
Method Article
Enrichissement des spermatozoïdes pachytenés et des spermatides à partir de testeurs de souris à l'aide d'équipement de laboratoire standard
* Ces auteurs ont contribué à parts égales
Dans cet article
Résumé
Présenté ici est un protocole pour enrichir les spermatozoïdes pachytenés, spermatides ronds, et spermatids allongeant des testicules adultes de souris utilisant un gradient discontinu de densité d'albumine de sérum bovin avec l'équipement standard de laboratoire.
Résumé
Pour caractériser chaque étape de la spermatogénèse, les chercheurs doivent séparer différentes sous-populations de cellules germinales des testicules. Cependant, isoler les populations discrètes est difficile, parce que le testicule adulte contient un mélange complexe de cellules germinales de toutes les étapes de la spermatogénèse avec certaines populations de cellules somatiques. Au cours des dernières décennies, différentes techniques telles que l'élutriation centrifuge, le tri cellulaire activé par la fluorescence (FACS) et le STA-PUT ont été appliquées avec succès à l'isolement des cellules germinales. Un inconvénient est qu'ils ont tous besoin d'appareils dédiés et de formation spécialisée. Suivant les principes sous-jacents à la méthode STA-PUT, un protocole simple a été développé pour l'isolement des spermatocytes de pachytene, des spermatides ronds, et des spermatides d'allongement des testicules de souris. Après avoir préparé une suspension cellulaire unique des cellules testiculaires, des populations cellulaires spécifiques sont enrichies par la sédimentation gravitationnelle par un gradient de densité d'albumine de sérum bovin discontinu (BSA). Les fractions cellulaires sont ensuite collectées manuellement et analysées au microscope. Ce gradient de densité modifié pour le protocole de sédimentation des spermatides ronds (MDR) peut être largement appliqué, car il ne nécessite que de l'équipement de laboratoire standard. En outre, le protocole exige un minimum de matériaux de départ, ce qui réduit son coût et l'utilisation d'animaux de laboratoire.
Introduction
On ignore encore beaucoup des événements moléculaires et biologiques qui se produisent pendant la spermatogénèse des mammifères, un processus complexe dans lequel les cellules souches spermatogonielles se transforment en spermatozoïdes hautement spécialisés1,2. La spermatogénèse a lieu à l'intérieur des tubules séminifères du testicule. Les tubules contiennent un mélange de cellules germinales de chaque étape de différenciation, y compris les cellules souches spermatogonielles, la division mitotique spermatogonie, les spermatocytes méiotiques et les spermatides postméiotiques (qui subissent une différenciation héloïde spermatides à l'allongement des spermatides, et enfin à la maturité spermatozoée). Les cellules somatiques des testicules comprennent les cellules Sertoli qui sont mélangées avec des cellules germinales à l'intérieur des tubules séminifères, les cellules myoïdes périubulaires qui forment les parois des tubules, et les cellules Leydig productrices de testostérone dans l'espace interstitiel entre les tubules.
L'étude des processus moléculaires et biochimiques pendant la spermatogénèse nécessite souvent la séparation des populations distinctes de cellules germinales d'un mélange complexe de cellules testiculaires. De nombreuses stratégies différentes ont été développées pour l'enrichissement cellulaire. Les méthodes les plus réussies sont la sédimentation de vitesse STA-PUT par gravité unitaire3,4,5,6, élutriation centrifuge basée sur la centrifugation de contre-flux7,8, et le tri des cellules activées par la fluorescence (FACS) qui sépare les cellules en fonction de la teneur en ADN et/ou de marqueurs spécifiques. Ces méthodes sont couramment utilisées parmi les chercheurs en spermatogénèse et permettent l'enrichissement efficace de types de cellules germinales spécifiques. Cependant, une limitation de ces techniques est qu'elles exigent le matériel spécialisé et coûteux qui exigent l'expertise.
Présenté ici est un protocole simple et peu coûteux pour isoler les populations enrichies des trois populations cellulaires les plus abondantes de testicules de souris: spermatides ronds, spermatocytes pachytene, et spermatides allongeant. Ce protocole est appelé le gradient de densité modifié pour les spermatides ronds (MDR), car il fonctionne particulièrement bien pour enrichir les spermatides ronds. La méthode MDR est basée sur les mêmes principes que la sédimentation de vitesse STA-PUT, mais elle ne nécessite que de l'équipement de laboratoire standard. Les cellules vivantes sont autorisées à se sédimenter à l'intermédiaire d'un gradient de densité d'albumine de sérum bovin discontinu (BSA) préparé manuellement à l'intérieur d'un tube standard de 50 ml sous le champ gravitationnel de la terre. Les cellules plus grandes se déplacent plus rapidement à travers le gradient, qui sépare différentes populations de cellules germinales. Après la sédimentation, des fractions enrichies des trois types de cellules sont collectées manuellement. La pureté de ces populations cellulaires enrichies est comparable à celles obtenues par STA-PUT et l'élutriation centrifuge.
En plus de couvrir la construction et l'utilisation du gradient BSA pour la sédimentation de vitesse, le protocole décrit également une méthode de digestion pour libérer les cellules testiculaires des tubules séminifères. Le protocole a été modifié à partir de celui développé par Romrell et al.9 et comprend des digestions séquentielles avec de la collagène IV et de la trypsine. Les digestions séquentielles combinées à l'utilisation d'un tampon de bicarbonate (c.-à-d., la solution Krebs) ont été montrées pour améliorer considérablement la séparation et la viabilité des cellules germinales9.
Pendant l'enrichissement de MDR, les cellules passent environ 4 h ensemble en dehors de l'environnement des tubules séminifères et ne sont pas soumises à des forces mécaniques stressantes, ce qui permet la collecte de fractions cellulaires hautement viables pour l'analyse en aval. En outre, semblable à l'élutriation centrifuge et STA-PUT, le protocole MDR ne nécessite aucun traitement chimique ou l'étiquetage des cellules, ce qui contribue également à maintenir leur viabilité. Fait important, aussi peu que deux testicules de souris adultes sont suffisants pour l'isolement MDR et, par conséquent, une souris adulte fournit suffisamment de cellules enrichies pour l'ARN et l'analyse des protéines. Le protocole standard STA-PUT recommande l'utilisation de 12 souris adultes pour l'isolement cellulaire6; bien que, basé sur l'expérience antérieure, il est connu que l'isolement réussi peut être fait de trois à quatre souris adultes. La plus faible quantité de matériel de départ déclarée suffisante pour l'élutriation centrifuge est de six testicules de souris (trois souris)8. Par conséquent, en plus d'éliminer le besoin d'équipement spécialisé coûteux, le protocole MDR réduit le nombre d'animaux de laboratoire requis.
Protocole
L'entretien des souris de laboratoire et toutes les expériences ont été effectués conformément aux lignes directrices et règlements pertinents pour le soin et l'utilisation des animaux de laboratoire.
1. Équipement et mise en place de réactif
- Fixer le bain d'eau à 37 oC.
- Définir l'incubateur de culture cellulaire à 34 oC, 5 % CO2,95 % d'humidité. Placez le rotateur de tube à l'intérieur de l'incubateur.
REMARQUE: Les incubateurs nécessitent beaucoup de temps pour changer la température interne. Si un incubateur constamment réglé à 34 oC n'est pas disponible, configurez-en un 1 jour avant l'expérience. - Préparer et étiqueter la quantité appropriée de diapositives en verre de microscopie. Dessiner un anneau de 1 cm de diamètre avec un stylo à graisse et laisser sécher la graisse.
- Préparer 1x tampon Krebs, pH 7.8 (tableau 1). Mettre deux tubes coniques de 50 ml de 1x Krebs à 34 oC pour réchauffer pendant les étapes 2,5 à 2,8. Conserver le reste des 1x Krebs à 4 oC ou sur la glace.
- Préparer des solutions BSA. Préparer d'abord 25 mL de 10% (w/v) solution BSA dans Krebs en dissolvant 2,5 g de BSA en tampon 1x Krebs à un volume final de 25 mL. Diluer la solution BSA de 10 % avec un tampon 1x Krebs pour obtenir différentes concentrations de BSA (tableau 2). Gardez toutes les solutions BSA à 4 oC.
REMARQUE: Préparer les solutions le jour même de la procédure et les conserver à 4 oC jusqu'à l'utilisation. - Préparer les enzymes de digestion en pesant la quantité correcte de trypsine et de collagène à 50 ml de tubes coniques (tableau 3).
2. Dissection animale et préparation de la suspension des cellules germinales
REMARQUE: Cela prend environ 1 h pour terminer.
- Sacrifiez une souris mâle adulte (âgée de 7 semaines ou plus, testicule pesant 80 à 120 mg selon la souche et l'âge) par luxation cervicale ou asphyxie au CO2.
- Vaporiser l'abdomen ventral de la souris avec 70% d'éthanol. Ouvrez la cavité abdominopelvic à l'aide de ciseaux, en faisant une ouverture en forme de V.
- Tirer sur le tampon de graisse épididymal avec des forceps, localiser les testicules et les enlever avec des ciseaux. Évitez de déranger la tunica albuginea. Déposer les testicules sur un plat Petri de 6 cm contenant 1x Krebs.
- Décapuiller les testicules et jeter la tunica albuginea. Disperser légèrement les tubules séminifères en les taquinant doucement à l'écart avec des forceps.
- Utiliser des forceps pour transférer les tubules sémiifères dans un tube conique de 50 ml contenant 2 ml de solution de collagène fraîchement préparée (tableau 3).
- Incuber les tubules dans le bain d'eau de 37 oC pendant 3 min. Agiter doucement en berçant le tube.
REMARQUE: Les tubules flottants devraient se produire dans les 3 minutes en raison de l'enlèvement des cellules interstitielles. La température physiologique des cellules testiculaires est de 34 oC; par conséquent, de longues digestions sont généralement faites à cette température. Cependant, la courte digestion de 3 minutes peut être effectuée à 37 oC (température recommandée par le fabricant). Notez que si l'on utilise 34 oC, le temps de digestion doit être réoptimisé. - Ajouter au moins 40 ml de Krebs chaud 1x et laisser les tubules dans les sédiments (1 min) à température ambiante (RT). Retirer le supernatant et répéter 1x.
- Ajouter 25 ml de solution de trypsine fraîchement préparée(tableau 3),placer le tube sur le rotateur du tube à l'intérieur de l'incubateur de 34 oC, et couver pendant 15 à 20 min (15 tr/min). Vérifier sporadiquement l'état des tubules. Une fois que la solution devient trouble et qu'il ne reste que de petits morceaux de tubules, placez le tube sur la glace et passez immédiatement à l'étape suivante.
REMARQUE: Pour éviter la surdigestion et la lyse cellulaire, passez rapidement aux étapes de lavage suivantes. Certains protocoles comprennent la désactivation de la trypsine par sérum bovin fœtal (FBS). Dans ce protocole, le traitement FBS est omis, et au lieu de cela, la trypsine est enlevée par centrifugation immédiate et lavages subséquents avec le 1x Krebs froid. - Filtrer la solution à l'intermédiaire d'une passoire cellulaire de 40 m dans un nouveau tube conique de 50 ml sur la glace.
- Centrifugeuse 600 x g pendant 5 min à 4 oC pour granuler les cellules.
REMARQUE: Centrifugation avec une force trop forte peut nuire aux cellules. - Retirez le supernatant en le versant soigneusement.
- Appuyez sur la pastille cellulaire pour resuspendre les cellules dans le reste de 1x Krebs.
- Ajouter au moins 40 ml de 1x Krebs froid aux cellules resuspension.
- Répétez les étapes 2.10 et 2.11.
- Appuyez sur le tube avec le granule cellulaire pour resuspendre les cellules. Ajouter 1 ml de 0,5% de BSA dans 1x Krebs et avec une pointe de pipette coupée resuspendre les cellules en pipetting la solution de haut en bas. Évitez de faire des bulles.
- Enfin, ajouter 1 3 ml de 0,5 % de BSA en 1x Krebs afin que le volume final soit de 3 ml. Filtrer la suspension des cellules germinales à travers une passoire cellulaire de 40 m et procéder immédiatement au chargement des cellules sur le gradient BSA.
3. Séparation des cellules germinales par le gradient BSA discontinu
REMARQUE: Cette section prend environ 2 h à remplir. Commencez à préparer le gradient BSA discontinu pendant les étapes de lavage (étapes 2.10-2.14) pour charger les cellules dès que le prétraitement est prêt.
- Accommoder un tube de 50 ml verticalement sur la glace afin qu'il soit possible de voir un côté du tube. Vous pouvez également exécuter le protocole dans une chambre froide à 4 oC, auquel cas aucune glace n'est nécessaire.
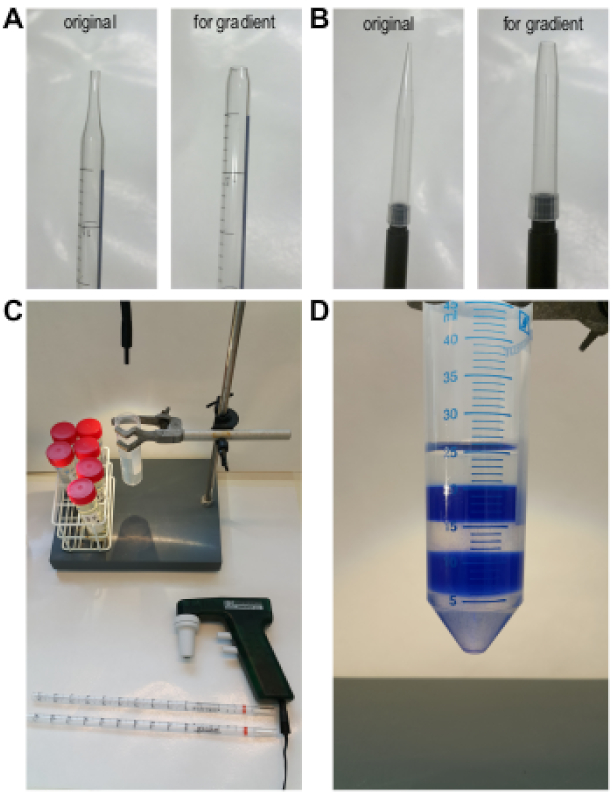
- Couper la pointe d'une pipette sérologique de 10 ml à environ 5 à 10 mm de la pointe pour obtenir une plus grande ouverture (figure 1A),et la monter sur le contrôleur de pipette (Figure 1C).
REMARQUE: Cela permettra de réduire la vitesse pendant le tuyauterie, ce qui facilite l'empilement des différentes solutions BSA. Vous pouvez également utiliser une pipette mécanique de 1 ml avec un piston lisse et couper les pointes de pipette avec une ouverture d'environ 3 mm de diamètre (Figure 1B). - Commencez par pipetting 5 mL de 5% de solution BSA au fond du tube de 50 ml.
- Lentement pipet 5 mL de 4% de solution BSA sur le dessus de la solution de 5%. Commencez par toucher doucement la surface de la solution de 5 % avec la pointe coupée de la pipette et superposez les deux solutions tout en maintenant soigneusement le contact avec la surface, en veillant à ne pas laisser la pointe de la tuyauterie s'immerger.
REMARQUE: Une ligne claire doit être visible entre les deux couches à la fin de cette étape. - Répéter l'étape 3.4 avec les autres solutions BSA obtenir un gradient discontinu de 5% à 1% (Figure 1D).
- Chargez soigneusement la suspension unicellulaire sur le dessus du gradient sans déranger. Laisser les cellules sédimenter à travers le gradient pendant 1,5 h à 4 oC ou sur la glace.
4. Collection de fractions de cellules germinales enrichies
REMARQUE: Cette section prend environ 30 minutes à compléter.
- Monter une pointe de pipette de 1 ml sur une pipette mécanique de 1 ml et couper la pointe de sorte que l'ouverture soit de 3 mm de diamètre(figure 1B).
- Recueillir soigneusement 1 ml de fractions dans des tubes séparés de 1,5 ml, à partir du haut du gradient BSA, et les stocker sur de la glace. Numéroter les tubes dans le même ordre que les fractions seront collectées.
REMARQUE: À ce stade, il devrait y avoir une série de 28 tubes contenant des suspensions cellulaires. Utilisez une pipette mécanique de 1 ml avec un piston lisse et coupez les pointes de pipette pour minimiser le risque de perturber le gradient BSA. - Centrifuger les fractions de cellules germinales à 600 x g pendant 10 min à 4 oC.
- Attention à ne pas déranger la pastille, jetez la plupart du supernatant et resuspendre la pastille cellulaire en faisant glisser le tube. Ajouter 1 ml de tampon de 1x Krebs à glace à chaque tube et répéter la centrifugation.
REMARQUE: Répétez l'étape de lavage si BSA interfère avec l'analyse en aval. - Jeter la plupart du supernatant, mais laisser 100 euros l après le lavage final. Resuspendre soigneusement la pastille cellulaire.
REMARQUE: Le fait de suspendre soigneusement les cellules garantit que l'échantillon prélevé dans cette solution représente toutes les cellules de la fraction donnée. Gardez les suspensions cellulaires sur la glace pendant la préparation des glissières.
5. Analyse des fractions cellulaires
REMARQUE: L'analyse prend 2 h à compléter.
- Un à la fois, pipet 20 'L de 4% de paraformaldéhyde (PFA) à l'intérieur de chaque anneau de stylo à graisse sur une diapositive de microscopie numérotée.
- Ajouter immédiatement 2 ll de la suspension cellulaire suspendue de la fraction correspondante. Répéter l'opération pour chaque fraction.
REMARQUE: Pendant la préparation des toboggans, gardez toutes les suspensions cellulaires sur la glace. - Séchez les toboggans à RT pendant 1 h jusqu'à la nuit (O/N).
REMARQUE: Au cours de cette étape, après avoir prélevé un échantillon de chaque fraction, les cellules peuvent être traitées pour des analyses en aval ou de stockage (section 6). - Rincer les diapositives une fois avec 1x PBS et monter avec le support de montage avec 4,6-diamidino-2-phenylindole (DAPI) (Tableau des matériaux).
- Analysez les diapositives sous un microscope à fluorescence pour estimer quel type particulier de cellules germinales est enrichi dans chaque fraction.
REMARQUE: Différents types de cellules peuvent être reconnus par leur morphologie nucléaire caractéristique visualisée par la coloration DAPI. Figure 2 A montre des fractions représentatives enrichies en spermatides allongés, spermatides ronds et spermatocytes pachytenés. Le noyau spermatide rond est petit (6 à 7 m de diamètre) et rond (Figure 2A). Les noyaux spermatidés ronds de souris sont également caractérisés par une structure unique et ronde d'hétérochromatine appelée le chromocenter. Les noyaux des spermatides allongés ont une forme allongée typique et une taille nucléaire réduite en raison de la condensation de chromatine (Figure 2A). Les noyaux de spermatocyte satéthyten sont beaucoup plus gros (diamètre de plus de 10 m) et de forme plus irrégulière, avec des zones de chromatine densément emballée réparties partout(figure 2A). -
Si nécessaire, effectuer immunostaining en plus de la coloration DAPI pour soutenir la reconnaissance des différents types de cellules.
- Dans ce cas, étalez 2 ll de suspension cellulaire avec 20 l d'une solution de fixation (4% PFA et 0,1% de surfactant nonionique en PBS) à l'intérieur d'un anneau de stylo à graisse. Après le séchage des diapositives, postfixer les cellules avec 4% PFA pendant 10 min à RT.
- Rincer avec PBS, puis perméabilize avec 0,1% de surfactant nonionic en PBS pendant 5 min. Rincer avec PBS et désactiver tout PFA restant en couve des diapositives dans 100 mM NH4Cl pendant 5 min.
- Rincer avec PBS et procéder à un protocole standard d'immunofluorescence consistant en le blocage et l'incubation avec des anticorps primaires spécifiques et des anticorps secondaires fluorochrome-étiquetés. Montez les toboggans avec un moniant antifade avec DAPI (Table of Materials) et laissez-les régler pendant 24 h.
REMARQUE: Des exemples de marqueurs utiles pour les incubations d'anticorps primaires comprennent l'agglutinine d'arachide (PNA; 1:2,000 dans la solution de blocage) qui tache l'acrosome dans les spermatides ronds et allongeants, anticorps anti-DDX4 (1:200 dans la solution de blocage) qui étiquette un seul granule cytoplasmique dans les spermatides ronds, et anticorps anti-H2AX (1:100 dans la solution de blocage) qui reconnaît les corps sexuels dans les spermatocytes de pachytene. Voir les résultats représentatifs et la figure 2C,D pour des exemples d'analyse d'immunofluorescence des fractions.
6. Traitement des échantillons restants pour le stockage
REMARQUE: La section 6 prend environ 20 minutes.
- Une fois qu'un échantillon de chaque fraction a été prélevé pour une diapositive de microscopie, ajouter 1 ml de 1x Krebs glacé à chaque fraction et centrifuger les cellules vers le bas à 600-13 000 x g pendant 10 min à 4 oC.
REMARQUE: Une centrifugation à basse vitesse entraîne une pastille lâche, ce qui rend difficile d'enlever complètement 1x Krebs, tandis qu'une centrifugation à grande vitesse peut nuire aux cellules. Choisissez la vitesse de centrifugation selon les exigences spécifiques du protocole en aval. - Retirez et jetez le supernatant et continuez avec l'analyse en aval préférée.
REMARQUE: À ce stade, les cellules peuvent être congelées dans de l'azote liquide et stockées à -70 oC. Une fois familiarisé avec cette technique, il est possible de continuer avec le protocole en aval de préférence directement, mais il est toujours conseillé de prendre un échantillon et d'assurer la pureté de chaque fraction. L'ARN total peut être extrait, quantifié et analysé par des méthodes telles que la réaction en chaîne de polymérase de transcription inversée (PCR) et le séquençage d'ARN. Des extraits totaux de protéines peuvent également être préparés à partir desquels l'immunoprécipitation ou l'analyse des ballonnements occidentaux peut être effectuée (figure 3).
Résultats
L'enrichissement suffisant d'un type de cellule germinale est généralement considéré comme supérieur à 80 %6. Le protocole MDR fonctionne particulièrement bien pour enrichir les spermatides ronds. Un nombre élevé de spermatides ronds purs de 'gt;90% peut être systématiquement obtenu en utilisant cette technique. Les fractions optimales des spermatozoïdes pachytenés et des spermatides allongés sont enrichies à 75 % et 80 %, respectivement. Les spermatides allongés ont tendance à rester au-dessus du gradient et sont recueillis avec la première fraction. Dans l'exemple montré, la fraction 1 contenait 80 % des spermatides allongés (figure 2B). La plupart des spermatides allongés obtenus avec cette technique ont condensé des noyaux, tandis que les spermatides allongés précoces non condensés sont rares (Figure 2A). Les fractions suivantes contiennent des spermatides ronds enrichis. Dans l'exemple, l'enrichissement des spermatides ronds était de plus de 90 % en fractions 2, 3 et 4, et l'enrichissement supérieur à 80 % a été observé au total en sept fractions (2 à 8) (figure 2B). En raison de leur grande taille, les spermatocytes de pachytene sédimentent plus rapidement et sont recueillis en dernier. Dans l'exemple de purification, l'enrichissement était d'environ 75 % en fractions 14 et 15(figure 2B).
Tandis que la morphologie nucléaire, visualisée par la coloration de DAPI, suffit habituellement pour la reconnaissance des cellules, l'analyse d'immunofluorescence peut être exécutée pour soutenir l'analyse. PNA tache les acrosomes en développement de spermatides ronds et allongés, et l'aspect acrosomique peut être exploité pour classer davantage les spermatides ronds dans les étapes 1-8 de la différenciation10. Distinguer les spermatides allongés et ronds teintés de PNA repose sur les différences dans leur forme nucléaire(figure 2C, panneau gauche). L'anticorps anti-DDX4 est un marqueur utile pour les fractions spermatides rondes puisqu'il visualise un seul granule périnucléaire DDX4-positif, le corps chromatoïde (CB), dans le cytoplasme de chaque spermatide rond. Cette coloration spécifique au CB est facile à distinguer de la coloration cytoplasmique plus largement distribuée dans les spermatocytes(Figure 2C, panneau moyen). Les spermatocytes de pachytene peuvent être reconnus par l'anticorps anti-H2AX qui étiquette le corps sexuel nucléaire apparaissant spécifiquement à la phase de pachytene de la première division méiotique (figure 2C, panneau droit). Dans cette purification, la coloration PNA a révélé que les fractions spermatées rondes enrichies de MDR contenaient un mélange de spermatides ronds à diverses étapes de différenciation, toutes contenant leur structure de signature, le CB DDX4-positif (Figure 2D, fraction RS). Les spermatozoïdes de pachytene d'enrichissement en fraction 16(figure 2D , fraction PSpc) ont validé les spermatozoïdes de pachytene d'enrichissement en fraction 16 ( figure 2D, fraction PSpc).
Le comptage cellulaire a révélé que le nombre de cellules dans les fractions rondes de spermatocytes spermatid et pachytené s'est adapté à diverses analyses en aval. Les fractions spermatées rondes (5 à 8) contenaient chacune environ 2,5 x 106 cellules. Par conséquent, en mettant en commun ces fractions, il est possible d'obtenir plus de 10 millions de cellules. Les fractions de spermatocyte de Pachytene (14 et 15) contenaient habituellement un peu moins de cellules. Dans cet isolement, environ 1,5 à 2,0 x 106 cellules par fraction ont été comptées. La première fraction contenait 0,75 x 106 spermatides allongés.
Comme le montre la figure 3A, la quantité totale d'ARN obtenue à partir de la majorité des fractions variait de 0,5 à 2,5 g, ce qui est suffisant pour les analyses d'ARN en aval telles que la transcription inversée PCR, le séquençage de l'ARN ou la visualisation de l'ARN sur un gel. La quantité de protéines obtenues à partir de chaque fraction varie généralement de 20 à 140 g (figure 3B), ce qui est suffisant pour plusieurs taches occidentales. Des lysates entiers de cellules ont été préparés à partir des fractions rassemblées, et l'analyse occidentale de tache a été exécutée utilisant des anticorps détectant DDX4, PIWIL1, et PIWIL2, qui sont tous fortement exprimés dans les spermatocytes de pachytene et les spermatides ronds aussi bien que l'omniprésent déshydrogénase exprimée de glycéraldéhyde 3-phosphate (GAPDH).
Dans ce protocole, 10 % des lysates protéiques dérivés de fractions uniques étaient suffisants pour détecter clairement toutes ces protéines sur un réglage standard de tache occidentale(figure 3C). La quantité de protéines dans une fraction s'est également avérée suffisante pour l'immunoprécipitation à l'aide d'un anticorps contre PIWIL1, ainsi que pour la détection de PIWIL2 co-immunopicié(figure 3D). En outre, ce protocole a été utilisé avec succès pour obtenir des fractions enrichies de spermatocytes pachytenés et de spermatides ronds à partir de souris témoins et génétiquement modifiées pour des applications en aval telles que la transcription inverse quantitative PCR11 et le séquençage de l'ARN à haut débit12.

Figure 1 : Préparation d'un gradient BSA discontinu. (A) Une pointe de pipette sérologique de 5 ml pour la préparation du gradient. (B) Une pointe de pipette de 1 ml pour la préparation du gradient et de la collecte des fractions des cellules germinales après la sédimentation. (C) L'équipement nécessaire à la préparation du gradient. (D) Vue latérale du gradient de densité BSA discontinu du 5 % (en bas) à la solution BSA de 1 % (en haut) ; les 2% et 4% BSA solutions sont de couleur bleue pour une meilleure visualisation. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 2 : Images représentatives des fractions cellulaires collectées et de l'analyse de l'enrichissement. (A) Cellules testiculaires tachées de DAPI. La rangée supérieure montre les cellules testiculaires dans l'épithélium séminifère intact d'une section de testis fixée par la paraffine (à gauche) ou en suspension (à droite). La rangée inférieure montre des fractions de spermatides allongés enrichis (ES), de spermatides ronds (RS) ou de spermatocytes pachytenés (PSpc). Barres d'échelle de 20 m pour la rangée supérieure et de 10 m pour les en-ttes de la rangée inférieure. (B) Quantification relative des différents types de cellules germinales dans chaque fraction. Les cellules ont été comptées manuellement à l'aide du logiciel ImageJ et classées en cellules RS, ES, PSpc, Sertoli et autres. Les nombres de fractions et les pourcentages respectifs de BSA dans le gradient sont indiqués sur l'axe X. (C) Analyse d'immunofluorescence des cellules testiculaires. Panneau gauche : PNA étiqueté rhodamine tache l'acrosome dans les deux ES (flèche jaune) et RS (flèche blanche). Panneau moyen : L'anticorps DDX4 tache un seul granule périnucléaire en RS, ce qui est facile à distinguer du signal cytoplasmique plus diffus dans les spermatocytes (flèche bleue). Aucun signal DDX4 n'est détecté dans ES (flèche orange). Panneau droit : L'anticorps H2AX reconnaît le corps sexuel nucléaire caractéristique présent uniquement dans les spermatocytes de pachytene (cellules négatives de H2AX marquées par un astérisque blanc). Des barres d'échelle de 10 m. (D) fractions de cellules enrichies mDR ont été étiquetées avec PNA (fraction RS), anti-DDX4 (fraction RS) et anti-H2AX (fraction PSpc) pour valider davantage l'enrichissement cellulaire dans chaque fraction. Barres d'échelle de 10 m. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 3 : Analyses en aval après enrichissement cellulaire MDR. (A) l'ARN a été extrait de chaque fraction après un enrichissement représentatif de cellules de MDR et quantifié. Les nombres de fractions et les pourcentages respectifs de BSA dans le gradient sont indiqués sur l'axe X. (B) Les protéines ont été extraites de chaque fraction en lysant la pastille cellulaire dans le tampon d'analyse de radioimmunoprécipitation (RIPA) puis quantifiées. (C) Des extraits entiers de protéine de cellules ont été préparés et analysés par le ballonnement occidental utilisant des anticorps contre DDX4, PIWIL1, PIWIL2, et GAPDH. 10 % du lysate était chargé à partir de chaque fraction indiquée. (D) L'immunoprécipitation a été effectuée à partir de fractions indiquées à l'aide d'anticorps anti-PIWIL1, suivis de ballonnements occidentaux avec des anticorps anti-PIWIL1 et anti-PIWIL2. L'échantillon d'entrée comprend un mélange de lysates protéiques provenant de différentes fractions, et l'immunoprécipitation de contrôle (IP) à l'aide du lapin IgG a également été effectuée à partir d'un lysate mixte. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 4 : Représentation schématique du protocole MDR et temps requis pour terminer chaque étape. Le matériel de départ est composé de deux testicules d'une souris adulte. Le nombre moyen de cellules et la quantité d'ARN et de protéines obtenues à partir d'une fraction sont indiqués. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}
| tampon | Réactifs | préparation | espace de stockage |
| Krebs tampon (10x) | 3,26 g KH2PO4 | Porter à 2 L avec H2O, filtrer 0,22 m et autoclave. | Peut être stocké à 4 oC pendant plusieurs mois |
| 139,5 g NaCl | |||
| 5,89 g MgSO4 '7H2O | |||
| 50 g de dextrose | |||
| 3,78 g CaCl2-2H2O | |||
| 7,12 g KCl | |||
| Krebs tampon (1x) | 4,24 g NaHCO3 | Dissoudre NaHCO3 à 100 ml de H2O, ajouter 200 ml de tampon De 10x Krebs et porter à 2 L avec H2O. | Pour être préparé frais |
| 200 mL 10x Krebs tampon |
Tableau 1 : Préparation du tampon Krebs.
| Concentration BSA (w/v) | 10% solution BSA | 1x Tampon Krebs |
| 0.50% | 0,5 ml | 9,5 ml |
| 1% | 1 mL | 9 mL |
| 2% | 2 mL | 8 mL |
| 3% | 3 mL | 7 mL |
| 4% | 4 mL | 6 mL |
| 5% | 5 mL | 5 mL |
Tableau 2 : Préparation des solutions BSA.
| Solution de digestion | Concentration de travail | Réactifs | préparation |
| Collagènase | 1 mg/ml | Collagène IV | Peser 2 mg de collagène IV à un tube conique de 50 ml, et ajouter 2 ml de tampon chaud de 1x Krebs juste avant la digestion (étape 2.3). |
| Trypsine | 0,6 mg/mL | Trypsine | Peser 15 mg de trypsine à un tube conique de 50 ml, et ajouter 25 ml de tampon chaud 1x KREBS et 40 oL de DNase I juste avant la digestion (étape 2.6). |
| 3,2 ku/mL | DNase I |
Tableau 3 : Préparation des enzymes de digestion.
Discussion
Présenté ici est un protocole simple et peu coûteux pour isoler les populations enrichies de spermatides ronds, spermatocytes pachytenés, et spermatids allongeant à l'aide d'équipement de laboratoire standard (aperçu du protocole montré à la figure 4). Bien qu'aucune expertise ou machinerie coûteuse ne soit requise, il y a quelques étapes critiques qui doivent être considérées pendant la digestion de tissu, la construction du gradient, et le chargement de la suspension de cellules sur le gradient.
Les cellules germinales sont libérées des tubules séminifères par deux digestions enzymatiques consécutives. La première digestion avec la collagène IV sépare les tubules séminifères en enlevant les cellules interstitielles. Un temps de digestion prolongé peut endommager les tubules et entraîner la perte de spermatides, car (s'ils sont libérés des tubules au cours de cette étape), ils seront jetés dans les étapes suivantes. La deuxième étape de digestion avec la trypsine libère des cellules germinales des tubules séminifères. Il peut y avoir une lyse cellulaire occasionnelle et généralement certaines amas se forment en raison de l'ADN génomique libéré. Il n'est pas conseillé de dépasser la durée ou la température suggérée de la digestion, car cela peut entraîner une viabilité plus faible, une lyse cellulaire accrue et un agglutination. Si l'agglutination légère se produit, les amas peuvent être ignorés. Cependant, dans les cas d'agglutination significative et de perte de cellules, le temps ou la concentration de digestion de trypsine devrait être réduit. Il convient également de noter que l'activité enzymatique de la trypsine peut varier d'un lot à l'autre et pendant des périodes prolongées de stockage. La quantité de DNase I pendant la digestion de trypsine peut également être augmentée pour enlever les amas excédentaires, mais ceci devrait être considéré une solution secondaire. Il est important d'obtenir une suspension homogène à une seule cellule à la fin du prétraitement, car les cellules agglomérées se sédimenteront plus rapidement, contaminant les fractions et perturbant le gradient.
La construction du gradient peut nécessiter une certaine pratique. S'il y a un inconfort à l'aide d'une pointe de pipette de 5 ml avec un contrôleur de pipette, il est recommandé d'utiliser une pipette mécanique normale de 1 ml avec un piston lisse, puis couper les pointes de pipette à une ouverture de 3 mm de diamètre (Figure 1B). Une ouverture plus large et un chargement fluide des solutions BSA diminueront le risque de mélanger le gradient. Une fois correctement préparé, il est possible de voir les limites entre les solutions BSA adjacentes en raison de leurs différents indices de réfraction. Le gradient doit être produit directement avant l'utilisation. Il convient également de noter que toute petite secousse ou vibration peut perturber le gradient, de sorte que le gradient doit être réglé dans un environnement où il ne sera pas dérangé.
Le chargement de la suspension cellulaire sur le gradient doit être fait très soigneusement. Après le chargement, la suspension cellulaire doit rester au-dessus du gradient, à partir de laquelle les cellules vont lentement commencer à sédiments à travers la première couche. Si de grands groupes de cellules sont vus se déplaçant rapidement à travers le gradient, les cellules n'ont probablement pas été suspendues soigneusement ou il ya un excès d'agglutination. Si les cellules ne restent pas au sommet du gradient BSA discontinu au chargement mais descendent immédiatement entre les couches BSA de 1 % et 2 % (étape 3.6), la suspension cellulaire est probablement trop dense. Ce protocole a été optimisé à l'aide de deux testicules d'une souris adulte (80-120 mg/testis) comme matériau de départ; bien que, des isolements réussis utilisant des quantités réduites de matériel de démarrage aient été exécutés. Pour mettre à niveau le protocole et obtenir des cellules germinales plus enrichies à partir d'un plus grand nombre de testicules, plus de tubes de 50 ml avec le gradient devraient être introduits.
Le protocole a été initialement développé et optimisé pour enrichir les spermatides ronds haploïdes des testicules adultes de souris, et la pureté des fractions spermatées rondes devrait être plus de 90%. En plus des fractions spermatides rondes très pures, des résultats satisfaisants pour l'enrichissement des spermatocytes de pachytene et des spermatides allongeantont ont été obtenus. Il convient de noter que les érythrocytes peuvent contaminer les fractions spermatées allongées, et d'autres mesures pour les éliminer devraient être prises si leur présence est censée interférer avec les analyses en aval. Nous n'avons pas été en mesure d'enrichir d'autres types de cellules telles que les cellules méiotiques préméidiques ou précoces (avant la phase de pachyène) à partir de souris adultes utilisant le protocole MDR.
En outre, la sédimentation STA-PUT a été utilisée avec succès pour obtenir des fractions enrichies de spermatogonie ou de préleptotene, de leptotene et de spermatocytes zygotene à l'aide de testicules juvéniles recueillis à des moments donnés après la naissance13. Cette approche tire parti de l'apparition de ces types de cellules lors de la première vague de spermatogénèse. La même approche peut probablement être appliquée à l'enrichissement du MDR, mais elle n'a pas encore été testée dans la pratique. Une autre méthode qui est une bonne option pour la purification des cellules préméidistes et méiotiques à des stades spécifiques de différenciation est FACS, qui a l'avantage important de permettre l'isolement de types de cellules spécifiques en fonction de la présence des marqueurs spécifiques14 ,15,16,17.
Dans l'ensemble, la sédimentation de la vitesse du MDR sert de méthode utile pour l'enrichissement des cellules germinales. Bien que cette méthode ne soit pas supérieure à d'autres méthodes bien établies en termes de pureté ou de quantité de cellules enrichies, ses avantages évidents sont sa simplicité et ses faibles coûts d'établissement. Ceci, avec la faible quantité de matériaux de départ requis, font de ce protocole une excellente option pour les chercheurs dans le domaine de la spermatogenèse et ceux dans d'autres domaines qui peuvent ne pas vouloir investir dans du matériel spécialisé ou de grands groupes d'animaux.
Déclarations de divulgation
Les auteurs n'ont rien à révéler.
Remerciements
Nous tenons à remercier tous les membres du laboratoire Kotaja pour leur contribution à l'élaboration du protocole, ainsi que pour l'utilisation active et l'essai du protocole dans leurs projets de recherche. Nous apprécions particulièrement la contribution de Jan Lindstram pour l'aide à l'optimisation du protocole. Cette étude a été soutenue par l'Académie de Finlande, la Fondation Sigrid Jusélius et le Programme doctoral de médecine moléculaire de Turku.
matériels
| Name | Company | Catalog Number | Comments |
| 4% Paraformaldehyde | Preference of researcher | ||
| AlexaFluor488 donkey anti-rabbit IgG | Thermo Fisher Scientific | A-21206 | |
| AlexaFluor647 donkey anti-mouse IgG | Thermo Fisher Scientific | A-31571 | |
| Bovine Serum Albumin (BSA) | Sigma | A9647 | |
| CaCl2·2H2O | Preference of researcher | ||
| Collagenase IV | Sigma | C5138 | |
| Complete protease inhibitor cocktail | Roche | 11836145001 | |
| DDX4 antibody | Abcam | ab13840 | |
| Dextrose | Preference of researcher | ||
| DNase I | Worthington | LS006355 | |
| GAPDH | HyTest | 5G4 | |
| HRP-linked anti-mouse IgG | GE Healthcare Life Sciences | NA931 | |
| HRP-linked anti-rabbit IgG | GE Healthcare Life Sciences | NA934 | |
| KCl | Preference of researcher | ||
| KH2PO4 | Preference of researcher | ||
| MgSO4·7H2O | Preference of researcher | ||
| NaCl | Preference of researcher | ||
| NaHCO3 | Preference of researcher | ||
| NH4Cl | Preference of researcher | ||
| Pierce BCA protein assay kit | Life Technologies | 23227 | |
| PIWIL1 | Cell Signaling Technology | G82 | |
| PIWIL2, clone 13E-3 | Millipore | MABE363 | |
| Prolong Diamond Antidafe Mountant with DAPI | Thermo Fisher Scientific | P36962 | for Alexa Fluor immunostainings |
| Rabbit IgG | Neomarkers | NC-100-P | |
| Rhodamine-labelled Peanut agglutinin (PNA) | Vector Laboratories | RL-1072 | |
| RIPA buffer | 50 mM Tris-HCl, pH 7.5, 1% NP-40, 0.5% w/v sodium deoxycholate, 0.05% w/v SDS, 1 mM EDTA, 150 mM NaCl, 1x protease inhibitor cocktail, 0.2 mM PMSF and 1 mM DTT | ||
| TRIsure | Bioline | BIO-38033 | |
| Triton X-100 | Preference of researcher | nonionic surfactant | |
| Trypsin | Worthington | LS003703 | |
| VECTASHIELD Antifade Mounting Medium with DAPI | Vector Laboratories | H-1200 | for standard DAPI analysis of cell fractions |
| γH2AX antibody | Millipore | 05-636 | |
| 0.22 µm filter | Sartorius | Sartolab BT 180C6 | or equivalent |
| 1 mL mechanical pipette | Preference of researcher | ||
| 1.5 mL or 2 mL tubes | Preference of researcher | ||
| 40 µm cell sieves for 50 mL tubes | Greiner Bio-One | 542040 | or equivalent cell strainer |
| 5 mL serological pipettes | Sarstedt | 86.1254.001 | or equivalent |
| 50 mL conical tubes | Preference of researcher | ||
| 6 cm Petri dishes | Preference of researcher | ||
| Cell culture incubator | Preference of researcher | ||
| Centrifuge for 50 mL tubes | Preference of researcher | ||
| Grease pen for microscopy glass slides | Preference of researcher | ||
| HulaMixer Sample Mixer | Thermo Fisher Scientific | 15920D | or equivalent cell rotator |
| Microdissection forcepts | Preference of researcher | ||
| Microdissection scissors | Preference of researcher | ||
| Microscopy glass slides and coverslips | Preference of researcher | ||
| Nanodrop 1000 | Thermo Scientific | ||
| Pipetboy Acu 2 | Integra | 155 000 | or equivalent pipette controller |
| Refrigerated centrifuge for 1.5 mL tubes | Preference of researcher | ||
| Tips for 1 mL mechanical pipette | Preference of researcher | ||
| Water bath | Preference of researcher | ||
| Widefield fluorescence microscope | Preference of researcher |
Références
- Hess, R. A., Renato de Franca, L. Spermatogenesis and cycle of the seminiferous epithelium. Advances in Experimental Medicine and Biology. 636, 1-15 (2008).
- Lehtiniemi, T., Kotaja, N. Germ granule-mediated RNA regulation in male germ cells. Reproduction. 155 (2), R77-R91 (2018).
- Lam, D. M., Furrer, R., Bruce, W. R. The separation, physical characterization, and differentiation kinetics of spermatogonial cells of the mouse. Proceedings of the National Academy of Sciences of the United States of America. 65 (1), 192-199 (1970).
- Meistrich, M. L., Bruce, W. R., Clermont, Y. Cellular composition of fractions of mouse testis cells following velocity sedimentation separation. Experimental Cell Research. 79 (1), 213-227 (1973).
- La Salle, S., Sun, F., Handel, M. A. Isolation and short-term culture of mouse spermatocytes for analysis of meiosis. Methods in Molecular Biology (Clifton, N.J.). 558, 279-297 (2009).
- Bryant, J. M., Meyer-Ficca, M. L., Dang, V. M., Berger, S. L., Meyer, R. G. Separation of spermatogenic cell types using STA-PUT velocity sedimentation. Journal of Visualized Experiments. (80), e50648(2013).
- Grabske, R. J., Lake, S., Gledhill, B. L., Meistrich, M. L. Centrifugal elutriation: separation of spermatogenic cells on the basis of sedimentation velocity. Journal of Cellular Physiology. 86 (1), 177-189 (1975).
- Barchi, M., Geremia, R., Magliozzi, R., Bianchi, E. Isolation and analyses of enriched populations of male mouse germ cells by sedimentation velocity: the centrifugal elutriation. Methods in Molecular Biology (Clifton, N.J.). 558, 299-321 (2009).
- Romrell, L. J., Bellvé, A. R., Fawcett, D. W. Separation of mouse spermatogenic cells by sedimentation velocity. A morphological characterization. Developmental Biology. 49 (1), 119-131 (1976).
- Kotaja, N., et al. Preparation, isolation and characterization of stage-specific spermatogenic cells for cellular and molecular analysis. Nature Methods. 1 (3), 249-254 (2004).
- Korhonen, H. M., et al. DICER Regulates the Formation and Maintenance of Cell-Cell Junctions in the Mouse Seminiferous Epithelium. Biology of Reproduction. 93 (6), 139(2015).
- Da Ros, M., et al. FYCO1 and autophagy control the integrity of the haploid male germ cell-specific RNP granules. Autophagy. 13 (2), 302-321 (2017).
- Bellvé, A. R., et al. Spermatogenic cells of the prepuberal mouse. Isolation and morphological characterization. The Journal of Cell Biology. 74 (1), 68-85 (1977).
- Mays-Hoopes, L. L., Bolen, J., Riggs, A. D., Singer-Sam, J. Preparation of spermatogonia, spermatocytes, and round spermatids for analysis of gene expression using fluorescence-activated cell sorting. Biology of Reproduction. 53 (5), 1003-1011 (1995).
- Suter, L., Koch, E., Bechter, R., Bobadilla, M. Three-parameter flow cytometric analysis of rat spermatogenesis. Cytometry. 27 (2), 161-168 (1997).
- Lima, A. C., et al. Multispecies Purification of Testicular Germ Cells. Biology of Reproduction. 95 (4), 85-85 (2016).
- Bastos, H., et al. Flow cytometric characterization of viable meiotic and postmeiotic cells by Hoechst 33342 in mouse spermatogenesis. Cytometry. Part A: the Journal of the International Society for Analytical Cytology. 65 (1), 40-49 (2005).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.