
Method Article
Измерение и анализ внеклеточной кислоты производства, чтобы определить гликолитических Оценить
В этой статье
Резюме
Glycolysis is a defining metabolic marker in multiple biological systems. Monitoring glycolysis by measuring the extracellular flux of H+ is common, but requires correction to be quantitative and unambiguous. Here, we demonstrate how to gather and correct extracellular flux data to distinguish between respiratory and glycolytic sources of extracellular acidification.
Аннотация
Extracellular measurement of oxygen consumption and acid production is a simple and powerful way to monitor rates of respiration and glycolysis1. Both mitochondrial (respiration) and non-mitochondrial (other redox) reactions consume oxygen, but these reactions can be easily distinguished by chemical inhibition of mitochondrial respiration. However, while mitochondrial oxygen consumption is an unambiguous and direct measurement of respiration rate2, the same is not true for extracellular acid production and its relationship to glycolytic rate 3-6. Extracellular acid produced by cells is derived from both lactate, produced by anaerobic glycolysis, and CO2, produced in the citric acid cycle during respiration. For glycolysis, the conversion of glucose to lactate- + H+ and the export of products into the assay medium is the source of glycolytic acidification. For respiration, the export of CO2, hydration to H2CO3 and dissociation to HCO3- + H+ is the source of respiratory acidification. The proportions of glycolytic and respiratory acidification depend on the experimental conditions, including cell type and substrate(s) provided, and can range from nearly 100% glycolytic acidification to nearly 100% respiratory acidification 6. Here, we demonstrate the data collection and calculation methods needed to determine respiratory and glycolytic contributions to total extracellular acidification by whole cells in culture using C2C12 myoblast cells as a model.
Введение
Общая цель этого метода является точно измерить скорость гликолиза клеток с использованием анализа внеклеточной потока. Количественное измерение скорости гликолиза, используя внеклеточный подкисления является искомым конечная точка многих экспериментах. Тем не менее, общая скорость внеклеточного подкисления является суммой двух составляющих: дыхательная подкисление, в виде СО 2 (который увлажняет Н 2 СО 3, то распадается на НСО 3 - + Н +), и гликолитического подкисления, в виде лактата - + Н +.
Вклады CO 2 в общей внеклеточной подкисления до недавнего было считать незначительным в измерительной платформы, используемой здесь, в XF24 анализатора 7. Тем не менее, очевидно, в нескольких других систем, которые CO 2 может быть одной из основных причин внеклеточной подкисления 4-5. Несколько статей признать этот конвклад, но не пытайтесь прямой количественный СО 2 -derived кислоты 3,8,9. Недавно мы показали, что количественно СО 2 производство является значительным источником внеклеточной подкисления в этой системе 6. Кроме того, хотя существует множество метаболических путей, которые генерируют СО 2 из катаболизма глюкозы, тем осуществляется матричных дегидрогеназ в цикле лимонной кислоты подавляющие Авторы и все другие источники генерации количество CO 2, которые в пределах экспериментальной ошибки 6.
Без коррекции СО 2 производства внеклеточного подкисления, следовательно, является неоднозначным показателем гликолитического скорости и не могут быть использованы в количественном отношении. Нашей предыдущей публикации подчеркивается несколько случаев, когда дыхательная СО 2 содержит основную часть общего сигнала подкисления, даже в клетках, как правило, как полагают, в основном, используют гликолиз 6. Кроме того,дыхательная СО 2 вклад в общую подкисления широко варьируется в ходе общих метаболических экспериментах профилирования, демонстрируя, что правильно сравнение гликолитического скоростью в разные части эксперимента требует коррекции для CO 2.
Для измерения скорости гликолиза клеток с использованием скорости внеклеточной подкисления, необходимо преобразовать изменение рН на изменения в общем H +, генерируемых и вычесть внеклеточный подкисление вызванное CO 2 выделяется при работе в цикле лимонной кислоты. Здесь мы опишем простой метод для измерения внеклеточной дебит протонов (с внеклеточной изменения рН и калиброванного буферизации власти анализа среды) и СО 2 производства (от внеклеточного изменений в O 2 концентрации), и продемонстрировать, как рассчитать гликолитический скорость На основе этих измерений.
Эта сила метод ENS утилиту внеклеточного подкисления измерения, используя его правильно рассчитать скорость гликолиза, как определено лактата. Без коррекции дыхательных CO 2 (или прямого измерения лактата), невозможно определить, если и в какой степени общий уровень кислотности отражает гликолитический скорость, не оправдав интерпретацию экспериментов, использующих общую внеклеточный подкисления в качестве прямого измерения лактата.
РАСЧЕТ
СО 2 и лактата, в пределах экспериментальной ошибки, только два вклад в производство внеклеточной кислоты, основанные на экспериментах с клетками миобластов 6. Таким образом, скорость общего внеклеточного подкисления (PPR, дебита протон) может быть определена как:
PPR PPR малыш = соответственно + PPR Glyc Уравнение 1
. _content ">, где малыш = общее; соответственно = дыхания; Glyc = гликолитическая гликолитических PPR такова:PPR PPR Glyc = малыш - PPR соответственно Уравнение 2
Вот,
PPR малыш = РВЦА малыш / ВР Уравнение 3
где РВЦА = внеклеточного скорости подкисление (миль / ч / мин), и ВР = буферизации питания (миль / ч / пмоль Н + в 7 мкл), а
ППР = соответственно (10 рН-PK 1 / (1 + 10 рН-PK 1)) (макс Н + / О 2) (OCR TOT - OCR об / MYX) Уравнение 4
где К 1 = комбинированный константа равновесия СО 2 гидратации и диссоциации в НСО 3 - + Н +; макс Н + / O 2 = йе СО 2 -derived подкисление для конкретного метаболической трансформации, таких как полное окисление глюкозы 6; OCR = скорость потребления кислорода (O 2 пмоль / мин), и OCR об / MYX = без митохондриальной OCR.
Уравнение 4 изолирует митохондриальной OCR путем вычитания любого не-митохондриальной OCR (определяется как OCR, которая устойчива к митохондриальной дыхательной ядов ротенона и myxothiazol) и счетов для максимального H +, образованное на O 2 потребляется для каждого субстрата (макс Н + / O 2 ) (см 6), а также доля CO 2 порождая H + на экспериментальной температуры и рН (10 рН-PK 1 / (1 + 10 рН-PK 1). Для полного окисления глюкозы, митохондриальная кислорода Потребление Оценить (OCR) в точности равна скорости CO 2, производства. В ограниченном объеме анализа измерения внеклеточной потока, CO 2, продуктыд дыханием остается в ловушке в среду для анализа. Большая часть захваченных СО 2 увлажненной, чтобы H 2 CO 3, который затем распадается на НСО 3 - + Н +. Небольшая часть остается растворенным, но не гидратированного, а другой небольшая часть является гидратированный, но не диссоциируют, как продиктовано термодинамически комбинированным константа равновесия СО 2 гидратации и диссоциации, чтобы HCO 3 - + Н + при температуре эксперимента (37 ° C) и рН (~ 7,4).
Таким образом, полное уравнение для расчета PPR г путем вычитания PPR PPR соотв от TOT является:
PPR Glyc = РВЦА малыш / ВР - (10 рН-PK 1 / (1 + 10 рН-PK 1)) (макс Н + / O 2) (OCR малыш - OCR об / MYX) Уравнение 5
яп Таким образом, темпы дыхания и гликолиза, а также связанных с ними темпы производства АТФ, может быть количественно определена из простых измерений (потребления кислорода, внеклеточный подкисления, буферная емкость) и импорта или расчета других необходимых значений (Н + / вывода 2 , п / о, а константа равновесия K 1) 6. Эксперимент, описанный здесь расширяется на стандартных методов для использования внеклеточной поток анализатор, таких как конек XF24 10,11; для других форматов измерений внеклеточного поток (например, XF е 96, или XFP), все объемы ниже должны быть расширены соответствующим образом.
Буферный мощность в среду для анализа может быть измерена путем построения калибровочной кривой либо непосредственно во внеклеточном платформы потока или отдельно с помощью калиброванного рН зонда. Здесь три варианта для измерения буферизации внеклеточным потока среды для анализа даны, в том числе с использованием всех injectiна портах внеклеточного анализатора потока с скважин образцов клеточных бесплатно, либо с использованием только последней инъекции порт в ячейке, содержащей скважин (раздел 1), или с помощью внешнего измерения рН (раздел 2). См.прикрепленную таблицу для полных расчетов, например, данные.
Для измерения буферизации питание с помощью рН-обнаружения возможности внеклеточного инструмента потока, это самый безопасный в использовании бесклеточных скважин, чтобы минимизировать изменение сигнала. Тем не менее, в пределах погрешности, статистической разницы не существует между бесклеточной и клеточно-содержащий скважин при выполнении этого измерения (данные не показаны). Примечание: Изменение описано на стадии 1.7 осуществляет преимущество составляет любых потенциальных изменений в буферизации, предоставляемых добавленных соединений или в присутствии клеток, с недостатком шумной сигнала. Однако, как указано выше, никаких существенных различий не было обнаружено в расчетной мощности между буферным конструкции бесклеточной показанном в таблице 1, идизайн после эксперимента в таблице 2 в экспериментальных условиях, описанных здесь.
Кроме того, на малых диапазонах ΔpH (<0,4 единиц; экспериментально лучше ограниченные до 0,2 единиц), то линейный наклон получается путем построения Д миль / ч / мкмоль H + адекватно приблизительно логарифмическая зависимость между ΔpH и [H +]. Наклон стандартной кривой, следовательно, представляет собой буферную силу в среду для анализа тестируемого рН / нмоль H + в 7 мкл или миль / ч / пмоль Н + 7 мкл. Мы рекомендуем увеличение средней мощности буферизации или уменьшением плотности клеток в образцах, которые превышают единичное изменение рН 0,2 в течение времени измерения. Время измерения также может быть уменьшена, но это может привести к сокращению установившейся скорости подкисления и ввести ошибку в расчете скорости.
протокол
1. Измерение Буферизация Сила в внеклеточной Flux инструмент: два варианта
Примечание: расчеты и способы, описанные здесь, были разработаны с использованием внеклеточного Flux Analyzer. Для других инструментов, объем измерения должны быть расширены соответствующим образом.
- Подготовка 0,1 М HCl в стандартной воде с использованием концентрата HCl (см Материалы и оборудование) в соответствии с инструкциями производителя.
Примечание: Пример расчета для приготовления инъекций HCl для использования во всех четырех инжекционных отверстий показано в таблице 1:

Таблица 1. Последовательные инъекции HCl в внеклеточного анализа потока также.
- Подготовьте разведения стандарта HCl в среде должны быть анализировали как описано в таблице 1 для ряда технических повторов, являющихся CarrIED в одной тарелке, плюс один, чтобы позволить ошибки пипетирования; например, для четырех технических повторов порта впрыска, подготовить запас (1,1 мкл х 5) 0,1 М HCl в (48,9 мкл х 5) анализа среды.
- Распределить 50 мкл в каждую Порт А картриджа измерения зонда. Повторите эту процедуру для оставшихся портов B, C и D.
- Запуск внеклеточный анализа потока 10 со стандартным циклом калибровки, после чего два цикла [2 мин смесь в течение 1 минуты ждать, и 5 измерения мин] для каждого из четырех портов дополнений (рисунок 2).
- Программирование эксперимент выше программного обеспечения прибора в соответствии с инструкциями программного обеспечения. Загрузите подготовленную картридж в аппарат и выполнить калибровку в соответствии с инструкциями программного обеспечения.
- При появлении в программе, удалите калибрующего содержащие пластину и вставьте пластину, содержащую анализа среды в каждую лунку в прибор; продолжить программу.
- U петь среднем 8-10 точек, полученных в стационарном состоянии (как правило, в последние 8-10 баллов) до и после каждого порта Кроме того, рассчитать совокупный () разница в рН (ΔpH), вызванного каждой инъекции стандартного кислоты.
- Участок ΔpH против нмоль Н +, содержащиеся в объеме 7 мкл захваченных измерительного зонда. Линейный наклон буферного питания (БП) в миль / ч / пмоль Н +.
- В качестве альтернативы к шагам 1,2-1,3, провести измерения ΔpH следующие анализе, в котором порты A, B, и С, используемого для проведения эксперимента с последующим инъекции HCl в Порт D. Как в таблице 2, четыре параллельных образца технические используется для генерации каждую точку стандартной кривой 5-балльной в 20 экспериментальных скважин (за исключением четырех фон коррекции температуры скважин) внеклеточного потока анализа пластины.
53464table2.jpg "/>
Таблица 2. Одноместный HCl инъекции в внеклеточного анализа потока скважины.
2. Измерение Буферизация питания Использование внешнего измерителя рН
Примечание: Для измерения буферизации мощность среде, используя внешнюю рН зонд, калибровку зонда при 37 ° С и поддерживают эту температуру в течение всех реагентов в ходе эксперимента.
- Подготовка 0,1 М HCl стандартный в воде с использованием концентрата HCl в соответствии с инструкциями производителя.
- Теплый рН зонд, стандарты рН, анализ среды, буферизация мощность должна быть измерена, и 0,1 М HCl до 37 ° С на водяной бане.
- Калибровка рН электрод при 37 ° С в соответствии с инструкциями производителя. Поддержание все реагенты при 37 ° С в течение всего теста с помощью тепла пластину или водяной бане.
- Аликвоты 10 мл среды для анализа в небольшой стакан или коническую трубку. Монитор рН постоянно используя погруженную рН зонда.
- Добавить 0,1 М HCl в качествеговорят среду в 10-20 мкл аликвоты.
- Убедитесь, смешивая с помощью мешалкой или вручную вращая контейнер после каждого добавления кислоты.
- Подождите несколько секунд для измерения рН для стабилизации, а затем записать рН после каждого добавления.
- Как показано в таблице 3, чтобы достаточное количество дополнений, чтобы обеспечить точный расчет наклона и охватывают диапазон рН ожидаемое в ходе эксперимента.

Таблица 3. Измерение энергии с использованием буферизации рН-метр. Данные представляют собой типичного эксперимента с шестью 20 мкл добавлением 0,1 М HCl.
- Участок ΔpH против нмоль H + добавлены в 7 мкл, давая линейный наклон, который представляет власть буферизации (рисунок 1).

Рисунок 1. Определение буферизации власть. Стандартная кривая HCl измеряли в таблице 1, Таблица 2 или (здесь), а в таблице 3. Наклон линейной аппроксимации кривой дает силу буферизации (рН / нмоль H + в 7 мкл). Каждая точка представляет среднее ± SEM п = 9 технические повторов.
- После того, как мощность буферизации известно через метод 1 или 2, преобразовать РВЦА сигнал (миль / ч / мин), чтобы PPR (пмоль Н + / мин / мкг белка) путем деления Ecar буферизацией мощности (ВР) (миль / ч / пмоль Н +) и масштабирование, чтобы содержание белка в каждой лунке:
PPR малыш (пмоль Н + / мин / мкг белка) = РВЦА (миль / ч / мин) / BP (миль / ч / пмоль Н + в 7 мкл) / белка на лунку (мкг) Уравнение 6 - В качестве альтернативы, используйте те же самые эксперименты в методес 1 или 2 для расчета стоимости буферную емкость (BC), используемый программным обеспечением прибора для автоматического расчета РНП во время сбора данных.
ПРИМЕЧАНИЕ: Руководство пользователя инструмент 12 (стр 107) содержит подробную информацию о расчете и использовании буферную емкость, где до н.э. описывается как
БК (моль / л) = моль H + / (ΔpH объем буфера х (L)) Уравнение 7
ПРИМЕЧАНИЕ: буферная емкость, как определено в уравнении 7, может быть рассчитана в инструмент или внешний рН зонда анализов, описанных выше. Преобразование между буферным власти и буферной емкости легко сделать (прилагается таблица):
BC = 1 х 10 -9 / BP ((миль / ч / пмоль Н + в 7 мкл) / 7 мкл) Уравнение 8
ПРИМЕЧАНИЕ: Если известны до проведения анализа, буферная емкость может быть введен непосредственно в программное обеспечение прибора во время эксперимента. - Применение этой процедуры и расчеты, используемые выше большинства обычных буферных систем, как описано в предыдущей публикации 6.
ПРИМЕЧАНИЕ: В таблице 4 перечислены электрические буферизации и буферная емкость из нескольких традиционных средств массовой информации.

Таблица 4. Буферизация мощности и буферная емкость отдельных средств массовой информации.
3. Выполнение внеклеточного Flux анализе с использованием клеток миобластов C2C12
Примечание: На шаге 3.4.3, не было никаких различий в наблюдаемых CO 2 не -derived производство кислоты зависит от наличия карбоангидразы в С2С12 культуры, предполагая, что его присутствие не требуется для полного превращения CO 2 в НСО 3 - + Н +. Тем не менее, для эмпирической проверки этого в разных экспериментальных системах рекомендуется перед омitting карбоангидразы.
- Культура мыши C2C12 миобластов 13 при 37 ° С в атмосфере 95% воздуха / 5% CO 2 в модифицированной Дульбекко среде Игла (DMEM) с 11,1 мМ глюкозы, 2 мМ глутамина, 10% об / об эмбриональной бычьей сыворотки (FBS), 100 Ед / мл пенициллина и 100 мкг / мл стрептомицина.
- 24 ч до анализа, пластина / семян клетки в 100 мкл той же культуральной среде при 20000 клеток / лунку в 24-луночный полистирольный внеклеточного поток Аналитический планшет (см Материалы и методы) без дополнительной покрытием.
- Развести олигомицин, FCCP и ротенон плюс myxothiazol и HCl (опция) до 10x конечной концентрации в Кребса Рингера фосфат HEPES (KRPH) анализа среды (2 мМ HEPES, 136 мМ NaCl, 2 мМ NaH 2 PO 4, 3,7 мМ KCl, 1 мМ MgCl 2, 1,5 мМ CaCl 2, 0,1% вес / объем жирных кислот, свободных бычий сывороточный альбумин, рН 7,4 при 37 ° С).
- Подготовка сотовый
- 30 мин перед анализом, мыть адгезивных клеток три раза путем аспирации с Gently удалить среды от скважины, а затем медленно добавляя 500 мкл KRPH.
- Инкубируйте клетки после третьего шага стирки при 37 ° С в атмосфере воздуха (не менее 5% CO 2, который будет изменить рН этой бикарбоната среде без).
- При запуске анализа, заменить KRPH в скважинах с 500 мкл свежей KRPH содержащей 500 U / мл карбоангидразы и либо глюкозу (10 мм) или только среду, без дополнительной подложке.
- Загрузка картриджа датчика
- Пипеткой 50 мкл аликвоты каждого 10x соединения, полученного на стадии 3.3 в патронных портов внеклеточного потока картриджа датчика следующим образом (конечные концентрации в тесте также приведены): порт A: 2 мкг / мл олигомицину, Порт B: 0,5 мкм FCCP, Порт C : 1 мкМ ротенон, 1 мкМ myxothiazol, порт D: HCl (если калибровке в-анализа кислоты, как описано выше, и в таблице 2).
ПРИМЕЧАНИЕ: для назначения полной торможения дыхательной цепи, описанные здесь, 1 мкМ Моиxothiazol могут быть использованы взаимозаменяемо с 1 мкМ антимицином А.
- Пипеткой 50 мкл аликвоты каждого 10x соединения, полученного на стадии 3.3 в патронных портов внеклеточного потока картриджа датчика следующим образом (конечные концентрации в тесте также приведены): порт A: 2 мкг / мл олигомицину, Порт B: 0,5 мкм FCCP, Порт C : 1 мкМ ротенон, 1 мкМ myxothiazol, порт D: HCl (если калибровке в-анализа кислоты, как описано выше, и в таблице 2).
- Внеклеточной анализ потока:
- Выполните стандартную внеклеточный анализа потока для определения дыхательного контроля, как описано в 10.
Примечание: Для каждого сегмента эксперимента, определить смесь, подождать, и время измерения необходимо, а также количество циклов в сегменте.
ПРИМЕЧАНИЕ: Данные в таблице 5 были собраны в течение двух циклов анализа по 2 мин смеси, 1 мин ожидания, и 5 мин меры для каждого сегмента, с трех циклов анализа происходящих после Порт D добавлением различных количеств HCl (для калибровки буферизации Мощность, как в таблице 2).

Таблица 5. внеклеточной поток конфигурации анализа.
4. Measuriнг конечная точка концентрации лактата
Примечание: Для проверки косвенного анализа, описанного здесь, в какой-то другой системе, точка концентрации лактата конец к концу внеклеточного эксперимента потока могут быть определены непосредственно в обычном 96-луночного планшета путем измерения начальной скорости (в течение 2 мин) снижения НАД + НАДН → катализируемой лактатдегидрогеназы, подробно описанной в предшествующих публикации 6. Для данных, представленных в Представитель Результаты, концентрация лактата конечная точка в содержащей глюкозу аналитические лунки был ~ 40 мкм.
- Подготовьте 2x гидразина среды: 1 М Трис, 20 мМ ЭДТА, 400 мМ гидразин, рН 9,8 при 22 ° С). Непосредственно перед началом анализа, добавить NAD + до 4 мм и лактатдегидрогеназы (ЛДГ) до 40 Ед / мл. Окончательный анализ состава среды (1x): 500 мМ Трис, 10 мМ ЭДТА, 200 мМ гидразина, 2 мМ НАД +, 20 ед / мл ЛДГ.
- Сразу же после внеклеточный анализа потока, удалить 100мкл среды для анализа из каждой лунки внеклеточного потока анализа пластины и передачи в лунку непрозрачной (черный) 96-луночного планшета.
- Для каждого образца, добавьте 100 мкл 2x гидразина среду.
- Сразу загрузить пластину в микропланшетного и начать мониторинг флуоресценции NADH при возбуждении 340 нм / 460 нм излучения.
- Запишите начальную скорость примерно на 2 мин.
- Запустите подобный эксперимент, чтобы построить стандартную кривую начальную скорость от концентрации лактата для добавленных концентрации лактата от 0 до 50 мкм.
- Рассчитать концентрацию лактата в каждой экспериментальной скважины с использованием стандартной кривой.
5. Измерение содержания белка
- Удалить остатки среды для анализа из каждой лунки аналитического планшета.
- Вымойте лунки три раза 250 мкл BSA-Free KRPH, будьте осторожны, чтобы свести к минимуму оплывание клеток из нижней поверхности скважины.
- Добавить 25 мкл лизис Рипасреда (150 мМ NaCl, 50 мМ Трис, 1 мМ EGTA, 1 мМ ЭДТА, 1% об / об Тритон Х-100, 0,5% вес / об дезоксихолат натрия, 0,1% об / об SDS, рН 7,4 при 22 ° С) В каждую лунку планшета для анализа.
- Инкубируйте пластины на льду в течение 30 мин.
- Перемешивают пластину на шейкере при 1200 оборотах в минуту в течение 5 мин.
- Измерение концентрации белка стандартными методами, например, путем анализа BCA, гарантируя, что состав буфера для лизиса совместимы с методом измерения. Содержание белка в эксперименте на рисунке 2 составляла ~ 4 мкг / лунку.
Результаты
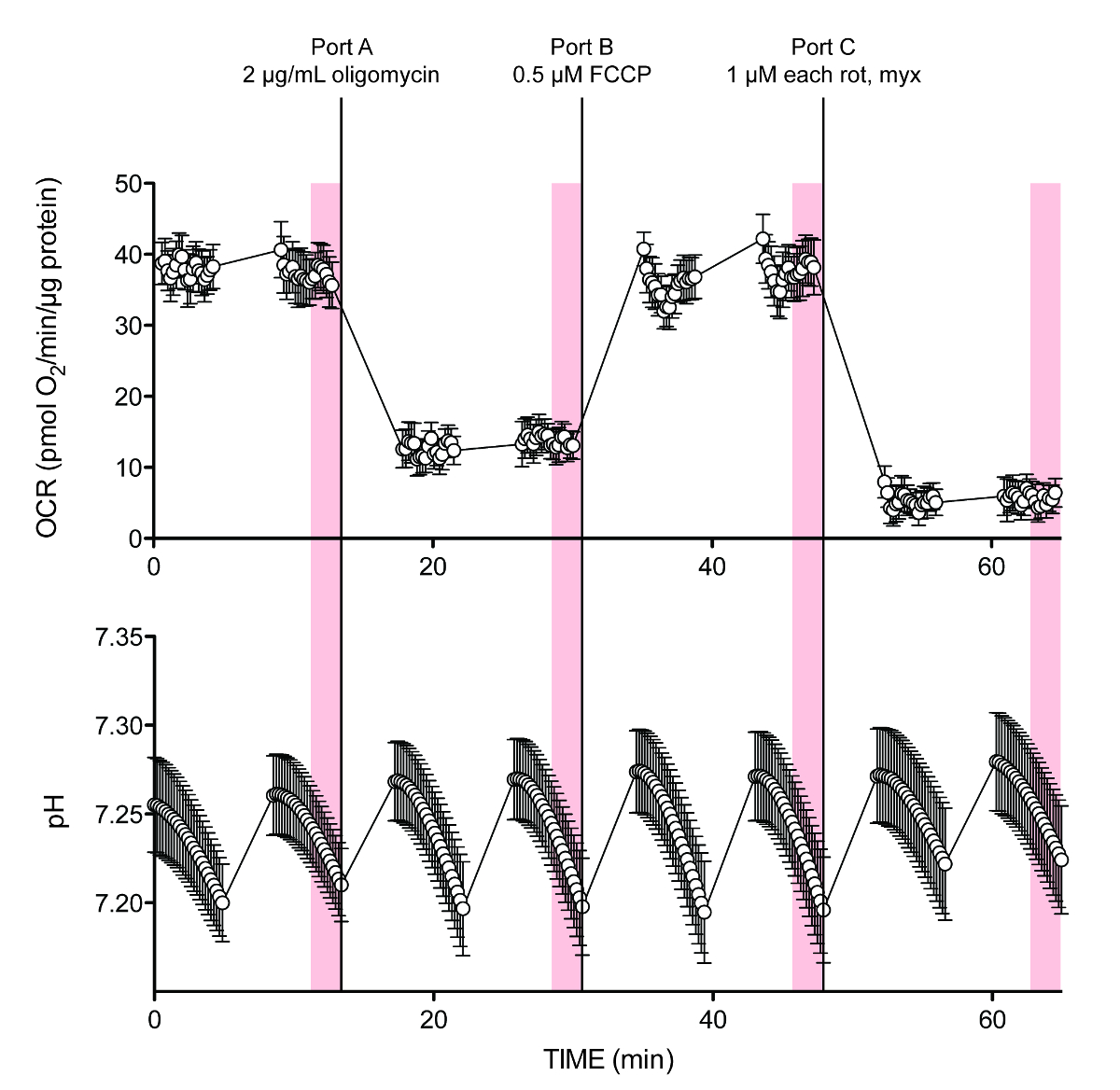
Рисунок 2 показывает исходные данные для типичного эксперимента. Последние 10 точек измерения от записи точка-точка, так и OCR (рН затененной вертикальные полосы) были использованы для расчетов. Первоначальные опасения, что среднее значение (измерение средняя точка) каждого цикла анализа не обеспечивают достаточную разрешение скорости для точного расчета, в частности, так как, казалось, было небольшое отставание между портом того и постоянной скоростью государственной подкисления, не оправдались, как это не появляется в значительной степени способствовать ошибки в расчетах (не показан). Кроме того, если правильно буферная емкость вводится во время экспериментальной установки, PPR можно прочитать непосредственно из инструмента сбора данных считывания, отображая выход PPR в программном обеспечении прибора или в формате PC-совместимый доступной в качестве одного из параметров выходных данных.
ftp_upload / 53464 / 53464fig2.jpg "/>
Рисунок 2. Типичные внеклеточные потока следы O 2 и H +. OCR и рН следы на примере эксперимента в таблице 5, содержащем 10 мМ глюкозы в начале анализа. Порт D имеют различные концентрации HCl для калибровки мощности буферизации (не показан на этих усредненных следов). Данные предыдущей публикации 6. Каждая точка представляет среднее ± SEM из N = 8 биологических повторяет. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
Анализ данных представительств результатов
Использование таблицы, показанной в таблице 6, который в качестве вложения, значения данных из отдельных скважин может быть введен в колонках с желтыми заголовками. Все шесть колонок в право вычисляются на основе этих записей. Пример в таблице 6 показаны расчеты PPR PPR соответственно и Glyc использованием Ecar и данные OCR из отдельных скважин для нативных условиях с добавлением или без добавления сахара, до порта A добавлением олигомицина. Технические повторов на каждую биопрепарата обычно в среднем, чтобы дать отдельные значения выходов в последних четырех столбцах, то данные из различных биологических препаратов в среднем с соответствующим распространения статистических данных об ошибках в ВР, и эти четыре значения.

Таблица 6. Расчет дыхательной и гликолитического подкисления. Колонны главе желтым указывают значения вводится из расчета (например, ВР, макс Н + / О 2), или от сбора данных (например, РВЦА малыш, OCR). PS: //www.jove.com/files/ftp_upload/53464/53464table6.jpg "целевых =" _blank "> Нажмите здесь, чтобы посмотреть большую версию этой таблице |. Пожалуйста, нажмите здесь, чтобы скачать этот стол в качестве таблицы Excel ,
Вклады гликолиза и дыхания в PPR после коррекции
Рисунок 3 показывает графический вывод данных, рассчитанных в таблице 6 в родных для ставок гликолиза и дыхания подкисления, ставки следующие олигомицин того (Порт A), и ставки следующие FCCP того (Порт B). Эти данные ясно показывают, как пропорции дыхательной и гликолитического изменения подкисления с выбором подложки (глюкозы по сравнению с контролем (CTL) ни с нами) и митохондриальной статуса (родной функции против фармакологически измененной функции).
">
Рисунок 3. Протон дебит (PPR) из гликолитических и дыхательной источников. PPR от дыхания (открытые участки) и гликолиза (заполненные участки) из С2С12 клеток, рассчитанных с использованием уравнения 5 с добавлением глюкозы (три левых бар) или без добавления сахара (три правых бар). Данные 6. Все данные представляют собой среднее ± SEM из N = 8 биологических повторяет.
Обсуждение
Внеклеточной подкисления является легко измерить указание сотовой метаболизма. Чтобы правильно определить скорость гликолиза сотовой (как определено лактата) важно знать буферизации силу в среду для анализа, и для преобразования внеклеточного измерений параметров потоков потребления кислорода и подкисления протона темпы производства. При выполнении этого расчета, подкисление в результате CO 2 выпущен в цикле лимонной кислоты могут быть вычтены, оставляя подкисление, которая является результатом лактата.
Несколько различных способов приведенные здесь, чтобы измерить силу буферизации для этой коррекции нести различные преимущества и недостатки. Внешний измерений с использованием рН электрод обладает высокой точным и воспроизводимым, но может не соответствовать небольшие различия в обнаружении рН введенной флуорофоров, содержащихся в планшет для анализа, добавлением соединений в процессе анализа, или в присутствии третон сами клетки. В-пластины измерения рН решать эти вопросы, но и ввести в разной степени экспериментального шума.
CO 2 поправка к РВЦА позволяет впервые однозначными и количественного расчета скорости гликолиза, и показывает изменение в дыхательной и гликолитического вклад в общую РВЦА в ходе эксперимента. Использование уравнения 5 и измерения OCR, РВЦА и буферизации власть, гликолитическая ставка может быть рассчитана с помощью простого таблицу представленную (таблица 6). При желании 6 Эта скорость может быть проверена путем измерения лактата после специальной. В клетках, где пентозофосфатный путь обладает высокой активностью, применение ингибиторов пути, такие как 6-aminonicotinamide может быть полезно, чтобы изолировать гликолитических скорость. Расчет взносов как CO 2 - и лактата, полученных Н + из общего измеряется внеклеточной подкислением Оценить и кислород Consumptионная Оценить является бесценным инструментом для использования внеклеточные данные потока сделать мощные и количественные заявления о метаболической активности.
Используя методики, описанные здесь, включая различные модификации для измерения буферизации мощности и оптимизации внеклеточный эксперимент потока для клеток исследуемых и данных желании скорость гликолиза в интактных клетках может быть определена количественно в широком диапазоне условий эксперимента. Этот метод ограничен клетками, которые могут расти в прикрепленной культуре на (или клетки или органеллы, которые могут быть придерживались) полистирольный поверхность. Это самый надежный при культивируемые клетки однородны и сливной, хотя полезные данные могут еще быть получены в диапазоне этих условиях. Расчеты требуют некоторые знания о метаболизме клеток, как максимум H + O 2 / колеблется от 0,65 до 1,0 для полного окисления различных субстратов и больше для частичного окисления 6, однако, если клетки кNown окислять глюкозу, значение 1.0 можно предположить.
Хотя отношение к метаболическим всех характеристик, этот способ может быть особенно полезным при использовании в системах, в которых сдвиг между дыхательной и гликолитического метаболизма, чтобы поддерживать питания сотовой АТФ является критическим фенотип, в том числе характеристики стволовых клеток и опухолевых полученных раковых клеток. Понимание метаболических нарушений контроля в этих и других контекстах позволит в большей степени сложности и точности в экспериментальной разработки и анализа этих типов клеток.
Раскрытие информации
Д-р Шона Мукерджи заявляет, что она не имеет конкурирующие финансовые интересы. Доктор Мартин Марка консультировал Морской конек биологических наук, который производит инструменты и реактивы, используемые в этой статье. Открытые сборы для доступа к этой статье были оплачены Морской конек биологических наук.
Благодарности
We thank David A. Ferrick and David G. Nicholls for contributing to project conception and presentation, Renata L.S. Goncalves and Akos A. Gerencser for data not shown here and for helpful discussions, Barbara Liepe for XF24 consumables, and Andy Neilson for input in developing Eq. (5).
Материалы
| Name | Company | Catalog Number | Comments |
| Pherastar FS | BMG | n/a | microplate reader |
| Seahorse XF-24 | Seahorse Bioscience | n/a | extracellular flux instrument |
| Seahorse XF assay plate | Seahorse Bioscience | V7-PS | consumable |
| XF Calibrant | Seahorse Bioscience | 100840-000 | solution |
| HCl standard | Sigma | 38280 | chemical |
| oligomycin | Sigma | O4876 | chemical |
| FCCP | Sigma | C2920 | chemical |
| Rotenone | Sigma | R8875 | chemical |
| Myxothiazol | Sigma | T5580 | chemical |
| DMEM | Corning | 10-013-CV | medium component |
| FBS | Corning | 35-010-CV | medium component |
| penicillin/streptomycin | Corning | 30-002-CI | medium component |
| carbonic anhydrase | Sigma | C2624 | chemical |
| 96-well assay plate | Corning | CLS3991 | consumable |
| NAD+ | Sigma | N7004 | chemical |
| LDH | Sigma | L1254 | chemical |
Ссылки
- Brand, M. D., Nicholls, D. G. Assessing mitochondrial dysfunction in cells. Biochem. J. 435 (2), 297-312 (2011).
- Gerencser, A. A., et al. Quantitative microplate-based respirometry with correction for oxygen diffusion. Anal. Chem. 81 (16), 6868-6878 (2009).
- Divakaruni, A. S., Paradyse, A., Ferrick, D. A., Murphy, A. N., Jastroch, M. Analysis and interpretation of microplate-based oxygen consumption and pH data. Meth. Enzymol. 547, 309-354 (2014).
- Renner, K., Jansen-Dürr, P., Gnaiger, E. Biphasic oxygen kinetics of cellular respiration and linear oxygen dependence of antimycin A inhibited oxygen consumption. Mol. Biol. Rep. 29 (1-2), 83-87 (2002).
- Helmlinger, G., Sckell, A., Dellian, M., Forbes, N. S., Jain, R. K. Acid production in glycolysis-impaired tumors provides new insights into tumor metabolism. Clin. Cancer Res. 8 (4), 1284-1291 (2002).
- Mookerjee, S. A., Goncalves, R. L. S., Gerencser, A. G., Nicholls, D. G., Brand, M. D. The contributions of respiration and glycolysis to extracellular acid production. Biochim. Biophys. Acta. 1847, 171-181 (2015).
- Wu, M., et al. Multiparameter metabolic analysis reveals a close link between attenuated mitochondrial bioenergetic function and enhanced glycolysis dependency in human tumor cells. Am. J. Physiol. 292 (1), C125-C136 (2006).
- Nadanaciva, S., et al. Assessment of drug-induced mitochondrial dysfunction via altered cellular respiration and acidification measured in a 96-well platform. J. Bioenerg. Biomembr. 44 (4), 421-437 (2012).
- Pelletier, M., Billingham, L. K., Ramaswamy, M., Siegel, R. M. Extracellular flux analysis to monitor glycolytic rates and mitochondrial oxygen consumption. Meth. Enzymol. 542, 125-149 (2014).
- Nicholls, D. G., Darley-Usmar, V. M., Wu, M., Jensen, P. B., Rogers, G. W., Ferrick, D. A. Bioenergetic profile experiment using C2C12 myoblast cells. J. Vis. Exp. (46), 2511 (2010).
- Rogers, G. W., et al. High throughput microplate respiratory measurements using minimal quantities of isolated mitochondria. PLoS ONE. 6 (7), e21746 (2011).
- Seahorse Biosciences. . F24 Extracellular Flux Analyzer and Prep Station Installation and Operation Manual. 1.7, (2010).
- Blau, H., Chiu, C., Webster, C. Cytoplasmic activation of human nuclear genes in human heterocaryons. Cell. 32, 1171-1180 (1983).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены