
Method Article
Sondes d’attache de jauge de tension pour quantifier la mécanique et l’adhérence de l’intégrine médiée par le facteur de croissance
Dans cet article
Résumé
TGT surface est une plate-forme innovante pour étudier la diaphonie facteur de croissance-intégrine. La conception flexible de la sonde, la spécificité du ligand d’adhérence et la modulation précise des conditions de stimulation permettent des évaluations quantitatives robustes de l’interaction EGFR-intégrine. Les résultats mettent en évidence l’EGFR en tant que mécanisme de réglage de la mécanique de l’intégrine«mécano-organisateur », influençant l’assemblage de l’adhérence focale et l’étalement des cellules.
Résumé
Les organismes multicellulaires s’appuient sur les interactions entre les récepteurs membranaires et les ligands apparentés dans la matrice extracellulaire (ECM) environnante pour orchestrer de multiples fonctions, notamment l’adhésion, la prolifération, la migration et la différenciation. Les forces mécaniques peuvent être transmises de la cellule via l’intégrine du récepteur d’adhésion aux ligands de l’ECM. La quantité et l’organisation spatiale de ces forces générées par les cellules peuvent être modulées par les récepteurs du facteur de croissance, y compris le récepteur du facteur de croissance épidermique (EGFR). Les outils actuellement disponibles pour quantifier les changements médiés par la diaphonie dans la mécanique cellulaire et les relier aux adhérences focales, à la morphologie cellulaire et à la signalisation sont limités. Des capteurs de force moléculaire à base d’ADN connus sous le nom d’attaches de jauge de tension (TGT) ont été utilisés pour quantifier ces changements. Les sondes TGT sont uniques dans leur capacité à moduler le seuil de force sous-jacent et à signaler les forces des récepteurs à l’échelle du piconewton sur toute la surface de la cellule adhérente à une résolution spatiale limitée par diffraction. Les sondes TGT utilisées ici reposent sur la dissociation irréversible d’un duplex d’ADN par des forces récepteur-ligand qui génèrent un signal fluorescent. Cela permet de quantifier la tension cumulée de l’intégrine (historique de force) de la cellule. Cet article décrit un protocole utilisant des TGT pour étudier l’impact de l’EGFR sur la mécanique de l’intégrine et la formation d’adhérence. L’assemblage de la plate-forme de détection mécanique TGT est systématiquement détaillé et la procédure d’imagerie des forces, des adhérences focales et de l’étalement des cellules est décrite. Dans l’ensemble, la capacité de moduler le seuil de force sous-jacent de la sonde, le ligand d’adhésion et le type et la concentration du facteur de croissance utilisé pour la stimulation en font une plate-forme robuste pour étudier l’interaction de divers récepteurs membranaires dans la régulation des forces médiées par l’intégrine.
Introduction
Les cellules ont la capacité intrinsèque de détecter, de générer et de répondre aux forces mécaniques, ce qui entraîne des changements dans le phénotype cellulaire et le remodelage du microenvironnement local 1,2. Les forces jouent un rôle crucial dans la régulation de nombreux aspects du comportement cellulaire, y compris l’adhésion, la migration, la prolifération, la différenciation et la cicatrisation des plaies 3,4. Les aberrations dans l’échange mécanique bidirectionnel entre une cellule et le microenvironnement peuvent conduire à des états malades, y compris le cancer5. De nombreux récepteurs membranaires sont impliqués dans le maintien de l’homéostasie de la matrice cellulaire; parmi ceux-ci, les intégrines et le récepteur du facteur de croissance épidermique (EGFR) ont une synergie robuste 6,7. Classiquement, les intégrines établissent le lien mécanique entre le microenvironnement et le cytosquelette intracellulaire tandis que l’EGFR régule la croissance, la prolifération et la survie des cellules 8,9. L’EGFR est une cible thérapeutique très étudiée, axée sur la régulation externe-in facilitant la signalisation intracellulaire. La diaphonie EGFR-intégrine a été établie génétiquement et biochimiquement pour réguler la progression de multiples maladies, y compris le cancer10,11. Bien que les études indiquent l’existence d’une interaction EGFR-intégrine, les résultats sont attribués aux voies de signalisation loin de la membrane plasmique 7,12,13,14. L’impact de l’EGFR, ou d’autres facteurs de croissance, sur la mécanique cellulaire reste largement inexploré en partie en raison du manque d’outils pour mesurer les forces cellulaires et les résultats de signalisation. Le défi consiste à identifier des outils appropriés pour étudier la communication entre ces paradigmes de signalisation parallèle et quantifier leurs contributions spécifiques à la mécanique cellulaire.
Plusieurs approches ont été développées pour mesurer les forces générées par les récepteurs d’adhésion cellulaire, et le lecteur est dirigé vers des examens approfondis de ces techniques15,16. En bref, la microscopie à force de traction et la détection de réseaux de micro-piliers reposent sur la déformation d’un substrat sous-jacent pour déduire les forces du nanonewton (nN), soit un ordre de grandeur supérieur aux forces de récepteurindividuelles 17,18. Les techniques à molécule unique, y compris l’AFM et la pince optique, sont sensibles aux forces du piconewton (pN) à protéine unique, mais ne mesurent qu’un seul récepteur à la fois et n’offrent pas une bonne (ou aucune) résolution spatiale. Les sondes de tension moléculaire à base d’ADN et les sondes d’attache de jauge de tension (TGT) offrent une résolution de force pN avec une résolution spatiale limitée à la diffraction (ou meilleure), ce qui leur confère un rôle unique dans l’étude des forces unicellulaires19,20 de divers types de cellules, y compris les fibroblastes, les cellules cancéreuses, les plaquettes et les cellules immunitaires 21,22,23,24 . Alors que les sondes de tension moléculaire ont un élément « ressort » extensible, idéal pour l’imagerie en temps réel, les sondes TGT se rompent de manière irréversible, laissant derrière elles un « historique de force » fluorescent. Les TGT modulent en outre le seuil de tension du substrat sous-jacent; une série de sondes avec des compositions chimiques similaires mais des forces de rupture différentes, ou des tolérances de tension (Ttol), peuvent être utilisées pour quantifier la tension minimale requise pour la formation d’adhérence focale et la propagation cellulaire. Les sondes TGT sont constituées de deux brins d’ADN complémentaires, l’un ancré à la surface et l’autre présentant un ligand à la cellule. Si un récepteur se lie au ligand et exerce une force supérieureau T tol de la sonde, les brins seront séparés. Ttol est défini comme la force constante nécessaire pour rompre 50% des sondes dans un intervalle de 2 s dans des conditions idéales. Dans les sondes TGT « turn-on », un quencher sur le brin supérieur peut être séparé d’un fluorophore sur le brin inférieur. Ce n’est que lorsque la sonde TGT a été rompue, vraisemblablement par des forces supérieures ou égales à Ttol, qu’un signal fluorescent sera généré. Les sondes TGT peuvent également être fixées, ce qui permet de manipuler facilement les systèmes biologiques et de tester plusieurs conditions. Pour ces raisons, des sondes TGT ont été utilisées dans ce travail.
Des sondes TGT ont été utilisées pour étudier comment l’adhésion cellulaire dépendante de l’intégrine et les forces mécaniques sont modulées par l’EGFR21 activé. Ce travail a établi l’EGFR en tant que « mécano-organisateur », accordant l’organisation de l’adhésion focale et la génération de tension. En outre, il a été constaté que la stimulation par l’EGF influençait la distribution et la maturité des adhérences focales et améliorait la propagation cellulaire. Cette approche pourrait être utilisée dans de futures études pour étudier comment les facteurs de croissance influencent les forces mécaniques dans la progression et la dynamique tumorales. Alors que le rôle de la diaphonie EGFR-intégrine dans la régulation de la transition épithéliale à mésenchymateuse est établi, le rôle des forces mécaniques dans ce processus reste sous-exploré10.
Ici, un protocole détaillé est présenté pour ces expériences couvrant la synthèse et l’assemblage de sondes TGT 56 pN, la génération de surfaces TGT sur des couvercles en verre, l’application de cellules Cos-7 sur la surface TGT et la stimulation avec EGF, la fixation et la coloration des cellules avec de la phalloïdine et un anticorps anti-paxilline, l’imagerie par fluorescence par réflexion interne totale (TIRF) à haute résolution et la microscopie de contraste par interférence de réflexion (RICM), et la quantification des images. Ce protocole, bien qu’écrit pour étudier la stimulation EGF des cellules Cos-7, est facilement adaptable à de nombreuses expériences basées sur TGT. Différents ligands, Ttol, types de cellules, paramètres de stimulation, protéines marquées après fixation et analyse quantitative peuvent être facilement substitués, ce qui rend ce protocole robuste et largement utile.
Protocole
1. Préparation d’oligonucléotides TGT
REMARQUE: Les détails de la synthèse de la sonde oligonucléotidique sont décrits ici. Veuillez noter que certaines modifications et étapes de purification peuvent être externalisées pour une synthèse personnalisée.
- Activer l’amine primaire du peptide cyclo[Arg-Gly-Asp-D-Phe-Lys(PEG-PEG)] avec le liant azide-NHS tel que décrit par Zhang et al22 en mélangeant dans un rapport 1:1,5 (100:150 nmoles) dans un volume final de 10 μL de diméthylformamide. Ajouter 0,1 μL de la base organique triéthylamine et incuber pendant 12 h à 4°C.
- Purifier le produit par CLHP en phase inverse en utilisant 0,1 M TEAA (solvant A) et 100% acétonitrile (solvant B) avec un débit de 1mL/ min et la condition initiale de 10% solvant B réglé à un gradient de 0,5%/min. Combiner les pics élués (absorbance à 203 nm) et vérifier par spectrométrie de masse MALDI-TOF. Le produit est cRGDfK-azide.
- Pour générer le brin supérieur TGT, combinez l’azoture cRGDfK et l’oligonucléotide alcyne-21-BHQ2 (brin supérieur TGT: 5Hexynyl/GTGAAATACCGCACAGATGCG/3BHQ_2) dans un rapport de 2:1 (͂200 μM: 100 μM) dans 100 μL de 1x solution saline tamponnée au phosphate (PBS) avec 5 mM d’ascorbate de sodium et 0,1 μM de Cu-THPTA préformé. Laisser la réaction se poursuivre pendant au moins 4 h à température ambiante (RT) ou pendant la nuit à 4 °C.
- Traiter le mélange à travers le gel de dessalage P2 pour éliminer l’excès de colorant, de sous-produits, de solvant organique et de réactifs non réagis. Préparez la colonne de centrifugeuse avec 650 μL de gel P2 préhydraté en la faisant tourner à 18 000 x g pendant 1 min. Jeter le liquide d’écoulement et ajouter le mélange réactionnel. Tournez à nouveau à 18 000 x g pendant 1 min et récupérez le débit. Porter le mélange réactionnel à un volume final de 300 μL avec de l’eau ultrapure.
REMARQUE: Pré-hydrater le gel P2 avec de l’eau pendant 6 h. - Purifier le mélange réactionnel dessalé par CLHP en phase inverse. Les solvants organiques utilisés pour cette purification comprennent 0,1 M TEAA enH2O(solvant A) et 100% MeCN (solvant B, ou phase mobile).
- Avant d’injecter le mélange, équilibrer la colonne avec une condition initiale de 10% de solvant B avec un gradient de 1% / min. Réglez le débit à 1 mL/min. Injecter le mélange réactionnel dans la boucle HPLC avec une aiguille d’injection de 500 μL.
- Collectez le produit en visualisant l’absorption maximale à 260 nm pour l’ADN et à 560 nm pour le quencher BHQ2. Sécher le produit élué pendant la nuit dans un concentrateur centrifuge sous vide.
- Utiliser la substitution nucléophile pour coupler le brin inférieur TGT à l’ester Cy3B-NHS comme décrit dans Ma et. al25. Mélanger 100 μM du brin inférieur TGT de 56 pN (5Biosg/TTTTTT/iUniAmM/CGCATCTGTGCGGTATTTCACTTT) avec 50 μg d’ester Cy3B-NHS pré-dissous dans 10 μL de DMSO. Ajuster le pH de ce mélange à 9 avec 0,1 M de bicarbonate de sodium et porter le volume final à 100 μL avec 1x PBS. Incuber le mélange réactionnel pendant la nuit à RT.
- Purifier le mélange séquentiellement à l’aide de la filtration sur gel P2 et de la CLHP en phase inverse pour séparer les réactifs, les sels et les solvants organiques qui n’ont pas réagi (décrits aux étapes 1.4 et 1.5).
- Estimer la concentration des conjugués oligonucléotides-colorants purifiés en enregistrant leur absorbance à 260 nm à l’aide d’un spectrophotomètre.
- Caractériser les produits purifiés par spectrométrie de masse MALDI-TOF. Dissoudre l’excès d’acide 3-hydroxypicolinique dans le solvant TA50 (acétonitrile à 50:50 v/v et TFA à 0,1 % dans ddH2O) pour préparer la matrice MALDI fraîche. Les masses moléculaires estimées et mesurées pour les produits étiquetés sont les suivantes : cRGDfK-1-BHQ2 - 8157,9 (calculé), 8160,1 (trouvé); Cy3B étiqueté 56 pN TGT - 10272.7 (calculé), 10295.8 (trouvé).
- Dissoudre les brins supérieur et inférieur séparément dans de l’eau sans nucléase à une concentration comprise entre 30 et 50 μM. Utilisez des embouts de pipette sans DNase pour éviter la contamination du stock. Conserver à 4 °C pour les applications à court terme ou à -20 °C pour les applications à long terme. La stabilité des oligonucléotides n’est pas affectée par les cycles répétés de gel-dégel.
2. Préparation de surface
Jour 1 :
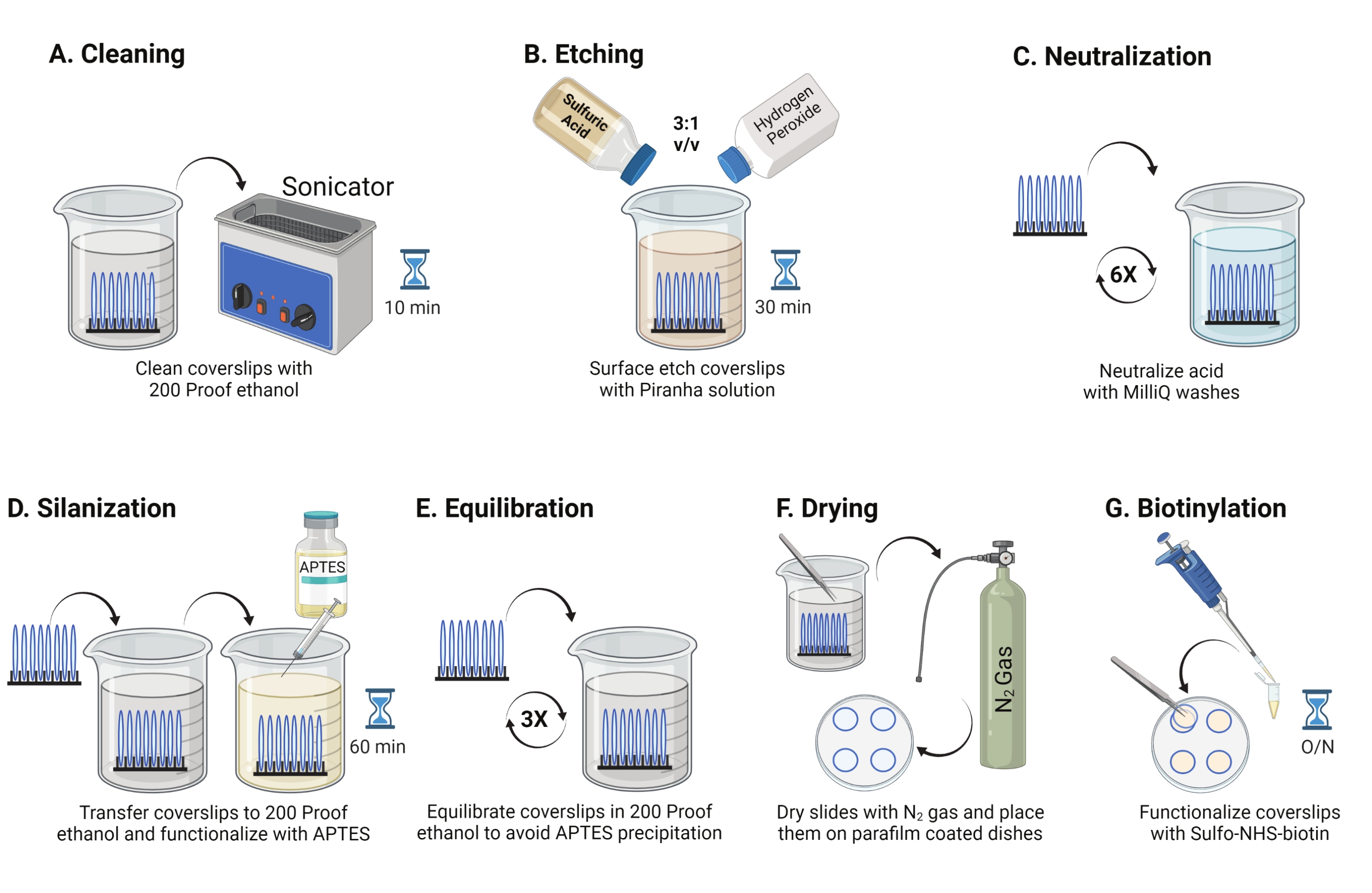
- Placez des couvercles en verre de 25 mm (jusqu’à 8) dans un rack en polytétrafluoroéthylène. Placer le rack dans un bécher borosilicate de 50 mL contenant 40 mL d’éthanol 200. Couvrir le bécher avec un film de paraffine pour éviter l’entrée d’eau et soniquer à une fréquence de fonctionnement de 35 kHz pendant 10-15 min à RT (Figure 1A).
- Remplir un bécher de 50 mL avec 40 mL de solution de piranha fraîchement préparée en mélangeant de l’acide sulfurique et du peroxyde d’hydrogène dans un rapport de 3:1 dans un bécher en pyrex. Remuer avec une pipette en verre. Transférer le porte-couvercle dans le bécher et incuber pendant 30 min à RT dans la hotte pour graver la surface du couvercle (Figure 1B).
REMARQUE: Portez un EPI complet, y compris un manteau de laboratoire, des gants et des lunettes, et travaillez dans la hotte de fumée chimique. Ajouter lentement le peroxyde d’hydrogène à l’acide pour éviter la surchauffe de la solution. - Après la gravure, utilisez une pince à épiler pour transférer le porte-couvercle dans un bécher avec de l’eau ultrapure. Répétez cette étape six fois à des intervalles de 15 s pour neutraliser complètement l’acide (Figure 1C).
REMARQUE: Laissez la solution de piranha dans la hotte chimique pendant la nuit avant de la jeter dans le conteneur à déchets acides. - Inspectez visuellement les couvercles pour vous assurer que les surfaces ont l’air propres sans motifs ni particules de poussière sur la surface du verre. Répétez les étapes 2.1 à 2.4 si des motifs ou de la poussière sont détectés.
REMARQUE: Tester l’hydrophilie de la surface en trempant les couvercles traités dans l’eau et en les retirant verticalement. L’eau sur les couvertures traitées se retire sous forme de feuille uniforme pour former les anneaux de Young par rapport aux plaques de couverture non traitées qui forment des plaques. - Transférer le porte-couvercle dans un bécher avec de l’éthanol résistant à 200 et laver deux fois pendant 15 s pour équilibrer les surfaces en solvant organique. Transférer le porte-couvercle dans une solution d’éthanol à 200 épreuves avec 3 % d’APTES pendant 1 h à RT pour silaniser les couvercles (Figure 1D). Couvrir le bécher avec un film de paraffine.
REMARQUE: Les paramètres de dépôt APTES varient en fonction de la méthode de nettoyage de surface, de la teneur en eau du solvant, de la concentration APTES, des temps d’incubation et de la température de recuit. - Immergez le rack dans un bécher propre avec une solution d’éthanol à l’épreuve des 200. Répétez ce lavage trois fois pendant 15 s chacun (Figure 1E).
- Sécher les couvercles à l’aide d’azote (N2) gazeux à faible pression de sortie. Placez les couvercles dans un plat en polystyrène de 10 cm avec un morceau de film de paraffine posé à plat à l’intérieur. Assurez-vous que les couvercles sont secs et séparés (Figure 1F).
- Ajouter 100 μL de solution de 2 mg/mL de NHS-biotine dans du DMSO à quatre couvercles placés sur un film de paraffine. Placer un « sandwich » avec les quatre autres couvercles sur le dessus (deux couvercles se faisant face avec la solution de fonctionnalisation entre les deux) et incuber le plat pendant la nuit à 4 °C (Figure 1G).
REMARQUE: À 4 ° C, le réactif NHS est plus stable, ce qui facilite la fonctionnalisation uniforme de la surface. De plus, le sandwich conserve les réactifs. Évitez d’ajouter un excès de solution dans le sandwich, car il pourrait s’échapper et faire glisser les couvercles.
Jour 2:
- Retirer le plat de 4 °C et séparer les couvercles en sandwich. Orientez les glissières dans le rack avec la surface revêtue l’une vers l’autre comme illustré à la figure 2A. Lavez-les avec une solution d’éthanol à l’épreuve de 200 trois fois pendant 15 s chacun. Sécher avec du gazN2 et les placer dans un nouveau plat avec un film de paraffine à l’intérieur.
REMARQUE: L’orientation des couvercles comme indiqué permet d’identifier la surface fonctionnalisée. - Lavez les couvercles avec 1 mL de 1x PBS trois fois pour les équilibrer à la phase aqueuse. Ajouter 800 μL d’albumine sérique bovine (BSA) à 0,1 % dans 1x PBS (p/v) à chacune des lames de couverture et incuber à RT pendant 30 min pour passer la surface et bloquer la liaison non spécifique des réactifs de fonctionnalisation ultérieurs (Figure 2B).
- Après l’incubation, laver les couvercles trois fois avec 1 mL de 1x PBS. Ajouter 800 μL de 1 μg/mL de streptavidine dans 1x PBS à RT pendant 45 à 60 min pour fonctionnaliser les couvercles (Figure 2C).
REMARQUE: Conservez un couvercle sans streptavidine pour vérifier l’efficacité de la passivation (facultatif). Ajouter 10 nM de molécules biotinylées et image en utilisant des conditions expérimentales. Cette intensité de surface doit être proche du bruit sombre de la caméra. - Simultanément à l’étape 2.11, assemblez les sondes TGT (haut : brin inférieur au rapport molaire 1:1) à une concentration finale de 50 nM dans 100 μL de 1 M NaCl dans un tube PCR à l’aide d’un thermocycleur. Dissocier les brins à 95 °C pendant 5 min, et recuit progressivement en réduisant la température à 25 °C et en la maintenant pendant 25 min (Figure 2D). Évitez l’exposition prolongée des sondes TGT à la lumière.
- Après l’incubation de la streptavidine, utilisez 1x PBS pour laver les couvercles trois fois. Ajouter 100 μL des sondes TGT pré-assemblées à quatre des couvercles et faire des sandwichs en utilisant les 4 couvercles restants avec le côté fonctionnalisé faisant face aux sondes (huit surfaces nécessitent quatre tubes de sondes TGT hybridées). Couvrir avec du papier d’aluminium et incuber pendant 1 h à RT pour permettre à la sonde de se lier à la surface (Figure 2E).
- Après l’incubation, séparez les sandwichs et lavez les couvercles avec 1x PBS trois fois. Les surfaces TGT sont maintenant prêtes pour l’imagerie. Assemblez soigneusement les couvercles dans des chambres d’imagerie prénettoyées et ajoutez 1x PBS pour garder les surfaces hydratées (Figure 2F).
REMARQUE: Un assèchement excessif des chambres fissurera la surface. Empêcher le séchage des surfaces.
3. Préparation des cellules et coloration
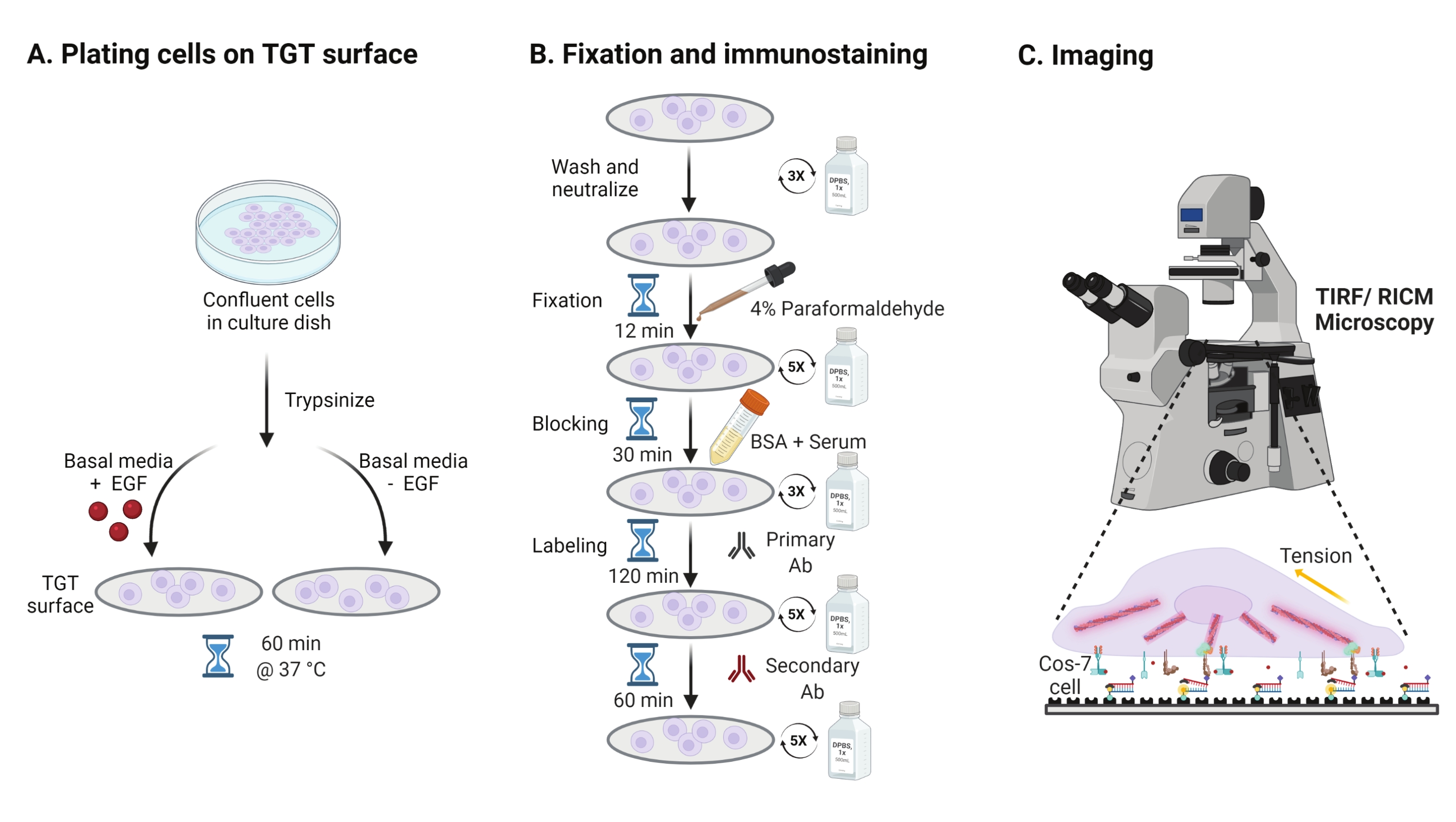
- Pour étudier l’effet de la stimulation du facteur de croissance épidermique (EGF) sur la mécanique du Cos-7, l’adhésion et la propagation cellulaire, trypsiniser les cellules Cos-7 avec 0,05% de trypsine-EDTA pendant 2 min. Neutraliser la trypsine en la lavant avec HBSS et en centrifugant à 800 x g pendant 5 min. Répétez l’étape de neutralisation une fois de plus.
- Cellules de plaque à une densité de 4 x 104 cellules sur les surfaces TGT assemblées dans le milieu Eagle modifié (DMEM) de Dulbecco complété par 50 ng / mL EGF ou DMEM sans EGF. Laisser les cellules se propager pendant 60 min à 37 °C avec 5 % de CO2 dans un incubateur de culture cellulaire (Figure 3A).
REMARQUE: Les cellules sont incubées dans le DMEM sans sérum pour éviter la stimulation des facteurs de croissance. L’EGF est dilué dans 10 mM d’acide acétique pour obtenir un stock de 1 mg/mL. Il est utilisé à 50 ng/mL dans le DMEM pour les expériences d’imagerie. - Après l’incubation, laver les cellules trois fois avec 1x PBS et fixer avec 2 mL de paraformaldéhyde à 4 % pendant 12 min à TA (Figure 3B).
REMARQUE: Toutes les étapes d’incubation sont effectuées sur un agitateur rotatif à ~ 35 tr / min pour une propagation uniforme des solutions. Protégez les surfaces TGT de la lumière en les recouvrant jusqu’à ce qu’elles soient prêtes pour l’imagerie. - Aspirer le fixateur et laver les couvercles cinq fois avec 1x PBS à des intervalles de 5 minutes à RT. Éventuellement, incuber les couvercles avec 50 mM NH4Cl dans 1x PBS pendant 30 min à 37 °C pour éteindre l’autofluorescence associée au paraformaldéhyde et laver trois fois avec 1x PBS à des intervalles de 5 min (Figure 3B).
- Ajouter le tampon A (1x PBS, 5% de sérum de cheval normal, 5% de sérum de chèvre normal, 1% de BSA, 0,025% de Triton X-100) et incuber pendant 30 min à 37 °C pour bloquer et perméabiliser les cellules. Lavez trois fois avec 1x PBS à des intervalles de 5 minutes (Figure 3B).
- Placez les chambres d’imagerie avec des couvercles dans un récipient d’humidité. Diluer l’anticorps anti-paxilline primaire (marqueur d’adhérence focale) à 1:250 dans un tampon bloquant (1x PBS, 5% de sérum de cheval normal, 5% de sérum de chèvre normal, 1% de BSA, 0,005% de Triton x-100). Incuber avec 200 μL de solution d’anticorps primaires par couvercle pendant 2 h à 37 °C (figure 3B).
REMARQUE: Ne laissez pas les surfaces sécher. - Lavez les couvercles cinq fois avec 1x PBS à des intervalles de 5 minutes et retournez-les dans le récipient d’humidité. Étiqueter les cellules simultanément avec un mélange d’anticorps secondaire anti-lapin conjugué au colorant à une dilution de 1:800 et de phalloïdine conjuguée au colorant (actine) à une dilution de 1:400 dans 200 μL de tampon bloquant par couvercle. Incuber à 37 °C pendant 60 min (Figure 3B).
- Lavez les surfaces cinq fois avec 1x PBS à des intervalles de 5 minutes et stockez-les à 4 °C jusqu’à ce qu’elles soient prêtes pour l’imagerie (Figure 3B).
REMARQUE: Échantillons d’image dans les 3 jours suivant la préparation de la surface pour éviter la dégradation du signal.
4. Acquisition d’images
- Utilisez un objectif 60x à immersion dans l’huile avec une ouverture numérique élevée (1,49) sur un microscope inversé avec une excitation TIRF 488, 561 et 647, une excitation RICM, un système de mise au point parfait et un appareil photo numérique.
- Ajoutez de l’huile à l’objectif, nettoyez le fond de la chambre d’échantillonnage et placez l’échantillon sur la scène. Concentrez-vous sur une cellule et engagez la concentration parfaite.
- Mettez le microscope en mode d’imagerie RICM avec excitation d’épifluorescence et un cube de filtre GFP avec le filtre d’émission retiré. Alignez le RICM en fermant et en centrant le diaphragme d’ouverture de l’épi-éclairage.
- Mettez le microscope en mode TIRF avec excitation laser et un cube de filtre TIRF à quatre passes. Concentrez le laser 488 nm sur un petit point au plafond de la pièce et augmentez l’angle d’incidence jusqu’à ce qu’il dépasse l’angle critique tout en surveillant la fluorescence sur la caméra en mode direct. Observez une forte réduction de la fluorescence de fond et un seul plan de mise au point lorsque l’angle critique est dépassé.
REMARQUE: TIRF excite une région mince (~ 100 nm) la plus proche de l’interface de glissement d’échantillon mettant en évidence les sondes TGT ouvertes et les adhérences focales tout en éliminant la fluorescence floue de l’intérieur de la cellule. Si le TIRF n’est pas disponible, l’épifluorescence peut être utilisée; cependant, les rapports signal/bruit seront plus faibles. - Identifiez les cellules pour l’imagerie à l’aide du mode « live » de la caméra à l’aide de RICM.
- Acquérir l’image RICM et les images TIRF de l’actine (640 nm ex), de la tension de l’intégrine (561 nm ex) et de la paxilline (488 nm ex). Obtenez des images séquentiellement en utilisant un temps d’exposition de 200 ms.
REMARQUE: Le temps d’exposition dépend de nombreux facteurs, notamment l’objectif, la puissance du laser, les filtres d’émission et la sensibilité de l’appareil photo. Le signal doit être au moins 2 fois supérieur à l’arrière-plan. L’arrière-plan est d’environ 1000 UA, de sorte que le signal doit être d’au moins 2000-3000 UA. - Répétez 4.4-4.5 pour au moins 30 cellules. Modifiez les capots, faites la mise au point et répétez 4.4-4.5.
5. Analyse des données
REMARQUE: Effectuer une analyse quantitative d’images à l’aide du logiciel Fidji et l’analyse à l’aide d’un logiciel de statistiques.
- Ouvrez le jeu d’images pour une cellule.
- Créez un masque de la zone de cellule (masque RICM) en traçant la limite de la cellule dans l’image RICM à l’aide de l’outil de sélection ImageJ Freehand. Enregistrez la région d’intérêt (ROI) dans le gestionnaire de retour sur investissement (Outils d’analyse > > gestionnaire de retour sur investissement) (Figure 4A1,2).
- Choisissez une zone représentative à l’extérieur de la cellule dans l’image de tension d’intégrine et dessinez un retour sur investissement d’au moins 200 x 200 pixels. Exclure toute autre cellule ou débris fluorescent du retour sur investissement. Mesurez la fluorescence de fond dans le retour sur investissement à l’aide de l’outil de mesure (Analyser > Mesure) (Figure 4A3).
- Soustrayez la fluorescence de fond moyenne obtenue à l’étape 5.2 de l’image de tension (Process > Math > Subtract) (Figure 4A4).
- Utilisez le masque RICM établi à l’étape 5.2 pour définir le signal de tension à l’intérieur de l’empreinte de la cellule (gestionnaire de retour sur investissement > Sélectionner > Appliquer le masque > Modifier > Effacer à l’extérieur) (Figure 4A5).
- Créez un masque de seuil pour l’image de tension à l’aide de la méthode de Huang pour le seuil automatique (Image > Ajuster > Seuil) (Figure 4A6). Assurez-vous que le masque de seuil représente le mieux la zone de tension d’intégrine générée. En règle générale, définissez le seuil sur 2x la fluorescence de fond moyenne.
- Créez une sélection du masque de tension seuillé (Modifier > Sélection > Créer) (Figure 4A7).
- Transférez le masque sélectionné sur l’image de tension générée à l’étape 5.4 et mesurez l’intensité intégrée des sondes ouvertes (gestionnaire de retour sur investissement > Select (masque de tension) > Analyser > Mesure > RawIntDen) (Figure 4C).
- Mesurez la zone des propriétés morphométriques de la cellule, la circularité et le rapport L/H à partir du masque RICM (gestionnaire de retour sur investissement > Sélectionner (masque RICM) > Appliquer le masque > Analyser > mesure) (Figure 4B).
- Mesurez la densité de rupture mécanique, définie comme le pourcentage de l’empreinte de la cellule avec les sondes rompues en sélectionnant l’image du masque de tension et en appliquant le masque RICM (roi manager > Select (RICM Mask) > Analyze > Measure > %Area) (Figure 4C).
- Exportez les mesures pour une analyse et une visualisation plus approfondies dans un logiciel de statistiques.
- Répétez 5.1-5.11 pour chaque cellule.
Résultats
Des sondes TGT activées ont été utilisées pour étudier l’effet du récepteur du facteur de croissance épidermique activé par ligand (EGFR) sur la mécanique cellulaire médiée par l’intégrine et la formation d’adhérence dans les cellules Cos-721. Les sondes présentent le ligand cyclique Arg-Gly-Asp-Phe-Lys (cRGDfK)21,23,25,26, qui est sélectif pour l’hétérodimère d’intégrine αVβ3 avec seulement une faible affinité pour les intégrines α5β1 27,28,29,30. La sonde TGT comprend un ADN duplex fonctionnalisé sur une surface de couverture en verre via le brin inférieur en utilisant la liaison biotine-streptavidine. Le brin supérieur affiche le ligand d’intégrine et est disponible pour se lier au récepteur de l’intégrine apparentée sur la membrane cellulaire (Figure 5A). Le brin inférieur est marqué avec un fluorophore et le brin supérieur avec un quencher, ce qui conduit à une fluorescence de fond minimale lorsque le TGT duplex est intact. Si une intégrine lie le ligand et applique une force d’une magnitude supérieureau T tol de la sonde, le duplex de l’ADN se séparera conduisant à la fluorescence (Figure 5A). Toute sonde TGT qui n’a pas été rompue par une force mécanique restera non fluorescente. Cette fluorescence d’activation sélective de force permet une cartographie systématique et quantitative des forces générées par l’intégrine à l’échelle pN à une résolution limitée par diffraction. Les sondes TGT modulent en outre le seuil de tension du substrat.
Voici un exemple représentatif d’une surface TGT avec un Ttol de 56 pN. Des cellules Cos-7 ont été plaquées sur cette surface TGT avec ou sans stimulation EGF pour étudier l’impact de l’activation de l’EGFR avec stimulation par ligand sur l’adhésion cellulaire et la mécanique de l’intégrine (Figure 5A, B). Les cellules ont été incubées avec ou sans EGF sur les surfaces TGT pendant 60 minutes, fixes et immuno-colorées pour afficher la distribution d’adhésion focale (paxilline) et l’organisation du cytosquelette (F-actine) (Figure 5B). Les cellules ont ensuite été imagées à l’aide de la microscopie RICM et TIRF. Comme on le voit clairement sur l’image RICM, l’étalement de cellules Cos-7 sur la surface TGT de 56 pN a été considérablement amélioré avec la stimulation EGF par rapport à sans stimulation. Cela a été quantifié pour 50 cellules dans chaque condition en mesurant la taille de la région de contact cellule-substrat à partir de l’image RICM (Figure 5C). La stimulation par l’EGF a entraîné une morphologie plus circulaire, représentative de la propagation et de la croissance des cellules Cos-7 dans leur environnement physiologique naturel (Figure 5D). La fluorescence des sondes ouvertes est également plus élevée avec la stimulation EGF comme observé dans l’image de fluorescence de tension. L’intensité intégrée des sondes ouvertes, qui est proportionnelle au nombre de sondes ouvertes, était beaucoup plus élevée avec la stimulation EGF que sans (Figure 5B,E). Il s’agit d’une représentation de tous les engagements récepteur-ligand où les intégrines ont appliqué une force supérieureà T tol (56 pN).
La coloration à la paxilline a montré que la distribution, le nombre, la maturation (taille) et l’organisation des adhérences focales étaient également influencés par la stimulation EGF. Les adhérences focales dans les cellules stimulées par l’EGF semblaient plus matures et orientées radialement par rapport à l’absence de témoins EGF. L’organisation du cytosquelette de la F-actine a également été améliorée par la stimulation EGF, évaluée par la coloration à la phalloïdine (figure 5B). Ces évaluations qualitatives ont été effectuées par comparaison visuelle d’images des deux groupes de traitement. L’analyse quantitative de l’adhésion focale peut être effectuée mais dépasse le cadre de ce protocole. Dans cette expérience, la surface TGT a fourni une plate-forme pour détailler systématiquement l’effet de l’activation de l’EGFR sur l’étalement cellulaire, la mécanique de l’intégrine et la formation d’adhérence focale.

Figure 1 : Schéma du jour 1 de la préparation de la surface TGT. (A) Nettoyez les couvercles. (B) Graver la surface du couvercle. C) Neutraliser la solution du piranha. (D) Silaniser la surface pour en faire des groupes amines réactifs. (E) Équilibrer les couvercles à la phase organique. (F) Sécher les couvercles avec un gaz inerte. (G) Biotinyler les groupes amines de surface. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2 : Schéma du jour 2 de la préparation de surface du TGT. (A) Nettoyez et séchez les couvercles pour éliminer toute biotine résiduelle de la veille. (B) Passivate avec BSA pour empêcher la liaison non spécifique du réactif dans les étapes ultérieures. (C) Fonctionnaliser les couvercles avec de la streptavidine. (D) Hybrider les sondes dans un thermocycleur. (E) Appliquer les sondes synthétisées sur les couvercles (F) Assembler les couvercles dans la chambre d’imagerie cellulaire. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 3 : Flux de travail général mettant en évidence les grandes étapes de l’ensemble de la configuration expérimentale. (A) Processus de détachement et de placage des cellules sur la surface TGT en milieu basal (DMEM) avec ou sans stimulation EGF. (B) Organigramme des étapes de la fixation et de l’immunocoloration post-fixation et de l’étalement sur la surface du TGT. C) Après la coloration, les cellules sont imagées sur un microscope à fluorescence inversée avec microscopie RICM et TIRF. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 4: Exemple de traitement de données et d’analyse quantitative. (A) Ventilation étape par étape du pipeline d’analyse utilisé aux Fidji (ImageJ) pour le RICM et la quantification de l’image de tension. (B) Un exemple représentatif des résultats morphométriques cellulaires analysés à l’aide du pipeline ci-dessus. (C) Exemples représentatifs de résultats mécaniques cellulaires analysés à l’aide du pipeline susmentionné. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 5 : Exemples de données issues d’une expérience TGT. (A) Diagramme mettant en évidence la zone de contact à l’interface de surface membrane cellulaire-TGT. Inset projette des intégrines interagissant avec son ligand apparenté cRGDfK avec (droite) ou sans (gauche) stimulation EGF. (B) Images RICM et TIRF de cellules Cos-7 étalées sur la surface TGT de 56 pN. Les images sont obtenues 60 min de post-placage avec ou sans stimulation EGF. Des images individuelles de RICM (telle qu’acquise), de tension d’intégrine (niveaux de gris), de paxilline (orange chaude) et de F-actine (bleu-vert) sont montrées avec des superpositions pour les deux conditions de stimulation. Barre d’échelle: 10 μm. L’encart met en évidence un ROI (région d’intérêt) zoomé détaillant la colocalisation de la tension d’intégrine générée aux sites de formation d’adhésion marqués par la paxilline, et l’organisation sous-jacente du cytosquelette subcellulaire marquée par l’actine. Barre d’échelle: 5 μm. (C-E) Nuages de points pour la zone d’étalement (empreinte cellulaire RICM) (C), la circularité (D) et la tension intégrée (E) pour les cellules Cos-7 avec ou sans stimulation EGF. Les barres indiquent la moyenne ± s.d. Les différences entre les groupes ont été évaluées statistiquement avec le test t de Student; P < 0,0001. n = 50 cellules dans trois expériences indépendantes. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 6 : Exemple de surfaces TGT avec différents problèmes possibles. (A) Images de tension et RICM d’une surface TGT idéale avec sonde assemblée trempée avant l’adhésion de la cellule. (B) Images de tension et RICM d’une surface TGT où la sonde TGT n’a pas le brin supérieur (quencher). L’image de tension montre une fluorescence uniforme du fluorophore ouvert dans le brin inférieur. (C) Images de tension et de RICM pour les cellules réparties sur une surface TGT idéale. (D) Images de tension et de RICM pour les cellules réparties sur une surface TGT mal faite avec une passivation limitée ou une sonde dégradée. (E) Les images de tension, de RICM et de fond clair pour les cellules plaquées sur une surface idéale avec un ligand cRGDfK indiquant les interactions cRGDfK-intégrine sont vitales pour la fixation cellulaire et la génération de tension. (F) Images de tension, de RICM et de champ lumineux pour les cellules plaquées sur une surface sans ligand cRGDfK sur le TGT. Alors que les cellules sont visibles dans l’image en champ lumineux, aucune fixation cellulaire ou tension d’intégrine générée n’est observée. Barre d’échelle: 10 μm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Discussion
Avec la procédure détaillée étape par étape décrite ci-dessus, on peut préparer des surfaces TGT pour quantifier la morphologie cellulaire et la tension d’intégrine générée par les cellules adhérentes lors de la fixation cellulaire et de la propagation après un traitement par EGF. La conception et la synthèse simples de la sonde et la préparation de surface ainsi que la configuration expérimentale simple ont fourni une plate-forme stable pour étudier l’interaction de l’EGFR et des intégrines. Dans l’ensemble, les résultats valident que l’activation de l’EGFR dépendante du ligand améliore la propagation cellulaire, ajuste les propriétés de force des récepteurs de l’intégrine et favorise l’organisation et la maturation de l’adhésion focale. Les résultats obtenus à l’aide de sondes TGT soutiennent l’hypothèse générale selon laquelle les facteurs de croissance, tels que l’EGFR, agissent comme des « mécano-organisateurs », augmentant la quantité et l’organisation spatiale de la tension de l’intégrine et régulant l’orientation et la mécanique des adhérences focales.
Lors de l’application sur la surface du TGT, les cellules atterrissent, se fixent et se propagent lorsque les récepteurs de l’intégrine (αVβ3) détectent et se lient au ligand cRGDfK. Ce faisant, les sondes TGT peuvent être rompues mécaniquement, générant une fluorescence sur le site d’engagement du ligand. La lecture est l'« historique de force » cumulatif de la cellule interagissant avec la surface. Il existe des problèmes courants avec les surfaces TGT qui peuvent être présents lors de ces expériences. La fluorescence de fond de surface élevée (Figure 6A, B), l’aspect de surface inégal, l’incapacité des cellules à générer un signal de tension (Figure 6C, D) et l’échec de la propagation des cellules (Figure 6E, F) peuvent être dus à des lacunes techniques avec la sonde TGT ou la synthèse de surface. Les solutions à ces problèmes courants sont présentées dans le tableau 1.
La conception simple des sondes TGT fournit aux biologistes cellulaires un outil puissant pour étudier les résultats spécifiques de la signalisation du facteur de croissance et de l’intégrine de manière isolée sans interférence d’autres récepteurs de surface cellulaire en ne fournissant que des ligands et des stimulations spécifiques. De plus, les sondes TGT permettent d’étudier le seuil de tension soulignant les récepteurs individuels de l’intégrine pendant l’adhésion cellulaire à la sensibilité pN. Les approches alternatives ne permettent pas de rapporter les forces exercées par des récepteurs individuels à haute résolution spatiale dans des échantillons fixes31. La microscopie à force de traction n’est sensible qu’aux forces nN, un ordre de grandeur supérieur aux forces appliquées par les récepteurs individuels de l’intégrine15, et les sondes de tension moléculaire mesurent les forces pN, mais parce qu’elles sont réversibles, elles ne résistent pas solidement à la fixation. Pour ces raisons, les sondes TGT sont un outil attrayant pour étudier la mécanique des interactions facteur de croissance-intégrine.
Il existe plusieurs nuances techniques associées aux sondes TGT qui doivent être prises en compte avant de concevoir une expérience. L’image de tension est un instantané dans le temps, représentant l’historique de la force et non un indicateur des engagements récepteur-ligand à un moment donné. Étant donné que la génération de signaux dépend de la séparation des sondes, la fluorescence TGT résulte de sondes ouvertes qui ne sont pas sous tension active en raison de l’engagement récepteur-ligand. Cela signifie que la lecture de la tension d’intégrine obtenue sur la surface TGT est de nature historique et cumulative représentant où il y avait des forces supérieuresà T tol; les emplacements des forces actuelles du récepteur-ligand inférieuresà T tol ne sont pas signalés19,32. Parce que la rupture TGT entraîne la fin de l’engagement récepteur-ligand, la propagation cellulaire est due aux interactions intégrine-ligand qui subissent des forces inférieuresà T tol. L’utilisateur doit donc être prudent lors de la définition du temps post-placage pour estimer les résultats mécaniques associés aux adhérences à base d’intégrine. Enfin, la signification de Ttol doit être considérée. Les sondes TGT utilisées ici ont un Ttol de 56 pN, où Ttol est la force constante nécessaire pour rompre 50% des sondes lorsqu’elles sont appliquées pendant 2 s. Lorsque l’on considère des systèmes biologiques complexes, les TGT connaissent probablement une gradation de force hétérogène et diversifiée avec des dépendances temporelles variables. Si les TGT sont rompus par des forces supérieuresà T tol, la fluorescence serait une sous-estimation de la tension totale. Alternativement, les forces inférieuresà T tol appliquées pour des durées plus longues peuvent rompre un nombre de sondes similaire aux forces de seuil élevé appliquées pour des temps plus courts. Ces deux scénarios peuvent entraîner la même lecture de l’intensité de fluorescence, ce qui rend difficile la résolution de l’amplitude ou de la dynamique exacte de la tension à l’aide des sondes TGT33,34.
Dans l’ensemble, les évaluations de la tension de l’intégrine avec stimulation du facteur de croissance doivent être effectuées avec soin en concevant des expériences avec des contrôles internes, en comparant les profils d’étalement sur d’autres surfaces revêtues de matrice, en effectuant des évaluations parallèles de la fluorescence TGT dans les cellules en présence ou en l’absence de stimulation du facteur de croissance, et en utilisant des TGT avec différents Ttol . Les TGT permettent de quantifier le rôle de la signalisation du facteur de croissance dans la régulation de la mécanique des récepteurs de l’intégrine, de la dynamique d’adhésion focale et de la propagation cellulaire. Ce protocole peut être utilisé comme modèle pour de nombreuses expériences basées sur TGT utilisant des sondes avec différents Ttol, différents ligands, différents types de cellules ou différentes conditions de stimulation. Toutes les protéines d’intérêt peuvent être étiquetées après fixation, et tout type d’analyse quantitative d’image peut être mis en œuvre. En tant que tel, nous présentons un modèle pour de nombreuses expériences TGT.
L’utilisation de sondes TGT ne se limite pas à l’étude des intégrines, mais peut être étendue à un large éventail de récepteurs de membrane cellulaire à travers différents types de cellules en modifiant le ligand. Les sondes TGT ont été utilisées pour étudier le rôle des forces dans la régulation de diverses cascades de signalisation des récepteurs, y compris l’identification du rôle mécanique de la mécanique des récepteurs Notch dans le développement embryonnaire et la neurogenèse35, les forces médiant l’identification et l’internalisation des antigènes par les récepteurs des cellules B36, et la capacité de relecture mécanique des récepteurs de surface des cellules T pour détecter les changements dans les forces afin d’augmenter la force et la spécificité du transfert de signal37 . Ensemble, ces résultats mettent en évidence l’immense potentiel des sondes TGT dans divers contextes expérimentaux.
Déclarations de divulgation
Les auteurs ne déclarent aucun conflit d’intérêts.
Remerciements
Les auteurs tiennent à remercier les membres du laboratoire Mattheyses pour leurs discussions et critiques fructueuses. Nous reconnaissons le financement accordé à A.L.M. par NSF CAREER 1832100 et NIH R01GM131099.
matériels
| Name | Company | Catalog Number | Comments |
| (3-Aminopropyl)triethoxysilane | Millipore Sigma | 440140 | Surface Preparation |
| 3-hydroxypicolinic acid (3-HPA) | Millipore Sigma | 56197 | Maldi-TOF-MS matrix |
| Acetic Acid, Glacial | Fisher Scientific | A38S | Diluting EGF |
| Acetonitrile (HPLC) | Fisher Scientific | A998SK | Oligonucleotide Preparation |
| Alexa Fluor 488 Phalloidin | Cell Signaling Technology | 8878S | Immunocytochemistry |
| Ammonium Chloride | Fisher Scientific | A687 | Immunocytochemistry |
| Anti-Paxillin antibody [Y113] | Abcam | ab32084 | Immunocytochemistry |
| BD Syringes only with Luer-Lok | BD bioscience | 309657 | Surface Preparation |
| Bio-Gel P-2 | Bio-Rad | 1504118 | Oligonucleotide Preparation |
| Bovine Serum Albumin (BSA) Protease-free Powder | Fisher Scientific | BP9703100 | Surface Preparation |
| Cos-7 cells | ATCC | CRL-1651 | Cell Culture, Passage numbers 11-20 |
| Coverslip Mini-Rack, for 8 coverslips | Fisher Scientific | C14784 | Surface Preparation |
| c(RGDfK(PEG-PEG)), PEG=8-amino-3,6-dioxaoctanoic acid | Vivitide | PCI-3696-PI | Oligonucleotide Preparation |
| Cy3B NHS ester | GE Healthcare | PA63101 | Oligonucleotide Preparation |
| Dimethylformamide | Millipore Sigma | PHR1553 | Oligonucleotide Preparation |
| DMEM with L-Glutamine, 4.5g/L Glucose and Sodium Pyruvate | Fisher Scientific | MT10013C | Cell Culture |
| Epidermal Growth Factor human EGF | Millipore Sigma | E9644 | Cell Culture |
| Ethanol, 200 proof (100%) | Fisher Scientific | 22032601 | Surface Preparation |
| Falcon Standard Tissue Culture Dishes | Fisher Scientific | 08-772E | Surface Preparation |
| Fetal Bovine Serum | Fisher Scientific | 10-438-026 | Cell Culture |
| Flurobrite DMEM | Fisher Scientific | A1896701 | Cell Culture |
| Goat anti-Rabbit IgG (H+L) Cross-Adsorbed Secondary Antibody, Alexa Fluor 647 | Invitrogen | A-21244 | Immunocytochemistry |
| Goat Serum | Fisher Scientific | 16-210-064 | Immunocytochemistry |
| Hank’s balanced salts (HBSS) | Fisher Scientific | 14-170-161 | Cell Culture |
| Horse Serum | Fisher Scientific | 16050130 | Immunocytochemistry |
| Hydrogen Peroxide | Fisher Scientific | H325-500 | Surface Preparation |
| Nanosep MF centrifugal devices | Pall laboratory | ODM02C35 | Oligonucleotide Preparation |
| NHS-azide | Fisher Scientific | 88902 | Oligonucleotide Preparation |
| Nitrogen Gas Cylinder | Airgas | Surface Preparation | |
| No. 2 round glass coverslips - 25 mm | VWR | 48382-085 | Surface Preparation |
| Parafilm M Laboratory Film | Fisher Scientific | 13-374-10 | Surface Preparation |
| Paraformaldehyde 16% | Fisher Scientific | 50-980-487 | Immunocytochemistry |
| PBS, 1X | Fisher Scientific | 21-030-CV | Surface Preparation/Immunocytochemistry |
| Penicillin-Streptomycin (5,000 U/mL) | Fisher Scientific | 15-070-063 | Cell Culture |
| PYREX Low Form Griffin Beakers | Fisher Scientific | 02-540G | Surface Preparation |
| Sodium Ascorbate | Fisher Scientific | 18-606-310 | Oligonucleotide Preparation |
| Sodium Bicarbonate | Fisher Scientific | S233 | Oligonucleotide Preparation |
| Sodium Chloride | Fisher Scientific | BP358 | Surface Preparation |
| Streptavidin | Fisher Scientific | 434301 | Surface Preparation |
| Sulfo-NHS-LC-Biotin | Fisher Scientific | 21335 | Surface Preparation |
| Sulfuric Acid | Fisher Scientific | A300-500 | Surface Preparation |
| TEAA | Fisher Scientific | NC0322726 | Oligonucleotide Preparation |
| Triethylamine | Millipore Sigma | 471283 | Oligonucleotide Preparation |
| Trifluoroacetic Acid (TFA) | Fisher Scientific | PI28901 | Oligonucleotide Preparation |
| THPTA | Fisher Scientific | NC1296293 | Oligonucleotide Preparation |
| Triton X 100 Detergent Surfact Ams Solution | Fisher Scientific | 85111 | Immunocytochemistry |
| Water, DNA Grade, DNASE, Protease free | Fisher Scientific | BP24701 | Oligonucleotide Preparation |
| Equipment | |||
| Agilent AdvanceBio Oligonucleotide C18 column, 4.6 x 150 mm, 2.7 μm | Agilent | 653950-702 | Oligonucleotide Preparation |
| High-performance liquid chromatography | Agilent | 1100 | Oligonucleotide Preparation |
| Low Speed Orbital Shaker | Fisher Scientific | 10-320-813 | Immunocytochemistry |
| Matrix-assisted laser desorption/ ionization time-of-flight mass spectrometer (MALDI-TOF-MS) | Voyager STR | Oligonucleotide Preparation | |
| Molecular Probes Attofluor Cell Chamber | Fisher Scientific | A7816 | Surface Preparation |
| Nanodrop 2000 UV-Vis Spectrophotometer | Thermo Fisher | Oligonucleotide Preparation | |
| Nikon Eclipse Ti inverted microscope | pe Nikon | Microscopy | |
| Nikon Perfect Focus System | Nikon | Microscopy | |
| NIS Elements software | Nikon | Microscopy | |
| ORCA-Flash4.0 V3 Digital CMOS camera | Hamamatsu | Microscopy | |
| Quad band TIRF 405/488/561/647 cube | CHROMA | Microscopy | |
| RICM Cube | CHROMA | Microscopy | |
| SOLA v-nIR Light Engine | Lumencor | Microscopy | |
| Thermo Forma Steri Cycle 370 CO2 Incubator | Fisher Scientific | Cell Culture | |
| VWR 75D Ultrasonic Cleaner | VWR | 13710 | Surface Preparation |
| Data Analysis | Use | ||
| Fiji (Image J) | https://imagej.net/software/fiji/downloads | Quantitative Analysis | |
| Graph Pad Prism | Graph Pad | Statistical Analysis | |
| Oligo name | 5'modification/ 3' modification | Sequence (5' to 3') | Use |
| Alkyne-21-BHQ2 | 5' Hexynyl/ 3' BHQ_2 | GTGAAATACCGCACAGATGCG | Top strand TGT probe |
| 56 pN TGT | 5' Biosg/TTTTTT/iUniAmM | CGCATCTGTGCGGTATTTCACTTT | Bottom strand TGT probe |
| 12 pN TGT | 5' AmMC6/ 3' BioTEG | CGCATCTGTGCGGTATTTCACTTT | Bottom strand TGT probe |
Références
- Lim, C. -. G., Jang, J., Kim, C. Cellular machinery for sensing mechanical force. BMB Reports. 51 (12), 623-629 (2018).
- Moraes, C., Sun, Y., Simmons, C. A. (Micro)managing the mechanical microenvironment. Integrative Biology. 3 (10), 959-971 (2011).
- Vogel, V., Sheetz, M. P. Mechanical forces matter in health and disease. From Cancer to Tissue Engineering. Nanotechnology. , 233-303 (2010).
- Wang, J. H. C., Li, B. Mechanics rules cell biology. BMC Sports Science, Medicine and Rehabilitation. 2 (1), 16 (2010).
- Moeendarbary, E., Harris, A. R. Cell mechanics: principles, practices, and prospects. Wiley Interdisciplinary Reviews: Systems Biology and Medicine. 6 (5), 371-388 (2014).
- Streuli, C. H., Akhtar, N. Signal co-operation between integrins and other receptor systems. Biochemical Journal. 418 (3), 491-506 (2009).
- Chiasson-MacKenzie, C., McClatchey, A. I. EGFR-induced cytoskeletal changes drive complex cell behaviors: The tip of the iceberg. Science Signaling. 11 (515), (2018).
- Kechagia, J. Z., Ivaska, J., Roca-Cusachs, P. Integrins as biomechanical sensors of the microenvironment. Nature Reviews Molecular Cell Biology. 20 (8), 457-473 (2019).
- De Luca, A., et al. The role of the EGFR signaling in tumor microenvironment. Journal of Cellular Physiology. 214 (3), 559-567 (2008).
- Javadi, S., Zhiani, M., Mousavi, M. A., Fathi, M. Crosstalk between Epidermal Growth Factor Receptors (EGFR) and integrins in resistance to EGFR tyrosine kinase inhibitors (TKIs) in solid tumors. European Journal of Cell Biology. 99 (4), 151083 (2020).
- Eliceiri, B. P. Integrin and growth factor receptor crosstalk. Circulation Research. 89 (12), 1104-1110 (2001).
- Dan, L., Jian, D., Na, L., Xiaozhong, W. Crosstalk between EGFR and integrin affects invasion and proliferation of gastric cancer cell line, SGC7901. OncoTargets and Therapy. 5, 271-277 (2012).
- Giancotti, F. G., Tarone, G. Positional control of cell fate through joint integrin/receptor protein kinase signaling. Annual Reviews: Cell and Developmental Biology. 19, 173-206 (2003).
- Ricono, J. M., et al. Specific cross-talk between epidermal growth factor receptor and integrin alphavbeta5 promotes carcinoma cell invasion and metastasis. Cancer Research. 69 (4), 1383-1391 (2009).
- Polacheck, W. J., Chen, C. S. Measuring cell-generated forces: a guide to the available tools. Nature Methods. 13 (5), 415-423 (2016).
- Hang, X., et al. Nanosensors for single cell mechanical interrogation. Biosensors and Bioelectronics. 179, 113086 (2021).
- Style, R. W., et al. Traction force microscopy in physics and biology. Soft Matter. 10 (23), 4047-4055 (2014).
- Schoen, I., Hu, W., Klotzsch, E., Vogel, V. Probing cellular traction forces by micropillar arrays: contribution of substrate warping to pillar deflection. Nano Letters. 10 (5), 1823-1830 (2010).
- Ma, V. P. -. Y., Salaita, K. DNA Nanotechnology as an Emerging Tool to Study Mechanotransduction in Living Systems. Small. 15 (26), 1900961 (2019).
- Kim, Y., Kim, K. A., Kim, B. C. Double-stranded DNA force sensors to study the molecular level forces required to activate signaling pathways. Journal of the Korean Physical Society. 78 (5), 386-392 (2021).
- Rao, T. C., et al. EGFR activation attenuates the mechanical threshold for integrin tension and focal adhesion formation. Journal of Cell Sciences. 133 (13), (2020).
- Zhang, Y., Ge, C., Zhu, C., Salaita, K. DNA-based digital tension probes reveal integrin forces during early cell adhesion. Nature Communications. 5, 5167 (2014).
- Liu, Y., et al. DNA-based nanoparticle tension sensors reveal that T-cell receptors transmit defined pN forces to their antigens for enhanced fidelity. Proceedings of the National Academy of Sciences of the United States of America. 113 (20), 5610-5615 (2016).
- Zhang, Y., et al. Platelet integrins exhibit anisotropic mechanosensing and harness piconewton forces to mediate platelet aggregation. Proceedings of the National Academy of Sciences of the United States of America. 115 (2), 325-330 (2018).
- Ma, V. P. -. Y., et al. Mechanically induced catalytic amplification reaction for readout of receptor-mediated cellular forces. Angewandte Chemie International Edition. 55 (18), 5488-5492 (2016).
- Wang, X., Ha, T. Defining single molecular forces required to activate integrin and notch signaling. Science. 340 (6135), 991-994 (2013).
- Chen, Y., Lee, H., Tong, H., Schwartz, M., Zhu, C. Force regulated conformational change of integrin αVβ3. Matrix Biology. 60-61, 70-85 (2017).
- Kantlehner, M., et al. Surface coating with cyclic RGD peptides stimulates osteoblast adhesion and proliferation as well as bone formation. ChemBioChem. 1 (2), 107-114 (2000).
- Kapp, T. G., et al. A comprehensive evaluation of the activity and selectivity profile of ligands for RGD-binding integrins. Scientific Reports. 7, 39805 (2017).
- Kok, R. J., et al. Preparation and functional evaluation of RGD-modified proteins as alpha(v)beta(3) integrin directed therapeutics. Bioconjugate Chemistry. 13 (3), 128-135 (2002).
- Li, I. T. S., Ha, T., Chemla, Y. R. Mapping cell surface adhesion by rotation tracking and adhesion footprinting. Scientific Reports. 7 (1), 44502 (2017).
- Wang, Y., et al. Force-activatable biosensor enables single platelet force mapping directly by fluorescence imaging. Biosensors and Bioelectronics. 100, 192-200 (2018).
- Murad, Y., Li, I. T. S. Quantifying molecular forces with serially connected force sensors. Biophysical Journal. 116 (7), 1282-1291 (2019).
- Yasunaga, A., Murad, Y., Li, I. T. S. Quantifying molecular tension-classifications, interpretations and limitations of force sensors. Physical Biology. 17 (1), 011001 (2019).
- Luca, V. C., et al. Notch-Jagged complex structure implicates a catch bond in tuning ligand sensitivity. Science. 355 (6331), 1320-1324 (2017).
- Spillane, K. M., Tolar, P. B cell antigen extraction is regulated by physical properties of antigen-presenting cells. The Journal of Cell Biology. 216 (1), 217-230 (2017).
- Brockman, J. M., Salaita, K. Mechanical proofreading: a general mechanism to enhance the fidelity of information transfer between cells. Frontiers in Physics. 7, 14 (2019).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.