte du golfe | Necturus beyeri | [des-Gly10, D- Ala6]-LhRH-RH hydrate de sel d'éthylatate | Ip | aucun | aucun | 0 (en) | Na | Lhrh | 100 g / 500 l | Stoops et coll., 2014 |
| Grenouille de cloche méridionale / grenouille d'herbe grognant | Litoria raniformis | des-Gly10, D- Ala6-[LHRH] | Sc | aucun | aucun | 0 (en) | Na | des-Gly10, D- Ala6-[LHRH] | 50 g | Mann et coll., 2010 |
| Le crapaud de Fowler | Anaxyrus fowleri | GnRH, HCG, P4 | Ip | Hcg | 3.7 UI/g | Browne et coll., 2006 | ||||
| Axolotl (salamandre mexicaine) | Ambystoma mexicanum | Hormones follicules-stimulantes | Im | aucun | aucun | 0 (en) | Na | Fsh | 400IU (en) | Trottier et Armstrong, 1974 |
| Grenouille à griffes africaines | Xenopus laevis | hCG et P4 | ajout d'eau; Ip | PMSG, HCG | Marcec , 2016 | |||||
| Salamandre de tigre | Ambystoma tigrinum | hCG, LH | ||||||||
| Le poulain du Wyoming | Anaxyrus baxteri | hCG, GnRHa, P4 | Ip | hCG et GnRHa | 100 UI 0,8 g | 1 Fois | 72 Annonces | hCG et GnRHa | 100 UI 0,8 g | Browne et coll., 2006 |
| Grenouille de léopard nordique | Lithobates pipiens | Extrait pituitaire (PE), P4, testostérone (T), corticostérone [C], Amphiplex, dompéridone (D) | SC, IP | aucun | aucun | 0 (en) | Na | PE, PE-T, PE-P4, PE-C; Amphiplex, GnRH et D | 100 UI (LHRH) en 1 ml; PE-0,002 g/L; PE 0,01 mg/50mL; PE-0.1mg/50mL; 0,4 g/g et 10 g/g; 0,4 g/g et D | Wright, 1961; Fort, 2000; Trudeau et coll., 2013 |
| Grenouille moulue | Lymnodynastes tasmeniensis | Extraits pituitaires, hCG, GnRHa, PZ | Ip | GnRHa (En anglais) | 0,9 à 1,2 g/g et PZ 10 g/g | 1 Fois | 20 Ans, états-unis | PE; PE et hCG; GnRH et PZ | PE vol; PE vol 100 UI hCG; GnRH (0.9-1.2g/g) 'PZ '10'g/g) | Clulow et coll., 2018 |
| Grenouille verte et d'or de cloche | Litoia aurea | Gnrh | Ip | GnRHa (En anglais) | 10 g | 1 Fois | 72 Annonces | GnRHa et hCG | 20 g et 300 UI | Clulow et coll., 2018 |
| Grande grenouille barrée | Fasciolatus Mixophyes | hCG et PMSG | Sc | PMSG, HCG | 50 UI et 25 UI; 1x100 UI | 2; 2 (en) | PMSG-144 et 96; hCG-24 | Hcg | 100IU (en) | Clulow et coll., 2012 |
| Pour plus de protocoles hormonaux et d'espèces voir Wright et Whitaker, 2001 | ||||||||||
Tableau 1 : Espèces d'amphibiens et certaines des hormones exogènes testées sur elles comme indiqué dans la littérature. Gonadotropine chorionique humaine (hCG); Gonadotropin libérant-hormone (GnRH); L'hormone de Lutenizing-libération d'hormone (LHRH); les lettres m, a et f représentent «mammifère», «aviaire» et «poisson»; gonadotropine enceinte de sérum de jument (PMSG); progestérone (P4); Hormone Follicle-stimulantE FSH); extrait pituitaire (PE); testostérone (T); corticostérone (C). Les antagonistes de la dopamine énumérés comprennent : la dompéridone (D); Pimozide (P); métoclopramide (MET). Amphiplex est le nom donné à un composé composé de GnRH et Metoclopramide27. Lucrin est un agoniste GnRH disponible dans le commerce avec l'ingrédient actif étant l'acétate de Leuprorelin. 4 ( en plus) , 7 Annonces , 17 Annonces , 18 ans, états-unis qui , 19 ans, états-unis qui , 20 Ans, états-unis , 26 Annonces , 27 Annonces , 38 Annonces , 39 Ans et plus qu'ils , 40 ans, états-unis ( , 41 Ans, états-unis ( , 42 Ans, états-unis ( , 43 Ans, états-unis ( , 44 Ans, en est à qui , 45 Annonces
| note | Statut reproducteur | description |
| 0 (en) | Non-gravid | Pas d'œufs visibles. |
| 1 Fois | Gravid précoce | Oeufs visibles (1-2mm de taille) aucune ligne échogénique distincte associée à l'œuf. |
| 2 (en) | Gravid moyen | Oeufs de 2 à 3 mm de taille, lignées échogéniques distinctes associées à chaque œuf. |
| 3 (en) | Gravid tardif | Oeufs de 4-5mm dans la taille, lignes échogéniques encore visibles, augmentation marquée de l'aspect anéchoïque de l'oeuf. |
| 4 ( en plus) | Oeufs conservés | Des degrés variables de matière échogénique présent dans la structure interne de l'œuf prennent une forme amorphique. Certains peuvent devenir très échogéniques et associés à la rétention de liquide dans la cavité corporelle. |
Tableau 2 : Système de classement utilisé pour marquer l'état reproducteur des femmes captives Necturus et Rana muscosa par ultrasonographie.
| drogue | Dosage et Route | Commentaires - référence |
| Buprénorphin | 50 mg/Kg (intracelomic) | Étude expérimentale dans un triette rouge-spotted de l'est (Notophthalmus viridescens). L'analgésie doit être administrée avant la chirurgie. (Koeller, 2009) |
| Butorphanol Butorphanol | 1 à 10 mg/kg (IM ou intracelomic) | Il y a plusieurs responsabilités spécifiques. Il est conseillé de commencer à 1 mg/kg. |
| Butorphanol Butorphanol | 0,5 mg/L (bain) | Étude expérimentale dans un triette rouge-spotted de l'est (Notophthalmus viridescens). (Koeller, 2009) |
| Fentanyl | 1 mg/kg | Analgésie -4 h, antagonisé par la naltrexone (Stevens, 1997) |
| Meloxicam | 0,1 à 0,2 mg/kg (IM) | (Minter, 2011) |
Tableau 3 : Protocoles pour l'analgésie chez les amphibiens.
| Rana muscosa | ||
| an | En 2014 | En 2015 |
| Non, ♀ | 18 ans, états-unis qui | 18 ans, états-unis qui |
| Développement d'œufs | 61 % | 94 % |
| Contrôle ♀ | 4 ( en plus) | 6 Annonces |
| ♀ amphiplex | 4 ( en plus) | 7 Annonces |
| Moyenne jour après Amphiplex à oviposit | 10,5 Annonces | 10,9 |
| Taux d'oviposition (Amphiplex) | 22,20 % | 33,33 % |
| Taux d'oviposition (contrôle) | 22,20 % | 38,88 % |
Tableau 4 : Comparaison des paramètres reproducteurs entre les amphiplex-traités par rapport aux femelles capturées de contrôle Rana muscosa en 2014 et 2015.
| Necturus Necturus sp. | |||||
| an | 1 Fois | 2 (en) | 3 (en) | 4 ( en plus) | 5 Annonces |
| Non, ♀ | 6 ans et plus | 7 Annonces | 7 ans et plus | 7 ans et plus | 7 ans et plus |
| Développement d'œufs | 83% | 100% | 86% | 86% | 86% |
| ♀ LHRH | 3 (en) | 5 Annonces | 3 (en) | 6 Annonces | 0 (en) |
| Contrôle ♀ | 2 (en) | 2 (en) | 3 (en) | 0 (en) | 6 Annonces |
| Jour post-LHRH à Oviposit | 5 Annonces | 7 Annonces | 5.5 (gamme 3 - 8) | 13 (en) | ne s'applique pas |
| Taux d'oviposition (LHRH) | 60% | 20% | 67 % | 17 % | ne s'applique pas |
| Taux d'oviposition (contrôle) | 50% | 50% | 100% | ne s'applique pas | 67 % |
| n 1 ♀ pas de développement d'œufs | |||||
Tableau 5 : Comparaison des paramètres de reproduction entre l'HRHS (GnRH) -traité et contrôle (eau stérile) femelle captive Necturus Necturus de trois espèces sur une période de 5 ans (2012-2017).

Figure 1 : Trois méthodes de tenue d'une grenouille. (A) Procédure 1. (B) Procédure 2. (C) Procédure 3. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 2 : Évaluations morphométriques. (A, B) SVL/SUL (C, D). dans R. muscosa et D. Necturus. (E) Mesure de taille avec étriers. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 3 : Le dimorphisme sexuel se distingue par des coussinets de pouce nuptiaux sur les mâles adultes de R. muscosa comparés aux femelles. (A) Femme (B) Homme. Le panneau inférieur montre la longueur du mâle contre la femelle. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 4 : Caractériser les comportements reproducteurs menant à l'oviposition dans R. muscosa (A) courtiser. (B) Amplexus. (C) Mâle serrant la femelle tandis qu'en amplexus. (D, E) Femelle ample dans un support de main. (F, G) Contractions abdominales et oviposition. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 5 : Ultrasons effectués sur R. muscosa A-B avec l'état reproducteur selon le stade de développement4. (A, B) Exécution de l'échographie sur Rana muscosa. (C) Grade 0. (D) Grade 1. (E) Grade 2. (F) Grade 3. (G) Grade 4 (ovulé s'il y a des œufs et conservés) Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 6 : Images ultrasons de Necturus. (A) Grade 1. (B) Grade 2. (C) Oeufs de 3e année. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 7 : Collecte de sang à R. muscosa. (A) Collecte de sang en perforant l'aérospatial orbitalis postérieure à la veine faciale juste au-dessus de la mâchoire au milieu de l'orbite. (B, C) Le sang est libéré sur la surface de la peau et est recueilli avec un tube capillaire héparinisé. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 8 : Méthodes d'injection chez les amphibiens. Selon la profondeur du type d'injection, l'angle et la profondeur de l'aiguille varient. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 9 : Injection hormonale dans R. muscosa. Induction de l'oviposition par traitement hormonal dans les femelles de Rana muscosa injectées avec l'amphiplex intra-peritoneally. Ovaries peuvent être trouvés dans la cavité coelomic S'il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 10 : Préparation avant l'opération. (A) Préparation aseptique de la zone chirurgicale utilisant la solution povidone-iode diluée (1/10), Resinifictrix de Trachycephalus. (B) Incision propre de scalpel dans un Laevis de Xenopus ou, (C) incision de peau de laser-diode, Lithobates catesbeianus. (D) Évitez d'endommager la veine mi-ventrale, Trachycephalus resinifictrix. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 11 : Ovariectomy dans Xenopus laevis. (A) Exposer et déplacer de gros corps gras pour découvrir la masse des œufs. (B) Excise une portion de la masse d'oeufs sans ligature tous les vaisseaux sanguins. (C) Cauterize surrounding blood vessels by electrocautery for complete ovariectomy. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 12 : Préparation préchirurgicale et ovarice crevaison dans A. mexicanum. (A) Gaze stérile trempée, solution de chlorhexidine de 0,75% appliquée au site chirurgical (B). Une ligne entre l'épaule et les membres postérieurs divise l'animal en trois parties égales et la tache bleue marque le site sur l'incision. (C) Rétracter les incisions coélomiques avec des rétracteurs de paupière. (D) Pour l'ovaricectomie complète, cautérisez les vaisseaux sanguins environnants par électrocautérisation. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 13 : Évaluation visuelle des stades de reproduction. (A, B) Évaluations visuelles de l'étape reproductrice par la peau semi-translucide, Necturus. (C) Peau translucide, Hyalinobatrachium (grenouille de verre). (D) Évaluation visuelle de R. muscosa avant (droite, ligne bleue) et après oviposition (gauche - ligne jaune). Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 14 : Rétention d'œufs. (A, B) Rana muscosa femelle avec le cas grave de la conservation d'oeuf. (C) L'échographie montre de vieux œufs dégénérants, des œufs (en haut) et des œufs plus gros (panneau moyen et inférieur) ovulés et piégés dans le coelom. (D) Oeufs conservés récupérés par décapage manuel. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 15 : Images échographiques d'œufs conservés dans Necturus qui (A) sont devenus échogéniques en apparence (cercle) et ont été associés à (B) rétention de liquide dans la cavité du corps (flèche). Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 16 : Pourcentage de Necturus femelles captives qui ont oviposited des embrayages pleins ou partiels (2013-2017) comparés à ceux qui n'ont pas oviposit. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}
Discussion
La manipulation directe, l'observation visuelle et les mesures morphométriques fournissent des techniques non invasives et sont les premiers critères d'évaluation pour déterminer l'étape de reproduction féminine. Cependant, cette étude montre que les ovaires gravidés ne peuvent pas toujours être identifiés de façon fiable par palpation. Selon l'espèce, les ovaires gravidpeuvent peuvent parfois être détectés visuellement par la peau semi-translucide (figure13A,B) ou complètement translucide sur le côté ventral de l'animal (Figure 13C). Les femelles qui ont terminé l'oviposition peuvent montrer des changements évidents à leur apparence par rapport aux femelles gravid (par exemple, la peau lâche et la perte de jusqu'à 30% de leur masse corporelle, Figure 13D). Pendant la reproduction, les mâles et les femelles afficheront certains comportements qui fournissent des informations sur la proximité de l'ovulation et de l'oviposition. Dans le cas de R. muscosa indications qu'une femelle est proche de l'oviposition commencent par la femelle entrant dans les poignées.
L'application de la technologie d'ultrason aux Anurans et aux Caudates permet le diagnostic de la présence ou de l'absence d'ovules et si l'oviposition a été associée à la libération complète ou partielle d'ovules développés. Ainsi, cette méthode fournit une évaluation plus complète et précise de l'état reproducteur sans se limiter à déterminer le statut gravid/non-gravid par l'intermédiaire d'une technique de visualisation qui varie par la transparence abdominale de peau, ou la cohérence épidermique parmi les différentes espèces d'amphibiens. L'échographie peut être effectuée avec une relative facilité et avec peu de stress pour les animaux (Figure 5 et Figure 13) et peut être utilisée pour caractériser les cycles reproducteurs et déterminer l'état reproducteur4. Il est essentiel de se familiariser avec l'espèce; cependant, cette étude a montré que Necturus et R. muscosa partagent des signes de développement communs dans leurs modèles reproducteurs permettant une classification similaire de l'étape de reproduction (figure 5). Grâce à cette technologie, il est maintenant prouvé que le développement d'œufs est élevé en captivité Necturus et R. muscosa et que ces deux espèces suivent un modèle saisonnier. Bien que les raisons de ces phénomènes soient inconnues et nécessitent une enquête plus approfondie, sans l'utilisation de l'échographie, plusieurs domaines de dysfonctionnement ovarien, tels que la rétention d'ovules et l'oviposition partielle, seraient passés inaperçus. Des applications futures à cette technique seront utilisées pour déterminer si les femelles devraient être sélectionnées pour la reproduction au cours d'une année donnée et si l'oviposition est terminée.
Un protocole de collecte de sang, tel que celui présenté dans R. muscosa, qui est à la fois efficace et cause une détresse minimale à l'animal, est optimal pour étudier les profils d'hormones chez les Anurans captifs et capturés à l'état sauvage (Calatayud, non publié). À ce jour, aucune information n'existe concernant les profils hormonaux annuels de la r. muscosa captive et, par conséquent, aucune connaissance sur la façon dont les hormones influencent leur santé et leur reproduction. En outre, avec la preuve que les femelles de cette espèce peuvent ne pas être des éleveurs annuels, le profilage hormonal sera une autre méthode pour suivre les cycles ovariens. Avec l'échographie, l'analyse hormonale peut conduire à une meilleure prédiction de ce que les femelles seront prêtes pour l'oviposition. En outre, au cours de la dernière année, deux cas d'intersexués dans la population captive de R. muscosa ont été documentés. En outre, le développement des coussinets de pouce a été noté sur quelques-unes des femelles fondatrices plus âgées. Les raisons de ceci sont actuellement à l'étude mais les résultats initiaux suggèrent qu'il peut se rapporter aux changements des niveaux de testostérone (Calatayud, non édité). Discerner les cycles hormonaux chez les femmes de différents âges nous aidera à comprendre pourquoi les femmes peuvent développer des caractéristiques sexuelles secondaires associées aux hommes et si cela doit être prévu dans une population vieillissante.
L'hormonothérapie exogène a été utilisée pour surmonter les dysfonctionnements reproducteurs fréquemment rencontrés chez les amphibiens captifs. Cependant, pour les populations de R. muscosa et de Necturus dans cette étude, aucune différence significative dans l'oviposition entre les femelles hormone-traitées et témoins n'a été détectée sur une période de 2 et 5 ans, respectivement. Ceci peut indiquer que le protocole d'administration d'hormone, les doses, l'amorçage et la combinaison d'hormone utilisés n'étaient pas proportionnels pour l'espèce. Une analyse plus approfondie des antécédents reproducteurs féminins individuels suggère que R. muscosa pourrait ne pas connaître de reproduction annuelle, ce qui pourrait également expliquer l'absence d'effet hormonal observé chez les femelles traitées. Étant donné qu'un certain pourcentage de femelles sautaient systématiquement la reproduction chaque année, la compréhension de l'histoire naturelle de l'espèce peut aider à déterminer s'il y a un besoin d'hormones exogènes et quand elles peuvent être les plus efficaces. Les procédures décrites dans cet article peuvent être appliquées à un certain nombre d'espèces, (tableau 1) et sont pour les anurans allant de 5 g à 150 g; les animaux plus gros peuvent avoir besoin de seringues et de jauges d'aiguille différentes. L'emplacement de l'injection varie selon certaines hormones nécessitant une injection intramusculaire, intrapéritole, sous-cutanée ou intradermique (figure 7).
La chirurgie aux fins de l'ovaricectomie est une méthode courante utilisée chez diverses espèces d'amphibiens pour obtenir des ovocytes pour des études embryologiques. L'ovaricectomie peut également être indiquée pour le contrôle de la population et les problèmes médicaux tels que la rétention des œufs. Dans le cas d'ovariectomies partielles dans lesquelles, la récolte d'ovocytes est effectuée à des fins de recherche, la chirurgie doit s'assurer que l'animal reste reproducteur. L'administration de PGF2 a montré une certaine promesse dans la résolution de la rétention d'œufs chez les femelles R. muscosa. Chez plusieurs personnes, Le PGF2 a obtenu le dépôt complet des oeufs précédemment retenus mais dans d'autres seulement le dépôt partiel s'est produit exigeant le décapage manuel pour enlever tous les oeufs. Tandis que PGF2 peut servir d'alternative à la chirurgie pour la conservation d'oeuf dans R. muscosa,sa capacité à remédier aux conditions pathologiques semblables dans d'autres amphibiens exigera la validation d'espèce-spécifique. Lorsque l'intervention chirurgicale est obligatoire pour le patient Anuran ou Caudate, il est nécessaire d'assurer un plan adéquat d'anesthésie avant les incisions sont faites. Des compétences d'observation astucieuses sont nécessaires pour évaluer et surveiller les réponses normatives d'induction et de rétablissement telles qu'décrites dans cette étude pour chacun des taxons. Une fois que l'on est familier avec l'anatomie spécifique, une approche chirurgicale appropriée, l'hémostasie, la manipulation douce de tissu et la gestion postopératoire adéquate, les chirurgies reproductrices ne posent aucun obstacle écrasant.
Déclarations de divulgation
Les auteurs n'ont rien à révéler.
Remerciements
Natalie Calatayud tient à remercier la Dre Barbara Durrant pour sa formation et son aide en échographie et à Exploradora de Immuebles, S.A. (EISA) pour avoir accordé une aide financière à mon poste d'associé de recherche chez SDZG. Merci à la Dre Kylie Cane pour ses commentaires sur le manuscrit ainsi qu'aux réviseurs officiels (qui qu'ils soient). Merci à Jonathan Dain notre boursier d'été 2018, San Diego Zoo Institute for Conservation Research pour fournir des photos (Figure 1A, B). Monica Stoops remercie l'Association of Zoos and Aquariums Conservation Endowment Fund et le Disney Worldwide Conservation Fund d'avoir fourni un soutien financier pour établir la population captive de Necturus. De plus, le soutien a également été reçu grâce à des dons privés de Mme Iris de la Motte, avocate des amphibiens. Merci, est remis à M. Christopher DeChant et le Dr Mark Campbell pour leur contribution importante à la recherche.
matériels
| Name | Company | Catalog Number | Comments |
| GE logiq Book XP and 8C-RS probe 4e10 MHz GE Medical Systems | GE medical systems | GE logiq Book XP | Ultrasound |
| Aloka 500 7.5mHz linear or IBEX multi-frequency (10-6mHz) micro-convex | GE medical systems | 8C-RS (10 MHz) | Ultrasound probe |
| BD disposable U-100 insulin syringe (28-29 G needle) | Mettler Electronics Corp CA | Sonigel | Ultrasound gel (water soluble, salt-free) |
| Hormone | |||
| Gonadotropin releasing hormone | BACHEM | 4012028 | synonym: [Des-Gly10, D-Ala6, Pro-NHEt9]-GnRH acetate abbreviation: GnRH |
| Lutenizing hormone releasing hormone | BACHEM, Sigma-Aldrich | 4033013; L1898 | synonym: Pyr-His-Trp-Ser-Tyr-Gly-Leu-Arg-Pro-Gly-NH2 acetate salt; [D-Ala6}-LHRH acetate salt hydrate abbreviation: LHRH |
| Human chorionic gonadotropin | BACHEM, Sigma-Aldrich | 4030270, 4018894; C1063, CG5, CG10 | synonym: Chorionic gonadotropin-β (109-145)(109-119); Choriogonin, HCG (2,500IU, 5,000IU, 10,000IU) abbreviation: hCG |
| Prostaglandin 2α | Sigma-Aldrich | P40424; | synonym: (5Z,9α,11α,13E,15S)-9,11,15-Trihydroxyprosta-5,13-dienoic acid tris salt, PGF2α−Tris; abbreviation: PGF2α |
| Follicle-stimulating hormone | Sigma-Aldrich | F4021, F8174 | synonym: porcine, sheep abbreviation: FSH |
| Progesterone | Sigma-Aldrich | 46665; P7556 | synonym: Vetranal; P4 water soluble abbreviation: P4 |
| Pituitary extract | na | synonym: Check papers for amphibian species derivation abbreviation: PE | |
| Pregnant Mare Serum Gonadotropin | Prospec; Lee Biosolutions; Sigma-Aldrich | HOR-272; 493-10; 9002-70-4 | synonym: Pregnant Mare Serum Gonadotropin abbreviation: PMSG |
| Metaclopromide | Sigma-Aldrich | M0763 | synonym: 4-Amino-5-chloro-N-[2-(diethylamino)ethyl]-2-methoxybenzamide, Methoxychloroprocainamide abbreviation: MET |
| Lucrin | BACHEM; Sigma-Aldrich | 4033014; L0399 | synonym: Leuprorelin acetate abbreviation: Lucrin |
| Lutalyse | Pfizer | synonym: PGF2α - Dinoprost tromethamine abbreviation: Lut | |
| Pimozide | Sigma-Aldrich | P1793 | synonym: 1-[1-[4,4-bis(4-Fluorophenyl)butyl]-4-piperidinyl]-1,3-dihydro-2H-benzimidazol-2-one abbreviation: PZ |
| Amphiplex | see above | synonym: Gonadotropin releasing hormone + metoclopramide abbreviation: GnRH + MET | |
| Ovopel | Ovopel | na | synonym: GnRHa + dopamine receptor antagonist (administered 1 pellet/ kg) abbreviation: Ovo |
| Ovaprim | Pentair aquatic eco-systems | Ova10 | synonym: Salmon gonadotropin + domperidone abbreviation: Ova |
| Domperidone | Sigma-Aldrich | D122 | synonym: 4-(5-Chloro-2-oxo-1-benzimidazolinyl)-1-[3-(2-oxobenzimidazolinyl)propyl]piperidine abbreviation: DOM |
Références
- Conde, D. A., Flesness, N., Colchero, F., Jones, O. R., Scheuerlein, A. An emerging role of zoos to conserve biodiversity. Science. 331 (6023), 1390-1391 (2011).
- Conde, D. A., et al. Zoos through the Lens of the IUCN Red List: A Global Metapopulation Approach to Support Conservation Breeding Programs. PLoS ONE. 8 (12), e80311(2013).
- Morrison, C., Hero, J. -M. Geographic variation in life-history characteristics of amphibians: a review. Journal of Animal Ecology. 72 (2), 270-279 (2003).
- Calatayud, N. E., Stoops, M., Durrant, B. S. Ovarian control and monitoring in amphibians. Theriogenology. , 70-81 (2018).
- National Research Council. Institutional Animal Care and Use Committee Guidebook. , (2010).
- Peig, J., Green, A. J. New perspectives for estimating body condition from mass/length data: the scaled mass index as an alternative method. Oikos. 118 (12), 1883-1891 (2009).
- Graham, K. M., Langhorne, C. J., Vance, C. K., Willard, S. T., Kouba, A. J. Ultrasound imaging improves hormone therapy strategies for induction of ovulation and in vitro fertilization in the endangered dusky gopher frog (Lithobates sevosa). Conservation Physiology. 6 (1), coy020(2018).
- Wright, K. M., Whitaker, B. R. Amphibian Medicine and Captive Husbandry. , Krieger Publishing Company. (2001).
- Forzán, M. J., Vanderstichel, R. V., Ogbuah, C. T., Barta, J. R., Smith, T. G. Blood collection from the facial (maxillary)/musculo-cutaneous vein in true frogs (family Ranidae). Journal of Wildlife Diseases. 48 (1), 176-180 (2012).
- Allender, M. C., Fry, M. M. Amphibian hematology. Veterinary Clinics of North America: Exotic Animal Practice. 11 (3), 463-480 (2008).
- Green, S. L., Parker, J., Davis, C., Bouley, D. M. Ovarian hyperstimulation syndrome in gonadotropin-treated laboratory South African clawed frogs (Xenopus laevis). Journal of the American Association for Laboratory Animal Science. 46 (3), 64-67 (2007).
- Jorgensen, C. B. External and internal control of patterns of feeding, growth and gonadal function in a temperate zone anuran, the toad Bufo bufo. Journal of Zoology. 210 (2), 211-241 (1986).
- Jørgensen, C. B. Growth and reproduction. Environmental Physiology of the Amphibians. , 439-466 (1992).
- Vu, M., Trudeau, V. L. Neuroendocrine control of spawning in amphibians and its practical applications. General and Comparative Endocrinology. 234, 28-39 (2016).
- Clulow, J., Trudeau, V. L., Kouba, A. J. Amphibian declines in the twenty-first century: why we need assisted reproductive technologies. Reproductive Sciences in Animal Conservation. , 275-316 (2014).
- Kouba, A., et al. Assisted reproductive technologies (ART) for amphibians. Amphibian Husbandry Resource Guide. 2, 60-118 (2012).
- Clulow, J., et al. Optimisation of an oviposition protocol employing human chorionic and pregnant mare serum gonadotropins in the Barred Frog Mixophyes fasciolatus (Myobatrachidae). Reproductive Biology and Endocrinology. 10 (1), 60(2012).
- Browne, R. K., Seratt, J., Vance, C., Kouba, A. Hormonal priming, induction of ovulation and in-vitro fertilization of the endangered Wyoming toad (Bufo baxteri). Reproductive Biology and Endocrinology. 4 (1), 34(2006).
- Calatayud, N. E., et al. A hormone priming regimen and hibernation affect oviposition in the boreal toad (Anaxyrus boreas boreas). Theriogenology. 84 (4), 600-607 (2015).
- Stoops, M. A., Campbell, M. K., Dechant, C. J. Successful captive breeding of Necturus beyeri through manipulation of environmental cues and exogenous hormone administration: a model for endangered Necturus. Herpetological Review. 45 (2), 251-256 (2014).
- Mc Creery, B. R., Licht, P. Induced ovulation and changes in pituitary responsiveness to continuous infusion of gonadotropin-releasing hormone during the ovarian cycle in the bullfrog, Rana catesbeiana. Biology of Reproduction. 29 (4), 863-871 (1983).
- Johnson, C. J., Vance, C. K., Roth, T. L., Kouba, A. J. Oviposition and ultrasound monitoring of American toads (Bufo americanus) treated with exogenous hormones. Proceedings of the American Association of Zoo Veterinarians. 299, 301(2002).
- Herbert, D. Studies of assisted reproduction in the spotted grass frog Limnodynastes tasmaniensis: ovulation, early development and microinjection (ICSI). , (2004).
- Michael, S. F., Buckley, C., Toro, E., Estrada, A. R., Vincent, S. Induced ovulation and egg deposition in the direct developing anuran Eleutherodactylus coqui. Reproductive Biology and Endocrinology. 2, 6(2004).
- Ogawa, A., Dake, J., Iwashina, Y., Tokumoto, T. Induction of ovulation in Xenopus without hCG injection: the effect of adding steroids into the aquatic environment. Reproductive Biology and Endocrinology. 9 (1), 11(2011).
- Silla, A. J. Effects of luteinizing hormone-releasing hormone and arginine-vasotocin on the sperm-release response of Günther’s Toadlet, Pseudophryne guentheri. Reproductive Biology and Endocrinology. 8 (1), 139(2010).
- Trudeau, V. L., et al. Hormonal induction of spawning in 4 species of frogs by coinjection with a gonadotropin-releasing hormone agonist and a dopamine antagonist. Reproductive Biology and Endocrinology. 8 (1), 36(2010).
- Krause, E. T., von Engelhardt, N., Steinfartz, S., Trosien, R., Caspers, B. A. Ultrasonography as a minimally invasive method to assess pregnancy in the fire salamanders (Salamandra salamandra). Salamandra. 49, 211-214 (2013).
- Browne, R. K., Li, H., Seratt, J., Kouba, A. Progesterone improves the number and quality of hormone induced Fowler toad (Bufo fowleri) oocytes. Reproductive Biology and Endocrinology. 4 (1), 3(2006).
- Bramucci, M., et al. Different modulation of steroidogenesis and prostaglandin production in frog ovary in vitro by ACE and ANG II. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 273 (6), R2089-R2096 (1997).
- Gobbetti, A., Zerani, M. Possible roles for prostaglandins E2 and F2α in seasonal changes in ovarian steroidogenesis in the frog (Rana esculenta). Journal of Reproduction and Fertility. 98 (1), 27-32 (1993).
- Gobbetti, A., Zerani, M., Carnevali, O., Botte, V. Prostaglandin F2α in female water frog, Rana esculenta: Plasma levels during the annual cycle and effects of exogenous PGF2α on circulating sex hormones. General and Comparative Endocrinology. 80 (2), 175-180 (1990).
- Guillette, L. J. Jr, Dubois, D. H., Cree, A. Prostaglandins, oviducal function, and parturient behavior in nonmammalian vertebrates. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 260 (5), R854-R861 (1991).
- Clulow, J., Mahony, M., Browne, R., Pomering, M., Clark, A. Applications of assisted reproductive technologies (ART) to endangered anuran amphibians. Declines and Disappearances of Australian Frogs'. Campbell, A. , 219-225 (1999).
- Browne, R. K., Wolfram, K., García, G., Bagaturov, M. F., Pereboom, Z. Zoo-based amphibian research and conservation breeding programs. Amphibian and Reptile Conservation. 5 (3), 1-14 (2011).
- Chai, N. Surgery in amphibians. Veterinary Clinics: Exotic Animal Practice. 19 (1), 77-95 (2016).
- Gentz, E. J. Medicine and surgery of amphibians. Ilar Journal. 48 (3), 255-259 (2007).
- Snyder, W. E., Trudeau, V. L., Loskutoff, N. M. 168 a noninvasive, transdermal absorption approach for exogenous hormone induction of spawning in the northern cricket frog, Acris crepitans: a model for small, endangered amphibians. Reproduction, Fertility and Development. 25 (1), 232-233 (2012).
- Kouba, A. J., et al. Emerging trends for biobanking amphibian genetic resources: the hope, reality and challenges for the next decade. Biological Conservation. 164, 10-21 (2013).
- Michael, S. F., Buckley, C., Toro, E., Estrada, A. R., Vincent, S. Induced ovulation and egg deposition in the direct developing anuran Eleutherodactylus coqui. Reproductive Biology and Endocrinology. 2 (1), 6(2004).
- Fort, D. J. Frog reproduction and development study 2000 rana pipiens reproduction and development study. , Environmental Protection Agency. (2003).
- Clulow, J., et al. Differential success in obtaining gametes between male and female Australian temperate frogs by hormonal review: A Review. General and Comparative Endocrinology. 265, 141-148 (2018).
- Trottier, T. M., Armstrong, J. B. Diploid gynogenesis in the Mexican axolotl. Genetics. 83 (4), 783-792 (1976).
- Marcec, R. M. Development of assisted reproductive technologies for endangered North American salamanders. , Mississippi State University. (2016).
- Wright, M. L. Melatonin, diel rhythms, and metamorphosis in anuran amphibians. General and Comparative Endocrinology. 4, (2002).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.