
Method Article
Microinjection de cicadelle de maïs, Peregrinus maidis, embryons pour l’édition du génome CRISPR / Cas9
Dans cet article
Résumé
Voici des protocoles pour la collecte et la microinjection d’embryons de cicadelles de maïs précellulaires dans le but de modifier leur génome via l’édition du génome basée sur CRISPR / Cas9 ou pour l’ajout d’éléments transposables marqués par transformation germinale.
Résumé
La cicadelle du maïs, Peregrinus maidis, est un ravageur du maïs et un vecteur de plusieurs virus du maïs. Des méthodes publiées précédemment décrivent le déclenchement de l’interférence ARN (ARNi) chez P. maidis par microinjection d’ARN double brin (ARNds) chez les nymphes et les adultes. Malgré la puissance de l’ARNi, les phénotypes générés par cette technique sont transitoires et manquent d’héritage mendélien à long terme. Par conséquent, la boîte à outils de P. maidis doit être élargie pour inclure des outils génomiques fonctionnels qui permettraient la production de souches mutantes stables, ouvrant la porte aux chercheurs pour apporter de nouvelles méthodes de lutte contre ce ravageur économiquement important. Cependant, contrairement aux ARNds utilisés pour l’ARNi, les composants utilisés dans l’édition du génome basée sur CRISPR / Cas9 et la transformation de la lignée germinale ne traversent pas facilement les membranes cellulaires. En conséquence, les ADN plasmidiques, les ARN et / ou les protéines doivent être microinjectés dans les embryons avant que l’embryon ne se cellularise, ce qui fait du moment de l’injection un facteur critique de succès. À cette fin, une méthode de ponte à base d’agarose a été mise au point pour permettre de récolter des embryons sur des femelles de P. maidis à des intervalles relativement courts. On y trouve des protocoles détaillés pour la collecte et la micro-injection d’embryons précellulaires de P. maidis avec des composants CRISPR (nucléase Cas9 qui a été complexée avec des ARN guides), et les résultats de l’élimination du gène Cas9 d’un gène de couleur des yeux de P. maidis , blanc, sont présentés. Bien que ces protocoles décrivent l’édition du génome CRISPR / Cas9 chez P. maidis, ils peuvent également être utilisés pour produire des P. maidis transgéniques via la transformation de la lignée germinale en modifiant simplement la composition de la solution injectable.
Introduction
La cicadelle du maïs, Peregrinus maidis, est un ravageur économiquement important du maïs 1,2,3. Ils causent des dommages physiques directs à la plante, à la fois en se nourrissant avec leurs pièces buccales perçantes-suceuses, et pendant la reproduction lorsqu’ils pondent leurs embryons directement dans les tissus végétaux 2,4. Malgré les multiples voies de dommages directs aux cultures, l’impact le plus important de ces insectes sur la santé des cultures est indirect, en agissant comme vecteur du virus de la mosaïque du maïs (MMV) et du virus de la bande du maïs 5,6. Le MMV est capable de se répliquer dans le corps de son vecteur P. maidis, ce qui permet au virus de persister chez les insectes individuels tout au long de leur vie, de sorte qu’ils peuvent continuer à propager le virus à de nouvelles plantes hôtes 7,8. Les méthodes les plus courantes pour lutter contre P. maidis, et donc les virus qu’elle vectère, sont les insecticides.
Malheureusement, la mauvaise gestion de ces produits a entraîné le développement d’une résistance chez l’organisme nuisible ciblé ainsi que la pollution de l’environnement9. Par conséquent, de nouvelles stratégies sont nécessaires pour réduire les pertes de récoltes dues à cette combinaison insecte/virus-ravageur. Des travaux antérieurs ont démontré que l’interférence ARN (ARNi) pourrait être une méthode de contrôle efficace pour P. maidis, car ils sont sensibles à la régulation négative de l’expression génique même lors de l’ingestion d’ARN double brin (ARNds)10. Cependant, le moyen le plus efficace d’administrer l’ARNds sur le terrain serait par les plantes dont les insectes se nourrissent; Par conséquent, les cultures pourraient toujours être sensibles aux virus que les insectes transportent déjà. Avec l’avènement de l’édition du génome CRISPR/Cas9, de nouvelles stratégies de lutte antiparasitaire sont possibles, y compris le forçage génétique11,12 basé sur Cas9, qui pourrait être utilisé pour réduire la taille d’une population de ravageurs, ou pour remplacer ladite population par des individus résistants aux virus qu’ils vecvent.
Cependant, le développement et le déploiement de tout type de système de forçage génétique nécessiteront le développement de techniques transgéniques. De telles méthodes n’étaient pas nécessaires pour effectuer des expériences d’ARNi chez P. maidis parce que les ARNds et/ou les siRNA sont présumés capables de traverser les membranes cellulaires en raison de l’efficacité de l’ARNi chez P. maidis10,13. Ce n’est pas vrai pour les ADN et / ou les protéines utilisés dans la transgénèse traditionnelle ou dans l’édition de gènes basée sur Cas9, qui seraient l’un ou l’autre précurseur de la création d’insectes porteurs d’un forçage génétique. Pour effectuer l’édition de gènes ou d’autres formes de transformation de la lignée germinale, ces ADN et protéines sont idéalement microinjectés dans les embryons au stade du blastoderme syncytial, avant la cellulasse de l’embryon de l’insecte. Le timing est critique, car le stade syncytial est la première partie du développement14,15. Comme les femelles de P. maidis pondent préférentiellement leurs œufs dans les tissus végétaux, l’extraction de quantités suffisantes d’embryons précellulaires pour les micro-injections peut nécessiter beaucoup de travail et de temps. Par conséquent, de nouvelles techniques ont été développées pour collecter et microinjecter rapidement des embryons de P. maidis avant la cellularisation.
Protocole
1. Élevage de P. maidis adultes au niveau des colonies
- Plantez un minimum de quatre pots de maïs par semaine par cage d’élevage, avec 3-4 graines par pot. Cultivez dans un environnement exempt d’insectes.
- Lorsque les plantes ont ~ 5 semaines, placez-les à l’intérieur d’une cage de 30 cm x 30 cm x 60 cm.
- Obtenir une quantité suffisante d’adultes de P. maidis (~500) d’un laboratoire de recherche ou dans la nature, et placez-les dans une cage à l’épreuve des insectes avec 9 à 12 plants de maïs (3-4 pots).
- Maintenir la colonie dans un incubateur d’élevage d’insectes à 25 °C (± 1 °C), avec au moins 70 % d’humidité et un cycle lumineux de 14:10.
- Pour générer une colonie calibrée selon l’âge, retirez tous les adultes initiaux après quatre jours de ponte et laissez les embryons pondus dans la cage éclore et vieillir naturellement.
- Déplacer les insectes P. maidis (adultes) âgés de 5 semaines vers des plants de maïs frais pour une sous-culture hebdomadaire en les recueillant à l’aide d’un aspirateur (figure 1). Ensuite, relâchez les adultes dans une cage propre avec des plants de maïs frais. Pour maintenir un approvisionnement régulier de jeunes adultes à des fins expérimentales, préparez chaque semaine de nouvelles cages calibrées selon l’âge.
- Arrosez les pots dans les cages deux fois par jour. Coupez périodiquement les tiges, enlevez le matériel végétal en décomposition et remplacez-les par des pots de maïs frais au besoin.
REMARQUE: Avec un entretien approprié, une colonie peut durer ~ 5 semaines (c’est-à-dire assez longtemps pour que les embryons pondus dans la cage deviennent adultes).
2. Chambre de ponte à base d’agarose
- Préparer des plats de collecte d’œufs (milieu de ponte) en versant 1 % p/v d’agarose dans de l’eau dans des boîtes de Petri propres de 100 mm x 15 mm. Conserver le milieu de ponte à 4 °C après sa solidification.
- Préparer une solution de saccharose à 10% p/v pour nourrir les adultes. Conservez la solution de saccharose à -20 °C jusqu’à un mois.
- Créez une chambre pour contenir les adultes en perçant un trou dans le fond d’une tasse de 1 oz (voir le tableau des matériaux) et en collant un écran sur le trou pour l’échange d’air (figure 2).
- Couper le film de cire de paraffine plastique en carrés de 5 cm x 5 cm; Réservez 2 carrés pour chaque tasse.
- Recueillir ~15 femelles adultes de 1 semaine dans une colonie de P. maidis calibrée selon l’âge. Pour sélectionner les femelles, examinez la face ventrale de l’abdomen et recherchez l’ovipositeur, qui est généralement plus foncé que le reste de l’abdomen (figure 3). Conservez les adultes jusqu’à une heure dans un flacon conique de 15 ml si vous installez plusieurs chambres de ponte. Refroidir brièvement les insectes sur la glace avant le sexage et les transférer dans le contenant pour adultes.
REMARQUE : Cet examen peut être effectué sans microscope. Les femelles adultes qui ont eu le temps de se nourrir et de s’accoupler ont aussi généralement un abdomen plus gros que les mâles adultes et sont plus dociles; Par conséquent, ils peuvent plus facilement être sélectionnés dans une population de cages. - Transférer les femelles dans un récipient pour adultes et sceller la tasse avec 1 couche de film de cire de paraffine plastique en l’étirant uniformément 3 à 4 fois sa taille d’origine (figure 4A,B).
- Appliquer 400 μL de solution de saccharose à 10 % p/v sur le dessus du joint de pellicule de cire de paraffine plastique et ajouter une deuxième couche de film de cire de paraffine plastique, en étirant le film de cire de paraffine plastique exactement comme ci-dessus (figure 4C,D).
REMARQUE: Le sandwich de film de cire de paraffine plastique étiré met sous pression la solution de saccharose, ce qui est très important pour l’alimentation des adultes, mais n’empêchera pas les femelles de percer leurs ovipositeurs jusque dans le milieu de ponte. - Placer la chambre adulte sur un plat de collecte d’œufs avec le côté du film de cire de paraffine plastique directement sur le milieu de ponte, et envelopper toute la chambre de ponte avec une pellicule de plastique sans couvrir les trous d’air, car ceux-ci sont nécessaires pour l’échange d’air (figure 5).
- Incuber chaque chambre de ponte à 25 °C avec 70% d’humidité et un cycle de lumière 14:10.
- Changez le sandwich de film de cire de paraffine plastique et de solution de saccharose à 10% p / v tous les jours, et retirez toute eau qui s’accumule à l’intérieur de la tasse.
3. Collecte et alignement des embryons dans un environnement très humide
- Mettre en place un système de micro-injection au stéréomicroscope dans un espace ou une hotte humidifié (hotte humidifiée; Graphique 6) pour s’assurer que l’environnement de travail atteint au moins 70% d’humidité tout au long du processus de micro-injection.
- Vérifiez le milieu de ponte pour les œufs après la période de ponte désirée. Faites-le dans une hotte humidifiée ou dans un autre environnement humide.
NOTE: La période de ponte généralement utilisée était de nuit, de 18 heures à 10 heures, d’une durée de ~ 16 h. - Si des œufs sont pondus dans l’agarose, utilisez des pinces fines pour les déterrer soigneusement et placez-les à la surface de l’agarose pour les garder humides (figure 7A).
- Appliquer une bande de ruban adhésif double face de 1 mm x 15 mm sur une lamelle de couverture de 22 mm x 30 mm (figure 7B). Placer le ruban adhésif vers le haut sur le milieu de ponte (figure 7C).
- Ramassez chaque œuf individuel de la surface de la gélose et passez au ruban adhésif double face à l’aide d’une brosse fine. Enlevez tous les œufs qui sont complètement blancs ou qui ont une coloration noire. Les œufs sains seront semi-transparents.
- Placez les œufs en forme de banane sur le côté, l’extrémité la plus grande étant collée sur le ruban adhésif double face (figure 7D).
REMARQUE: Conservez toujours les œufs dans un environnement très humide, comme une boîte de Petri coulée avec une couche de gélose à 1% sur le fond.
4. Préparation des réactifs CRISPR et des aiguilles d’injection
- Tirez les aiguilles de quartz à l’aide d’un extracteur de micropipettes de type Flaming/Brown.
- Biseauter les aiguilles de quartz à l’aide d’un biseauteur à micropipette.
- Utilisez du ruban adhésif double face pour fixer les aiguilles tirées dans un contenant transparent, comme une boîte de Pétri, jusqu’à ce qu’elles soient prêtes à être utilisées.
- Préparer la solution injectable en combinant 0,5 μL de protéine Cas9 (solution mère de 5 μg/μL) et 0,5 μL de sgRNA (solution mère de 4 μg/μL; voir le tableau des matériaux) avec 1 μL de tampon rouge de phénol dans un volume final de 5 μL. Pour précipiter les particules qui pourraient obstruer l’aiguille, vortex brièvement la solution et centrifuger pendant 3 minutes à la vitesse maximale.
- Remplissez l’aiguille d’injection en prenant soin de laisser le mélange d’injection près de l’extrémité effilée de l’aiguille. Enlevez les bulles, le cas échéant, du bout de l’aiguille.
- Placez soigneusement l’aiguille remblayée dans le porte-aiguille et serrez le collier en acier inoxydable pour maintenir l’aiguille solidement en place pendant la micro-injection.
- Générez un flux fiable de solution d’injection à partir de l’aiguille en caressant doucement l’extrémité biseautée avec un pinceau fin et humidifié, tout en délivrant des rafales de pression d’air à l’aiguille avec le système d’injection.
REMARQUE: L’aiguille est prête pour l’injection lorsque le mélange d’injection peut quitter l’embout en petites quantités.
5. Micro-injection et soins post-injection
- Préparez une plate-forme de micro-injection en remplissant une boîte de Petri propre de 100 mm x 15 mm avec une gélose à 1% pour former une couche plane de gélose au ras du dessus de la boîte.
- Placer une lamelle de couverture préalablement préparée avec ~25 embryons sur la plate-forme de gélose (Figure 8A).
REMARQUE: Toutes les étapes d’injection doivent être effectuées à l’intérieur d’une hotte humidifiée (~ 70% d’humidité). - Vérifiez la pression d’injection en plaçant la pointe de l’aiguille dans une goutte d’eau et en initiant le cycle d’injection.
REMARQUE : Une petite quantité de solution injectable doit se disperser dans l’eau si le réglage de pression est correct (figure 8B). - Insérez l’aiguille dans l’extrémité la plus grande de l’embryon, en vous approchant du côté gauche de la lamelle de couverture (figure 8C). Livrez la solution d’injection dans l’œuf et retirez l’aiguille rapidement.
- Une fois tous les œufs injectés, placez la lamelle de couverture sur la surface d’une nouvelle boîte de gélose 1 % et transférez-la dans une chambre d’humidité (figure 9).
6. Incubation et éclosion des embryons
- Placez la chambre d’éclosion dans un incubateur à 25 °C pendant 6 jours.
- Transférer tous les embryons survivants, à l’aide d’eau propre et d’une brosse fine, dans une boîte de Petri de 35 mm x 10 mm avec du papier filtre humidifié à l’eau recouvrant le fond de la boîte. Sceller la boîte de Petri avec un film de cire de paraffine plastique et maintenir à 25 °C pour permettre aux embryons d’éclore. Commencez à vérifier la survie des embryons 6 jours après l’injection.
NOTE: Les nymphes du premier stade commenceront à éclore vers le jour 8. - Transférer les nymphes, à l’aide d’un pinceau fin, dans une boîte de Petri contenant des coupures de feuilles. Couvrir le plat et sceller avec un film de cire de paraffine plastique.
- Incuber le plat scellé des nouveau-nés sur boutures de feuilles pendant 48 h à 25 °C.
- Transférer toutes les nymphes âgées de 2 jours d’une série d’injections dans une cage d’élevage avec des plants de maïs à l’aide d’une brosse fine. Si les personnes injectées avec un phénotype visible sont récupérées en nombre suffisant, élevez-les séparément pour maximiser la récupération du caractère cible dans la génération suivante. Sinon, effectuez un accouplement en masse de tous les injectés.
REMARQUE : Placez doucement les nouveau-nés dans la spire du plant de maïs pour leur fournir un refuge et assurer une humidité adéquate de leur environnement immédiat. - Élever les insectes dans les conditions décrites ci-dessus, en assurant une température, une humidité et des transferts réguliers vers les plants de maïs frais.
- Cribler la descendance pour les phénotypes attendus. Placez les individus présentant le phénotype désiré dans leur propre cage pour établir des lignées homozygotes.
Résultats
La chambre de ponte a été spécialement conçue pour permettre aux femelles de P. maidis de se nourrir tout en ovipositant dans un milieu protecteur à partir duquel leurs œufs pourraient facilement être récupérés. En utilisant cette méthode, des quantités suffisantes d’embryons précellulaires ont été récupérées pour la micro-injection d’ADN, d’ARN et / ou de protéines. Les femelles adultes de P. maidis pondent habituellement des œufs à l’intérieur du tissu foliaire des plants de maïs, ce qui rend difficile l’obtention de suffisamment d’œufs en peu de temps, car elle nécessite beaucoup de dissection foliaire. L’environnement artificiel de ponte offre une solution pour surmonter ces problèmes. Comme le montre le tableau 1, 6 483 œufs ont été prélevés sur un total de 645 femelles en 4 semaines. Les femelles commencent généralement à pondre après le jour 2 et fournissent la plupart des œufs du jour 4 au jour 6. L’activité de ponte a ralenti au jour 9. Chaque chambre de ponte a été mise en place le vendredi et vérifiée pour les œufs du dimanche au dimanche suivant. Le respect de cet horaire a permis de collecter la plupart des ovules pour des micro-injections pendant la semaine de travail.
La première application pratique de ce système de ponte a été de tester l’efficacité de l’élimination du gène médiée par Cas9, en utilisant l’orthologue P. maidis du gène de la couleur des yeux, blanc (Pmw), comme cible. On sait que les mutations du blanc entraînent une perte substantielle de pigmentation oculaire chez d’autres espèces d’insectes, et le blanc est autonome par les cellules, ce qui permet de détecter des mutations chez les individus injectés16,17. Pour augmenter le risque que même une petite mutation puisse entraîner une perte de fonction, les ARN guides ont été conçus pour couper dans la région de la cassette de liaison à l’ATP, ce qui est nécessaire pour la fonction blanche16. Les embryons de P. maidis ont été injectés soit avec 20% de rouge de phénol (tampon d’injection), tampon d’injection avec Cas9 à une concentration finale de 800 ng / μL (témoin Cas9), ou Cas9 dans le tampon d’injection avec trois ARN guides ajoutés à une concentration de 400 ng / μL chacun. La combinaison de trois guides au sein d’un mélange d’injection visait à maximiser davantage les chances de générer des mutants, à la fois en créant une délétion importante et en compensant la possibilité qu’un guide puisse être inefficace pour la coupe. Les taux de développement pour chaque traitement étaient comparables (tableau 2), 50 à 60 % des personnes injectées montrant des signes de développement. Les taux d’éclosion pour les contrôles tampon et Cas9 étaient également comparables; Cependant, les taux d’éclosion des personnes recevant le mélange de trois guides étaient relativement plus faibles. À l’heure actuelle, il n’est pas clair si la survie réduite est le résultat de la perte de la fonction blanche ou le résultat des conséquences imprévues de la combinaison de trois guides, telles que des effets hors cible (voir la section de discussion). Cependant, aucune des personnes présentant une perte complète de pigmentation oculaire (c.-à-d. knockout complet) n’a éclos, et aucune des personnes issues de l’injection n’avait les yeux blancs. L’efficacité sur cible de la mutagénèse à base de Cas9 a été vérifiée de deux façons. Tout d’abord, les personnes injectées ont été examinées pour détecter une perte de pigmentation oculaire.
Sur les 71 individus ayant reçu une injection guidée qui se sont développés, 23 ont montré un certain degré de perte de pigment (figure 10), et 9 de ces individus ont éclos, ce qui a entraîné un taux d’élimination de ≥32%. Aucune perte de pigment oculaire n’a été observée dans les deux traitements témoins. Deuxièmement, les mutations chromosomiques ont été confirmées par réaction en chaîne de la polymérase (PCR)18 et séquençage19. Parce qu’une lignée mutante n’a pas pu être récupérée, l’ADN génomique a été analysé à partir de pools d’embryons injectés avec le mélange à trois guides ou le tampon. Le mélange à trois guides devrait éliminer ~180 paires de bases du locus blanc . Cela peut être vu dans les produits de PCR amplifiés à partir de l’ADN génomique isolé des individus injectés, ainsi que les données de séquence associées générées à partir de ces produits (Figure 11). Ces preuves combinées indiquent que les embryons ont été injectés avant la cellularisation.

Figure 1 : Aspirateur. Un aspirateur efficace peut être assemblé en fixant une pompe à vide à l’admission, via un tube en plastique, à un tube conique en plastique de 15 mL. Environ 0,5 cm doit être soigneusement retiré du fond du tube conique. Une boule de coton doit être placée dans le tube conique, au-dessus de l’ouverture du tube en plastique, pour attraper les adultes de P. maidis au fur et à mesure qu’ils sont collectés et les garder hors de la pompe à vide. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2 : Construction de conteneurs pour adultes. (A) Les fournitures nécessaires (dans le sens des aiguilles d’une montre à partir du haut à gauche): écran, pistolet à colle chaude, lame de rasoir, récipient de 1 oz. (B) Un grand trou doit être coupé dans le fond du récipient de 1 oz, et un carré de moustiquaire est coupé juste assez grand pour couvrir ce trou. (C) L’écran est ensuite collé sur le trou à l’aide de colle chaude. (D) Une fois la colle prise, tout excès de maille doit être enlevé. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 3 : Sexage de P. maidis adultes. Les faces ventrales des adultes mâles (gauche) et femelles (droite) de P. maidis sont représentées. L’ovipositeur, visible au-dessus de l’abdomen féminin, est l’indicateur le plus clair du sexe d’un individu. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 4 : Sceller les adultes dans des contenants. (A) Un carré de 5 cm x 5 cm de film de cire de paraffine plastique. (B) Le film doit être étiré uniformément à 3-4 fois sa taille d’origine. (C) Une fois que les adultes ont été placés dans le contenant pour adultes, le film étiré doit être placé sur l’ouverture pour fixer les adultes. Une goutte de 400 μL de solution de saccharose à 10 % p/v doit ensuite être placée sur le film. (D) Pour fournir une pression alimentaire adéquate aux adultes, un deuxième carré de 5 cm x 5 cm de film de paraffine plastique doit être étiré de la même manière et placé sur la goutte de saccharose. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 5 : Mise en place d’une chambre de ponte. (A) Les fournitures nécessaires (dans le sens des aiguilles d’une montre à partir du haut à gauche) : pellicule de plastique, récipient complet pour adultes (avec adultes) et boîte de Petri contenant 1 % d’agarose (milieu de ponte). (B) Le récipient pour adultes doit être placé sur l’agarose avec le film de paraffine plastique/« sandwich » de saccharose à 10 % placé directement sur le milieu de ponte. (C) Une pellicule de plastique est utilisée pour fixer le contenant adulte au milieu de ponte. Cela empêche le milieu de sécher trop rapidement. (D) Il faut prendre soin d’éviter de couvrir l’écran du contenant pour adultes, afin que l’échange d’air puisse continuer à se poursuivre. E) Schéma de la chambre de ponte. Veuillez cliquer ici pour voir une version agrandie de cette figure.
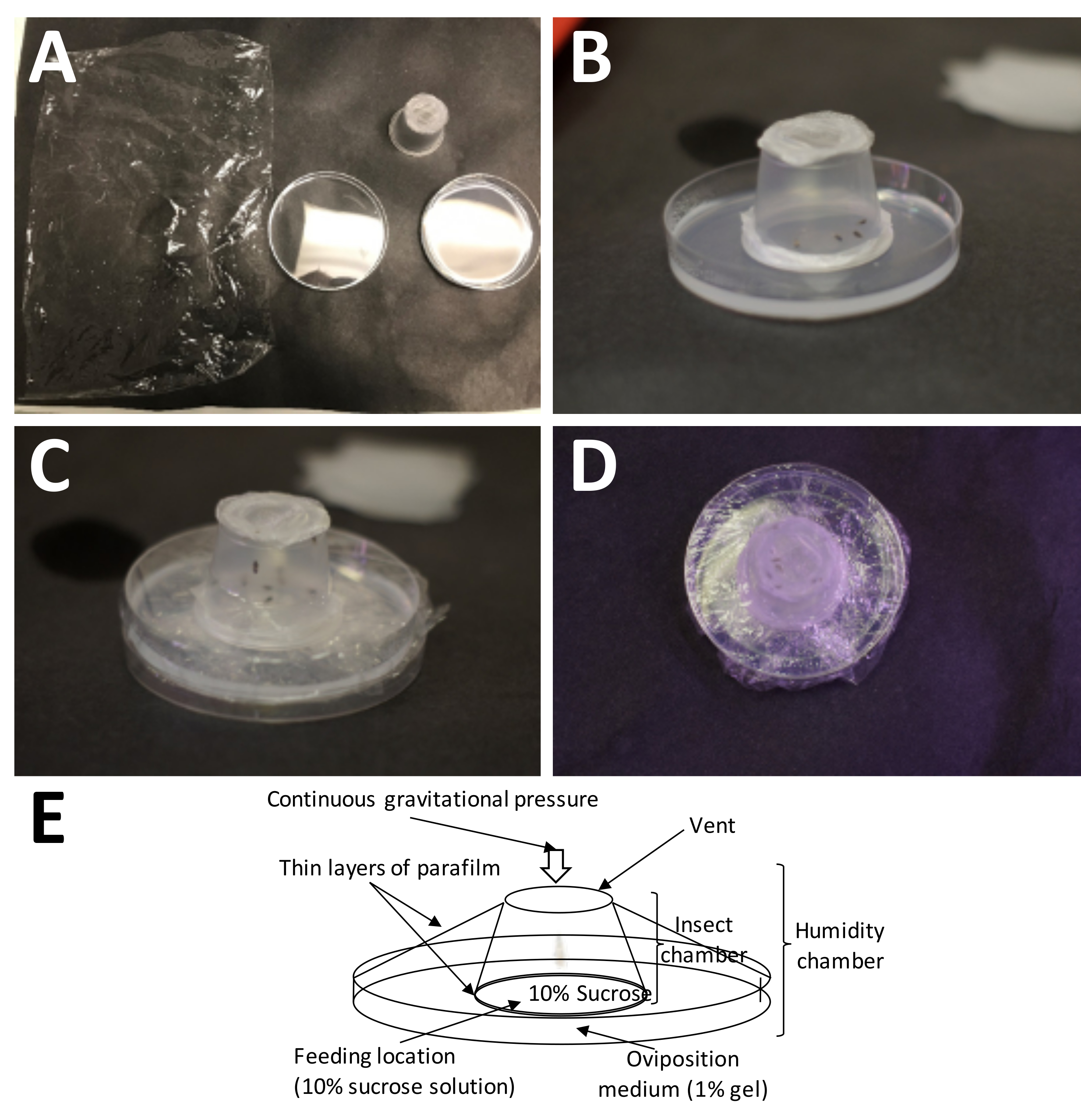{kind=link}

Figure 6 : Hotte humidifiée. Une hotte équipée d’un humidificateur a été installée autour de la lunette d’injection pour minimiser les courants d’air et maintenir l’humidité pendant la manipulation des embryons. Les volets peuvent être rabattus sur l’entrée une fois que le travailleur est en place, pour aider à maintenir des niveaux d’humidité appropriés. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 7: Collecte d’embryons en vue d’injections . (A) Embryons qui ont été déposés dans le milieu de ponte. Une paire de pinces fines est utilisée pour extraire les embryons du milieu et les placer à sa surface. (B) Une bande étroite de ruban adhésif double face de 1 mm x 15 mm sur une glissière de couverture de 22 mm x 30 mm. (C) La feuille de couverture peut être placée sur le milieu de ponte pour faciliter le transfert des embryons de la surface du milieu à la bande sur la lamelle de ponte. (D) Les embryons de P. maidis sont en forme de banane, avec une extrémité plus étroite que l’autre (extrémité étroite indiquée par une pointe de flèche rouge; extrémité plus large indiquée par une pointe de flèche jaune dans l’exemple embryonnaire). L’extrémité large de l’embryon doit être placée sur la bande. Veuillez cliquer ici pour voir une version agrandie de cette figure.
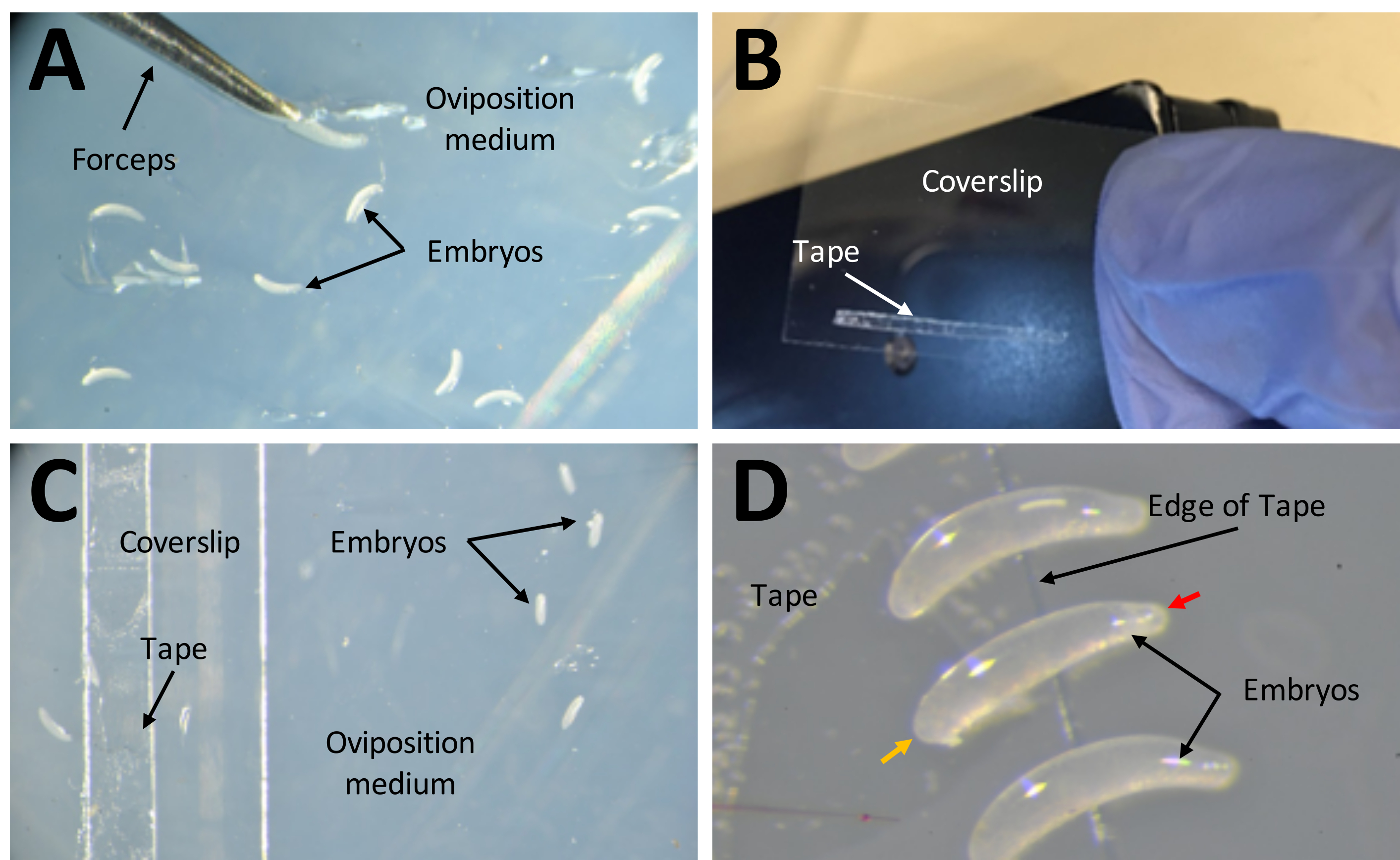{kind=link}

Figure 8 : Injection. (A) La plate-forme d’injection est une boîte de Petri remplie à ras bord de gélose à 1%. La lamelle de couverture avec une bande de ruban adhésif contenant des embryons doit être placée directement sur la surface de la plate-forme d’injection. (B) La pression d’injection doit être testée avant l’injection d’embryons en « injectant » une petite quantité de solution injectable dans une goutte d’eau. Cette méthode peut également être utilisée à tout moment pendant le processus d’injection pour vérifier la présence de sabots dans l’aiguille. (C) Les embryons doivent être injectés en insérant l’aiguille dans la plus grande extrémité de l’embryon. La solution injectable doit être visible si l’injection a réussi. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 9 : Soins post-injection. (A) Une fois que tous les embryons d’une lamelle de couverture ont été injectés, la lamelle de couverture doit être placée dans une boîte de Petri fraîche contenant 1% d’agarose. (B) La boîte de Petri avec la lamelle de couverture peut ensuite être maintenue dans une chambre d’humidité (comme celle montrée) jusqu’à l’éclosion des embryons. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 10 : Phénotype knockout de Pmw. (A) Témoins appariés selon l’âge et (B) embryons knock-out PMW, avec des yeux en développement indiqués par des pointes de flèches noires. L’embryon en B est en mosaïque, car une petite bande de pigmentation peut être vue. (C) Témoins appariés selon l’âge et (D) nouveau-nés knock-out Pmw, avec des encarts montrant un angle différent sur les yeux. Le nouveau-né en D est également en mosaïque. Une flèche blanche pointe vers une zone de l’image principale montrant une perte de pigmentation. Veuillez cliquer ici pour voir une version agrandie de cette figure.
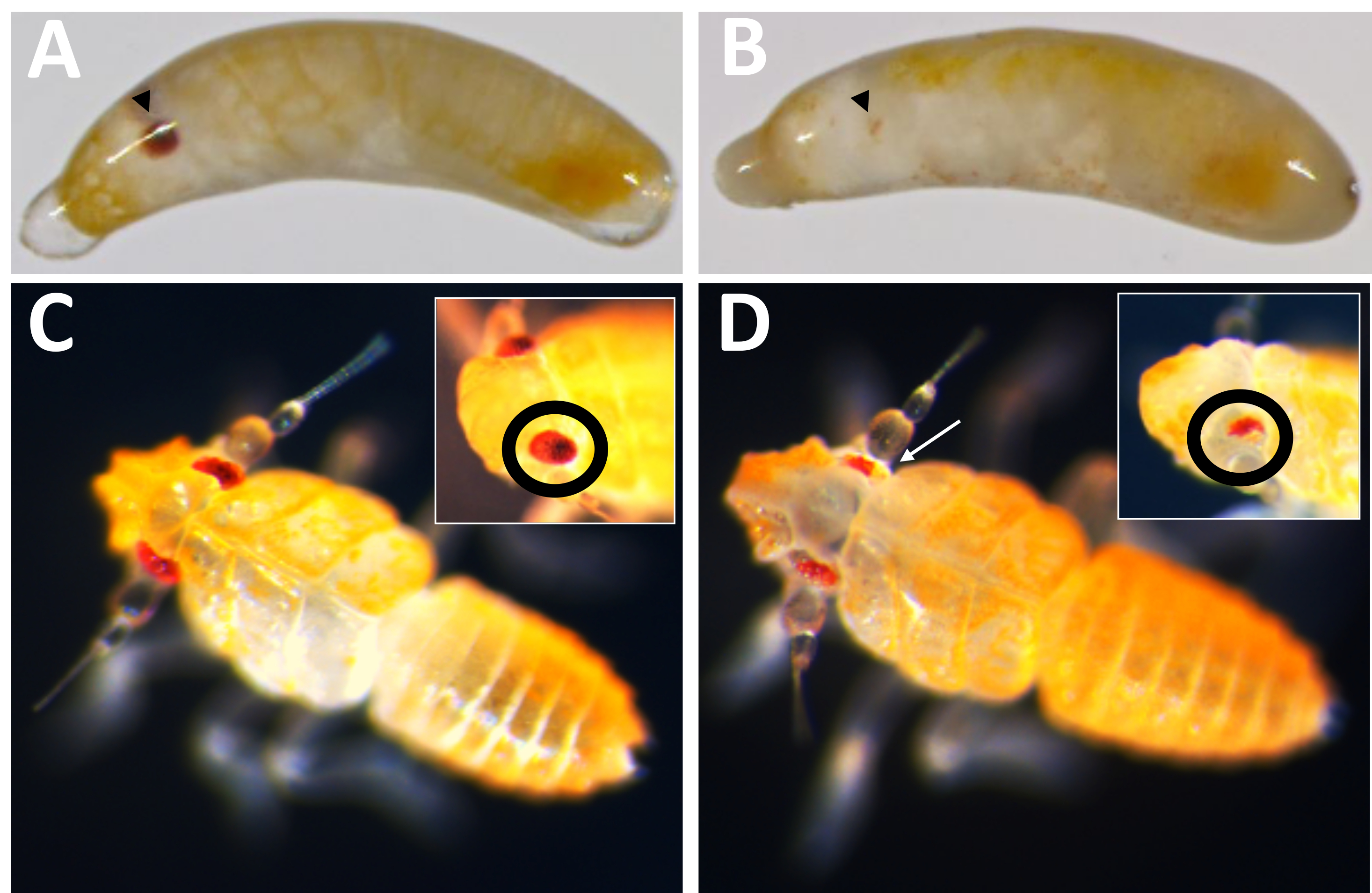{kind=link}

Figure 11 : Séquence éliminatoire de Pmw. (A) Modèle à l’échelle de l’ARNm Pmw, marqué par incréments de 500 pb, avec les emplacements des sites de liaison de l’ARNg indiqués: G1, bleu; G2, jaune; G3, rose. Toute mutation de décalage d’image générée à ce stade perturbera la majorité du produit de traduction. (B) Contexte génomique des sites d’ARNg, le tout en un seul exon (texte en majuscules en gras). Les sites de liaison de guidage sont mis en surbrillance dans les mêmes couleurs que A, et les PAM sont soulignés. L’envergure est de ~300 pb. La traduction dans le cadre de l’exon est montrée ci-dessus, sous forme d’abréviations d’une seule lettre en majuscules. Deux motifs spécifiques aux transporteurs de pigments oculaires sont marqués. Le motif CDEPT du domaine fonctionnel Walker B est encadré en violet, et le motif IHQP du domaine de la boucle H est encadré en vert. Les deux domaines sont essentiels à la fonction de transporteur ATP. (C) La région cible de PMW a été amplifiée à l’aide de deux cycles de PCR. Le produit de deuxième ronde a été examiné sur un gel pour détecter un changement de taille dû à l’élimination réussie de la région entre les guides. Couloirs: L = échelle de 100 pb; 1 = PCR contrôle de l’eau; 2 = oeufs injectés tampon; 3-4 = deux séries distinctes d’œufs injectés avec un mélange à trois guides. Seuls les embryons recevant le mélange à trois guides produisaient à la fois la bande WT (flèche rouge) et la bande résultant d’une excision complète (flèche blanche). (D) Pour confirmer l’identité de la bande inférieure (flèche blanche), cet ADN a été purifié, cloné et séquencé. La ligne supérieure est la séquence de type sauvage. Les deux autres lignes sont des séquences de deux clones. Trois clones supplémentaires correspondaient à la séquence inférieure. La surbrillance bleue indique le site de liaison du Guide 1, tandis que la surbrillance rose indique le site de liaison du Guide 3. Dans les deux allèles, toute la région située entre ces deux sites guides a été supprimée. Abréviations : Pmw = gène blanc de Peregrinus maidis; ARNg = ARN guide; PAM = motif adjacent protospacer ; ATP = adénosine triphosphate; PCR = réaction en chaîne de la polymérase; WT = type sauvage; KO = KO. Veuillez cliquer ici pour voir une version agrandie de cette figure.
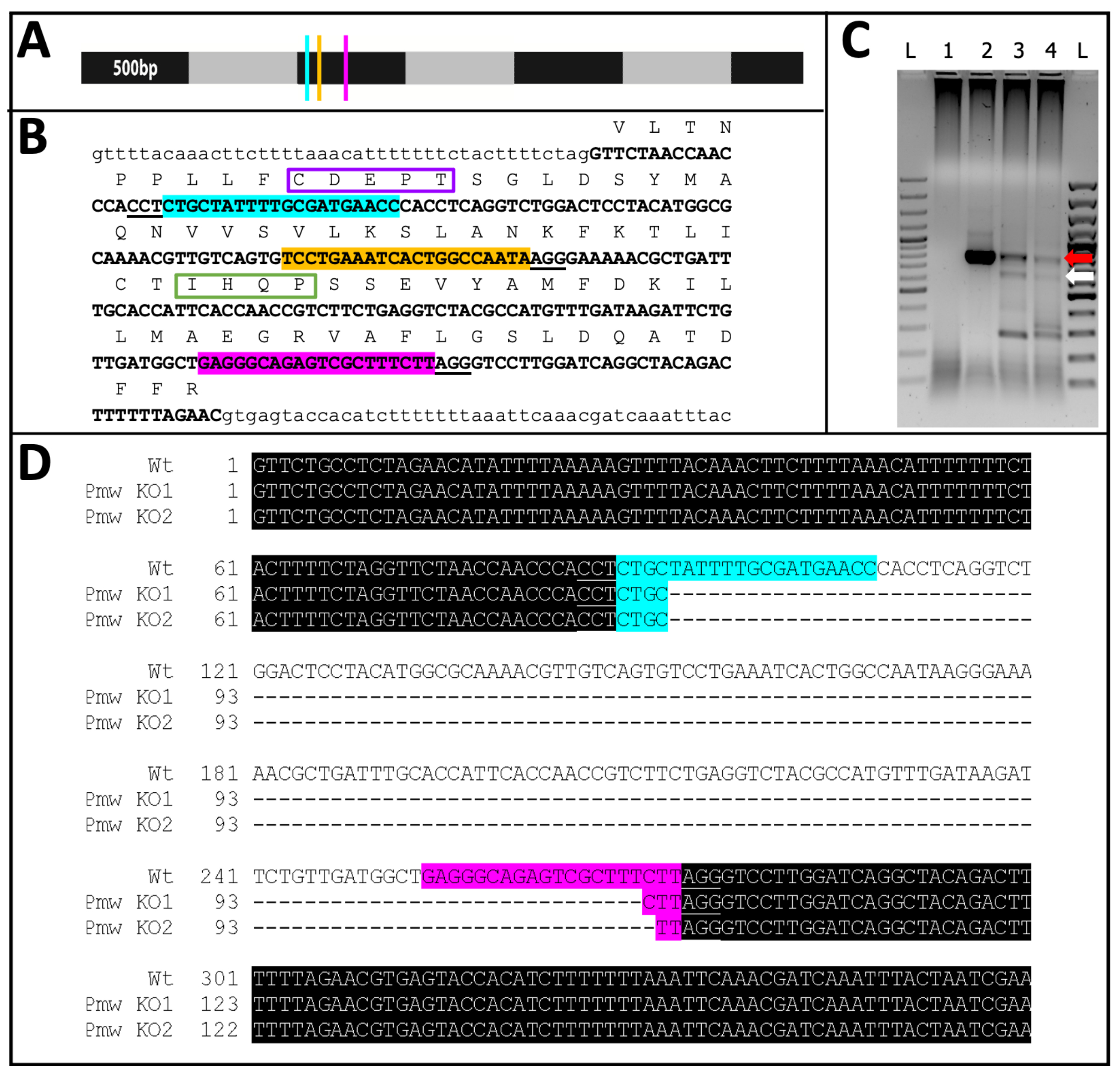{kind=link}
| Poser | # de tasses | # de femelles dans chaque tasse | # d’oeufs | # total d’oeufs | |||||||
| Jour 2 | Jour 3 | Jour 4 | Jour 5 | Jour 6 | Jour 7 | Jour 8 | Jour 9 | ||||
| 1 | 10 | 15 | 0 | 26 | 166 | 355 | 530 | 193 | 91 | 37 | 1398 |
| 2 | 15 | 15 | 22 | 238 | 489 | 699 | 520 | 379 | 203 | 58 | 2608 |
| 3 | 8 | 15 | 0 | 57 | 230 | 190 | 116 | 80 | 34 | 1 | 708 |
| 4 | 10 | 15 | 0 | 226 | 446 | 519 | 301 | 179 | 24 | 15 | 1710 |
| Total | 43 | 15 | 23 | 547 | 1331 | 1763 | 1467 | 831 | 352 | 111 | 6483 |
Tableau 1 : Collections d’œufs représentatives provenant d’un environnement de ponte artificiel. Les données de quatre ensembles de gobelets de collecte d’œufs sont présentées, avec des comptages d’œufs commençant le deuxième jour après l’installation et se poursuivant jusqu’au neuvième jour.
| Traitement par injection | Total injecté | Total développé | Nombre total d’éclos | Taux de développement (%) | Taux d’éclosion (%) |
| Tampon | 39 | 20 | 12 | 51 | 31 |
| Cas9 | 39 | 24 | 14 | 61 | 36 |
| ARNg Cas9 +Pmw | 121 | 71 | 28 | 59 | 28 |
Tableau 2 : Taux de survie et d’élimination des injections de 3 mélanges d’injections différents.
Discussion
Qualité de la ponte et nutrition
Récemment, des chercheurs travaillant avec une espèce apparentée, Nilaparvata lugens, ont obtenu les œufs qu’ils utilisaient pour les micro-injections directement à partir de la feuille, gardant les œufs injectés dans le tissu foliaire jusqu’à leur éclosion17. Bien que cette méthode à base de feuilles fournisse un environnement plus naturel pour le développement embryonnaire, elle a également augmenté les risques d’infections et de dommages aux œufs pendant le processus d’élimination. Le système de ponte artificielle présenté ici offre un environnement plus uniforme et réduit les risques d’endommagement des œufs lors de la manipulation. En installant les gobelets de ponte le vendredi, la majorité des ovules pondus ont été collectés au cours d’une semaine de travail typique, au profit de ceux qui effectuent le travail de micro-injection. Une mise en garde à cette méthode, cependant, est que le manque de nutriments dans le régime de solution de saccharose à 10% finira par affecter la santé des insectes, et les femelles dans les tasses commencent généralement à mourir après seulement 10 jours. La qualité des œufs commence également à baisser après 6 jours, comme en témoigne une augmentation des œufs morts ou malsains. Par conséquent, il est important d’être sélectif des œufs utilisés pour les micro-injections et de ne pas garder les femelles après le jour 6.
Taux de survie et humidité
Deux facteurs semblent être essentiels à la survie embryonnaire grâce au processus de micro-injection. L’aspect le plus difficile de la manipulation des embryons de P. maidis est de les empêcher de se dessécher après le retrait du milieu de ponte et tout au long de la micro-injection. Comme les œufs sont généralement pondus à l’intérieur des tissus végétaux, ils n’ont pas de coquille adéquate pour prévenir la déshydratation. Même dans le capot humidifié, des ensembles entiers d’œufs ont été perdus à cause de la dessiccation. Cependant, une humidité trop élevée pourrait également affecter les micro-injections si de l’eau s’est accumulée sur le ruban double face ou sur la lunette. Malheureusement, la déshydratation des ovules n’était généralement pas facile à remarquer pendant le processus de micro-injection, et ils semblaient souvent normaux jusqu’à 2 ou 3 jours plus tard, quand ils sont devenus complètement transparents, ne montrant aucun signe de développement.
La qualité des aiguilles semble également jouer un rôle important dans la survie. L’aiguille doit être biseautée pour minimiser les dommages inutiles à l’œuf. Lorsque l’aiguille est bloquée, l’utilisation de la fonction de dégagement de l’injecteur tout en caressant doucement le bout de l’aiguille avec un pinceau humidifié (voir l’étape 4.7) ramène généralement l’aiguille à un état fonctionnel. Quoi qu’il en soit, il est recommandé de ne mettre que de petites quantités de solution d’injection (~ 0,25 μL) dans chaque aiguille et de passer à une nouvelle aiguille toutes les quelques lames (~ 50-60 œufs) pour s’assurer que la qualité de l’aiguille est maintenue tout au long du processus d’injection.
Génération réussie d’un phénotype knockout
Pour réussir à transformer les cellules germinales, les micro-injections d’embryons doivent généralement être effectuées le plus tôt possible avant la cellularisation. Selon l’espèce d’insecte, la fenêtre de temps pour compléter les micro-injections varie de seulement quelques heures à une journée complète14,15,20. On ne sait toujours pas quand les embryons de P. maidis subissent une cellularisation. L’élimination médiée par Cas9 a été testée sur des embryons aussi jeunes que 4 h après la ponte (pel) jusqu’à 16 h pel, et les phénotypes attendus ont été observés dans toutes les expériences, suggérant que toutes les micro-injections ont été effectuées dans la fenêtre de précellurisation.
L’orthologue P. maidis du gène de la couleur des yeux, blanc, a été choisi parce que le phénotype knockout devait être facile à dépister chez les injectés en raison de sa nature autonome cellulaire. En effet, comme prévu, les knockouts mosaïques et totaux étaient clairement identifiables parmi les embryons recevant le mélange d’injection contenant Cas9 et des ARN guides. Malheureusement, aucun injecté n’a éclos complètement et un accouplement massif de survivants n’a pas réussi à générer une progéniture aux yeux blancs. Cependant, une lignée mutante a ensuite été générée avec succès en ciblant un gène différent (Klobasa et al., en cours). Cela suggère que l’échec de l’établissement d’une lignée mutante blanche est probablement dû soit à des effets hors cible (c.-à-d. Cas9 coupant des régions importantes ailleurs dans le génome) générant une mutation létale étroitement liée, soit à un rôle critique imprévu pour le blanc chez P. maidis.
Les données phénotypiques et moléculaires (Figure 8 et Figure 9) affirment qu’un knockout significatif dans le locus blanc a été créé dans un échantillon d’embryons injectés, ce qui entraînerait une perte totale de la fonction génique. De plus, alors que les mutations chez le blanc sont viables chez certaines espèces, il existe un précédent pour la réduction de l’activité blanche étant préjudiciable21,22. Cela dit, des effets hors cible ne peuvent être complètement exclus. La prédiction des cibles probables hors cible nécessite des données précises sur le séquençage du génome23, ce que l’état actuel des ressources génomiques chez P. maidis rend impossible à faire à l’heure actuelle. Quoi qu’il en soit, avec ces nouvelles méthodes, il est possible de tester d’autres gènes cibles en toute confiance, même en s’orientant vers une transgénèse plus traditionnelle dans le but d’apporter de nouveaux outils génétiques à ce ravageur pernicieux.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Le département d’entomologie et de pathologie végétale de l’Université d’État de Caroline du Nord fait partie d’une équipe soutenant le programme Insect Allies de la DARPA. Les points de vue, opinions et/ou conclusions exprimés sont ceux des auteurs et ne doivent pas être interprétés comme représentant les points de vue ou les politiques officiels du Département de la Défense ou du gouvernement des États-Unis. Les auteurs ne déclarent aucun intérêt concurrent. MDL, DR et AEW ont conçu le projet et ont fourni l’acquisition de fonds, l’administration du projet et les ressources. FC, WK, NG et MDL ont conçu et conçu les expériences de micro-injection; OH a conçu et conçu la méthode de ponte. FC et WK ont effectué les expériences; FC et WK ont analysé les résultats; et FC, WK, NG et MDL ont écrit le manuscrit. Les auteurs tiennent à remercier tout particulièrement Kyle Sozanski et Victoria Barnett pour leur aide dans le maintien des colonies de P. maidis.
matériels
| Name | Company | Catalog Number | Comments |
| 1 oz Containers | Dart | P100N | Adult container for egg-laying setup |
| 15 mL Conical Tubes | Olympus | Genesee 28-103 | Serves as collection tube on vacuum aspirator setup |
| 15 mL Conical Tubes | Olympus | Genesee 28-106 | For making 10% sucorose solution and for holding adults when chilling before screening |
| Aspirator | Bioquip | 1135A | For handling planthoppers |
| Vacuum Aspirator | Fischer Technical | LAV-3 | Vacuum for aspirating larger numbers of insects |
| Blue Spectrum LED Lights | Home Depot | GLP24FS/19W/LED | Grow lights for potted corn plants hoppers are feeding on |
| Cas9 | TrueCut Cas9 Protein v2 | A36498 | Endonuclease for cutting planthopper genes |
| Clear Vinyl Tubing | Home Depot | 3/8 in. I.D. x 1/2 in. O.D. x 10 ft. | Connects collection tube to pump on vacuum aspirator setup |
| Corn planthoppers | North Carolina State University | N/A | Request from Dr. Anna Whitfield's lab |
| Cotton balls | Genessee | 51-101 | Serves as a filter/insect catcher in collection tube on vacuum aspirator setup |
| Double sided tape | Scotch Double Sided Tape | NA | Holding eggs for microinjection |
| Early Sunglow corn | Park Seed Company | 05093-PK-N | Corn for rearing planthoppers |
| epTIPS Microloader Tips | Eppendorf | C2554691 | Backfilling needle loading tips |
| Femtojet Microinjection System | Eppendorf | 5247 | Controls injection pressure (12-20 psi, depending on needle bore size) |
| Nutri-Fly Drosophila Agar | Genessee | 66-103 | Substrate for everything except egg-laying dish |
| Fine forceps | Bioquip | 4731 | Egg handling |
| General Purpose LE Agarose | Apex | 20-102 | Substrate inn egg-laying dish (oviposition medium) |
| Guide RNA 1 - GGUUCAUCGCAAAAUAGCAG | Synthego | CRISPRevolution sgRNA EZ Kit (1.5 nmol) | RNA guides for targeting planthopper white gene |
| Guide RNA 2 - UCUGAAAUCACUGGCCAAUA | Synthego | CRISPRevolution sgRNA EZ Kit (1.5 nmol) | RNA guides for targeting planthopper white gene |
| Guide RNA 3 - GAGGGCAGAGUCGCUUUCUU | Synthego | CRISPRevolution sgRNA EZ Kit (1.5 nmol) | RNA guides for targeting planthopper white gene |
| Humidifyer | Homedics | UHE-CM45 | For providing humidity in humidified hood |
| Humidity chamber | Billups-Rothenberg | MIC-101 | For holding injected embryos until hatching |
| Insect rearing cages | Bioquip (special order) | Close to 1450 L (has plastic front and mesh fabric sides) | Cage for planthoppers on corn |
| Laser-based Micropipiette Puller | Sutter Instruments | P-2000/G | For making injection needles / Heat = 700, FIL = 4, VEL = 40, DEL = 170, PUL = 160 |
| Leica M165 FC Fluorescence Stereomicroscope | Leica | M165 FC | Planthopper screening |
| Microinjection Scope | Leica | MZ12-5 | Microinjection scope outfited with an XY stage |
| Micromanipulator | Narishige | MN-151 | For positioning microinjection needle |
| Micropipette beveler | Sutter Instruments | FG-BV10-D | For beveling injection needles / Used 'fine' graded plate at 20° angle |
| Microscope Stage | AmScope | GT100 X-Y Gliding Table | For positioning and moving embryos under microscope |
| Miniature Paint Brush | Testor #2 8733 | Sold in 3 pack 281206 | Fine paintbrushes for embryo handling |
| Needle Holder | Narishige | HI-7 | For holding the microinjection needle |
| Percival Incubator | Percival | I41VLH3C8 | Rearing injectees until hatch |
| Petri Dishes (100 x 15 mm) | VWR | 89038-968 | Making agar dish for egg-lay |
| pGEM-T Easy Vector System I cloning kit | Promega | A1360 | Cloning Pm white target site |
| Phenol Red | Sigma | 143-78-8 | Microinjection buffer |
| Plain Microscope Slides or coverslip | Fisher Scientific | 12-549-3 | Hold eggs for microinjection |
| Plasmid DNA Midi Kit | Zymo | D4200 | Purification of injection-ready plasmid DNAs |
| Plastic paraffin film | Pechiney Plastic Packaging | PM-996 | Roll size 4 in. x 125 ft |
| Plastic wrap | Glad ClingWrap Plastic Wrap | NA | Wrap the entire egg-laying chamber |
| Primer - PmW CRISPR check F1 - AAGGAATTTCTGGAGGTGAAA | IDT | 25 nmole DNA Oligo | First-round Primer for amplifing across target site within the Pm white gene |
| Primer - PmW CRISPR check R1 - GATTCCTCGCTGTTGGGT | IDT | 25 nmole DNA Oligo | First-round Primer for amplifing across target site within the Pm white gene |
| Primer - PmW CRISPR check F3 - TCACAGACCCTGGTGCTAATC | IDT | 25 nmole DNA Oligo | Second-round Primer for amplifing across target site within the Pm white gene |
| Primer - PmW CRISPR check R3 - GTCCACAATCCACACTTCTGA | IDT | 25 nmole DNA Oligo | Second-round Primer for amplifing across target site within the Pm white gene |
| Quartz capillaries | Sutter Instruments | QF100-50-10 | For making microinjection needles / O.D. 1 mm, I.D. 0.7 mm, 10 cm length |
| Screen (White Organza Fabric) | Joann Fabrics | 16023889 | For covering the adult container |
| Sparkleen | Fisher Scientific | 04-320-4 | Wash dishes |
| Sucrose | Fisher Scientific | BP220-1 | To make 10% sucorose solution |
Références
- Namba, R., Higa, S. Y. Host plant studies of the corn planthopper, Peregrinus maidis (Ashmead) in Hawaii. Proceedings of the Hawaiian Entomological Society. 21, 105-108 (1971).
- Singh, B. U., Seetharama, N. Host plant interactions of the corn planthopper, Peregrinus maidis Ashm.(Homoptera: Delphacidae) in maize and sorghum agroecosystems. Arthropod-Plant Interactions. 2 (3), 163-196 (2008).
- Tsai, J. Occurrence of a corn disease in Florida transmitted by Peregrinus maidis. Plant Disease Reporter. 59 (10), 830-833 (1975).
- Chelliah, S., Basheer, M. Biological studies of Peregrinus maidis (Ashmead) (Araeopidae: Homoptera) on sorghum. Indian Journal of Entomology. 27, 466-471 (1965).
- Lastra, J., Esparza, J. Multiplication of vesicular stomatitis virus in the leafhopper Peregrinus maidis (Ashm.), a vector of a plant rhabdovirus. Journal of General Virology. 32 (1), 139-142 (1976).
- Nault, L. R., Ammar, E. -. D. Leafhopper and planthopper transmission of plant viruses. Annual Review of Entomology. 34 (1), 503-529 (1989).
- Ammar, E. -. D., Tsai, C. -. W., Whitfield, A. E., Redinbaugh, M. G., Hogenhout, S. A. Cellular and molecular aspects of rhabdovirus interactions with insect and plant hosts. Annual Review of Entomology. 54, 447-468 (2009).
- Barandoc-Alviar, K., Ramirez, G. M., Rotenberg, D., Whitfield, A. E. Analysis of acquisition and titer of Maize mosaic rhabdovirus in its vector, Peregrinus maidis (Hemiptera: Delphacidae). Journal of Insect Science. 16 (1), 14 (2016).
- Tsai, J. H., Steinberg, B., Falk, B. W. Effectiveness and residual effects of seven insecticides on Dalbulus maidis (Homoptera: Cicadellidae) and Peregrinus maidis (Homoptera: Delphacidae). Journal of Entomological Science. 25 (1), 106-111 (1990).
- Yao, J., Rotenberg, D., Afsharifar, A., Barandoc-Alviar, K., Whitfield, A. E. Development of RNAi methods for Peregrinus maidis, the corn planthopper. PloS One. 8 (8), 70243 (2013).
- Esvelt, K. M., Smidler, A. L., Catteruccia, F., Church, G. M. Emerging technology: concerning RNA-guided gene drives for the alteration of wild populations. Elife. 3, 03401 (2014).
- Gantz, V. M., Bier, E. The mutagenic chain reaction: a method for converting heterozygous to homozygous mutations. Science. 348 (6233), 442-444 (2015).
- Yao, J., Rotenberg, D., Whitfield, A. E. Delivery of maize mosaic virus to planthopper vectors by microinjection increases infection efficiency and facilitates functional genomics experiments in the vector. Journal of Virological Methods. 270, 153-162 (2019).
- Kimelman, D., Martin, B. L. Anterior-posterior patterning in early development: three strategies. Wiley Interdisciplinary Reviews: Developmental Biology. 1 (2), 253-266 (2012).
- Mito, T., Nakamura, T., Noji, S. Evolution of insect development: to the hemimetabolous paradigm. Current Opinion in Genetics & Development. 20 (4), 355-361 (2010).
- Grubbs, N., Haas, S., Beeman, R. W., Lorenzen, M. D. The ABCs of eye color in Tribolium castaneum: orthologs of the Drosophila white, scarlet, and brown Genes. Genetics. 199 (3), 749-759 (2015).
- Xue, W. H., et al. CRISPR/Cas9-mediated knockout of two eye pigmentation genes in the brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Insect Biochemistry and Molecular Biology. 93, 19-26 (2018).
- Lorenz, T. C. Polymerase chain reaction: basic protocol plus troubleshooting and optimization strategies. Journal of Visualized Experiments: JoVE. (63), e3998 (2012).
- Sanger, F., Coulson, A. R. A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. Journal of Molecular Biology. 94 (3), 441-448 (1975).
- Chu, F. C., Wu, P. S., Pinzi, S., Grubbs, N., Lorenzen, M. D. Microinjection of Western Corn Rootworm, Diabrotica virgifera virgifera, embryos for germline transformation, or CRISPR/Cas9 genome editing. Journal of Visualized Experiments: JoVE. (134), e57497 (2018).
- Brent, C. S., Hull, J. J. RNA interference-mediated knockdown of eye coloration genes in the western tarnished plant bug (Lygus hesperus Knight). Archives of Insect Biochemistry and Physiology. 100 (2), 21527 (2019).
- Khan, S. A., Reichelt, M., Heckel, D. G. Functional analysis of the ABCs of eye color in Helicoverpa armigera with CRISPR/Cas9-induced mutations. Scientific Reports. 7 (1), 1-14 (2017).
- Manghwar, H., et al. CRISPR/Cas systems in genome editing: methodologies and tools for sgRNA design, off-target evaluation, and strategies to mitigate off-target effects. Advanced Science. 7 (6), 1902312 (2020).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.