
Method Article
מדידה וניתוח של תאי ייצור חומצה לקביעת glycolytic דרג
In This Article
Summary
Glycolysis is a defining metabolic marker in multiple biological systems. Monitoring glycolysis by measuring the extracellular flux of H+ is common, but requires correction to be quantitative and unambiguous. Here, we demonstrate how to gather and correct extracellular flux data to distinguish between respiratory and glycolytic sources of extracellular acidification.
Abstract
Extracellular measurement of oxygen consumption and acid production is a simple and powerful way to monitor rates of respiration and glycolysis1. Both mitochondrial (respiration) and non-mitochondrial (other redox) reactions consume oxygen, but these reactions can be easily distinguished by chemical inhibition of mitochondrial respiration. However, while mitochondrial oxygen consumption is an unambiguous and direct measurement of respiration rate2, the same is not true for extracellular acid production and its relationship to glycolytic rate 3-6. Extracellular acid produced by cells is derived from both lactate, produced by anaerobic glycolysis, and CO2, produced in the citric acid cycle during respiration. For glycolysis, the conversion of glucose to lactate- + H+ and the export of products into the assay medium is the source of glycolytic acidification. For respiration, the export of CO2, hydration to H2CO3 and dissociation to HCO3- + H+ is the source of respiratory acidification. The proportions of glycolytic and respiratory acidification depend on the experimental conditions, including cell type and substrate(s) provided, and can range from nearly 100% glycolytic acidification to nearly 100% respiratory acidification 6. Here, we demonstrate the data collection and calculation methods needed to determine respiratory and glycolytic contributions to total extracellular acidification by whole cells in culture using C2C12 myoblast cells as a model.
Introduction
המטרה הכוללת של שיטה זו היא למדוד את קצב glycolytic של תאים באמצעות ניתוח שטף תאי מדויק. מדידת כמותית של שיעור glycolytic באמצעות החמצה תאית היא נקודת הסיום הרצויה של ניסויים רבים. עם זאת, השיעור הכולל של החמצה תאית הוא הסכום של שני מרכיבים: החמצת נשימה, בצורה של CO 2 (שמוסיף נוזלים לH 2 CO 3 אז dissociates לHCO 3 - + H +), והחמצת glycolytic, בצורה של חומצת חלב - + + H.
התרומות של CO 2 להחמצה תאית כוללת שעד לאחרונה נחשבו זניחות בפלטפורמת המדידה המשמשת כאן, מנתח XF24 7. עם זאת, ברור במספר רב של מערכות אחרות שCO 2 יכולים להיות תורם עיקרי להחמצת 4-5 תאיים. ניירות מרובים להכיר קון זהtribution, אבל אל תנסה לכמת ישיר של CO 2 -derived 3,8,9 חומצה. לאחרונה הפגינו כמותית שCO 2 ייצור הוא מקור משמעותי של החמצה תאית במערכת זו 6. יתר על כן, למרות שיש מספר רב של מסלולי מטבוליים שיוצרים CO 2 מפירוק גלוקוז, המתבצע על ידי dehydrogenases מטריצה במעגל החומצה הציטרית הם התורמים המכריע וכל המקורות האחרים לייצר כמויות של CO 2 הנמצאות בניסוי השגיאה 6.
ללא תיקון לCO 2 ייצור, החמצה תאית לכן מחוון משמעי של שיעור glycolytic ולא ניתן להשתמש בו כמותית. הפרסום הקודם שלנו מדגיש כמה מקרים שבם הנשימה CO 2 כוללים את חלק הארי של כוללת אות החמצה, גם בתאים האמינו בדרך כלל להשתמש גליקוליזה 6 בעיקר. בנוסף,CO 2 תרומת נשימה להחמצה כוללת משתנית מאוד במהלך ניסויי פרופיל מטבולים משותפים, הוכחה כי השוואה נכונה של שיעור glycolytic במהלך חלקים שונים של ניסוי דורשת תיקון לCO 2.
כדי למדוד את קצב glycolytic של תאים תוך שימוש בשיעור של החמצה תאית, יש צורך להמיר שינויי pH לשינויים בסך הכל + H שנוצרו, וכדי להפחית את החמצה תאית הנגרמת על ידי CO 2 שפורסמה במהלך פעולה של מעגל החומצה הציטרית. כאן אנו מתארים שיטה פשוטה למדידת קצב ייצור פרוטון תאי (משינויים תאיים בpH והכוח המכויל החציצה של מדיום assay) ו- CO 2 ייצור (משינויים תאיים בO 2 ריכוז), ולהדגים כיצד לחשב את שיעור glycolytic באמצעות מדידות אלה.
כוח בשיטה זו ens השירות של מדידת החמצה תאית על ידי השימוש בו כדי לחשב כראוי שיעור glycolytic כפי שהוגדר בייצור חומצת החלב. ללא תיקון לCO 2 הנשימה (או מדידה ישירה של חומצת החלב), שאי אפשר לקבוע אם ובאיזו מידה הכוללת שיעור החמצה משקף שיעור glycolytic, בלבול הפרשנות של ניסויים המשתמשים בהחמצה תאית כוללת, כפי שמדידה ישירה של ייצור חומצת החלב.
חישובים
CO 2 וחומצת חלב הם, בתוך שגיאה ניסיונית, שני התורמים רק לייצור חומצה תאי, המבוססים על ניסויים עם תאים והיצמדות למנגנון 6. לכן, שיעור החמצה מוחלטת תאית (PPR, קצב ייצור פרוטון) יכול להיות מוגדר כ:
טוט PPR משוואת glyc = PPR שו"ת + PPR 1
. _content "> בי טוט = כולל; שו"ת = נשימה; = PPR glycolytic glyc glycolytic היא כך:glyc PPR = PPR טוט - PPR שו"ת משוואה 2
כאן,
טוט טוט = ECAR PPR / BP משוואה 3
שם ECAR = שיעור תאי החמצה (MPH / min), וBP = כוח באגירה (MPH / H pmol + 7 μl), ואילו
PPR שו"ת = (10 1 / (1 + 10 pH-PK 1) pH-PK) (H מקסימום + / O 2) (OCR טוט - OCR ריקבון / MYX) משוואה 4
כאשר K 1 = קבוע שיווי משקל משולב של CO 2 לחות וניתוק לHCO 3 - + H +; המקסימום H + / O 2 = ההדואר CO 2 החמצת -derived לשינוי מטבולים מסוים כגון חמצון של גלוקוז 6 מלא; = שיעור OCR חמצן הצריכה (O pmol 2 / min), וריקבון OCR / MYX = OCR לא המיטוכונדריה.
משוואה 4 מבודדת OCR המיטוכונדריה על ידי הפחתת כל OCR לא המיטוכונדריה (מוגדר כOCR כי הוא עמיד לrotenone המיטוכונדריה הנשימה הרעלים וmyxothiazol) ומהווה H המרבי + שנוצר לO 2 נצרך לכל מצע (H מקסימום + / O 2 ) (ראה 6), כמו גם את חלקם של CO 2 והוליד + H בטמפרטורה הניסיונית וpH (1 / (1 + 10-PK pH pH 10-PK 1). לחמצון מלא של גלוקוז, חמצן המיטוכונדריה שיעור הצריכה (OCR) הוא בדיוק שווה לשיעור של CO 2 ייצור. במרותק נפח assay של מדידת שטף תאית, CO 2 תוצרתד על ידי נשימה נשאר לכודים במדיום assay. רוב CO 2 לכודים הוא התייבשות לH 2 CO 3, אשר לאחר מכן dissociates לHCO 3 - + H +. חלק קטן נשאר מומס אבל לא התייבשות, ועוד חלק קטן הוא התייבשות אבל לא ניתק, כפי שהוכתב על ידי thermodynamically קבוע שיווי משקל בשילוב של CO 2 לחות וניתוק לHCO 3 - + H + בטמפרטורת ניסוי (37 ° C) ו- pH (~ 7.4).
לפיכך, המשוואה המלאה לחישוב g PPR על ידי הפחתת שו"ת PPR מטוט PPR הוא:
טוט PPR glyc = ECAR / BP - (PK-pH 1 / (1 + 10 pH-PK 1) 10) (H מקסימום + / O 2) (OCR טוט - OCR ריקבון / MYX) משוואה 5
אניn בדרך זו, שיעורי נשימה וגליקוליזה, כמו גם שיעורי ייצור ATP הקשורים, ניתן לקבוע כמותית ממדידות פשוטות (צריכת חמצן, החמצה תאית, קיבולת חציצה) ויבוא או חישוב ערכים הנדרשים אחרים (H + / O 2 , P / O, ושיווי המשקל K הקבוע 1) 6. הניסוי המתואר כאן מרחיב על טכניקות סטנדרטי לשימוש תאי שטף Analyzer כגון סוסון הים XF24 10,11; לפורמטים אחרים מדידת שטף תאית (למשל, דואר XF 96, או XFP), כל הכרכים שלהלן יש לשנותם כראוי.
הכח באגירה של מדיום assay ניתן למדוד על ידי הבנייה של עקומה סטנדרטית במישרין בפלטפורמת השטף תאית או בנפרד באמצעות בדיקה pH מכוילת. הנה, שלוש אפשרויות למדידת חציצה על ידי מדיום assay שטף תאי מקבלות, כולל שימוש בכל להזריקביציאות של מנתח השטף תאי עם בארות תא ללא מדגם, או באמצעות יציאת הזריקה האחרונה רק בבארות המכילות תא (סעיף 1) או באמצעות מדידה חיצונית pH (סעיף 2). ראה הגיליון האלקטרוני המצורף לחישובי הנתונים מלאים של דוגמא.
כדי למדוד את כוח החציצה באמצעות יכולת זיהוי-pH של מכשיר השטף תאי, זה בטוח להשתמש בארות תא ללא למזער וריאציה אות. עם זאת, בתוך השגיאה, אין הבדל סטטיסטי קיים בין ובארות בעת ביצוע מדידה זו המכילה תא-תא ללא (מידע לא מוצג). הערה: הווריאציה המתוארת בשלב 1.7 נושאת את היתרון של טיפול חשבונאי בשינויי פוטנציאל אגירה שמעניק תרכובות נוספות או על ידי הנוכחות של תאים, עם החסרון של אות רועשת. עם זאת, כאמור לעיל, לא נמצאו הבדלים משמעותיים בכח באגירה מחושב בין עיצוב התא ללא מוצג בטבלה 1 והעיצוב שלאחר הניסוי בטבלה 2 תחת תנאי הניסוי המתוארים כאן.
בנוסף, על טווחים קטנים ΔpH (<0.4 יחידות; בניסוי הטוב ביותר המוגבלים ל -0.2 יחידות), המדרון ליניארי שהתקבל על ידי התוויית קמ"ש / H pmol Δ + כראוי קרוב היחסים בין לוגריתמי ΔpH ו[ + H]. השיפוע של עקומת סטנדרט זה ולכן מייצג את הכח באגירה של מדיום assay הנבדק ב+ H pH / nmol ב -7 μl, או + H קמ"ש / pmol ב -7 μl. אנו ממליצים להגדיל את כוח חציצה בינוני או הפחתת צפיפות תא לדגימות שתעלינה על שינוי יחידת 0.2 pH בזמן המדידה. זמן המדידה גם עשוי להיות ירידה, אבל זה עשוי לקצר את שיעור החמצת מצב היציב ולהציג את השגיאה בחישוב השיעור.
Protocol
1. מדידת חציצת כוח בתאי שטף מכשיר: שתי וריאציות
הערה: החישובים ושיטות שתוארו כאן פותחו באמצעות תאי שטף Analyzer. למכשירים אחרים, מדידת הנפח חייב להיות מדורגים כראוי.
- הכן 0.1 M HCl הסטנדרטי במים באמצעות להתרכז HCl (ראה חומרים וציוד) לפי הוראות יצרן.
הערה: חישוב דוגמא להכנת זריקות HCl לשימוש בכל ארבע יציאות ההזרקה מוצג בטבלה 1:

טבלת 1. זריקות HCl ברציפות לassay שטף תאי גם.
- הכן דילולים של תקן HCl במדיום להיות assayed כמו בטבלה 1 למספר חזרות הטכניות להיות קארמטען בצלחת אחת, ועוד אחד כדי לאפשר לשגיאת pipetting; למשל, לארבע חזרות טכניות של נמל הזרקה, להכין מלאי של (1.1 x μl 5) 0.1 M HCl ב( 48.9 μl x 5) בינוני assay.
- הפץ 50 μl לכל יציאה של מחסנית הבדיקה מדידה. חזור על תהליך זה עבור יציאות שנותרו B, C, ו- D.
- הפעל assay תאי שטף 10 עם מחזור כיול סטנדרטי, ואחריו שני מחזורים של [תערובת 2 דקות, לחכות 1 דקות, ו -5 דקות מדידה] לכל אחד מארבע תוספות הנמל (ראה איור 2).
- לתכנת את הניסוי מעל בתוכנת המכשיר בהתאם להוראות תוכנה. טען את המחסנית מוכנה למכונה ולבצע כיול על פי הוראות תוכנה.
- כאשר תתבקש על ידי התכנית, להסיר את הצלחת המכילה calibrant והכנס את הצלחת המכילה מדיום assay בכל טוב למכשיר; להמשיך בתכנית.
- U לשיר הממוצע של 8-10 נקודות נתונים המתקבלות במצב יציב (בדרך כלל 8-10 הנקודות האחרונות) מלפני ואחרי כל יציאה בנוסף, לחשב את ההבדל (במצטבר) בpH (ΔpH) הנגרם על ידי כל הזרקה של חומצה סטנדרטית.
- העלילה ΔpH נגד H nmol + כלול ב7 μl לכוד על ידי הבדיקה מדידת הנפח. המדרון ליניארי הוא הכח באגירה (BP) ב+ H קמ"ש / pmol.
- לחלופין לצעדים 1.2-1.3, לבצע מדידות ΔpH הבאות assay שבנמלים, B, ו- C משמשים לביצוע ניסוי, ואחרי הזרקת HCl בנמל ד כמו בטבלה 2, ארבע חזרות הטכניים המשמש לייצור כל נקודה של עקומה סטנדרטית של 5 נקודות ב -20 הבארות הניסיוניות (לא כולל בארות תיקון טמפרטורת ארבעה רקע) של צלחת assay שטף תאית.
53464table2.jpg "/>
טבלת 2. זריקה אחת HCl לassay שטף תאי גם.
2. מדידת חציצת כוח שימוש חיצוני pH מטר
הערה: כדי למדוד את הכח באגירה של מדיום באמצעות בדיקה pH חיצונית, לכייל את הבדיקה על 37 מעלות צלזיוס ולשמור על טמפרטורה זו לכל המגיבים במהלך הניסוי.
- הכן 0.1 M HCl הסטנדרטי במים באמצעות להתרכז HCl על פי הוראות יצרן.
- בדיקה חמה pH, תקני ה- pH, בינוני assay החציצה שהכח יימדד, ו0.1 M HCl עד 37 מעלות צלזיוס באמבט מים.
- לכייל בדיקה pH על 37 מעלות צלזיוס בהתאם להוראות יצרן. לשמור את כל חומרים כימיים על 37 מעלות צלזיוס לאורך כל assay באמצעות צלחת חום או אמבט מים.
- Aliquot 10 מיליליטר של מדיום assay לתוך כוס קטנה או צינור חרוטי. צג ה- pH ברציפות באמצעות בדיקה pH שקועה.
- להוסיף 0.1 M HCl כל אומר בינוני ב10-20 aliquots μl.
- להבטיח ערבוב באמצעות סרגל מערבבים או על ידי מתערבל את המכל באופן ידני לאחר כל הוספת חומצה.
- לאפשר כמה שניות למדידת pH לייצוב, אז להקליט את ה- pH לאחר כל ההוספה.
- כפי שמודגם בטבלה 3, לעשות מספר מספיק של תוספות כדי להבטיח חישוב שיפוע מדויק וכדי לכסות את טווח pH הצפוי במהלך הניסוי.

טבלה 3. מדידת כוח החציצה באמצעות מד pH. הנתונים מייצגים ניסוי טיפוסי עם שש 20 תוספות μl של 0.1 M HCl.
- העלילה ΔpH נגד + H nmol הוסיפה ל7 μl, נותנת מדרון ליניארי שמייצג את הכח באגירה (איור 1).
.within עמודים = "תמיד"> 
איור 1. קביעת כוח החציצה. עקומה סטנדרטית HCl נמדדת כבטבלת 1, טבלה 2 או (כאן) כמו בטבלה 3. השיפוע של העקומה ליניארית בכושר נותן כוח באגירה (pH / H nmol + 7 μl). כל נקודה מייצגת ממוצע ± SEM של n = 9 משכפל טכני.
- ברגע שהכח באגירה ידוע באמצעות שיטת 1 או 2 לעיל, להמיר את אות ECAR (MPH / min) למקום מגורי הקבע (H + / דקות / חלבון מיקרוגרם pmol) על ידי חלוקת ECAR ידי חציצת כוח (BP) (MPH / + H pmol) ומדרוג לתכולת החלבון של כל טוב:
טוט (H pmol + / דקות / חלבון מיקרוגרם) PPR = ECAR / BP (MPH / H pmol + 7 μl) / חלבון בכל טוב (מיקרוגרם) משוואת 6 (MPH / min) - לחלופין, להשתמש באותה שיטה בניסוייםשל 1 או 2 כדי לחשב את ערך קיבולת חציצה (BC) המשמש את תוכנת המכשיר כדי לחשב באופן אוטומטי PPR במהלך איסוף הנתונים.
הערה: המדריך למשתמש מכשיר 12 (עמוד 107) מספקת מידע מפורט על חישוב ושימוש בקיבולת חציצה, שבו לפני הספירה מתוארת כ
לפנה"ס (mol / L) = H שומות + / (ΔpH חיץ x נפח (L)) משוואת 7
הערה: קיבולת החציצה כהגדרתו במשוואה 7 ניתן לחשב במבחני המכשיר או בדיקה חומציות חיצונית שתוארו לעיל. המרה בין כוח באגירה וקיבולת חציצה נעשה בקלות (ראה גיליון מצורף):
BC = 1 x 10 -9 / BP ((MPH / H pmol + 7 μl) / 7 μl) משוואה 8
הערה: אם ידוע לפני ביצוע assay, קיבולת החציצה ניתן להזין ישירות לתוך תוכנת המכשיר במהלך התקנה ניסיונית. - החל הליך זה והחישובים בשימוש מעל למרבית מערכות חיץ קונבנציונליות, כפי שתואר בפרסום קודם 6.
הערה: טבלה 4 רשימות הכח באגירה וקיבולת חציצה של כמה כלי תקשורת קונבנציונליות.

לוח 4. כוח החציצה וקיבולת חציצה של המדיה שנבחרה.
3. ביצוע תאי שטף Assay באמצעות תאי C2C12 Myoblast
הערה: בשלב 3.4.3, לא היו הבדלים שנצפו בCO 2 -derived ייצור חומצה תלוי בנוכחות של קרבוניק אנהידראז בתרבות C2C12, המצביע על כך את נוכחותה אינה נדרשת להמרה מלאה של CO 2 לHCO 3 - + H +. עם זאת, באופן אמפירי בדיקה זו במערכות ניסיוניות שונות מומלצת לפני אוםקרבוניק אנהידראז התלוש.
- תרבות העכבר C2C12 myoblasts 13 על 37 מעלות צלזיוס מתחת 95% אוויר / 5% CO 2 במדיום השתנה Dulbecco הנשר (DMEM) עם 11.1 גלוקוז מ"מ, 2 גלוטמין מ"מ, 10 V% / V בסרום שור עוברי (FBS), 100 U / פניצילין מיליליטר ו -100 מיקרוגרם / מיליליטר סטרפטומיצין.
- 24 שעות לפני assay, תאי צלחת / זרע מאותו מדיום התרבות 100 μl ב20,000 תאים / גם בצלחת 24 גם קלקר תאי שטף assay (ראה חומרים ושיטות) ללא ציפוי נוסף.
- לדלל oligomycin, FCCP, וrotenone תוספת myxothiazol, וHCl (אופציונאלי) ל10x ריכוז סופי בקרבס רינגר פוספט HEPES (KRPH) בינוני assay (2 מ"מ HEPES, 136 מ"מ NaCl, 2 מ"מ לאא 2 PO 4, 3.7 מ"מ KCl, 1 מ"מ MgCl 2, 1.5 מ"מ CaCl 2, 0.1% w / v אלבומין חומצות שומן חופשי שור בסרום, pH 7.4 על 37 מעלות צלזיוס).
- הכנת תא
- 30 דקות לפני assay, לשטוף תאים חסיד שלוש פעמים על ידי aspirating לGEntly להסיר את המדיום מן הבאר ולאחר מכן הוספת 500 KRPH μl לאט.
- דגירה תאים לאחר השלב השלישי לשטוף על 37 מעלות צלזיוס באוויר (לא מתחת לגיל 5% CO 2, שישנה את ה- pH של מדיום ללא יקרבונט זה).
- בתחילת assay, להחליף KRPH בבארות עם KRPH הטרי 500 μl מכיל קרבוניק אנהידראז 500 U / ml וגם גלוקוז (10 מ"מ) או בינוני בלבד, ללא מצע נוסף.
- טעינת מחסנית החיישן
- Pipet 50 aliquots μl של כל מתחם 10x מוכן בשלב 3.3 ליציאות מחסנית של מחסנית תאית שטף חיישן כדלקמן (ריכוזים סופיים בassay ניתנו גם): נמל: 2 מיקרוגרם / מיליליטר oligomycin, נמל B: 0.5 מיקרומטר FCCP, נמל C : Rotenone 1 מיקרומטר, myxothiazol 1 מיקרומטר, הנמל D: HCl (אם ביצוע כיול חומצה ב- assay כפי שתואר לעיל ובטבלה 2).
הערה: לצורך עיכוב שרשרת נשימה מלא מתואר כאן, 1 מיקרומטרxothiazol ניתן להשתמש לסירוגין עם א antimycin 1 מיקרומטר
- Pipet 50 aliquots μl של כל מתחם 10x מוכן בשלב 3.3 ליציאות מחסנית של מחסנית תאית שטף חיישן כדלקמן (ריכוזים סופיים בassay ניתנו גם): נמל: 2 מיקרוגרם / מיליליטר oligomycin, נמל B: 0.5 מיקרומטר FCCP, נמל C : Rotenone 1 מיקרומטר, myxothiazol 1 מיקרומטר, הנמל D: HCl (אם ביצוע כיול חומצה ב- assay כפי שתואר לעיל ובטבלה 2).
- assay שטף תאי:
- בצע assay שטף תאי סטנדרטי לקביעת שליטה בדרכי הנשימה כפי שתואר ב 10.
הערה: לכל קטע של הניסוי, לקבוע את התמהיל, לחכות, ופעמים מדידה רצויה, כמו גם את מספר המחזורים במגזר.
הערה: הנתונים בלוח 5 נאספו על שני מחזורי assay של 2 דקות לערבב, לחכות 1 דקות, ו -5 דקות מדד לכל מגזר, עם שלושה מחזורי assay מתרחשים לאחר תוספת הנמל D של כמויות שונות של HCl (לכיול של חציצה כוח כמו בטבלה 2).

לוח 5. תצורת assay שטף תאית.
4. Measuring נקודת סיום ריכוז חומצת חלב
הערה: כדי לאמת את assay העקיף שתואר כאן בכמה מערכת שונה, ריכוז חומצת החלב נקודת סיום בסוף ניסוי שטף תאי ניתן לקבוע ישירות בצלחת 96-היטב קונבנציונלית על ידי מדידת המהירות ההתחלתית (מעל 2 דקות) של הפחתה NAD + → NADH מזורז על ידי דהידרוגנז חומצת החלב, שתואר בפירוט בפרסום הקודם שלנו 6. לנתונים שהוצגו בנציגי תוצאות, ריכוז חומצת החלב נקודת סיום בבארות assay המכיל גלוקוז היה ~ 40 מיקרומטר.
- הכן בינוני הידרזין 2x: 1 M טריס, 20 מ"מ EDTA, 400 מ"מ הידרזין, pH 9.8 על 22 מעלות צלזיוס). מייד לפני תחילת assay, להוסיף NAD + עד 4 מ"מ ודהידרוגנז חומצת החלב (LDH) 40 U / ml. הרכב בינוני assay סופי (1x): 500 מ"מ טריס, 10 mM EDTA, 200 מ"מ הידרזין, 2 מ"מ NAD +, 20 LDH / מיליליטר U.
- מייד לאחר assay השטף תאי, להסיר 100μl של מדיום assay מכל טוב של הצלחת תאי שטף assay והעברה לבאר אטום (שחור) צלחת 96-היטב.
- לכל דגימה היטב, להוסיף 100 μl 2x בינוני הידרזין.
- מייד לטעון את הצלחת לקורא microplate ולהתחיל ניטור הקרינה NADH בעירור 340 ננומטר / 460 פליטת ננומטר.
- רשום את המהירות הראשונית לכ -2 דקות.
- להפעיל ניסוי דומה לבניית עקומת סטנדרט ידי התוויית מהירות ראשונית נגד ריכוז חומצת החלב לריכוזי חומצת החלב הוסיפו 0-50 מיקרומטר.
- חישוב ריכוז חומצת החלב בכל ניסוי גם באמצעות העקומה סטנדרטית.
5. מדידת תכולת חלבון
- הסר בינוני assay שנותר מכל טוב של צלחת assay.
- לשטוף בארות שלוש פעמים עם 250 μl KRPH ללא BSA, להיות זהיר, כדי למזער את דחיקת רגליהם של תאים מפני השטח והתחתון.
- להוסיף תמוגה Ripa 25 μlהבינוני (150 מ"מ NaCl, 50 מ"מ טריס, 1 מ"מ EGTA, 1 mM EDTA, 1% V / V Triton X-100, 0.5% w / v deoxycholate נתרן, 0.1% נ / SDS נ ', pH 7.4 על 22 מעלות צלזיוס) היטב בכל צלחת assay.
- דגירה צלחת על קרח למשך 30 דקות.
- להתסיס את צלחת על שייקר צלחת ב1,200 סל"ד במשך 5 דקות.
- למדוד ריכוז חלבון על ידי שיטות סטנדרטיים, למשל, על ידי assay BCA, להבטיח כי הרכב מאגר תמוגה תואם את שיטת המדידה. תכולת החלבון בניסוי באיור 2 ~ 4 מיקרוגרם / טובה הייתה.
תוצאות
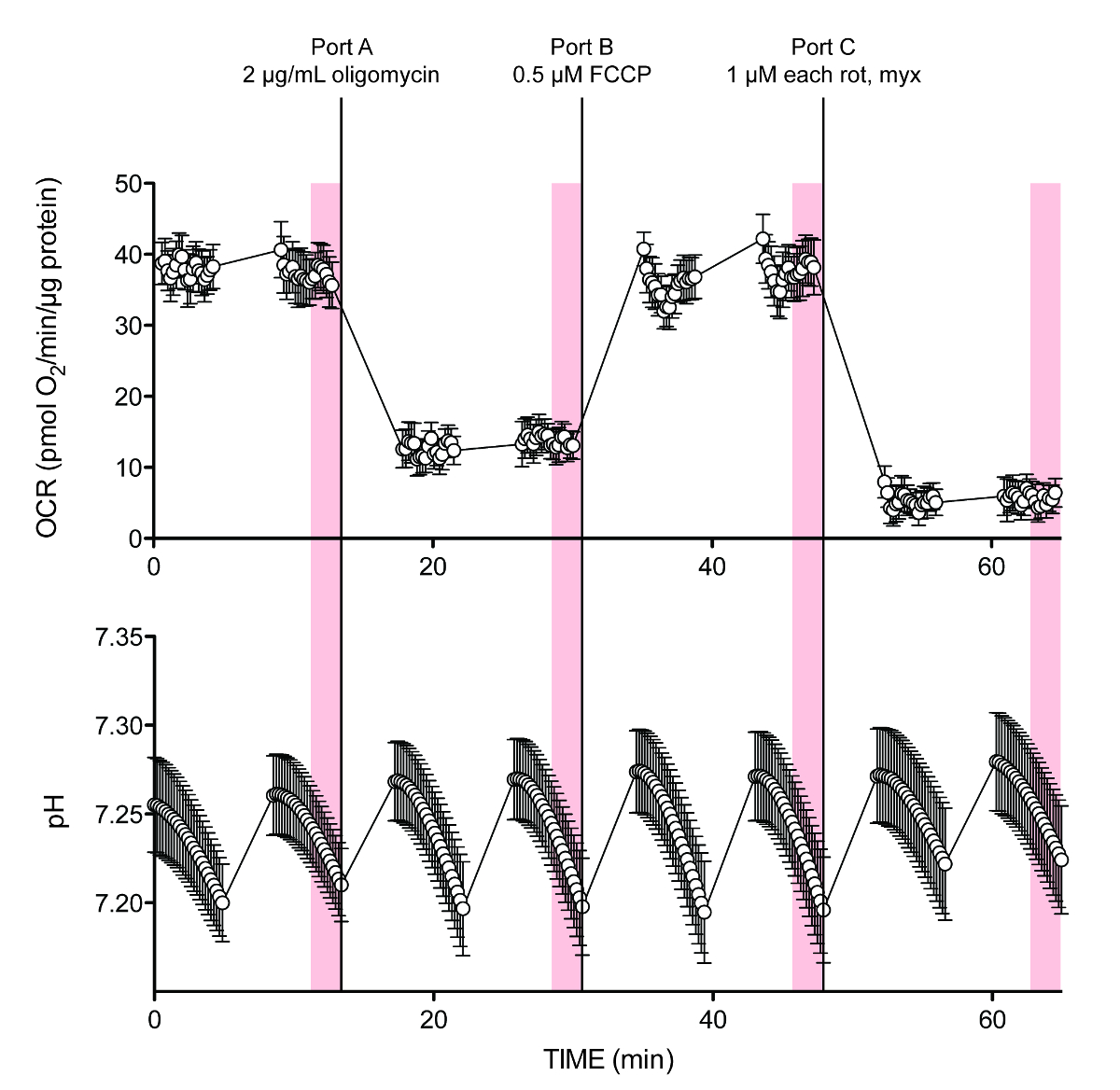
איור 2 מציג את הנתונים הגולמיים לניסוי טיפוסי. נקודות מדידת 10 שעברה מרישום נקודה לנקודה של שני OCR ו- pH (מוצל קווים אנכיים) שמשו לחישובים. חששות ראשוניים שהערך הממוצע (מדידת נקודת אמצע) של כל מחזור assay לא לספק רזולוציה מספקת של שיעור לחישוב מדויק, במיוחד כפי שיש שנראה כפיגור קל בין בנוסף נמל ושיעור החמצת מצב יציב, לא התממשו, כמו זה אינו מופיע לתרום באופן משמעותי לשגיאת חישוב (לא מוצג). לחלופין, אם קיבולת החציצה הנכונה הוא נכנס במהלך התקנה ניסיונית, ניתן לקרוא PPR ישירות מקריאת נתוני איסוף נתוני מכשיר על ידי הצגת פלט PPR בתוכנת המכשיר או במחשב האישי בפורמט תואם זמין כאחת מהגדרות פלט נתונים.
ftp_upload / 53,464 / 53464fig2.jpg "/>
איור 2. עקבות שטף תאי נציג של O 2 ו+ H. OCR ו- pH עוקב לניסוי הדוגמא בלוח 5, המכילים 10 מ"מ גלוקוז בתחילת assay. היה לי הנמל D ריכוזי HCl שונים לכיול של כוח באגירה (לא מוצג בממוצע עקבות אלה). נתונים מפרסום קודם 6. כל נקודה מייצגת ממוצע ± SEM של n = 8 חזרות ביולוגיות. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}
ניתוח נתונים של תוצאות נציג
שימוש בגיליון האלקטרוני מוצג בטבלה 6 ובתנאי כקובץ מצורף, נתונים ערכים מבארות בודדות ניתן להזין בעמודות מוצגות עם כותרות צהובות. כל שישה העמודים ל הזכות מחושבת מערכים אלה. הדוגמא בלוח 6 מציגה את החישובים של שו"ת PPR וglyc PPR באמצעות ECAR ונתונים OCR מבארות בודדות לתנאים המקומיים עם או בלי גלוקוז הוסיף, לפני יציאת תוספת של oligomycin. משכפל טכני על כל הכנה ביולוגית בדרך כלל בממוצע לתת ערכים יחידים של התפוקות בארבעת הטורים האחרונים, אז הנתונים מהכנות ביולוגיות שונות בממוצע עם התפשטות מתאימה לסטטיסטיקה שגיאה בBP וארבעת ערכים אלה.

חישוב לוח 6. של נשימה והחמצת glycolytic. עמודות בראשות בצהוב מצביעים על ערכים שנכנסו מחישוב (למשל, BP, מקסימום H + / O 2), או מאיסוף נתונים (למשל, טוט ECAR, OCR). נ.ב.: "target =" _ //www.jove.com/files/ftp_upload/53464/53464table6.jpg blank "> לחץ כאן כדי לצפות בגרסה גדולה יותר של שולחן זה |. אנא לחץ כאן כדי להוריד את הטבלה כגיליון אלקטרוני Excel .
תרומות של גליקוליזה ונשימה למקום מגורי הקבע לאחר תיקון
איור 3 מציג את הפלט הגרפי של נתונים מחושבים כבלוח 6 לשיעורי יליד החמצת glycolytic ונשימה, שיעורים הבאים oligomycin בנוסף (נמל), ושיעורים הבאים בנוסף FCCP (הנמל B). נתונים אלה מראים בבירור כיצד הפרופורציות של שינוי הנשימה וglycolytic החמצה עם בחירה של מצע (גלוקוז לעומת השליטה (CTL) עם אף הוסיפו) ועם מצב של המיטוכונדריה (פונקצית ילידים לעומת פונקציה פרמקולוגית שונה).
">
איור 3. שיעור פרוטון ייצור (PPR) ממקורות glycolytic ונשימה. PPR מהנשמה (חלקים פתוחים) וגליקוליזה (מנות מלאות) של תאי C2C12 מחושבים באמצעות משוואה 5 עם גלוקוז הוסיף (שלושה ברים שמאל) או בלי גלוקוז הוסיף (שלושה ברים מימין). נתונים מ 6. כל הנתונים מייצגים ממוצע ± SEM של n = 8 חזרות ביולוגיות.
Discussion
החמצה תאית היא אינדיקציה נמדדת בקלות של קצב חילוף חומרים תאי. כדי לקבוע כראוי השיעור של גליקוליזה הסלולרית (כהגדרתו על ידי ייצור חומצת החלב) זה קריטי לדעת את הכח באגירה של מדיום assay, ולהמיר את מדידות שטף תאית של צריכת חמצן והחמצה לפרוטון שיעורי ייצור. על ידי ביצוע חישוב זה, יכול להיות מופחת החמצה נובעת מCO 2 שוחרר במעגל החומצה הציטרית, עוזבת את החמצה שנובעת מייצור חומצת החלב.
הדרכים שונות מרובות הניתנות כאן כדי למדוד כוח באגירה לתיקון זה לשאת יתרונות וחסרונות שונים. מדידה חיצונית באמצעות בדיקה pH היא מדויקת מאוד לשחזור, אבל עלול שלא לשקף הבדלים קטנים בזיהוי pH הוצג על ידי fluorophores בתוך צלחת assay, התוספת של תרכובות בassay, או הנוכחות של tהוא התאים עצמם. מדידות pH לטפל בצלחת נושאים אלה, אלא גם להציג את הדרגות שונות של רעש ניסיוני.
CO 2 התיקון לECAR מאפשר בפעם הראשונה החישוב חד-משמעי וכמותי של שיעור glycolytic, ומגלה וריאציה בתרומת נשימה וglycolytic לECAR הכולל במהלך ניסוי. שימוש במשוואה 5 והמדידות של OCR, ECAR, וחציצת כוח, שיעור glycolytic יכול להיות מחושב באמצעות הגיליון האלקטרוני פשוט סיפק (לוח 6). שיעור זה יכול להיות מאומת על ידי מדידת חומצת החלב פוסט הוק-אם תרצה 6. בתאים שבו מסלול פנטוז פוספט הוא פעיל מאוד, השימוש במעכבי מסלול כגון 6-aminonicotinamide עשויים להיות שימושיים לבודד את שיעור glycolytic. חישוב התרומות של שני CO 2 - ו+ H-נגזר חומצת החלב מהסך נמדדו סלולרי במיוחד החמצת דרג וחמצן Consumptיון דרג הוא כלי רב ערך עבור שימוש בנתוני שטף תאיים להצהיר הצהרות חזקות וכמותי על פעילות המטבולית.
שימוש בהליכים שתוארו כאן, כולל שינויים שונים למדידת כוח באגירה, ואופטימיזציה של ניסוי השטף תאי לתאים תחת חקירה והנתונים הרצויים, השיעור של גליקוליזה בתאים שלמים ניתן לכמת במגוון רחב של תנאי ניסוי. שיטה זו מוגבלת לתאים שיכולים לגדול בתרבות חסיד ב( או תאים או אברונים שניתן דבקו) משטח קלקר. זה אמין ביותר כאשר תאים בתרבית הם הומוגנית ומחוברות, אם כי ייתכן שעדיין ניתן לקבל מידע שימושי על פני טווח של תנאים אלה. החישובים דורשים ידע מסוים בחילוף החומרים של התאים, כמו מקס H + / O 2 טווחים 0.65-1.0 לחמצון מלא של מצעים שונים ויותר לחמצון חלקי 6, לעומת זאת, אם התאים הם known לחמצן גלוקוז, ערך של 1.0 ניתן להניח.
למרות שרלוונטי לכל האפיון מטבולים, שיטה זו עשויה להיות מועילה במיוחד בעת שימוש במערכות שבמשמרת בין חילוף חומרים בדרכי הנשימה וglycolytic כדי לשמור על אספקת ATP הסלולרית היא פנוטיפ קריטי, כולל האפיון של תאי גזע ותאים סרטניים שמקורם בגידול. הבנת שינויי שליטת חילוף חומרים בהקשרים אלה ואחרים תאפשר מידה רבה יותר של תחכום ודיוק בעיצוב והניתוח הניסיוני של סוגי התאים.
Disclosures
ד"ר השונה Mookerjee מצהיר כי אין לה אינטרסים כלכליים מתחרים. ד"ר מרטין המותג התייעץ לסוסון הים Biosciences, אשר מייצר מכשירים וחומרים כימיים המשמשים בסעיף זה. דמי גישה פתוחים לכתבה זו מומנו על ידי Biosciences סוסון הים.
Acknowledgements
We thank David A. Ferrick and David G. Nicholls for contributing to project conception and presentation, Renata L.S. Goncalves and Akos A. Gerencser for data not shown here and for helpful discussions, Barbara Liepe for XF24 consumables, and Andy Neilson for input in developing Eq. (5).
Materials
| Name | Company | Catalog Number | Comments |
| Pherastar FS | BMG | n/a | microplate reader |
| Seahorse XF-24 | Seahorse Bioscience | n/a | extracellular flux instrument |
| Seahorse XF assay plate | Seahorse Bioscience | V7-PS | consumable |
| XF Calibrant | Seahorse Bioscience | 100840-000 | solution |
| HCl standard | Sigma | 38280 | chemical |
| oligomycin | Sigma | O4876 | chemical |
| FCCP | Sigma | C2920 | chemical |
| Rotenone | Sigma | R8875 | chemical |
| Myxothiazol | Sigma | T5580 | chemical |
| DMEM | Corning | 10-013-CV | medium component |
| FBS | Corning | 35-010-CV | medium component |
| penicillin/streptomycin | Corning | 30-002-CI | medium component |
| carbonic anhydrase | Sigma | C2624 | chemical |
| 96-well assay plate | Corning | CLS3991 | consumable |
| NAD+ | Sigma | N7004 | chemical |
| LDH | Sigma | L1254 | chemical |
References
- Brand, M. D., Nicholls, D. G. Assessing mitochondrial dysfunction in cells. Biochem. J. 435 (2), 297-312 (2011).
- Gerencser, A. A., et al. Quantitative microplate-based respirometry with correction for oxygen diffusion. Anal. Chem. 81 (16), 6868-6878 (2009).
- Divakaruni, A. S., Paradyse, A., Ferrick, D. A., Murphy, A. N., Jastroch, M. Analysis and interpretation of microplate-based oxygen consumption and pH data. Meth. Enzymol. 547, 309-354 (2014).
- Renner, K., Jansen-Dürr, P., Gnaiger, E. Biphasic oxygen kinetics of cellular respiration and linear oxygen dependence of antimycin A inhibited oxygen consumption. Mol. Biol. Rep. 29 (1-2), 83-87 (2002).
- Helmlinger, G., Sckell, A., Dellian, M., Forbes, N. S., Jain, R. K. Acid production in glycolysis-impaired tumors provides new insights into tumor metabolism. Clin. Cancer Res. 8 (4), 1284-1291 (2002).
- Mookerjee, S. A., Goncalves, R. L. S., Gerencser, A. G., Nicholls, D. G., Brand, M. D. The contributions of respiration and glycolysis to extracellular acid production. Biochim. Biophys. Acta. 1847, 171-181 (2015).
- Wu, M., et al. Multiparameter metabolic analysis reveals a close link between attenuated mitochondrial bioenergetic function and enhanced glycolysis dependency in human tumor cells. Am. J. Physiol. 292 (1), C125-C136 (2006).
- Nadanaciva, S., et al. Assessment of drug-induced mitochondrial dysfunction via altered cellular respiration and acidification measured in a 96-well platform. J. Bioenerg. Biomembr. 44 (4), 421-437 (2012).
- Pelletier, M., Billingham, L. K., Ramaswamy, M., Siegel, R. M. Extracellular flux analysis to monitor glycolytic rates and mitochondrial oxygen consumption. Meth. Enzymol. 542, 125-149 (2014).
- Nicholls, D. G., Darley-Usmar, V. M., Wu, M., Jensen, P. B., Rogers, G. W., Ferrick, D. A. Bioenergetic profile experiment using C2C12 myoblast cells. J. Vis. Exp. (46), 2511 (2010).
- Rogers, G. W., et al. High throughput microplate respiratory measurements using minimal quantities of isolated mitochondria. PLoS ONE. 6 (7), e21746 (2011).
- Seahorse Biosciences. . F24 Extracellular Flux Analyzer and Prep Station Installation and Operation Manual. 1.7, (2010).
- Blau, H., Chiu, C., Webster, C. Cytoplasmic activation of human nuclear genes in human heterocaryons. Cell. 32, 1171-1180 (1983).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved