
Method Article
הליך RNA הנפתח כדי לזהות מטרות RNA לונג ללא קידוד RNA
In This Article
Summary
שיטה זו נפתחים RNA מאפשר זיהוי המטרות RNA של RNA לונג ללא קידוד (lncRNA). על סמך הכלאה תוצרת בית, מעוצב תחושה אנטי DNA oligonucleotide הגששים הספציפי הזה lncRNA רקמות כראוי קבוע או שורת התאים, ביעילות מאפשר לכידת כל המטרות RNA של lncRNA.
Abstract
זמן ללא קידוד RNA (lncRNA), אשר מהווים רצף נוקלאוטידים יותר מ-200 ללא מסגרת הקריאה מוגדר, שייך המשפחה של הרגולציה ללא קידוד RNA. למרות תפקודים ביולוגיים שלהם נשארים אינו מודע לקיומם, המספר של אלה lncRNAs עלה בהתמדה, עכשיו מעריכים כי בני אדם יכול להיות יותר מ-10,000 תעתיקים כזה. חלק מהם ידועים להיות מעורב חשוב מסלולים הרגולציה של ביטוי גנים אשר מתקיימים ברמת תעתיק, אלא גם על שלבים שונים של RNA co - ועם בגרות post-transcriptional. במקרים האחרונים, RNAs המיועדים מאת lncRNA חייב להיות מזוהה. זו הסיבה למה זה שימושי לפתח שיטה המאפשרת הזיהוי של RNAs הקשורים ישירות או בעקיפין עם lncRNA של עניין.
פרוטוקול זה, אשר היה בהשראת הפרוטוקולים שפורסמו בעבר ומאפשר את ניתוקה של lncRNA יחד עם רצפי כרומטין המשויכת שלה, הותאם להתיר את הבידוד של RNAs המשויך. אנחנו נקבע כי שני צעדים הם קריטיים עבור היעילות של פרוטוקול זה. הראשון הוא העיצוב של ספציפי תחושה אנטי DNA oligonucleotide הגששים מסוגלים hybridize כדי lncRNA עניין. למטרה זו, המבנה משני lncRNA היה שמנבאת ביואינפורמטיקה, תחושה אנטי oligonucleotide הגששים תוכננו עם משיכה חזקה עבור אזורים המציגים הסתברות נמוכה של זיווג בסיס פנימי. צעד קריטי השני של ההליך מסתמך על התנאים לשבועיים של רקמות או תאים בתרבית שיש לשמר את הרשת בין כל השותפים מולקולרית. יחד עם רצפי RNA תפוקה גבוהה, פרוטוקול זה נפתחים RNA יכול לספק את interactome RNA שלם של lncRNA עניין.
Introduction
המטרה הכוללת של השיטה המתוארת כאן היא לזהות השותפים מולקולרית RNA RNA noncoding זמן (lncRNA). LncRNA תואמות רצפי נוקלאוטידים יותר מ-200 ללא מסגרת הקריאה מוגדר. חלק מהם הוכחו להיות מעורב ביטוי הכונה, לא רק ברמה תעתיק, אלא גם על שלבים שונים של RNA co - ועם בגרות post-transcriptional. במקרים האחרונים, מולקולרית של lncRNA השותפים RNAs כי הם זוהו. שיטה המאפשרת הזיהוי של RNAs הקשורים ישירות או בעקיפין עם lncRNA של עניין ואז יהיה חיוני לפתח.
שפורסמו קודם לכן בשיטות, כגון כרומטין בידוד על ידי RNA טיהור (בטוח)1,2 ו לכידת הכלאה ניתוח של RNA מטרות (תרשים)3,4, לאפשר גילוי תפוקה גבוהה RNA מכורך חלבונים, אתרי קישור גנומית של lncRNA ספציפי. ב שתי שיטות אלו, lncRNA עניין היה hybridized קודם כדי oligonucleotides משלים biotinylated, המתחם היה אז מבודדים באמצעות streptavidin חרוזים. ההבדל העיקרי בין שתי שיטות אלה קשורה העיצוב של הגששים כי המטרה lncRNAs. . אה, בהשראת דגים RNA, כללה תכנון בריכה של הגששים oligonucleotide DNA משלימים קצר כדי לפרוש לכל אורכה של lncRNA האסטרטגיה. לעומת זאת, בתרשים, המחברים הותאם assay מיפוי של RNase H ב- lncRNAs כדי לחקור אתרים לרשות הכלאה.
ההליך המוצע כאן עיצוב oligonucleotide biotinylated חוש אנטי DNA שימושים הגששים ביואינפורמטיקה מידול של מבנה שניוני lncRNA5 לבחירת רגשים עם משיכה חזקה עבור אזורים המציגים הסתברות נמוכה של פנימי בסיס זיווג. הליך זה יש את היתרון של להיות זול יותר מאשר אלה המבוססים על בריכות של ריצוף oligonucleotide הגששים2 וצורכת פחות זמן מאשר אלה בהתבסס על רגישות RNAse-H4.
מאחר גוף גדל והולך של ראיות הכונה post-transcriptional lncRNAs6, זה מאוד שימושי לפתח גישה המאפשרת לכידתו של RNAs כי הם היעדים lncRNA. יתר על כן, ניתנת לשימוש עבור מרבית היישומים, הגישה היה מותאם הן תאים בתרבית רקמה תמציות.
Protocol
כל ההליכים בוצעו בהתאמה קפדנית עם הקהילה הכלכלית האירופית עבור טיפול והשתמש של חיות מעבדה (86/609/EEC ו- 2010/63/מורה), תחת רישיון כמובן מאליו ל ד Becquet (Préfecture des בוש דו רון, אישור מס 13-002).
1. בדיקה עיצוב
-
באמצעות רצף lncRNA עניין ראשי, ליצור את מבנה שניוני על ה "שרת אינטרנט RNAstructure"5.
הערה: האתר, אלגוריתמים שונים יכול לשמש. כלי חיזוי שלושה לתת את התוצאות הטובות ביותר עבור בדיקה העיצובים הם: "קיפול" (מבנה אנרגיה חופשית הנמוך ביותר), "MaxExpect" (זוגות בסיס ובסבירות גבוהה) ו "Probnot" (כולל pseudoknots זוגות סביר בסיס). אלה שלושה ניתוחים ניתן יהיה לבצע, בהשוואה. שרתי אינטרנט אחרים, כגון the websuite וינה RNA7, יכול גם לשמש.- בחר אזורים להציג הסתברות נמוכה של זיווג בסיס פנימי ועיצוב תחושה אנטי oligonucleotide הגששים הבסיסים 25 עם משיכה חזקה עבור אזורים אלה.
הערה: התוכן GC של רגשים אלו צריך כמורכבת בין 40 ל- 60%. באמצעות כלי חיפוש יישור (פיצוץ), ודא כי הגששים oligonucleotide תחושה אנטי שנבחרו אינם מזהים נוקלאוטיד רצפי RNA אחרים הביעו במערכת התא שבחרת.
- בחר אזורים להציג הסתברות נמוכה של זיווג בסיס פנימי ועיצוב תחושה אנטי oligonucleotide הגששים הבסיסים 25 עם משיכה חזקה עבור אזורים אלה.
- עיצוב גם שאינם ספציפיים DNA oligonucleotide בדיקה של בסיסים 25 אשר מציג גם זיקה עבור lncRNA את עניין ולא עבור רצפי RNA אחרים בגנום של עניין.
- להזמין את הגששים עם ביוטין בקצה 3'.
הערה: כדי לצמצם את מכשול הסטריים, המרחק בין oligonucleotide לבין ביוטין צריך להיות מוגברת עם כרווח triethyleneglycerol. לקבלת תוצאה מיטבית, כדי שניתן יהיה להעריך את ייחודה של תוצאות נפתחים, מומלץ לבצע עיצוב 3 שונים רגשים אנטי-תחושה oligonucleotide ולאחר מכן להשוות השפעול היעילות שלהם.
2. cross-linking
-
Cross-linking בתרבית תאים
- תרבות GH4C1 sommatolactotroph תאים יותרת המוח במדיום F10 של חזיר, בתוספת 15% הסוס סרום ו- 2% עגל עוברית סרום. לגדל את התאים עד למפגש ב- 78.5 ס מ2 תרבות צלחות. זה תואם לתאים7 עונה 1 פרק 10.
- להסיר את המדיום תרבות תא מצלחת confluent GH4C1 78.5 ס מ2 תרבות, לאחר מכן יש לשטוף עם 1 x נפח בינוני של פוספט buffered תמיסת מלח (PBS)
- לתקן את התאים עם פתרון paraformaldehyde 1% ב- PBS (10 מ"ל מנות2 ס מ 78.5); הפתרון צריך להיות מוכן טרי פתרון מניות של paraformaldehyde 4%. קשר צולב תחת עצבנות 10 דקות בטמפרטורת החדר (RT).
התראה: Paraformaldehyde (PFA) הוא רעיל, יש לטפל בזהירות. - להרוות את הפעולה paraformaldehyde על-ידי הוספת 1/10 נפח של גליצין 1.25 מ' (1 מ"ל לכל 10 מ"ל של פתרון paraformaldehyde); להתסיס 5 דקות ב- RT.
- למחוק את התקשורת על ידי השאיפה ולשטוף שתי פעמים (5 דקות) עם 1 X נפח בינוני של PBS.
- להוסיף נפח של PBS התואם ל- 1/10th של אמצעי האחסון של מדיה, לאסוף תאים עם תא במגרדת ולאחר מכן העברת שפופרת צנטרפוגה.
- ספין-510 גרם ב 4 מעלות צלזיוס למשך 5 דקות.
- הסר את תגובת שיקוע רב ככל האפשר.
- לאחסן כדורי ללא הגבלת זמן כנדרש ב-80 מעלות צלזיוס.
-
Cross-linking רקמות
- לשים 5 מ"ג של רקמת בלוטת יותרת המוח העכבר טרי בריכוז של 1% paraformaldehyde מדולל ב- PBS (כ 10 x נפח הרקמה), להתסיס 10 דקות ב- RT.
- להרוות את הפעולה paraformaldehyde על-ידי הוספת פתרון גליצין 1.25 מ' (1 מ"ל לכל 10 מ"ל של פתרון paraformaldehyde), להתסיס 5 דקות ב- RT.
- למחוק את התקשורת על ידי השאיפה ולשטוף פעמיים עם PBS (כ 10 x נפח הרקמה). הסר את תגובת שיקוע רב ככל האפשר.
- לאחסן ברקמות צולבים ללא הגבלת זמן-80 מעלות צלזיוס.
3. תאים או רקמות פירוק
- הכנת המאגר פירוק (50 מ מ טריס-HCl pH 7.0, 10 מ מ EDTA, 1% מרחביות בתוספת 200 U/mL של פתרון מעכב RNAse ולאחר קוקטייל של פרוטאזות מעכב 5 µL/mL).
- כדי להשיג דגימות lysed ללא מפשיר מוקדמת, מחדש להשעות את כדורי תאים או רקמות צולבים עם המאגר (כ 1 מ"ל לכל 100 מ ג של גלולה תאים או רקמות). גלולה התא המתקבל עונה 1 פרק 107 תאים להצמיח מדגם lysed המכיל בערך 20 מ ג חלבון.
הערה: בהתאם הרקמה בשימוש, צעד של שיבוש מכני צריך להתווסף. במקרה זה, חשוב להימנע חימום של הדגימות במהלך שלב נוסף זה.
4. sonication
-
אופטימיזציה של התנאים sonication
- לתכנת את sonicator עם 2-5 סדרת 30 s ON ו- 30 s חופש.
- לבצע בדיקות כדי למטב את התנאים sonication על דגימות lysed מדולל (דילול גורם ½ או ¼ המקביל כ 10 או 5 מ ג חלבון). המקום מדולל דגימות lysed באמבט מים 4 ° C ולהתחיל הסדרה sonication.
- לטהר RNAs עם ערכת טיהור RNA או עם ריאגנט בידוד של RNA (למשל, Trizol).
- לטעון את הטוטאליות של RNA מטוהרת על 1% agarose בג'ל במאגר TBE, לבדוק את אורך קטעי RNA. ככה צריך נעות בין 200 ל-800 bp.
הערה: בהתאם לגודל קטע RNA, היעילות של הגששים אנטי-תחושה oligonucleotide יכול להשתנות. אז מומלץ לבדוק את היעילות של הגששים בתנאים שונים של sonication.
-
Sonication של הדגימות lysed
- דוגמאות lysed את המקום המתאים 20 מ ג של חלבונים (שהושג לאחר שלב 3.2) ב- 4 ° C רשמים ולהתחיל הסדרה sonication כפי אופטימיזציה בשלב 4.1.
- מיד לאחר sonication, צנטריפוגה למשך 5 דקות ב g 12,000-4 מעלות צלזיוס. העברת supernatants צינורות צנטריפוגה חדש.
הערה: כדי להבטיח הומוגניות, שכפל supernatants יכול להיות איחדו, וליידע בשלב זה.
5. RNA נפתחים
-
יום 1 – שלב הכלאה
- להוסיף 2 כרכים של הכלאה מאגר (50 מ מ טריס-HCl pH 7.0, 750 מ"מ NaCl, 1 מ"מ EDTA, מרחביות 1%, 15% Formamide הוסיף extemporaneously) כדי supernatants שנאספו לאחר השלב sonication. מערבולת.
- העברת µL 20 של כל מדגם בשפופרת צנטרפוגה (דגימות קלט) ולאחסן ב-20 ° C.
- להוסיף 100 pmol של הגששים oligonucleotide biotinylated (ספציפי או שאינם ספציפיים; ראה טבלה 1) כל דגימה. דגירה 4 עד 6 h תחת עצבנות מתון על מסובב שפופרת-RT.
- הוסף µL 50 של חרוזים streptavidin מגנטי בתוספת 200 U/mL של פתרון מעכב RNAse וקוקטייל של פרוטאזות מעכב 5 µL/mL....
- דגירה בין לילה תחת עצבנות מתון על מסובב צינור-RT.
-
יום 2 – שלב בידוד ה-RNA
- השתמש תמיכה מגנטי כדי להפריד בין החרוזים מתא lysate למחוק את תגובת שיקוע, לשטוף את החרוזים עם 900 µL שטיפת מאגר (מרחביות 0.5%, האס 2 x). חזור 5 פעמים בשריקות 5 דקות עצבנות-מסובב-RT.
- לאחר השטיפה האחרונה decant בפעם האחרונה ולהוסיף µL 95 מאגר Proteinease K (10 מ מ טריס-HCl pH 7.0, 100 מ מ NaCl, 1 מ"מ EDTA, 0.5% מרחביות) ו- 5 µL של proteinase-K (20 מ"ג/מ"ל) את הדגימות.
- על הקרח להפשיר את דגימות קלט (20 μL) ולהוסיף µL 75 מאגר Proteinease K 5 µL של proteinase-K (20 מ"ג/מ"ל).
- דגירה כל הדגימות עם proteinase K למשך 45 דקות ב- 50 מעלות ואז 10 דקות ב 95 ° C.
- מקררים את הדגימה על קרח למשך 3 דקות לפני להפריד את החרוזים בין RNAs עם תמיכה מגנטי. לשמור את תגובת שיקוע וזורקים את החרוזים.
- לטהר RNAs עם ערכת טיהור RNA, שתכלול את צעד עיכול הדנ א. חנות RNAs ב-80 מעלות צלזיוס.
- לבצע qPCR שעתוק במהופך (RT-qPCR) באמצעות ערכת RT ואחריו qPCR באמצעות תחל ספציפי (טבלה 1).
- לבנות שתי ספריות DNA המתאימים לבריכות RNA שני שהושג עם כל אחד הגששים Neat1 ספציפי (טבלה 1). לבצע רצף על מערכת רצף הדור הבא.
תוצאות
מספר מחקרים שנעשו לאחרונה הראו כי lncRNAs ממלאות תפקיד חיוני כמעט בכל תהליך ביולוגי חשוב, כי תפקיד זה מושגת באמצעות השליטה של ביטוי גנים המתרחשים הן את תעתיק והן את רמות post-transcriptional מראה במקרה זה האחרון כי RNAs עשוי להיות המטרה של lncRNAs6.
LncRNA גרעיני מועשר בשפע בתמליל 1 (Neat1) הוא מעורב neuropathologies שונה ומובהקת, נוירודגנרטיביות או אפילפסיה-8,-9,-10, הוא גם misregulated סוגי סרטן שונים11,12.
LncRNA זה ידוע גם להיות המרכיב המבני של גופים גרעיני ספציפי, paraspeckles, וכדי להיות מעורב בוויסות היממה post-transcriptional של ביטוי גנים13. Paraspeckles מצויים בגרעין כל תא, נוצרות סביב לא רק Neat1, אשר יש צורך והמבנה שלהם, אלא גם סביב חלבונים (RBP) של איגוד ה-RNA מספר14, אכן ידועים להיות מסוגל לשמור על מטרות RNA בתוך גרעין15 . היווצרות של paraspeckles מושגת דרך האגודה של הרכיבים השונים. צורה זו הוצגה להצגת תבנית קצבית היממה נהיגה של השמירה גרעיני קצבית של RNA מטרות13. השמירה הגרעין של RNA מטרות על-ידי paraspeckles עלולה להתרחש באמצעות קשירה RBP או ישירות דרך האגודה RNA/RNA, אך היקף RNAs ממוקד על ידי paraspeckles היה צריך להיקבע. כדי לזהות את הרנ א ממוקד ישירות או בעקיפין על-ידי Neat1,. פרוטוקול נפתחים RNA תוכנן המאפשרת הבידוד וזיהוי של כל המטרות Neat1 RNA בתאים בתרבית באותה מידה כמו דגימות רקמה (ראו איור 1 גרפי מצגת של הטכניקה).
הפרוטוקול הוחל גם בהצלחה על הזיהוי של RNA היעדים lncRNA אחר גרורות הקשורים ריאות אדנוקרצינומה התעתיק 1 (Malat1). Malat1 הוא מאוד שנשמרת, ביטוי lncRNA נמצאו ספאקלס הגרעין יחד עם מספר RNA שחבור גורמים. Malat1 ידוע להיות מעורב ברגולציה של שחבור של nascent pre-mRNA מספר16,17.
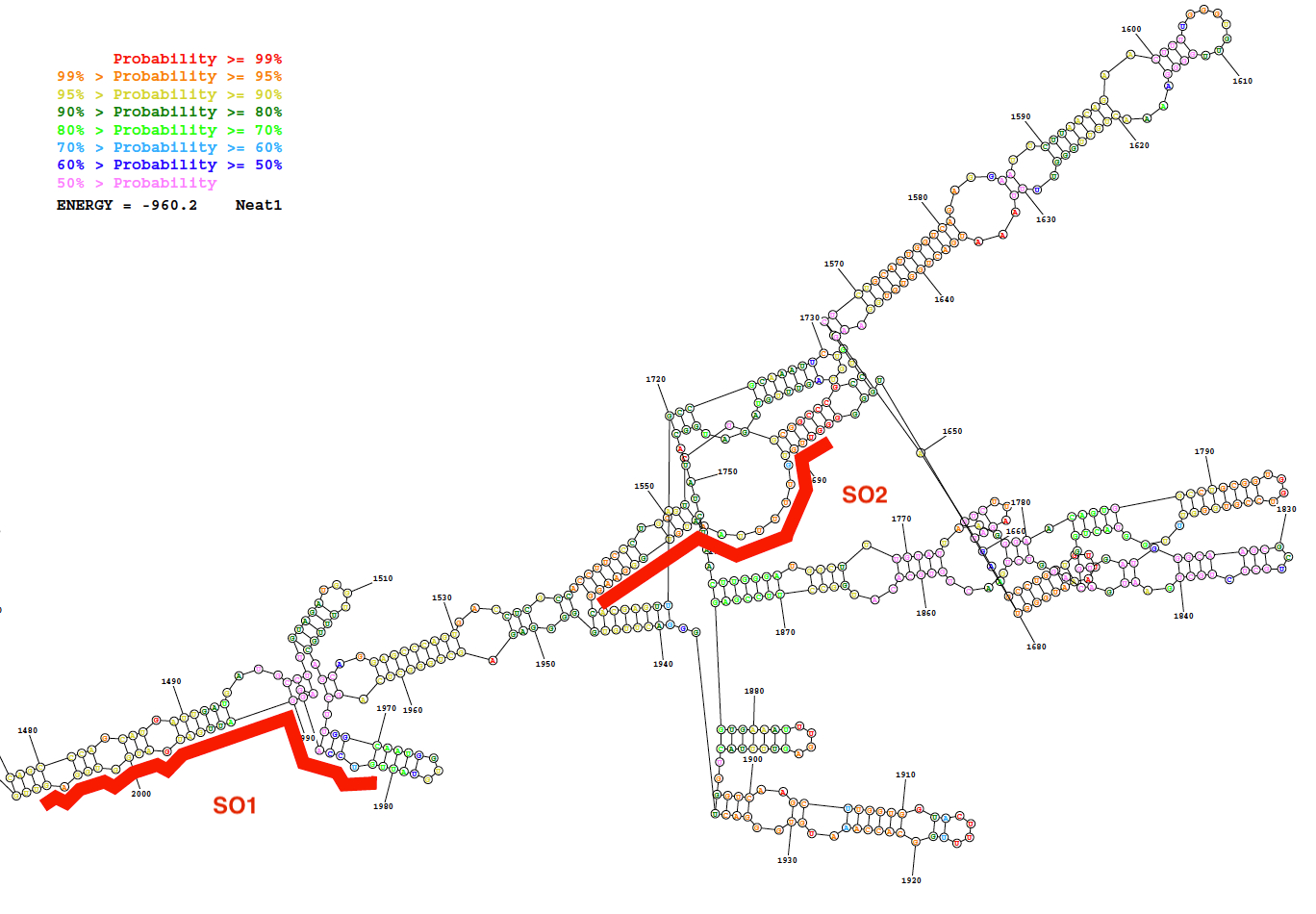
ספציפי (אז) וזונדים שאינם ספציפיים oligonucleotide (NSO) נוצרו באמצעות בדיקה עיצוב האסטרטגיה המתואר כאן. אסטרטגיה זו מסתמכת על הבחירה של אזורים המציגים הסתברות נמוכה של בסיס פנימי הזיווג כפי שחזיתי מאת המבנה משני של lncRNA ואת על העיצוב של הגששים ספציפי עם משיכה חזקה עבור אזורים אלה. כתוצאה מכך נציג של תחזיות אלה ביואינפורמטיקה, תמונה של המבנה משני החזוי של רצף של Neat1 (נוקלאוטידים 1,480 עד 2,000) יחד עם המיקום של שני תוכנן כך הגששים נותנים באיור2.
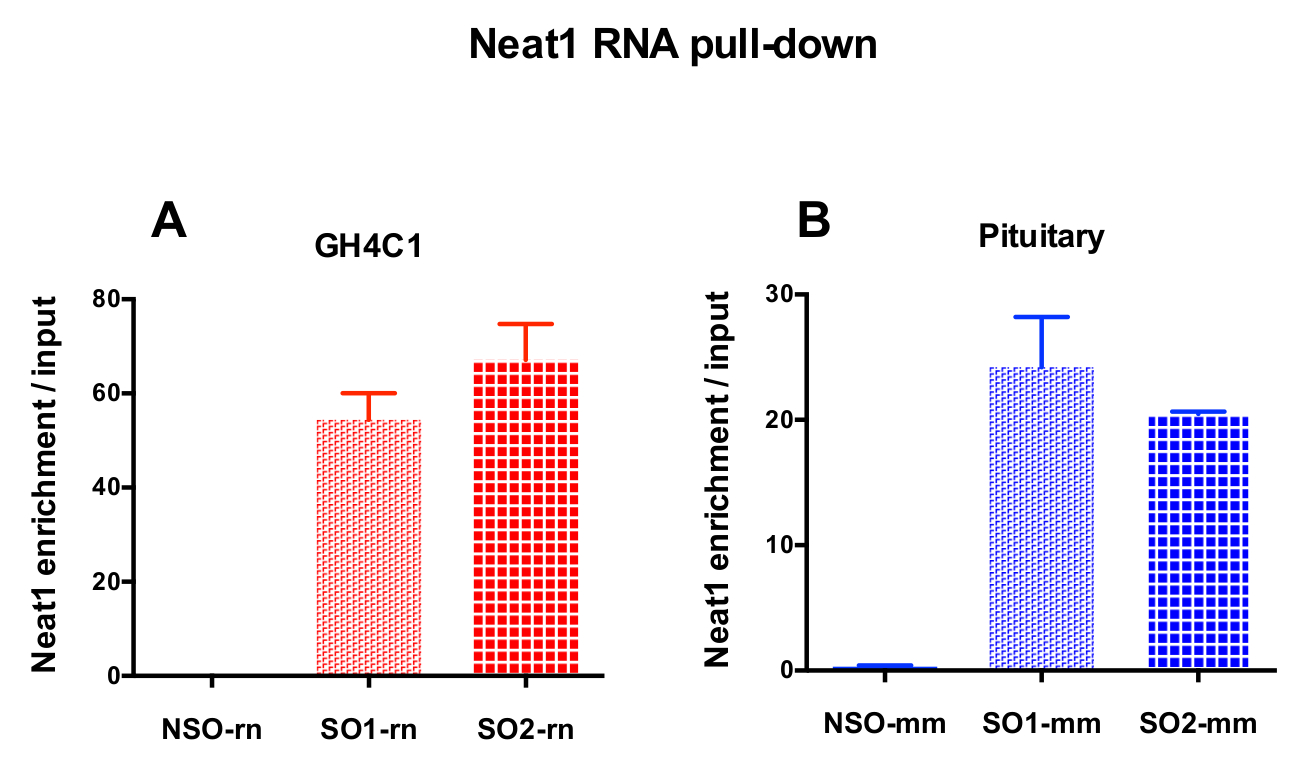
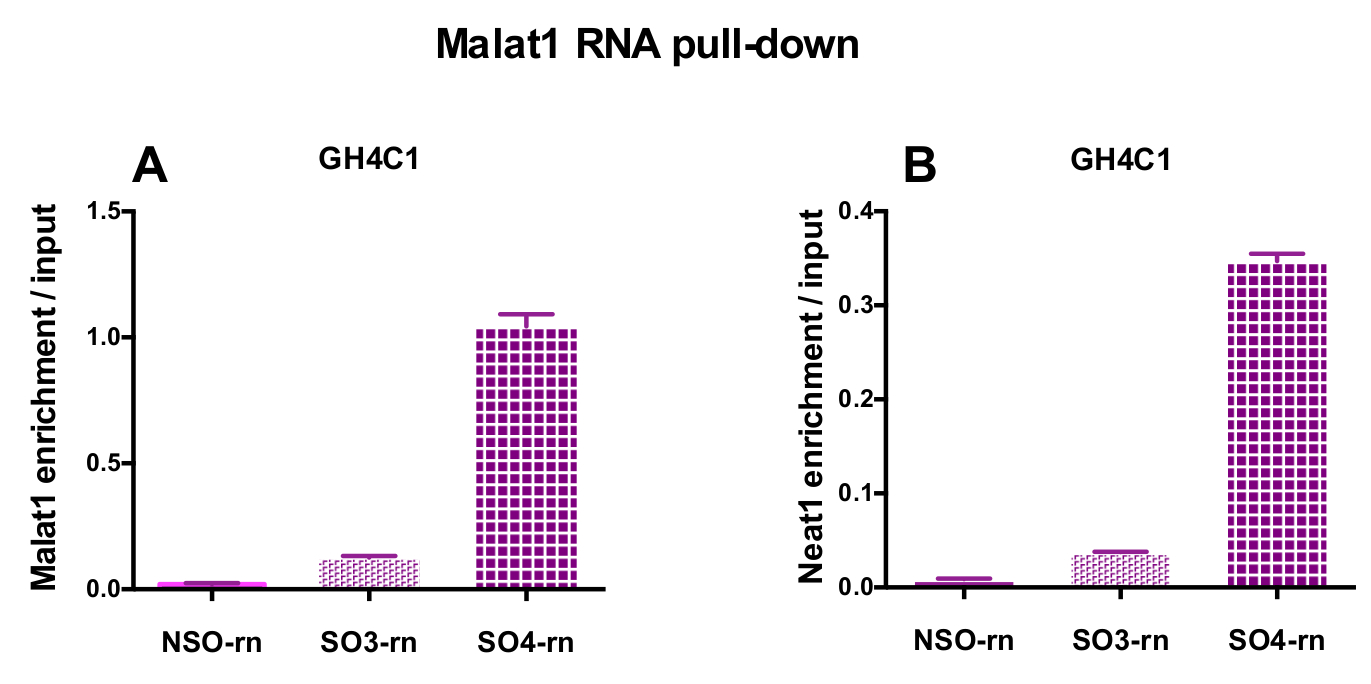
הגששים מעוצב הופנו להלשין Neat1 או Malat1 עבור תאים GH4C1 תרבותי וכדי בעכבר Neat1 על רקמת יותרת המוח תמציות (טבלה 1). העשרת יחסי Neat1 או Malat1 מחושב עבור הגששים שאינם ספציפיים וספציפיות ביחס בין דגימות קלט. איור 3 מראה את היעילות של הגששים ספציפיים כדי Neat1 נפתחים בחולדה שורת התאים יותרת המוח GH4C1 (איור 3 א) ותמצית עכבר רקמת יותרת המוח (איור 3B). בעת שימוש בפרוטוקול עיצוב בדיקה כדי ליצור oligonucleotide ספציפיים (אז) הגששים למזהים ייחודיים Malat1, אחד המכשיר היעיל הושג בזמן אחר היה לא מספיק יעיל, שנמחקו (איור 4A).
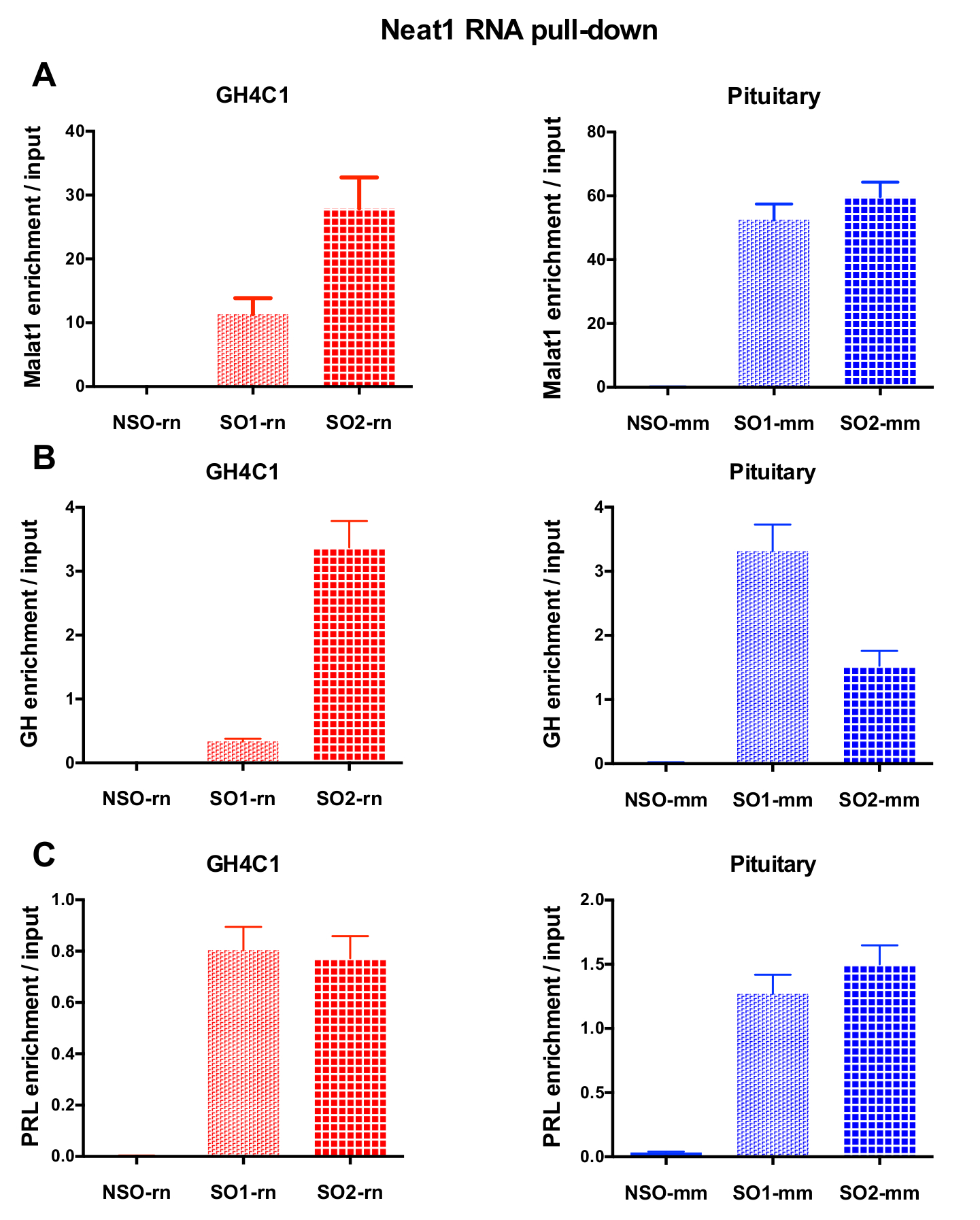
לאחר RNA הליך נפתחים ואחריו RT-qPCR הניסויים, כמה RNAs העריך עם תחל ספציפי (טבלה 1) הוצגו להיות מזוהה עם Neat1 או Malat1 ב GH4C1 תמציות. RNAs המשויך Neat1 ב GH4C1 תא תמציות הוצגו גם להיות מזוהה עם Neat1 של רקמת יותרת המוח תמציות. ואכן, לאחר Neat1 RNA נפתחים, Malat1 נמצאה ניתן לפלח לפי Neat1 של הקו תא GH4C1 והן העכבר יותרת המוח רקמות תמציות (איור 5A). קישור הדדי, Neat1 היה משמעותי מועשר לאחר Malat1 RNA נפתחים הופיעה עם מכשיר בדיקה ספציפית בתאי GH4C1 (איור 4B). על-ידי הדגשת הקשר ההדוק בין lncRNAs שני, תוצאות אלו עקביים עם התפקיד co-regulatory הפוטנציאלי של Neat1 ו- Malat1 שהוצעה על ידי עכברים knockout Malat1 המציגות וריאציות ב- Neat1 RNA ביטוי18, 19. ההעתקים של שני הורמוני יותרת המוח המרכזי, הורמון גדילה (Gh) (איור 5B) ואת פרולקטין (Prl) (איור 5C) היו באופן משמעותי מועשר בעקבות Neat1 RNA נפתחים הגששים ספציפי GH4C1 תאים והן יותרת המוח תמציות, רומז תקנה אפשרי של שני הורמונים על ידי Neat1. בעת השוואת שני רגשים ספציפיים השתמשו, נראה כי היעילות שלהם עשויים להשתנות בהתאם RNA המטרה נחשבת (איור 5B , איור 5C). תוצאות אלה להדגיש את הצורך בעיצוב מספר הגששים ספציפיים על מנת לבחור את אלה להצגת לא רק את היעילות הכי טוב העשרת lncRNA נפתחים, אלא גם את היעילות ביותר העשרת של מטרותיו RNA.
השיטה נפתחים RNA יכולה להיות מלווה גם רצפי RNA תפוקה גבוהה כדי לקבל את רשימת מטרות RNA lncRNA של עניין13מקיף. ניתוח ה-RNA-seq בתאי GH4C1 יותרת המוח לאחר Neat1 RNA הנפתח באמצעות שני רגשים ספציפיים המתוארים לעיל בוצעה. יצוין כי פקד שליליים באמצעות NSO גם אפשר יהיה נתון RNA-seq ניתוח, אם רמת ה-RNA התאושש לאחר ה-RNA נפתחים עם NSO הוא מספיק כדי לאפשר הקמת ספריות. זה לא היה המקרה הניסיון הקודם13. ספריות נוצרו לאחר שימוש של הגששים ספציפי נותחו באמצעות Tophat/חפתים צינור20 וגיליון הציונים רק עם ערכים של קטעים kilobase לכל מיליון הקריאות ממופה (FPKM) גבוהה יותר מאשר 1 נלקחו בחשבון. הרשימות שהושג עם שני רגשים ספציפי למזהים ייחודיים Neat1 (טבלה 1) היו שלובות להעריך יחודיות של התוצאות. גנים 4,268 נמצאו המשויך paraspeckles, אשר ייצג 28% של הפרוטוקולים ביטוי תאים GH4C113. בקנה אחד עם התוצאות המתקבלות באמצעות ניתוח qPCR (איור 5A-C), התמלילים של Gh, Prl Malat1 נמצאו להיות מזוהה עם Neat1. השיטה נפתחים RNA ולכן הוכח להיות כלי יעיל על מנת לחקור את האינטראקציה בין lncRNAs לבין מטרות RNA שלהם.

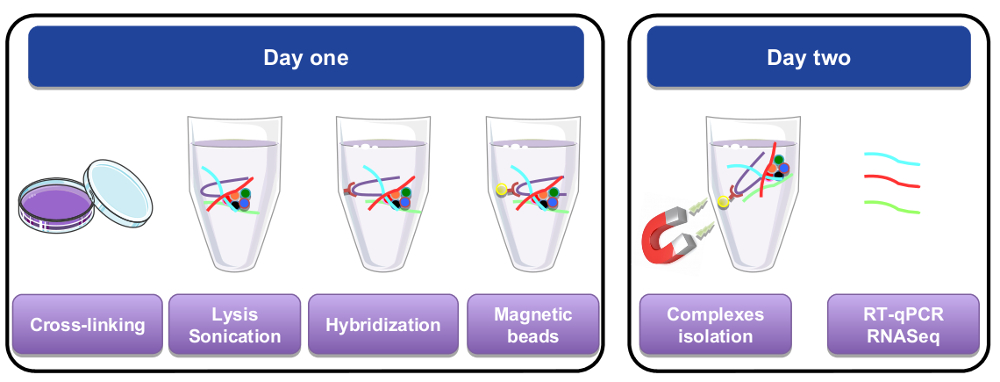
איור 1: ייצוג גרפי של RNA למשוך את ההליך. ביום הראשון, תאים או רקמות היו קישורים צולבים עם paraformaldehyde, lysed, sonicated לפני השלב הכלאה שבוצעה על-ידי הוספת הגששים ספציפית biotinylated. Streptavidin מגנטי חרוזים נוספו ואז להפרדת החומר הספציפי משאר חלקי התא lysate. ביום השני, חרוזים היו מבודדים על ידי מגנט ושטף מספר פעמים. צעד דה-crosslinking מותר שחזור של RNAs היו מטוהרים, המשמש RT-qPCR או ניתוח RNA-seq. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 2: מבנה שניוני של רצף Neat1 (נוקלאוטיד 1,480 עד 2,000) כפי שחזיתי על-ידי משאב לביואינפורמטיקה (לשרת RNAstructure; מבנה אנרגיה חופשית הנמוך). המבנה הוא בצבע על פי מידת ההסתברות של זיווג הבסיס. שני רגשים oligonucleotides (SO1 ו- SO2) באדום ממוקמות לאורך מבנה הרנ א Neat1. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 3: אימות qPCR Neat1 העשרה לעומת קלט. qPCR אימות של העשרה Neat1 לעומת קלט לאחר Neat1 RNA למשוך למטה על-ידי שני שונים ספציפיים רגשים (SO1-rn, SO2-rn עבור תאים GH4C1 ו- SO1-מ מ ו- SO2-מ מ על רקמת יותרת המוח) לעומת הלא-ספציפי אחד (NSO-rn עבור תאים GH4C1 ו- NSO-מ מ עבור רקמת יותרת המוח) בתאים עכברוש GH4C1 (א), יותרת המוח העכבר רקמות תמציות (B). התוצאות הן זאת אומרת ± SEM שהושגו בניסויים 3 עד 10. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 4: אימות qPCR העשרה Malat1 ו- Neat1 לעומת קלט, לאחר Malat1 RNA לנתץ. qPCR אימות של Malat1 (א) ו- Neat1 העשרה (B) לעומת קלט לאחר Malat1 RNA למשוך למטה על-ידי שני שונים ספציפיים רגשים (SO3-rn ו SO4-rn) לעומת הלא-ספציפי אחד (NSO-rn) בתאים עכברוש GH4C1. התוצאות הן זאת אומרת ± SEM שהושגו בניסויים 3. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 5: qPCR אימות של Malat1, Gh, Prl העשרה לעומת קלט לאחר Neat1 RNA לנתץ. qPCR אימות של Malat1 (א), Gh (B), העשרה Prl (C) לעומת קלט לאחר Neat1 RNA לנתץ באמצעות הגששים ספציפיים שונים לעומת אדם שאינם ספציפיים בתאים עכברוש GH4C1 ותמצית רקמת יותרת המוח העכבר. התוצאות הן זאת אומרת ± SEM שהושגו בניסויים 3-8. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}
| בדיקת שמות | רצפים | |
| NSO-Rn | TAAAATACCATTTGATGTTTGAAATTAT | |
| SO1-Rn | CTCCACCATCATCAATCCTCTGGAC | |
| SO2-Rn | GCCTTCCCACATTTAAAAACACAAC | |
| SO3-Rn | AACTCGTGGCTCAAGTGAGGTGACA | |
| SO4-Rn | AAGACTCTCAGGCTCCTGCTCATTC | |
| NSO מ מ | GTTTGTGGTTTAACAGTGGGAAGGC | |
| SO1 מ מ | GCCTTCCCACTGTTAAACCACAAAC | |
| SO2-מ מ | CTCACCCGCACCCCGACTCCTTCAA | |
| qPCR תחל: | ||
| Rattus norvegicus | ||
| Neat1 | AAGGCACGAGTTAGCCGCAAAT | |
| TGTGCACAGTCAGACCTGTCATTC | ||
| Malat1 | GAAGGCGTGTACTGCTATGCTGTT | |
| TCTCCTGAGGTGACTGTGAACCAA | ||
| Gh1 | CCGCGTCTATGAGAAACTGAAGGA | |
| GGTTTGCTTGAGGATCTGCCCAAT | ||
| Prl | TGAACCTGATCCTCAGTTTGGT | |
| AGCTGCTTGTTTTGTTCCTCAA | ||
| ס. מאסכאלאס | ||
| Neat1 | TGGGCCCTGGGTCATCTTACTAGATA | |
| CACAGCTGTTCCAATGAGCGATCT | ||
| Gh1 | CTCGGACCGTGTCTATGAGAAACTGA | |
| TTTGCTTGAGGATCTGCCCAACAC | ||
| Prl | TGAACCTGATCCTCAGTTTGGT | |
| AGCTGCTTGTTTTGTTCCTCAA | ||
טבלה 1: רצפי DNA oligonucleotide המחקרים וכל qPCR primers
Discussion
זמן noncoding RNAs (lncRNAs) על ידי מספר והמגוון שלהם לייצג תחום רחב של המחקר, רוב התפקידים שלהם הם עדיין להתגלות. רבים lncRNAs אלה יש לוקליזציה של הגרעין, ביניהם, כמה הם ערבו במשעולים הרגולציה של ביטוי גנים באמצעות מנגנונים תעתיק או posttranscriptional. אחד האתגרים הנוכחיים בתחום זה יש להבין את הרלוונטיות של האלה lncRNAs בעיבוד post-transcriptional של RNAs. למטרה זו, RNAs ממוקד על ידי lncRNAs צריך להיות מזוהה. בהשראת מחקרים קודמים התמקדו התאחדות lncRNAs עם כרומטין, פיתחנו תהליך המאפשר זיהוי RNAs המשויך lncRNA. ההצלחה של פרוטוקול זה, בשם RNA נפתחים, תלויה בעיקר שני צעדים חיוניים, כלומר העיצוב של חוש אנטי DNA oligonucleotide רגשים זה צריך במיוחד. ובלעדית hybridize עם lncRNA את עניין, התנאים של רקמה או קיבוע תא שיש כדי לשמור על השלמות של הרשת בין כל השותפים מולקולרית.
בעבר פרסם פרוטוקולים סיפק הליכים כדי לבודד את lncRNA יחד עם רצפי כרומטין המשויכת שלו (אה1,2,3,תרשים4). בפרוטוקולים אלה, הועסקו אסטרטגיות שונות לתכנון ש-oligonucleotide biotinylated חוש אנטי DNA רגשים. בהליך מעוצבנת, המחברים משמש מאגר של ה-DNA-biotinylated-oligonucleotide-הגששים המקיף לכל אורכה של lncRNA עניין לאחר ניכוי של הגששים שאינם ספציפיים של כל1,2. בפרוטוקול תרשים, המחברים זיהו את מחוזות lncRNA נגישים יותר של הכלאה ועוצב oligonucleotides לכידת המכוון על אזורים אלה. אזורים אלה נבחרו על סמך את רגישותם RNase-H. ואכן, באמצעות המאפיין של RNAse-H כדי hydrolyze RNAs באתרים של איגוד ה-RNA-DNA, oligonucleotides זה hybridize לאתרים נגישים ב lncRNA לייצר מכוניות היברידיות RNA-DNA ולהוביל lncRNA. המחשוף אנזימטי. המחברים נבחר שלושה אלה oligonucleotides לכידה של המועמד, המשמשים אותם קוקטייל3,4.
ההליך נהגנו עיצוב חוש אנטי DNA biotinylated oligonucleotide הגששים היתה קרובה בשימוש בפרוטוקול תרשים, אבל מחוזות lncRNA הרצוי הכלאה-זמין לא נבחרו על סמך את רגישותם RNAse-H, אבל לפי הסתברות נמוכה שלהם זיווג בסיס פנימי כפי שנקבע על ידי ביואינפורמטיקה מידול של מבנה שניוני lncRNA. זה בטח הבחין כי מבנים משניים שונים ניתן לחזות באמצעות אלגוריתמים שונים, הגששים להיבחר צריך להיות כאלה hybridize כדי רצפים זמינים של lncRNA בהמספר הגדול ביותר של מבנים משני חזה. אותן התוצאות התקבלו באמצעות קוקטייל של שלושה תוכנן, הגששים ספציפי או בדיקה אחת בנפרד. הנחיה זו השימוש של שני רגשים נפרד, הספציפיים, השיקול של תוצאות חיוביות כמו אלה משותפים אלה שני רגשים. לבסוף, ולכן מומלץ בתחילת הפיתוח של השיטה, לקבלת תוצאה מיטבית, כדי שניתן יהיה להעריך את ייחודה של התוצאות של המשיכה-למטה, לעצב 3 oligonucleotide אנטי-תחושה שונה רגשים ולאחר מכן להשוות השפעול היעילות שלהם, במיוחד מאחר יעילות המכשיר עשויה להשתנות על ידי תא lysate הכנה. למרות זאת, ההליך של עיצוב המכשיר המבוסס על ביואינפורמטיקה מידול של lncRNA משני מבנה השתמשנו נותרה פחות יקר זה מבוסס על בריכות של ריצוף oligonucleotide הגששים2, וזה היה פחות לארוך זמן רב יותר השיטה מבוסס על רגישות RNAse-H4.
פקד שלילי גם להתבצע באמצעות לכידת שלילי oligonucleotide גם חוש דנ א biotinylated oligonucleotide רגשים או מקושקשות הגששים oligonucleotide, או oligonucleotides נגד של RNA לא קשורים. בגלל הקיום של תעתיקים antisense טבעי lncRNAs, לפעמים ייתכן שימוש הגיוני oligonucleotide הגששים לקוי. ללא קשר החללית oligonucleotide שנבחר עבור הפקד שלילי, זה הכרחי כדי לבדוק אם הפיצוץ כי זה לא hybridize עם RNA ידועים וכדי לזכור כי oligonucleotide הזה יכול hybridize כדי lncRNA עדיין האו ם המבואר.
Lysates תא שימוש בניסויים משיכה אלה RNA, התקבלו 106 תאים7 10 בעת עבודה עם תאים בתרבית, מ 1 עד 10 מ ג כאשר עובדים עם רקמה. הכנת תא lysates צריך להיות מותאם על פי סוג הרקמה או תא בשימוש בשני שלבים העיקרי צריך להיות מותאם: כלומר, השלב cross-linking המאפשרת היווצרות קשרים הדדיים בין lncRNA את שותפיה מולקולרית, ו sonication שלב זה מפחית את צמיגות by גיזום כרומטין.
מטרת השלב cross-linking נועד להבטיח כי כל המטרות RNA להישאר סגור בפני lncRNA על ידי גרימת היווצרות של רשת בין כל השותפים מולקולרית. שלב הטיפול paraformaldehyde שיהוו קשרים הדדיים בין lncRNA את השותפים שלה מאפשר שהרשת תהיה רשת לסופרמרקטים. בפרוטוקול תרשים, הוצע אם על כל עובד עם lncRNA גרעינית, לבצע טיפול הראשון עם paraformaldehyde תא lysate, שנייה טיפול על שבר nucleic מבודד3,4. הבחנו כי צעד משלים זה ירד את היעילות של הגששים, כנראה על-ידי הפחתת הנגישות lncRNA בתאים. לכן, מידת ' רשתית ' מאת paraformaldehyde חייב להיות מותאם לוקח בחשבון את התא או סוג הרקמה בשימוש, הלוקליזציה של lncRNA את עניין, ואת היעילות של הגששים מעוצב.
בזמן lysing את התאים, כרומטין מופץ ב- lysate ומגביר צמיגות שלה; זה ואז יש צורך לגרוס את כרומטין מאת sonication להגדיל את הנזילות של הדגימות, ומכאן לקדם את הנגישות של oligonucleotide רגשים כדי lncRNA עניין. עם זאת, sonication גם לגרוס את RNAs חילוץ עם lncRNA עניין. אז חשוב למזער את הזמן sonication כך בזמן זה מפחית ביעילות את צמיגות lysate, שהיא גם מאפשרת שהקניית RNA קטעים באורך מורכבת בין 200-800 bp. שימו לב הפעם sonication תהיה מאוד תלוי הכמות והן את הסוג של רקמה או תאים בתרבית בשימוש.
לסיכום, ההליך המתואר כאן מאפשרת ב- 2-3 ימים לכידתו של RNA המטרות של lncRNA הרצוי. יחד עם RT-qPCR, שיטות אלה יאפשר מחפש של ויסות mRNA וארגון ספציפי על ידי lncRNA הרצויה כמו הגישה של המועמד. לגישה הגנום כולו, ניתן לנתח RNA נפתחים ניסויים על ידי RNA בתפוקה גבוהה-המאפשר שליפת כל RNAs המשויך lncRNA הרצוי רצף. לא משנה מה, אנליטי האסטרטגיה שנבחרה, ההליך נפתחים RNA צריך לספק ידע משמעותי חדש על RNA הרגולציה על-ידי lncRNAs.
Disclosures
המחברים אין לחשוף.
Acknowledgements
עבודה זו נתמכת על-ידי אוניברסיטת Aix-מרסיי ו CNRS, שמומן על ידי מענק של פייזר מעבדות.
Materials
| Name | Company | Catalog Number | Comments |
| Bioruptor Plus | Diagenode | B01020001 | Sonicator |
| Dynabeads My One | Thermo-Fisher | 65001 | Magnetic streptavidin beads |
| Formamide | Thermo-Fisher | 15515-026 | |
| Gel electrophoresis apparatus | Advance | Mupid-One | Gel electrophoresis apparatus |
| Proteinase K | Sigma | P2308 | |
| RNA XS purification kit | Macherey-Nagel | 740902 | RNA purificationkit |
| RNAseOUT | Thermo-Fisher | 10777-019 | RNAse inhibitor |
| Trizol | Thermo-Fisher | 15596018 | RNA purification |
| Tube Rotator | Stuart | SB2 | Eppendorf tube rotator |
| RNA to DNA | Thermo-Fisher | 4387405 | Reverse transcription kit |
| iTaq Universal SYBR Green Supermix | BioRad | 1725124 | qPCR reagent |
| Applied 7500 Fast | Thermo-Fisher | 4351107 | qPCR apparatus |
| Illumina TruSeq Stranded mRNA Sample Preparation kit | Illumina | 20020594 | DNA library construction kit |
| Illumina NextSeq 500 | Illumina | SY-415-1002 | NGS system |
References
- Chu, C., Qu, K., Zhong, F. L., Artandi, S. E., Chang, H. Y. Genomic maps of long noncoding RNA occupancy reveal principles of RNA-chromatin interactions. Mol Cell. , 667-678 (2011).
- Chu, C., Quinn, J., Chang, H. Y. Chromatin isolation by RNA purification (ChIRP). J Vis Exp. , e3912 (2012).
- Simon, M. D., Wang, C. I., Kharchenko, P. V., West, J. A., Chapman, B. A., Alekseyenko, A. A., Borowsky, M. L., Kuroda, M. I., Kingston, R. E. The genomic binding sites of a noncoding RNA. Proc Natl Acad Sci U S A. , 20497-20502 (2011).
- Simon, M. D. Capture hybridization analysis of RNA targets (CHART). Curr Protoc Mol Biol. , (2013).
- Reuter, J. S., Mathews, D. H. RNAstructure: software for RNA secondary structure prediction and analysis. BMC Bioinformatics. 11, 129 (2010).
- Sun, X., Haider Ali, M. S. S., Moran, M. The role of interactions of long non-coding RNAs and heterogeneous nuclear ribonucleoproteins in regulating cellular functions. Biochem J. , 2925-2935 (2017).
- Gruber, A. R., Lorenz, R., Bernhart, S. H., Neuböck, R., Hofacker, I. L. The Vienna RNA websuite. Nucleic Acids Res. , W70-W74 (2008).
- Tollervey, J. R., Curk, T., Rogelj, B., Briese, M., Cereda, M., Kayikci, M., König, J., Hortobágyi, T., Nishimura, A. L., Zupunski, V., Patani, R., Chandran, S., Rot, G., Zupan, B., Shaw, C. E., Ule, J. Characterizing the RNA targets and position-dependent splicing regulation by TDP-43. Nat Neurosci. , 452-458 (2011).
- Riva, P., Ratti, A., Venturin, M. The long non-coding RNAs in neurodegenerative diseases: novel mechanisms of pathogenesis. Curr Alzheimer Res. (27338628), (2016).
- Barry, G., Briggs, J. A., Hwang, D. W., Nayler, S. P., Fortuna, P. R., Jonkhout, N., Dachet, F., Maag, J. L., Mestdagh, P., Singh, E. M., Avesson, L., Kaczorowski, D. C., Ozturk, E., Jones, N. C., Vetter, I., Arriola-Martinez, L., Hu, J., Franco, G. R., Warn, V. M., Gong, A., Dinger, M. E., Rigo, F., Lipovich, L., Morris, M. J., O'Brien, T. J., Lee, D. S., Loeb, J. A., Blackshaw, S., Mattick, J. S., Wolvetang, E. J. The long non-coding RNA NEAT1 is responsive to neuronal activity and is associated with hyperexcitability states. Sci Rep. , 40127 (2017).
- Adriaens, C., Standaert, L., Barra, J., Latil, M., Verfaillie, A., Kalev, P., Boeckx, B., Wijnhoven, P. W., Radaelli, E., Vermi, W., Leucci, E., Lapouge, G., Beck, B., van den Oord, J., Nakagawa, S., Hirose, T., Sablina, A. A., Lambrechts, D., Aerts, S., Blanpain, C., Marine, J. C. p53 induces formation of NEAT1 lncRNA-containing paraspeckles that modulate replication stress response and chemosensitivity. Nat Med. , (2016).
- Fang, J., Qiao, F., Tu, J., Xu, J., Ding, F., Liu, Y., Akuo, B. A., Hu, J., Shao, S. High expression of long non-coding RNA NEAT1 indicates poor prognosis of human cancer. Oncotarget. , (2017).
- Torres, M., Becquet, D., Blanchard, M. P., Guillen, S., Boyer, B., Moreno, M., Franc, J. L., François-Bellan, A. M. Circadian RNA expression elicited by 3'-UTR IRAlu-paraspeckle associated elements. Elife. , (2016).
- Chen, L. L., DeCerbo, J. N., Carmichael, G. G. Alu element-mediated gene silencing. EMBO J. , 1694-1705 (2008).
- Tripathi, V., Ellis, J. D., Shen, Z., Song, D. Y., Pan, Q., Watt, A. T., Freier, S. M., Bennett, C. F., Sharma, A., Bubulya, P. A., Blencowe, B. J., Prasanth, S. G., Prasanth, K. V. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Mol Cell. , 925-938 (2010).
- Engreitz, J. M., Sirokman, K., McDonel, P., Shishkin, A. A., Surka, C., Russell, P., Grossman, S. R., Chow, A. Y., Guttman, M., Lander, E. S. RNA-RNA Interactions Enable Specific Targeting of Noncoding RNAs to Nascent Pre-mRNAs and Chromatin Sites. Cell. , 188-199 (2014).
- Nakagawa, S., Ip, J. Y., Shioi, G., Tripathi, V., Zong, X., Hirose, T., Prasanth, K. V. Malat1 is not an essential component of nuclear speckles in mice. RNA. , (2012).
- Zhang, B., Arun, G., Mao, Y. S., Lazar, Z., Hung, G., Bhattacharjee, G., Xiao, X., Booth, C. J., Wu, J., Zhang, C., Spector, D. L. The lncRNA Malat1 is dispensable for mouse development but its transcription plays a cis-regulatory role in the adult. Cell Rep. , 111-123 (2012).
- Trapnell, C., Roberts, A., Goff, L., Pertea, G., Kim, D., Kelley, D. R., Pimentel, H., Salzberg, S. L., Rinn, J. L., Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat Protoc. , 562-578 (2012).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved