
Method Article
세포 외 산 생산의 측정 및 분석은 당분 속도를 확인하는
요약
Glycolysis is a defining metabolic marker in multiple biological systems. Monitoring glycolysis by measuring the extracellular flux of H+ is common, but requires correction to be quantitative and unambiguous. Here, we demonstrate how to gather and correct extracellular flux data to distinguish between respiratory and glycolytic sources of extracellular acidification.
초록
Extracellular measurement of oxygen consumption and acid production is a simple and powerful way to monitor rates of respiration and glycolysis1. Both mitochondrial (respiration) and non-mitochondrial (other redox) reactions consume oxygen, but these reactions can be easily distinguished by chemical inhibition of mitochondrial respiration. However, while mitochondrial oxygen consumption is an unambiguous and direct measurement of respiration rate2, the same is not true for extracellular acid production and its relationship to glycolytic rate 3-6. Extracellular acid produced by cells is derived from both lactate, produced by anaerobic glycolysis, and CO2, produced in the citric acid cycle during respiration. For glycolysis, the conversion of glucose to lactate- + H+ and the export of products into the assay medium is the source of glycolytic acidification. For respiration, the export of CO2, hydration to H2CO3 and dissociation to HCO3- + H+ is the source of respiratory acidification. The proportions of glycolytic and respiratory acidification depend on the experimental conditions, including cell type and substrate(s) provided, and can range from nearly 100% glycolytic acidification to nearly 100% respiratory acidification 6. Here, we demonstrate the data collection and calculation methods needed to determine respiratory and glycolytic contributions to total extracellular acidification by whole cells in culture using C2C12 myoblast cells as a model.
서문
이 방법의 전체 목표는 정확하게 유속 세포 분석을 이용하여 세포의 당분 속도를 측정하는 방법이다. 세포 외 산성화하여 당분 레이트의 정량적 측정은 많은 실험을 원하는 종점이다. 그러나, 세포 외 산성화의 총 비율은 두 성분의 합이다 : 호흡기 산성화, CO (2 CO 3 다음 HCO 3 해리되어 H로 촉촉 - + H +) (2)의 형태로, 그리고 당분 산성화 형태 + 시간 + - 락 테이트.
총 세포 외 산성화에 CO 2의 기여는 최근까지 여기에 사용되는 측정 플랫폼, XF24 분석기 7 무시할 수있다. 그러나, 2는 세포 외 산성화 4-5의 주요 원인이 될 수있는 CO 여러 다른 시스템에서 명백하다. 다수의 논문이 사기꾼을 인정기여도를,하지만 산 3,8,9을 - 유래 이산화탄소의 직접 정량을 시도하지 않습니다. 우리는 최근에 이산화탄소 생산이 시스템 6 세포 외 산성화의 중요한 원천이다 정량적으로 보여 주었다. 포도당 분해 대사에서 CO 2를 생성 다중 대사 경로도 있지만 게다가, 사람들은 시트르산 사이클에서 매트릭스게나 의해 수행은 압도적 기여자와 모든 다른 소스는 실험 오차 6 내에 CO 2의 양을 생성한다.
이산화탄소 생산을 보정하지 않고, 세포 외 산성화 따라서 당분 속도의 모호한 지표 및 정량적으로 사용할 수 없습니다. 이전 공보 호흡기 CO 2에도 일반적으로 주로 당분 6을 사용하는 것으로 세포에서, 전체 산성화 신호의 대부분을 포함하는 여러 인스턴스를 강조한다. 또한,총 산성화 호흡기 CO 2 기부는 실험의 다른 부분 동안 당분 레이트 정확한 비교 CO 2 보정이 필요하다는 보여주는 공통 대사 프로파일 실험 과정 중에 광범위하게 변한다.
세포 외 산성화 속도를 이용하여 세포의 당분 속도를 측정하기 위해, 생성 된 총 H의 +의 변화 pH 변화로 변환하고, 시트르산 회로의 동작 동안 방출 CO 2에 의한 세포 외 산성화를 감산 할 필요가있다. 여기서는 (O 2 농도의 세포 외 변화로부터) 및 CO 2 제조 (pH의 외 변경 및 보정 분석 배지의 완충력부터) 외 양성자 생성 속도를 측정하기위한 간단한 방법을 설명하고, 당분 레이트를 계산하는 방법을 보여 이러한 측정을 사용.
이 방법은 강도 락 테이트 생산에 의해 정의 된대로 올바르게 당분 속도를 계산하는 데 사용하여 세포 외 산성화 측정 유틸리티 ENS. 호흡기 CO 2 (또는 락트산의 직접 측정)에 대한 보정없이 그것이 판별 불가능하면과 총 산성화 레이트 락트산 생산 직접 측정으로 전체 세포 외 산성화를 이용해 실험 해석 교란, 당분 레이트를 반영 어느 정도이다.
용 계산
이산화탄소와 젖산은 실험 오차 내에서,이다, 근육 모세포의 세포 6 실험을 기반으로 세포 외 산 생산에 두 참여자,. 따라서, 총 세포 외 산성화 (PPR, 양성자 생산 속도)의 속도는 다음과 같이 정의 될 수있다 :
PPR의 TOT = PPR RESP + PPR의 glyc의 식 (1)
. _content "> TOT = 총, RESP = 호흡은, glyc = 당분 당분 PPR은 이렇게이다 :PPR의 glyc = PPR TOT - PPR RESP 식 (2)
이리,
中, TOT = ECAR의 TOT / BP 식 3
여기서 ECAR = 세포 외 산성화 속도 (MPH / 분), 및 BP = 완충력 (MPH / pmol의 H +의 7 μL에서), 반면
PPR RESP = (10의 pH - 피크 1 / (1 + 10의 pH - 피크 1)) (최대 H + / O 2) (OCR TOT - OCR 부패 / MYX) 식 (4)
여기서 K 1 = 이산화탄소 수화와 HCO 3 해리의 결합 평형 상수 - + H +; 최대 H + / O 2 = 일전자 CO 포도당 (6)의 완전 산화 같은 특정 대사 변환 2 유래 산성화; OCR = 산소 소비 속도 (pmol의 O를 2 / 분), 그리고 OCR 부패 / MYX = 비 미토콘드리아 OCR.
수학 식 4의 (미토콘드리아 호흡 독극물 로테 논 및 myxothiazol에 내성 OCR로 정의) 및 최대 H 차지 + 각각의 기판 (최대 H + / O 2 소비 O 2 당 생성 비 미토콘드리아 OCR을 감산함으로써 미토콘드리아 OCR을 분리 ()) 6 참조뿐만 아니라 실험 온도 및 산도 (10 산도 - 피크 1 / (1 + 10 산도 - 피크 1 H의 + 상승을주는 CO 2의 비율). 글루코스의 전체 산화 미토콘드리아 산소 소비율 (OCR)이 CO 2 생성 속도에 정확히 동일하다. 세포 광속 측정의 제한된 분석 부피, CO 2 농산물D는 호흡에 의한 분석 배지에 갇혀 유지된다. H + + - 포획 CO 2 대부분이어서 HCO 3 해리되어 H 2 CO 3로 수화. 작은 부분 용해 남아 있지만 수화하지 않으며, HCO 3 CO 2 수화와 해리의 결합 평형 상수에 의해 열역학적으로 지시로 다른 작은 부분은, 수화하지만 해리 아니다 - + H + 실험 온도 (37 ° C) 및 pH에서 (~ 7.4).
따라서, PPR의 TOT는 PPR로부터의 RESP를 감산하여 PPR g를 계산하기위한 완전한 방정식 :
PPR glyc = ECAR의 TOT / BP - (10 산도 - 피크 1 / (1 + 10의 pH - 피크 1)) (최대 H + / O 2) (OCR TOT - OCR 부패 / MYX) 식 (5)
나는N이 방법, 호흡과 당분,뿐만 아니라 연관된 ATP 생성 비율의 비율은, 정량적으로 직접적인 측정 (산소 소모, 세포 외 산성화, 완충 능력) 및 다른 필요한 값의 수입 또는 계산 (H + / O (2)로부터 결정될 수있다 , P / O, 및 평형 상수 K 1) 6. 여기에 설명 된 실험은 해마 XF24 10, 11 등의 세포 외 유출 분석기는 사용하기위한 표준 기술을 확장; 다른 세포 외 플럭스 측정 형식 (예를 들어, XF 전자 (96), 또는 XFP)를 위해, 아래의 모든 볼륨을 적절하게 조절해야한다.
분석 배지의 완충력은 교정의 pH 프로브를 사용하여 직접적으로 세포 외 플럭스 플랫폼에 개별적으로 또는 표준 곡선의 구성에 의해 측정 될 수있다. 여기서, 외 플럭스 분석 배지로 버퍼링을 측정하기위한 세 가지 옵션 모두를 사용 injecti 포함한 주어진다무 세포 샘플 웰, 또는 세포를 함유하는 웰 (제 1) 또는 외부 pH 측정 (제 2)를 이용하여 마지막 주사 포트를 사용하여 세포 외 플럭스와 분석기 포트. 예를 들어 데이터의 전체 계산에 부착 스프레드 참조.
세포 외 플럭스 인스트루먼트의 pH 검출 기능을 이용하여 버퍼링 전력을 측정하기 위해, 신호의 변화를 최소화하기 위해, 무 세포 웰을 사용하는 것이 안전하다. 그러나, 오차 내에 통계적 차이는 무 세포 및 세포 - 함유이 측정을 수행 할 때 우물 (데이터는 미도시) 사이에 존재하지 않는다. 주 : 단계 1.7에 기재된 변이는 잡음이 신호의 단점과 함께, 추가의 화합물 또는 세포의 존재에 의해 부여되는 버퍼링에 대한 전위 변화를 차지하는 장점을 운반한다. 그러나, 상기 한 바와 같이, 큰 차이는 표 1에 도시 된 무 세포 간의 설계 계산 완충력에서 발견되었고여기에 기술 된 실험 조건으로 표 2의 실험 후 디자인.
또한, 작은 ΔpH 범위 (<0.4 단위, 실험적으로 최고 0.2 단위로 제한)을 통해, 선형 기울기는 Δ의 MPH / pmol의 H를 플롯하여 얻은 + 적절하게 ΔpH와 [시간 +] 사이의 대수 관계가 가깝다. 이 표준 곡선의 기울기에 따라서 pH를 7 μL / nmol의의 H +의 피 시험 분석 배지의 완충력을 나타내고, 또는 마일 / pmol의 H 종 + 7 μL이다. 우리는 중간 버퍼링 전력을 증가 또는 측정 시간 동안 0.2의 pH 변화 부 초과 샘플 세포 밀도를 감소시키는 추천. 측정 시간은 또한 감소 될 수 있지만, 이는 정상 상태의 산성화 속도를 단축 율 계산에 오류를 초래할 수있다.
프로토콜
1. 세포 외 유출 악기에 버퍼링 전력을 측정 : 두 개의 변주
참고 : 계산 및 여기서 설명하는 방법은 세포 외 유출 분석기를 사용하여 개발되었다. 다른 악기를 들어, 측정 량을 적절히 스케일링되어야한다.
- 제조업체의 지침에 따라 (재료 및 장비를 참조) 염산 농축액을 사용하여 물에 0.1 M 표준 염산을 준비합니다.
주 : 모든 네 주입구에 사용 염산 주사를 준비하기위한 예시적인 계산은 표 1에 도시된다 :

표 아니라 세포 외 플럭스 분석에 1. 연속 염산 주사.
- 매체에 염산 표준 희석액을 제조는 카 인 기술의 복제 수는 표 1에서와 같이 분석 될한 판에서 IED, 더하기 하나는 피펫 오류 수 있도록, 예를 들어, 포트의 네 가지 기술 복제 주입 (48.9 μL × 5) 분석 매체 (1.1 μL × 5) 0.1 M 염산의 재고를 준비합니다.
- 측정 프로브 카트리지의 각 포트에 50 μl를 배포합니다. 나머지 포트 B, C 및 D에 대해이 절차를 반복
- 네 개의 포트 가산의 각 [2 분간 혼합, 1 분간 대기 한 5 분의 측정]의 2 사이클이어서 표준 교정 사이클 외 플럭스 분석 (10), 실행 (도 2 참조).
- 소프트웨어 명령어에 따라 구 소프트웨어에서 상기 실험 프로그램. 기계로 제조 된 카트리지를로드하고 소프트웨어 지침에 따라 보정을 수행합니다.
- 프로그램에 연결되면, calibrant 함유 플레이트를 제거하고 잘 악기로 각각 분석 매체를 포함하는 플레이트를 삽입; 프로그램을 계속.
- 유 앞에서 정상 상태에서 얻어 8-10 데이터 포인트 (전형적으로 80-10 마지막 점)의 평균과 각각의 포트 부를 첨가 한 후, 각각의 표준 산 주입으로 인한 산도 (ΔpH)에서 (누적)의 차이를 계산한다.
- nmol의 H에 대해 플롯 ΔpH + 측정 탐침 (7)에 의해 포획 μL 부피 함유 하였다. 선형 기울기는 MPH / pmol의 H 종 +에서 버퍼링 전원 (BP)입니다.
- 대안 단계 1.2~1.3로, 표 2와 포트 D.에서 HCl을 주입 하였다 포트가, B, 및 C는 실험을 행할 때 사용되는 분석 다음 ΔpH 측정을 수행하는 네 개의 기술적 복제는 세포 외 플럭스 분석 플레이트의 (4 배경 온도 보정 웰은 제외) 20 실험 웰에 5 점 표준 곡선의 각 점을 생성하는 데 사용된다.
53464table2.jpg "/>
잘 세포 외 플럭스 분석에 표 2. 단일 염산 주입입니다.
2. 외부의 pH 측정기를 사용하여 버퍼링 전력을 측정
주 : 외부의 pH 프로브를 사용하여 매체의 완충력을 측정 37 ℃에서 탐침을 보정하고 모든 시약 실험 동안이 온도를 유지한다.
- 제조업체의 지침에 따라 염산 농축액을 사용하여 물에 0.1 M 표준 염산을 준비합니다.
- 따뜻한의 pH 탐침, 기준 산도, 그 버퍼링 전력 측정하는 분석 배지, 수욕에서 37 ℃로 0.1 M 염산.
- 제조업체의 지침에 따라 37 ℃에서의 pH 프로브를 보정합니다. 열판이나 수조를 사용하여 분석에 걸쳐 37 ℃에서 모든 시약을 유지한다.
- 분취 작은 비커 또는 원추형 튜브에 분석 배지 10 ㎖. 지속적 몰입의 pH 프로브를 사용하여 pH를 모니터링합니다.
- 같은 0.1 M 염산 추가10 ~ 20 μL 분취 중간을 말한다.
- 교반 막대를 사용하거나 수동으로 각각의 산 부가 한 후 컨테이너를 소용돌이로 혼합해야합니다.
- pH 측정을 안정화시키기 위해 몇 초, 각 첨가 후 pH를 기록 할 수 있습니다.
- 표 3에서 증명 된 바와 같이, 추가의 충분한 수의 정확한 기울기 계산을 보장하고 실험 기간 동안 예상되는 pH 범위를 커버 할 수 있도록.

표 3. pH 미터를 이용하여 버퍼링은 전력을 측정. 데이터는 0.1 M HCl을 여섯 20 μL를 추가 전형적인 실험을 나타낸다.
- 을 nmol의 시간 +에 대한 플롯 ΔpH 버퍼링 전력 (그림 1)을 나타내는 직선의 기울기를주고, 7 μL 당 덧붙였다.

도 1 버퍼링 파워를 결정. 염산 표준 곡선은 표 3과 표 1, 표 2 (여기)와 같이 측정 하였다. 선형 피팅 곡선의 기울기는 완충력 (PH / nmol의 H +의 7 μL에서)를 제공한다. 각 점은 N의 ± SEM = 9 기술 복제를 의미 나타냅니다.
- 완충력이 방법 1 또는 상기 (2)를 통해 알려지면, 전력을 버퍼링하여 ECAR 나누어 PPR (pmol의의 H + / 분 / μg의 단백질)에 ECAR 신호 (MPH / 분)로 변환 (BP) (MPH / pmol의 H 종 +) 각 웰의 단백질 함량에 스케일링 :
PPR의 TOT (pmol의의 H + / 분 / μg의 단백질) = ECAR (MPH / 분) / BP (MPH / pmol의의 H + 7 μL에서) 잘 (μg의) 당 / 단백질 식 (6) - 대안 적 방법에서 동일한 실험을 사용S (1) 또는 (2)가 자동으로 데이터를 수집하는 동안 PPR를 계산하는 장비 소프트웨어가 사용하는 완충 능 (BC) 값을 산출한다.
참고 :이 장비는 사용자 설명서 12 (107 페이지)을 계산하고 BC는 다음과 같이 설명되어 완충 능력을 사용하는 방법에 대한 자세한 정보를 제공합니다
BC (몰 / L) = 몰의 H + / (ΔpH X 버퍼 용량 (L)) 식 (7)
주 : 수학 식 7에 정의 된 버퍼링 용량은 상기 외부 기기의 pH 프로브 분석에서 계산 될 수있다. 완충 능력과 완충 능력 사이의 변환을 쉽게 (첨부 된 스프레드 시트를 참조) 수행 :
BC = 1 × 10-9 / BP ((MPH / pmol의의 H + 7 μL에서) / 7 μL) 식 (8)
주 : 상기 분석을 수행하기에 앞서 공지 된 경우, 완충 능력이 실험 중에 설치 장비 소프트웨어에 직접 입력 할 수있다. - 이전 발행 6에 기술 된 바와 같이,이 절차 대부분의 기존의 버퍼 시스템 위에서 사용 된 계산을 적용합니다.
참고 : 표 4는 완충 능력과 여러 기존 미디어의 완충 능력.

표 4. 버퍼링 전력과 선택한 미디어의 완충 능력.
3. C2C12 근육 모세포의 세포를 사용하여 세포 외 유출 분석 실험을 수행
참고 : 단계 3.4.3에서, CO 2에는 관찰 된 차이는 존재가 HCO 3 CO 2의 완전한 전환을 위해 요구되지 않는다는 것을 시사 C2C12 문화에 탄산 탈수 효소의 존재에 산 생산에 의존 - 유래없는 있었다 - + H +. 그러나 경험적으로 다른 실험 시스템이 테스트를하는 것은 OM 전에 추천itting 탄산 탈수 효소.
- 배양 마우스 C2C12 11.1 mM의 글루코오스, 2 mM의 글루타민, 10 % V / V 소 태아 혈청 (FBS), 100 U 가진 둘 베코 변성 이글 배지 (DMEM)에서 95 % 공기 / 5 % CO 2 하에서 37 ℃에서 13 아세포 / ml의 페니실린과 100 μg의 / ㎖ 스트렙토 마이신.
- 24 시간 이전에 분석에 20,000 세포에서 같은 배양 배지 100 ㎕에 판 / 종자 세포 / 웰의 추가 코팅 24 웰 폴리스티렌 세포 외 플럭스 분석 플레이트 (재료 및 방법 참조).
- , FCCP을 oligomycin 희석하고, 크렙스 링거 인산 HEPES (KRPH) 분석 배지 (2 mM의 HEPES, 136 mM의 NaCl을, 2 밀리미터의 NaH 2 PO 4, 3.7 밀리미터의 KCl, 1 로테 플러스 myxothiazol, 10 배 최종 농도의 HCl (선택 사항) 밀리미터의 MgCl 2, 37 ° C에서 1.5 mM의 CaCl2를, 0.1 % V / w 지방산 무 소 혈청 알부민, pH 7.4의).
- 셀 준비
- 30 분 전에 분석에, GE에 흡입에 의해 부착 세포를 세 번 씻는다ntly 우물에서 매체를 제거하고 천천히 500 μL KRPH를 추가.
- 공기 하에서 37 ℃에서 세 번째 세척 단계 후, 세포를 부화 (되지 5 % 중탄산이없는 배지의 pH를 변경한다 CO 2 이하).
- 분석 시작시, 추가 기판, 500 U / ㎖ 탄산 탈수 효소 및 중 포도당 (10 mM)을 또는 유일한 매체를 포함하는 500 μL 신선한 KRPH와 우물에 KRPH를 교체합니다.
- 센서 카트리지 넣기
- 포트 : 2 μg의 / ㎖ oligomycin, 포트 B : 0.5 μm의 FCCP, 포트 C (분석에서 최종 농도가 아니라 주어진) 다음과 같은 세포 외 유량 센서 카트리지의 카트리지 포트에 단계 3.3에서 제조 된 각각의 10 배 화합물의 피펫 50 μL 씩 1 μM의 로테 논, 1 μM의 myxothiazol, 포트 D : 염산 (상기 표 2에 기재된 분석에서의 산 - 교정을 행하는 경우).
주 : 완전 호흡 쇄 억제 목적을 위해 여기에 설명 된, 제 1 μMxothiazol 1 μM의 antimycin의 A.와 상호 교환 적으로 사용될 수있다
- 포트 : 2 μg의 / ㎖ oligomycin, 포트 B : 0.5 μm의 FCCP, 포트 C (분석에서 최종 농도가 아니라 주어진) 다음과 같은 세포 외 유량 센서 카트리지의 카트리지 포트에 단계 3.3에서 제조 된 각각의 10 배 화합물의 피펫 50 μL 씩 1 μM의 로테 논, 1 μM의 myxothiazol, 포트 D : 염산 (상기 표 2에 기재된 분석에서의 산 - 교정을 행하는 경우).
- 세포 외 플럭스 분석 :
- 10에 기술 된 바와 같이 호흡 제어를 결정하기위한 표준 외 플럭스 분석을 수행한다.
참고 실험의 각 세그먼트에 대해, 혼합 결정 기다린 측정 시간 세그먼트 당 사이클뿐만 아니라, 숫자, 소망.
주 : 표 5의 데이터는 세 가지 분석 사이클 버퍼링 교정 용 염산 (상이한 양의 포트 D를 첨가 한 후 발생하여, 2 분간 혼합, 1 분간 대기하고, 각 세그먼트에 대한 5 분 계수의 두 분석 사이클 동안 수집 된 표 2에서와 같이 전원).

표 5. 세포 외 플럭스 분석 구성.
4. MeasuriNG 엔드 포인트 젖산 농도
주 : 환원 (2 분 이상)의 초기 속도를 측정함으로써 기존의 96 웰 플레이트에서 직접 결정될 수있다 외 플럭스 실험의 끝에서 약간 다른 시스템, 엔드 포인트 젖산 농도 여기서 설명 간접 분석법를 검증 NAD + → NADH는 당사의 사전 출판 (6)에 자세히 설명 젖산 탈수소 효소에 의해 촉매. 대표 결과에 제시된 데이터, 포도당 - 함유 분석 웰에서 종점 젖산 농도는 ~ 40 μM이었다.
- 배 히드라진 매체를 준비 : 1 M 트리스, 20 mM의 EDTA, 400 mM의 히드라진, pH가 9.8를 22 ° C에서). 즉시 분석을 시작하기 전에, 40 U / ㎖로 4 밀리미터 및 젖산 탈수소 효소 (LDH)에 NAD +를 추가합니다. 최종 분석 배지 조성물 (1X) : 500 mM 트리스, 10 mM의 EDTA, 200 mM의 히드라진, 2 mM의 NAD +, 20 U / ㎖의 LDH.
- 즉시 세포 외 플럭스 분석을 다음, 100을 제거불투명 (블랙)의 웰에 세포 외 플럭스 분석 플레이트 및 전사의 각 웰로부터의 분석 배지를 96- 웰 플레이트의 μL.
- 물론 각각의 샘플에 100 μL 배 히드라진 매체를 추가 할 수 있습니다.
- 즉시 마이크로 플레이트 리더로 플레이트를 로딩하고 340 nm의 여기 / 460 nm의 방출에서 NADH 형광 모니터링을 시작.
- 약 2 분의 초기 속도를 기록한다.
- 0 내지 50 μM 첨가 농도에 대한 락 테이트 락 테이트 농도에 대해서 초기 속도를 플로팅하여 표준 곡선을 구성하는 유사한 실험을 실행.
- 각각의 실험이 잘 표준 곡선을 사용하여 젖산 농도를 계산한다.
5. 단백질 함량을 측정
- 분석 플레이트의 각 웰에서 남은 분석 매체를 제거합니다.
- 바닥 아니라 표면에서 세포의 빠지을 최소화하기 위해주의하고, BSA없는 KRPH μL 우물 (250)로 3 회 세척 할 것.
- 25 μL RIPA 용해 추가매체 (150 mM의 염화나트륨, 50 mM 트리스, 1 mM의 EGTA, 1 mM의 EDTA, 1 % V / V 트리톤 X-100, 0.5 % V / W 나트륨 데 옥시 콜레이트, 0.1 % V / V SDS, 22 ° C에서의 pH 7.4) 분석 플레이트의 각 웰.
- 30 분 동안 얼음 접시를 품어.
- 5 분 동안 1,200 rpm에서 판 쉐이커에 판을 선동.
- 용해 완충액의 조성의 측정 방법과 호환되는 것을 보장 BCA 분석으로 예를 들어 표준 방법에 의해 단백질 농도를 측정한다. 도 2의 실험에서 단백질 함량은 4 ~ μg의 / 웰이었다.
결과
그림 2는 일반적인 실험에 대한 원시 데이터를 보여줍니다. OCR 및 산도 (수직선 음영)의 두 지점 간 기록의 마지막 10 측정 포인트는 계산을 위해 사용 하였다. 초기 문제는 각 시험주기의 평균값 (중간 점 측정), 확증되지 않은 포트 가산 및 정상 상태 산성화 속도 사이에 약간의 지연이있을 등장 특히로서, 정확한 계산 속도의 충분한 해상도를 제공하지 않을 것이라고 이는 계산 오차에 크게 기여하는 것으로 보이지 않는 (도시되지 않음). 버퍼링 용량은 실험 셋업 동안 입력되는 경우 선택적으로, PPR은 장비 소프트웨어 또는 데이터 출력 설정 중 하나로서 사용할 PC 호환 포맷 PPR 출력을 표시하여 기기의 데이터 수집 판독로부터 직접 판독 될 수있다.
ftp_upload / 53464 / 53464fig2.jpg "/>
그림 O 2 시간 + 2. 대표 외 플럭스 흔적. OCR 및 pH는 시험 시작 10 mM의 포도당을 포함, 표 5의 예 실험에 대한 추적합니다. 포트 D는 (이들 평균 트레이스에 도시되지 않음) 완충력 교정 용 다른 염산 농도를 가지고 있었다. 이전 발행 6의 데이터. 각 점은 N = 8 생물 복제의 ± SEM을 의미 나타냅니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
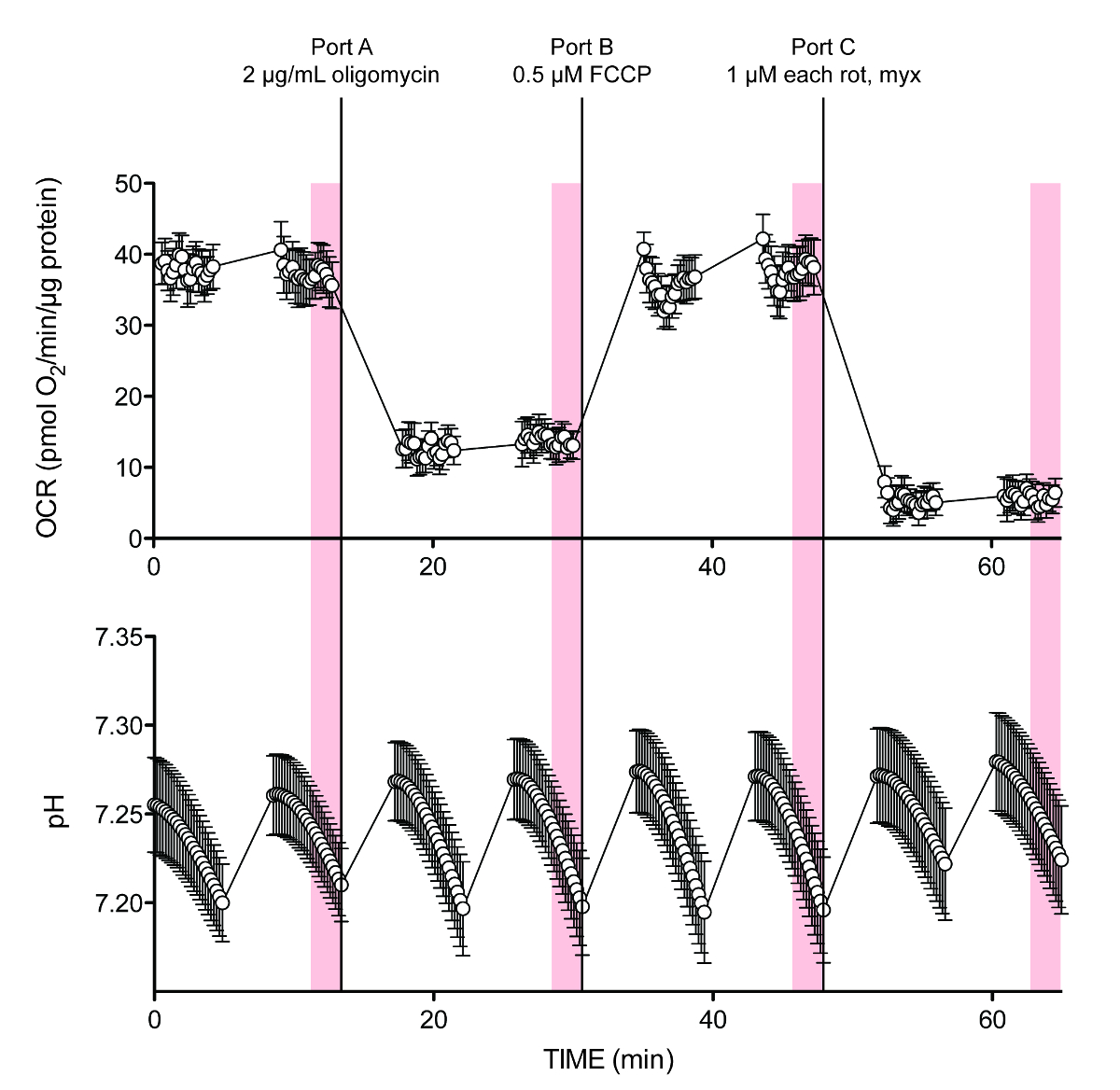{kind=link}
대표적인 결과의 데이터 분석
스프레드 시트의 표 6에 나타낸 바와 같이 부착을 제공하여, 각각의 웰로부터의 데이터 값은 황색 헤더 도시 칼럼에 입력 될 수있다. 여섯 컬럼 오른쪽은이 항목에서 계산됩니다. 표 6의 예는 PPR의 RESP 및 또는 추가 포도당없이 기본 조건, oligomycin의 사전 포트에 또한 개별 우물에서 ECAR 및 OCR 데이터를 사용하여 PPR의 glyc의 계산을 보여줍니다. 각각의 생물학적 제제에 기술 복제는 일반적으로 마지막 네 열 출력의 단일 값을 제공하기 위해 평균을, 그 다음 다른 생물학적 제제의 데이터는 오류 BP 통계 이러한 네 가지 값의 적절한 전파로 평균된다.

표 6. 호흡과 당분 산성화의 계산. 노란색으로 향했다 열 값이 계산에서 입력 할 수 표시 (예를 들어, BP, 최대 H + / O 2), 또는 데이터 수집에서 (예를 들어, ECAR 어린 아이, OCR). PS : //www.jove.com/files/ftp_upload/53464/53464table6.jpg "대상 ="_ 빈 ">이 테이블의 더 큰 버전을 보려면 여기를 클릭하십시오 |. 여기를 클릭하십시오를 Excel 스프레드 시트로이 표를 다운로드 .
보정 후 PPR에 해당 작용과 호흡의 기여
그림 3은 FCCP 추가 (포트 B) 다음과 같은 기본 당분과 호흡 산성화 속도, 또한 (포트) oligomycin 다음과 같은 요금 및 요금은 표 6과 계산 된 데이터의 그래픽 출력을 보여줍니다. 이러한 데이터는 명확하게 기판의 선택과 호흡과 당분 산성화 변화의 비율 (없음과 제어 (CTL 대 포도당)를 첨가)하는 방법을 보여줍니다 및 미토콘드리아 상태 (기본 기능 대 약리학 변경된 기능)와 함께.
">
당분과 호흡 소스에서 그림 3. 양자의 생산 속도 (PPR). 호흡에서 PPR (오픈 부분) 및 추가 포도당 (세 왼쪽 막대) 또는 추가 포도당 (세 바로 바)없이 식 (5)을 사용하여 계산 C2C12 세포의 당분 (채워진 부분). 6의 데이터. 모든 데이터는 N의 ± SEM = 8 생물학적 복제를 의미 나타냅니다.
토론
세포 외 산성화는 세포 신진 대사 속도를 쉽게 측정 지표이다. (락트산 생산에 의해 정의 됨) 적절히 그것을 분석 배지의 완충력을 아는 것이 중요하다 셀룰러 해당 작용의 속도를 결정하고, 생산 속도를 프로톤 산소 소비 및 산성화의 세포 외 플럭스 측정 변환. 이 계산을 행함으로써, 시트르산 사이클에 출시 CO 2에서 얻어진 산성화 락 테이트 생산으로 인한 산성화 남겨 감산 될 수있다.
이 보정에 대한 완충력을 측정하기 위해 여기에 제공된 여러 가지 방법이 서로 다른 장단점을 가지고. 산도 프로브를 사용하여 외부 측정 고도로 정확하고 재현성이지만 분석 플레이트, 분석 중에 화합물 첨가 또는 t의 존재에 포함 형광체에 의해 도입 pH를 검출 작은 차이를 반영하지 않을그는 세포 그 자체. 인 - 판 pH 측정은 이러한 문제를 해결뿐만 아니라, 실험적인 소음의 변화도 소개합니다.
ECAR에 CO 2 보정은 처음 당분 레이트 모호 정량 계산이 가능하게하고, 실험의 과정 전체에 ECAR 호흡과 당분 기여도의 변동을 보여준다. 수학 식 5 및 OCR, ECAR의 측정치를 이용하여, 전력을 버퍼링, 당분 레이트 제공된 간단한 스프레드 시트 (표 6)를 사용하여 계산 될 수있다. 6을 원하는 경우,이 비율은 사후 락트산 측정에 의해 검증 될 수있다. 오탄당 포스페이트 경로는 고도로 활성 인 세포, 예컨대 6 aminonicotinamide 같은 경로 억제제의 사용은 당분 레이트를 분리하는 데 유용 할 수있다. 모두 CO 2의 기여의 계산 - 총과 젖산 파생 H +를 세포 외 산성화 속도와 산소 Consumpt 측정이온 비율은 신진 대사 활동에 대한 강력한 양적 진술을하는 세포 외 플럭스 데이터를 사용하기위한 매우 중요한 도구입니다.
버퍼링 전력을 측정하고, 조사 및 원하는 데이터에서 세포 외 플럭스 실험을 최적화하기위한 각종 변형을 포함하여 여기에 기재된 절차를 사용하여, 그대로 세포에서 해당 작용의 비율은 실험 조건의 넓은 범위에 따라 정량화 될 수있다. 이 방법은 폴리스티렌 표면 (에 부착 될 수 있거나 또는 세포 소기관)에 접착 배양에서 성장할 수있는 세포에 한정되어있다. 배양 된 세포는 균질하고 합류 할 때 유용한 데이터가 여전히 이러한 조건의 범위에서 얻어 질 수 있지만이, 가장 신뢰할 수있다. 계산은 K 세포가있는 경우에는 다른 기판과 부분 산화 6 이상 전체 산화 0.65-1.0 최대 H + / O 등의 2 범위, 세포의 신진 대사에 대한 약간의 지식이 필요합니다nown 포도당을 산화시키기 위해, 1.0의 값을 가정 할 수있다.
시스템에서 사용될 때 모든 대사 특성화와 관련이,이 방법이 특히 유용 할 수 있지만되는 호흡기 및 당분 해 대사 사이의 변화는 세포 ATP 공급 줄기 세포 및 종양 - 유도 된 암세포의 특성을 포함하여, 중요한 표현형이다 유지한다. 이들과 다른 상황에서 대사 제어의 변화를 이해하는 것은 이러한 세포 유형의 실험 설계 및 분석의 정교함과 정확성의 큰 학위를 허용합니다.
공개
박사 쇼나어 Mookerjee 그녀는 더 경쟁 금융 이익이 없다는 것을 선언합니다. 닥터 마틴 브랜드는이 문서에서 사용 기기 및 시약을 생산하는 해마 생명 과학에 대한 상담을하고있다. 이 기사에 대한 오픈 액세스 수수료 해마 생명 과학에 의해 지불했다.
감사의 말
We thank David A. Ferrick and David G. Nicholls for contributing to project conception and presentation, Renata L.S. Goncalves and Akos A. Gerencser for data not shown here and for helpful discussions, Barbara Liepe for XF24 consumables, and Andy Neilson for input in developing Eq. (5).
자료
| Name | Company | Catalog Number | Comments |
| Pherastar FS | BMG | n/a | microplate reader |
| Seahorse XF-24 | Seahorse Bioscience | n/a | extracellular flux instrument |
| Seahorse XF assay plate | Seahorse Bioscience | V7-PS | consumable |
| XF Calibrant | Seahorse Bioscience | 100840-000 | solution |
| HCl standard | Sigma | 38280 | chemical |
| oligomycin | Sigma | O4876 | chemical |
| FCCP | Sigma | C2920 | chemical |
| Rotenone | Sigma | R8875 | chemical |
| Myxothiazol | Sigma | T5580 | chemical |
| DMEM | Corning | 10-013-CV | medium component |
| FBS | Corning | 35-010-CV | medium component |
| penicillin/streptomycin | Corning | 30-002-CI | medium component |
| carbonic anhydrase | Sigma | C2624 | chemical |
| 96-well assay plate | Corning | CLS3991 | consumable |
| NAD+ | Sigma | N7004 | chemical |
| LDH | Sigma | L1254 | chemical |
참고문헌
- Brand, M. D., Nicholls, D. G. Assessing mitochondrial dysfunction in cells. Biochem. J. 435 (2), 297-312 (2011).
- Gerencser, A. A., et al. Quantitative microplate-based respirometry with correction for oxygen diffusion. Anal. Chem. 81 (16), 6868-6878 (2009).
- Divakaruni, A. S., Paradyse, A., Ferrick, D. A., Murphy, A. N., Jastroch, M. Analysis and interpretation of microplate-based oxygen consumption and pH data. Meth. Enzymol. 547, 309-354 (2014).
- Renner, K., Jansen-Dürr, P., Gnaiger, E. Biphasic oxygen kinetics of cellular respiration and linear oxygen dependence of antimycin A inhibited oxygen consumption. Mol. Biol. Rep. 29 (1-2), 83-87 (2002).
- Helmlinger, G., Sckell, A., Dellian, M., Forbes, N. S., Jain, R. K. Acid production in glycolysis-impaired tumors provides new insights into tumor metabolism. Clin. Cancer Res. 8 (4), 1284-1291 (2002).
- Mookerjee, S. A., Goncalves, R. L. S., Gerencser, A. G., Nicholls, D. G., Brand, M. D. The contributions of respiration and glycolysis to extracellular acid production. Biochim. Biophys. Acta. 1847, 171-181 (2015).
- Wu, M., et al. Multiparameter metabolic analysis reveals a close link between attenuated mitochondrial bioenergetic function and enhanced glycolysis dependency in human tumor cells. Am. J. Physiol. 292 (1), C125-C136 (2006).
- Nadanaciva, S., et al. Assessment of drug-induced mitochondrial dysfunction via altered cellular respiration and acidification measured in a 96-well platform. J. Bioenerg. Biomembr. 44 (4), 421-437 (2012).
- Pelletier, M., Billingham, L. K., Ramaswamy, M., Siegel, R. M. Extracellular flux analysis to monitor glycolytic rates and mitochondrial oxygen consumption. Meth. Enzymol. 542, 125-149 (2014).
- Nicholls, D. G., Darley-Usmar, V. M., Wu, M., Jensen, P. B., Rogers, G. W., Ferrick, D. A. Bioenergetic profile experiment using C2C12 myoblast cells. J. Vis. Exp. (46), 2511 (2010).
- Rogers, G. W., et al. High throughput microplate respiratory measurements using minimal quantities of isolated mitochondria. PLoS ONE. 6 (7), e21746 (2011).
- Seahorse Biosciences. . F24 Extracellular Flux Analyzer and Prep Station Installation and Operation Manual. 1.7, (2010).
- Blau, H., Chiu, C., Webster, C. Cytoplasmic activation of human nuclear genes in human heterocaryons. Cell. 32, 1171-1180 (1983).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유