
Method Article
Microgravimetry dissipative ללמוד את הדינמיקה איגוד של פוספוליפיד איגוד חלבון Annexin A2 כדי Bilayers השומנים הנתמכות על-ידי מוצק באמצעות מהוד קוורץ
* These authors contributed equally
In This Article
Summary
כאן, אנו מציגים טיפול נסיוני זה יכול להיות מועסק כדי לקבוע את הזיקות מחייב ואת מצב של האינטראקציה של פוספוליפיד מחייב חלבון ללא תווית annexin A2 עם קיבוע הנתמכות על-ידי מוצק bilayers (סאט גלובל) על ידי מדידת בו זמנית ספיגת המוני, המאפיינים viscoelastic של החלבון annexin A2.
Abstract
הטכניקה microbalance dissipative גביש הקוורץ הינה גישה פשוטה ונטולת תווית למדוד בו זמנית את המאפיינים ספיגת ו- viscoelastic המוני של מסה נספג/ותשמרו על משטחים חיישן, המאפשר את המידות של האינטראקציה החלבונים משטחים הנתמכות על-ידי מוצק, כגון bilayers השומנים, בזמן אמת, עם רגישות גבוהה. Annexins הם קבוצה מאוד ההכפלה של פוספוליפיד מחייב חלבונים המקיימים אינטראקציה הפיכה עם headgroups טעונים שלילית דרך התיאום של יונים של סידן. כאן, אנו מתארים את פרוטוקול הועסק לנתח באופן כמותי את הכריכה של annexin A2 (AnxA2) את השומנים מישורי bilayers שהוכנו על פני השטח של חיישן קוורץ. פרוטוקול זה ממוטבת כדי לקבל נתונים לשחזור ועמיד, כולל תיאור מפורט שלב אחר שלב. השיטה ניתן ליישם מחייב קרום חלבונים ויצירות bilayer אחרים.
Introduction
ממברנות הסלולר הן דינמיות ומורכבות מאוד מבנים. תערובת מורכבת של שומנים ממברנה, יחד עם ממברנה באופן עקיף ו/או ומקווים המשויך חלבונים, להרכיב את טופס microdomains. תהליכים פיזיולוגיים מפתח1מעורב ארגון מרחבי טמפו מתואמת של אלה microdomains ממברנה. קרום microdomain dynamics מונעים על-ידי הגומלין של קרום שומנים, וכן על ידי היכולת של חלבוני ממברנה פריפריאלי לזהות ולקיים אינטראקציה עם שומנים מועשר microdomains. הגיוס של חלבונים ליפידים ספציפי לעתים קרובות מושגת באמצעות זיהוי השומנים מודולים, כגון הומולוגיה של pleckstrin (PH) או C2 תחומים2,3. שיטות אנליטיות biophysical באמצעות מודל ממברנות הם המפתח להבנת העקרונות הבסיסיים המסדירים את התהליכים האלה ברמה המולקולרית.
Annexins, multigene-משפחה גדולה, ידועים ביכולתם לאגד ממברנה טעונים שלילית שומנים, בעיקר phosphatidylserine (PS), ב- Ca2 +-נשלט באופן2. סימן ההיכר השני של משפחת annexin היא הנוכחות של מקטע מבניים שנשמרת, annexin. חוזר, כלומר נוכח ארבעה או שמונה פעמים בנמלים Ca2 +- ו אתרי פוספוליפיד מחייב4. Ca2 +-השומנים תלויים באינטראקציה ממקם את annexins בעמדה מצוינת כדי לחוש ומשדרות Ca2 +-מתווכת איתות לממברנות היעד. באופן עקבי, annexins מסוגלים לגרום להיווצרות microdomains מועשר ב כולסטרול, phosphatidylinositol-4, 5-bisphosphate (PI (4.5) P2) ו- PS, הן מערכות ממברנות הסלולר ו/או מלאכותי5. פרוטוקול זה מתאר גישה זו כדי לנתח את האינטראקציה annexin-ממברנה בעזרת גביש הקוורץ Microbalance פיזור (QCM-D)6,7,8.
המרכיב הבסיסי ב microbalance הזה הוא קריסטל נדנוד משמש המשטח חיישן. ספיחה ו/או איגוד של מולקולות על פני השטח חיישן של הקטנת תהודה תדירות (נ) פרופורציונליים להעלייה במסה. אם פני השטח באופן שווה מצופה סרט, הכריכה של חומרים נוספים עלולים להפריע השלמות. המבנית של שכבה זו, שינויים ב- viscoelasticity (אנרגיה פיזור הגורם D) ניתן לנטר בנוסף. זוהי טכניקה נפוצה את האינטראקציה של חלבונים עם השומנים bilayers. בגישה זו, השומנים שלפוחית נספגים על גבי המשטח חיישן מצופה בהתאם. היווצרות ליפיד bilayer היא חיובית על חומרים מבוססי-סיליקה9,10, כמו שלפוחית לעיתים קרובות לא להיקרע על משטחים הידרופילית אחרים, כגון זהב11 מחומצן לאחר חשיפה UV-אוזון, TiO212או Al 2O313. שבר השלפוחיות המוגלתיות התגבשו משחרר שלב מימית, מובילים לשינויים האופייניים מסה והוללות. הדור של הנתמך על-ידי מוצק bilayers (סאט גלובל) מאת בועית פיוז'ן חזקה ופשוט והוא יכול לשמש כדי ליצור דגמים מורכבים המחקים ממברנות הסלולר.
גביש קוורץ dissipative microbalance היא טכניקה ללא תווית ורגיש. היתרון העיקרי הוא האפשרות לקבל מעיל חומר שיוצר סרט דק במידה מספקת על גבי המשטח, ובכך לספק מגוון רחב של יישומים בתחומים מגוונים מחקר. האינטראקציה חלבון-הממברנה הוא ציין בזמן אמת, ולא ניתן לנתח את התוצאות ישירות. השטח חיישן אותו יכול לשמש במדידות עוקבות (לאחר ביצוע ניקוי מינימלי כפי שמתואר פרוטוקול זה), ובכך מאפשר בקרה פנימית מדויקת, של comparability בין analytes.
Protocol
הערה: מאגרי לסנן באמצעות מסנן 0.22-מיקרומטר, degassed על ידי ואקום לשעה.
1. השומנים שלפוחית הכנה
- השתמש צינורות זכוכית 2-mL. להמיס כל השומנים, 1,2-dioleoyl-sn-glycero-3-phosphocholine (DOPC), 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC), 1-palmitoyl-2-oleoyl-sn-glycero-3-phospho-L-serine (אבא), כולסטרול (חול), בתערובת של כלורופורם / מתנול (50: 50 v/v) כדי להכין פתרון השומנים ברור 5 מ מ. להמיס 1,2-dioleoyl-sn-glycero-3-phospho-(1'-myo-inositol-4',5'-bisphosphate) (PI (4.5) P2) בתערובת של כלורופורם/מתנול/מים (20:9:1 v/v).
- לשלב ליפידים מומס ביחס טוחנת הרצוי של סבא/POPC (80/20), P POPC/PI (4.5)2 (95/5), אבא/POPC/חול (60/20/20), /Chol2POPC/אבא/PI (4.5) P (60/17/3/20) ו- P POPC/DOPC/אבא/PI (4.5)2(/Chol (37/20/20/3/20) טבלה 1) צינורות זכוכית 10-mL.
הערה: הסכום הסופי של השומנים הכולל הוא 500 µg. - להתנדף ממיסים אורגניים באמצעות זרם יבש של חנקן. להשאיר את התערובת השומנים מערכת ואקום גבוה (lyophilization) עבור 3 שעות להסיר את עקבות שיורית של ממיסים.
הערה: התוצאה סרט ברור יבש. - Resuspend הסרט השומנים ב 1 מ"ל של מאגר ציטראט (10 מ מ trisodium-ציטראט, 150 מ מ NaCl, pH 4.6). דגירה השומנים ההשעיה ב 60 מעלות צלזיוס (טמפרטורה זו הוא בסביבות 10 ° C מעל טמפרטורת המעבר לשלב השומנים ההיתוך הגבוהה ביותר בתערובת) במשך 30 דקות אמבט מים, ו מערבולת זה נמרצות כל 5 דקות.
הערה: התוצאה היווצרות שלפוחית גדול, multilamellar (MLVs). שמור את המתלים מעל טמפרטורת המעבר. - מחממים את מכבש (ב 60 מעלות צלזיוס, במקרה זה) מצוידים עם קרום גודל הנקבוביות פוליקרבונט קוטר 50-nm מעל טמפרטורת המעבר (אשר נמצא כאן 40-50 מעלות צלזיוס) למשך 30 דקות.
- לטעון את המתלים קמפנילה MLV לתוך המתקן preheated ולהעביר בעדינות את תערובת x 31 דרך קרום פוליקרבונט כדי ליצור unilamellar קטן שלפוחית (רכבי השטח)14. לשמור על הטמפרטורה מעל טמפרטורת המעבר.
- להעביר את המתלים רכב שטח כלי פלסטיק תגובה 2 מ"ל ולהוסיף למאגר ציטראט (ראה שלב 1.4) כדי להביא את עוצמת הקול הסופי 2 מ"ל.
הערה: זו תניב ריכוז השומנים הסופי של 250 µg/mL.
2. טיפול החיישנים קוורץ
הערה: תמיד לנהל את החיישנים קוורץ עם פינצטה.
- דגירה ארבעה חיישני הוכנס בעל טפלון בפתרון מרחביות 2% עבור ≥ 30 דקות לשטוף אותם בהרחבה עם מים אולטרא טהורים לחלוטין להסיר זמנים תוססים ולתת להן להתייבש באמצעות זרם של ארגון יבש או חנקן.
- השתמש מערכת לניקוי פלזמה כדי להסיר לחלוטין את כל מזהמים. הוסף את החיישנים יבש בבית הבליעה לניקוי פלזמה, לפנות את החדר, לשטוף את זה 3 x עם חמצן. הפעל הפלזמה מנקה על. השתמש בפרמטרים תהליך הבאים: עונה 1 פרק 10-4 טנדר של גוה של לחץ כוח גבוהה בתדר רדיו (RF), 10 דקות של זמן תהליך. כבה את המכשיר, להוציא את החיישנים.
3. microbalance מבצע
הערה: מערכת microbalance עם ארבע צ'יימברס זרימה מבוקרי טמפרטורה בתצורת מקבילים, מחוברת משאבה סחרור והגדר את קצב זרימה של 80 µL/דקה, נעשה שימוש. במצב זרימה פתוחה, המאגר היה שאוב מן המאגר מזין לתוך הטנק המקבל. במצב לולאה, הטנק המקבל היה מקושר עם המאגר מזין כדי ליצור לולאות סגורות. הטמפרטורה הוגדר 20 º C.
- בזהירות לעגן את החיישנים ניקיתי פלזמה לתוך התאים זרימה 4, באמצעות פינצטה. למנוע לחץ על או פיתול של התאים, הצינורות שעלול לגרום דולף.
- לרוקן את המערכת עם מאגר ציטראט (10 מ מ trisodium-ציטראט, 150 מ מ NaCl, pH 4.6) במצב של זרימה פתוחה במשך 10 דקות.
הערה: זה דורש בדיוק 3.2 מ"ל של המאגר, אבל מומלץ להשתמש עודף של מאגר (10 מ"ל). - הפעל את התוכנית. להתחיל להקליט כל שינוי תדירות, פיזור של הצליל הבסיסי הראשון (n = 1), גוון (n = 3-13) באמצעות התוכנה, עד התוכניות הבסיסיות של תדירות והוללות יציבים (זה לוקח בערך 40-60 דקות).
הערה: רמת הרעש תדר (שיא אל שיא) צריך להיות נמוך מ- 0.5 הרץ, ובמשך את פיזור, נמוך יותר 0.1·10-6, עם הסחף מקסימלי (ב תמיסה מימית) של 1 הרץ/h בתדר, 0.3·10-6/h בפיזור. - כאשר התוכניות הבסיסיות של יציבה, להחיל את המתלים רכב שטח במאגר ציטראט (מ 2 בשפופרת קטנה). באמצעות כלי התגובה, להסיר 1.5 מ"ל נפח מת. לאחר מכן סגור את המערכת במצב של זרימה לולאה. להקליט את המשמרת תדר/פיזור לעוד 10 דקות.
הערה: במהלך תקופה זו, השלפוחיות המוגלתיות התפשטה על פני2 SiO, נתיך. לגבש bilayer רציף15,16 (שלב 2 ב 1 דמויות איור 2, איור 3). ספיחה רכבי שטח על פני השטח של החיישן הוא דו-phasic ויש לו תדר טיפוסית מינימום ומקסימום ב פיזור. בסיס יציב תדר/פיזור חדש עם שינוי התדירות האופיינית (תלוי הרכב השומנים) מ 26-29 הרץ (ראה טבלה 1) מציין bilayer מתמשך על פני השטח. - מתי סאט גלובל יציב (ראה שלב 3.4), equilibrate את המערכת עם המאגר המצטבר (10 מ מ HEPES, 150 מ מ NaCl, pH 7.4) בריכוזים הנדרש Ca2 + (ועד 50 מיקרומטר עד 1 מ"מ CaCl2, בהתאם הניסוי) במצב זרימה פתוחה עבור 40 דקות.
- להוסיף את החלבון (כאן, AnxA2) בריצה מאגר האישורים המכילה2 + (ראה שלב 3.5). לבצע את היישום של החלבון במצב זרימה לולאה עד למצב יציב של שיווי משקל (שלב 3 איור 1, איור 2, איור 3).
הערה: ריכוז חלבון עלול בטווח של 1 עד 400 ננומטר. ספיחה חלבון גורמת לשינוי ריכוז תלוית תדירות המשקף את ספיחה מסה (חלבון). - מביצועם החלבון המאוגד על-ידי chelating Ca2 + יונים עם 5 מ מ EGTA במאגר פועל במצב זרימה פתוחה (שלב 4 דמויות 1 , איור 2).
הערה: החלמה של תדר, פיזור לקו הבסיס סאט גלובל מציינת של הפיכות הכולל של חלבון מחייב. האגודה-דיסוציאציה מחזורים יכול לחזור כדי להשוות ריכוזים שונים או חלבונים.
4. microbalance ניקוי
הערה: לבצע הליך ניקוי מינימלי אחרי כל מדידה.
- צור מחדש את המערכת microbalance עם 50 מ של ddH2O במצב פתוח זרימה רציפה, להסיר את הצינורות מיכל המים, לאפשר למערכת להתייבש.
- להסיר את החיישן קריסטל ובזהירות לנקות אותו עם הפתרון מרחביות 2% באמצעות בעל טפלון (ראה שלב 2.1).
- יבש את חלקי הפנים מודול הזרימה שבו הוצב החיישן גלוי.
הערה: לבצע אינטנסיבי של ניקוי הליך לאחר סדרה של 10 מדידות. - שתנקה את המערכת עם פתרון מרחביות 2% (50 מ"ל) ב- 40 ° צ' (התקנה לתוכנה) במצב זרימה רציפה פתוח (tube), באמצעות קצב זרימה של µL 20/min עבור המורחבת במגע עם הנוזל.
- נקה עם 250 מ של ddH2O במצב פתוח זרם בקצב הזרימה של µL 160/min.
הערה: לאחר 4 חודשים, לבצע את שגרת ניקוי מקיף על-פי הוראות היצרן.
תוצאות
הירידה לתדר המתאים (Δf) מופיע בצורה ליניארית עם המסה הספוחה (Δm), כפי שהוגדר על-ידי המשוואה Sauerbrey. 17

. הנה, f הוא לתדר המתאים, Cf הוא קבוע התלוי המאפיינים הגיאומטריים והפיזי של המינרל נתון לתדר המתאים, A הוא שטח הפנים חיישן.
ברוב היישומים, השכבה הספוחה אינה נוקשה לגמרי אבל viscoelastic. לשבור וכתוצאה מכך של תנודה חיישן קוורץ הוא המכונה פיזור (D). פיזור בפיקוח שינויים (ΔD) לתאם עם המאפיינים viscoelastic של המוני מאוגד18 והם מוגדרים כדלקמן8.

כאן, Eהתפוגג היא אנרגיה איבדה בתקופה תנודה אחת, Eמאוחסנת האנרגיה הכוללת של החיישן בחופשיות נדנוד.
כדי לנתח ולכמת את הפרמטרים מחייב, תדירות isotherms מופקות באמצעות התוויית את המשמרות תדירות שיווי משקל (ΔΔfe) כנגד ריכוז חלבון. ΔΔfe מוגדרת בתור

כאן, Δft1 מייצג את תחילת החלבון ספיחה, Δft2 המדינה שיווי משקל. לא-ליניאריות פריסטלטיות יכול להתבצע באמצעות הרחבה של הגבעה של המשוואה לאנגמיור כדלקמן6,8.

כאן, ΔΔfמקסימום הוא ה-ΔΔfe ריכוז חלבון וכתוצאה מכך מרבי (קולח) מחייב, Kd הוא קבוע דיסוציאציה נראית לעין עבור החלבון/קרום מורכבים, n הוא המקדם היל.
המקדם היל (n) מתאר את cooperativity של האיגוד. עבור n = 1, המודל ספיחה היל הוא איזותרמה לאנגמיור פשוטה (האתרים מחייב שוויון ומולקולות כל לאגד בנפרד ליפידית). אם בשל n 1, ליגנד מאוגד משתנה קרום מחייב אהדה ליגנדים אחרים, גם הגדלה (n > 1, cooperativity חיובי) או הקטנה (n < 1, cooperativity שלילי) את הזיקה.
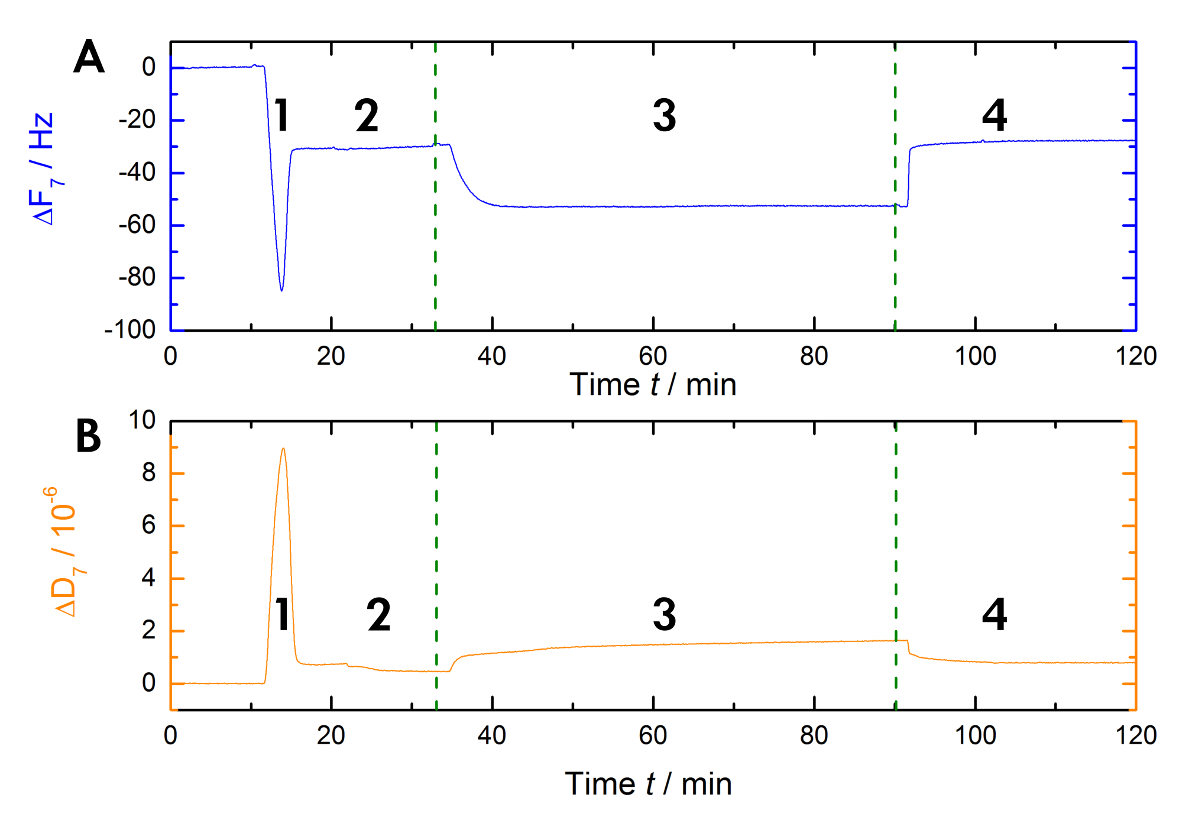
איור 1 מראה סכימטי של זרימת העבודה ניסיוני המשמש במעבדה שלנו כדי למדוד את המשמרות של תהודה ותדר במהלך Ca2 +-מחייבים תלות, על שחרורו של AnxA2 כדי ליפידית בשלב נוזלי. הקלטת למופת מוצג באיור2. איור 2 א מציג את ההקלטה של העקומה תדירות ומציג איור 2B את המשמרות פיזור. ירידה בולטת תדר על התוספת של ליפוזומים (איור 2א [שלב 1]) מציין ספיחה שלהם. בגלל השלפוחיות המוגלתיות מאגר מלא לא נוקשה, אבל viscoelastic, פיזור מגדילה (איור 2B [שלב 1]). לאחר מכן, השלפוחיות המוגלתיות התגבשו קרע. המהדורה בו-זמני של המאגר בתוך השלפוחיות המוגלתיות מקטין את המסה הספוחה עד רמה יציבה (איור 2א [שלב 2]). ראוי לציין, התוספת של שלפוחית תוצאות שינוי פיזור גבוה, בזמן המשמרת בתגובה bilayer הוא הרבה יותר קטן בגלל אופי הומוגני קשיח סאט גלובל (איור 2B [שלב 2]). שלב 3 ברשומות איור 2A ו- 2B הכריכה של AnxA2 כדי ליפידים, אשר מוסיפה מסה, כפי שנראה את המשמרת תדר נקי, אך לא מפריע המבנה bilayer, כמצוין על-ידי שינוי קטן רק פיזור. כאשר Ca2 + מוסר מאת מכלט EGTA (איור 1 , איור 2 [שלב 4]), AnxA2 dissociates מהסרט השומנים. התדירות, וכן הקלטות פיזור, משמרת את רמות אובחנה bilayer בלבד (השווה בשלבים 2 ו- 4 איור 2A ו- 2B), המציינת כי איגוד AnxA2 תלויה לחלוטין Ca2 + , וכי הסרט שומנים בדם נשאר שלם.
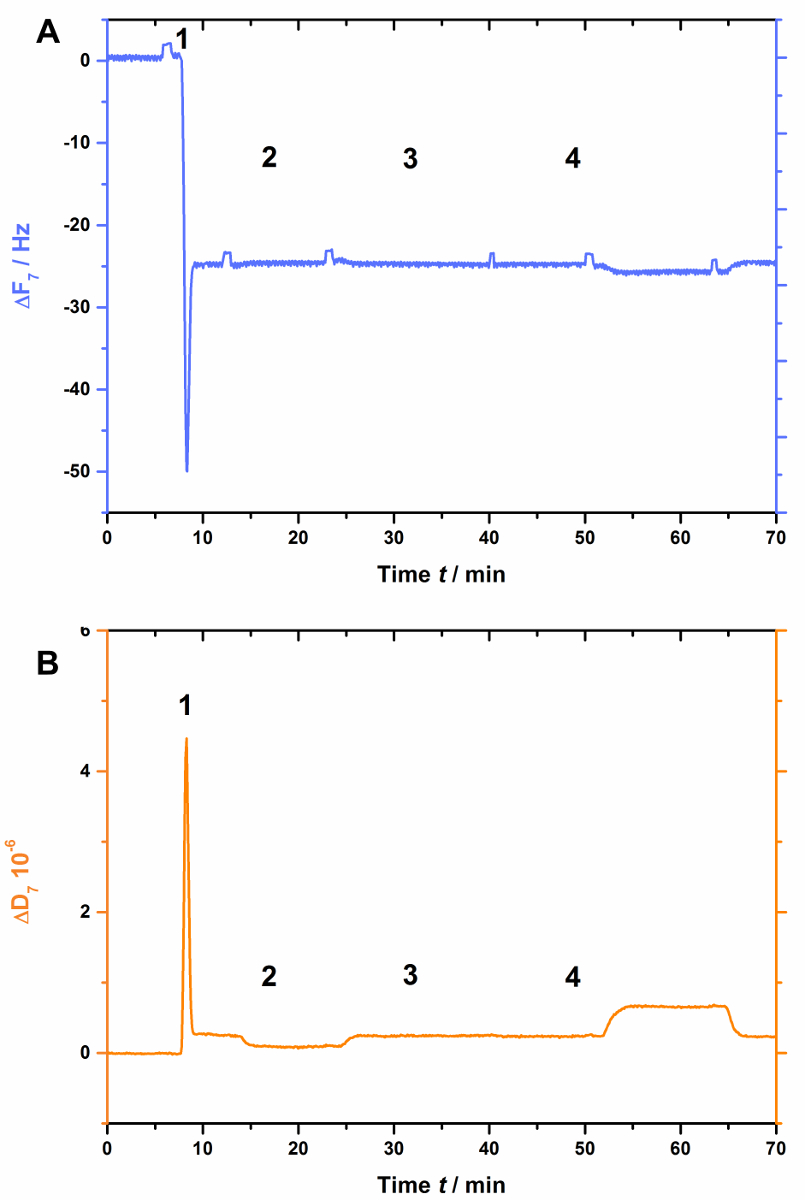
AnxA2, כמו ביותר של annexins, תלוי ליפידים טעונים שלילית כגון PS. זה נראה בבירור מתי אבא נעדר ב ליפידית (איור 3). איור 3 א מציג את ההקלטה של העקומה תדירות ומציג איור 3 את המשמרות פיזור. הערה כי התדירות משתנה על בסיס יציב ב-25 הרץ, עדיין פיזור אינו משתנה (איור 3B [שלב 2]), מצביע על כך צורה bilayer נאותה. עם זאת, שאין שינוי תדר (איור 3א') או פיזור (איור 3ב) הם נצפו אחרי תוספת של AnxA2 בנוכחות Ca2 + (איור 3A ו- 3B [שלב 3]) או EGTA (איור 3A ו- 3B [שלב 4]), כפי AnxA2 לא יכול לקיים אינטראקציה עם הסרט שומנים בדם.

איור 1 : מודל גרפי של זרימת העבודה ניסיוני. זרימת עבודה זו ממחישה את הספיגה שלפוחית השטח חיישן הידרופיליות (שלב 1), שלפוחית הפיוז'ן/העלתה שמוביל היווצרות סאט גלובל (שלב 2), ו- Ca2 +-תלויים ספיחה (שלב 3), EGTA תלוית desorption של AnxA2 (שלב 4). אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 2 : הקלטה למופת. אלה מראות (A) תלויי-זמן הפיקוח על התדר תהודה צליל עילי 7, (B) פיזור משמרות של החיישן קוורץ במהלך המדידה. היישום של ליפוזומים גורמת ירידה מהירה הבסיס תדר, ואילו הבסיס פיזור מגדילה (שלב 1). הייצוב של התוכניות הבסיסיות של מצביע על היווצרות bilayer (שלב 2). AnxA2 (200 ננומטר) ספיחה (בנוכחות Ca2 +) על גבי ליפידית סבא המכילות מוסיף מסה מבלי לשנות באופן משמעותי את פליטת, המציין כי הסרט השומנים לא מוטרד (שלב 3). ההתאוששות של הבסיס תדר על Ca2 + קלאציה עם EGTA מציין את desorption הכולל של החלבון (שלב 4). אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 3 : ניסוי בקרה שלילית, הוכחת כי AnxA2 לא לאגד SLBs בהיעדרו של אבא. לוחות אלה מראים התוספת של ליפוזומים ויצירת סאט גלובל (שלבים 1 ו- 2). שאין שינוי תדר (א) או (B) פיזור ניכרים לאחר התוספת של AnxA2 (שלב 3; 200 ננומטר, בנוכחות Ca2 +) או EGTA (שלב 4). אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}
| קומפוזיציה | ΔΔF הרץ לאחר היווצרות SLBs | ΔΔD * 10-6 לאחר היווצרות סאט גלובל |
| POPC/אבא (80: 20) | 26.3 ± 0.2 | 0.26 ± 0.03 |
| POPC/PI (4.5) P2 (95: 5) | 26.5 ± 0.5 | 0.31 ± 0.02 |
| POPC/אבא/חול (60: 20: 20) | 29.2 ± 0.2 | 0.45 ± 0.09 |
| POPC/אבא/פאי /Chol2P (4.5) (60: 17: 3:20) | 29.6 ± 0.6 | 0.43 ± 0.10 |
| POPC/DOPC/אבא/פאי /Chol2P (4.5) (37: 20: 20: 3:20) | 29.4 ± 0.4 | 0.39 ± 0.14 |
טבלה 1: נתוני הרכב ואת היווצרות ליפיד סאט גלובל 7 .
Discussion
כדי לענות על שאלות לגבי היחס מבנה פונקציה של ממברנות הסלולר הן באופן כמותיים ואיכותניים, תא הרווחים ביולוגיה לאין שיעור משימוש של גישות biophysical בהתבסס על ומבוססת, בשימוש נרחב בטכניקות , כולל אטומי לכפות מיקרוסקופ (AFM), משטח פלזמון תהודה (SPR), והעסיק הטכניקה QCM-D כאן. הראנו במחקרים קודמים חלבונים annexin לאגד ב Ca2 +-תלוי באופן למוח קיבוע עם זיקה גבוהה. אנו משתמשים בתדר, פיזור משמרות של 7 צליל עילי (Δf7) כי זה מייצג את הפשרה הטובה ביותר של זיהוי רגישות, תנודה יציבות.
טכניקה זו מאפשרת גם תיאור כמותי של אינטראקציה חלבון ממברנלי. AnxA2 מחייב הקרום מאופיינת cooperativity חיובי זה מתווך על ידי קבוצת המחשבים ליבת annexin שנשמרת ותלוי הנוכחות של כולסטרול. הנתונים הכמותיים שהושג עבור AnxA2 ו- AnxA8 הם דיווחו פירוט במקום6,8.
ישנם שלבים קריטיים רבים פרוטוקול זה. להשתמש ליפוזומים באופן מיידי; אחרת, שלפוחית קטנה אולי הפתיל לתוך שלפוחית גדול יותר עם פחות מתח, שמוביל עיכוב של היווצרות bilayer השומנים. לשמור על טמפרטורת חום קבועה במהלך המדידות. בכל בסטיות הקטנות בטמפרטורה יוצרת שינוי תדירות והוללות לא זניח. להימנע בועות אוויר; אחרת, המערכת יציב והם לא יקימו תוכנית בסיסית.
המשוואה Sauerbrey מאפשרת המרה ישירה של התדר שנצפה שינוי שינויים במסה, ולכן, בשימוש נרחב. ההנחה של מתאם ליניארי בין שינוי תדר תהודה המסה נוספה רק מחזיקה נכון עבור רכיבי ויוצרים סרט נוקשה ובלתי אחיד על פני השטח חיישן. המשוואה Sauerbrey אינם יכולים לשמש עבור viscoelastic adsorbents כגון מים עתירי חלבון סרטים, שכבות השומנים במים מואגד, או אפילו הספוחה תאים. . הנה, מודלים מתמטיים מורכבים יותר נדרשים. לכן, חשוב מאוד לעקוב בו-זמנית אחר השינויים בתדירות והוללות. כדי לזהות שינויים מבניים במהלך המדידה, ΔD לעומת Δf יחסי ניתן להתוות, עם קו ישר המציין אין שינויים הסתגלותי.
היתרון העיקרי של טכניקה זו היא האפשרות לשימוש במגוון רחב מאוד של חומרים דיאלקטריים. יתר על כן, זה שיטה אמינה וישירה ללמוד מגוון רחב של אינטראקציות macromolecular, כמו היווצרות נאותה של הסרטים ציפוי פני השטח (כגון סאט גלובל), כמו גם אינטראקציות חלבון-השומנים נוספות, ניתן לנטר באופן מקוון.
פרוטוקול זה יכול להיות מוחל על חלבונים אחרים ממברנה-אינטראקציה, לדוגמה, בר תחום חלבונים19, ERM (אזרין, radixin ו moesin) חלבון המשפחה בעלת תפקיד חשוב בממברנה-שלד התא הצמדה20,21 ,22, או חלבונים המכילים דומיינים C2 או PH. יתר על כן, מגוון רחב של יישומים של טכניקה זו ללמוד חומרים ביולוגיים בהצלחה שפורסם, ובכך לבסס QCM כפלטפורמה ניסיוני ולסדרם ללמוד את האינטראקציות מכלולים מורכבים יותר macromolecular או אפילו תאים23,24.
Disclosures
המחברים אין לחשוף.
Acknowledgements
עבודה זו נתמכה על ידי פתוח תחת מענקים SFB 858/B04, סיפורה ה 1003, SFB 1348/A04, SFB 1348/A11.
Materials
| Name | Company | Catalog Number | Comments |
| Chemicals | |||
| Calciumchloride | Merck | 017-013-00-2 | 99% |
| Chloroform | Roth | 4432.1 | 99% |
| DOPC | Avanti | 850375P | |
| EGTA | PanReac AppliChem | A0878 | 99% |
| HEPES | PanReac AppliChem | A1069 | |
| Methanol | PanReac AppliChem | A3493 | |
| PiP2 | Avanti | 850155P | |
| POPC | Avanti | 850457P | |
| POPS | Avanti | 840034P | |
| Sodiumchloride | PanReac AppliChem | A1149 | |
| SDS | Roth | 183 | |
| Trisodium citrate | PanReac AppliChem | A3901 | |
| Equipment | |||
| Extruder Liposofast | Avestin | ||
| Qsense E4 Analyzer | Qsense | ||
| QSense Dfind | Qsense | ||
| Pump IPC 4 | Ismatec | ISM 930 | |
| QSX 303 SiO2 Silicon dioxide 50nm | Qsense | QSX 303 | |
| PC Membranes 0.05μm | Avanti polar lipids | 610003 | |
| OriginPro | OriginLab Corporation | Version 8 and 9 |
References
- Simons, K., Toomre, D. Lipid rafts and signal transduction. Nature Reviews Molecular Cell Biology. 1 (1), 31-41 (2000).
- Gerke, V., Creutz, C. E., Moss, S. E. Annexins: linking Ca2+ signalling to membrane dynamics. Nature Reviews Molecular Cell Biology. 6, 449-461 (2005).
- Mim, C., Unger, V. M. Membrane curvature and its generation by BAR proteins. Trends in Biochemical Science. 37, 526-533 (2012).
- Rescher, U., Ruhe, D., Ludwig, C., Zobiack, N., Gerke, V. Annexin 2 is a phosphatidylinositol (4,5)-bisphosphate binding protein recruited to actin assembly sites at cellular membranes. Journal of Cell Science. 117, 3473-3480 (2004).
- Gerke, V., Moss, S. E. Annexins: from structure to function. Physiological Reviews. 82, 331-371 (2002).
- Heitzig, N., et al. Cooperative binding promotes demand-driven recruitment of AnxA8 to cholesterol-containing membranes. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1863 (4), 349-358 (2018).
- Drücker, P., Grill, D., Gerke, V., Galla, H. J. Formation and characterization of supported lipid bilayers containing phosphatidylinositol-4,5-biphosphate and cholesterol as functional surfaces. Langmuir. 30, 14877-14886 (2014).
- McConnell, H. M., Watts, T. H., Weis, M. R., Brian, A. A. Supported planar membranes in studies of cell-cell recognition in the immune system. Biochimica et Biophysica Acta - Reviews on Biomembranes. 864, 95-106 (1986).
- Anderson, T. H., et al. Formation of supported bilayers on silica substrates. Langmuir. 25, 6997-7005 (2009).
- Pfeiffer, L., Petronis, S., Koper, I., Kasemo, B., Zach, M. Vesicle adsorption and phospholipid bilayer formation on topographically and chemically nanostructured surfaces. Journal of Physical Chemistry B. 114, 4623-4631 (2010).
- Cho, N. J., Jackman, J. A., Liu, M., Frank, C. W. pH-Driven assembly of various supported lipid platforms: a comparative study on silicon oxide and titanium oxide. Langmuir. 27, 3739-3748 (2011).
- Jackman, J. A., Tabaei, S. R., Zhao, Z., Yorulmaz, S., Cho, N. J. Self-Assembly formation of lipid bilayer coatings on bare aluminium oxide: overcoming the force of interfacial water. ACS Applied Materials & Interfaces. 7, 959-968 (2015).
- Olson, F., Hunt, C. A., Szoka, F. C., Vail, W. J., Papahadjopoulos, D. Preparation of liposomes of defined size distribution by extrusion through polycarbonate membranes. Biochimica et Biophysica Acta(BBA) - Biomembranes. 557 (1), 9-23 (1979).
- Richter, R., Mukhopadhyay, A., Brisson, A. Pathways of lipid vesicle deposition on solid surfaces: a combined QCM-D and AFM study. Biophysical Journal. 85 (5), 3035-3047 (2003).
- Richter, R. P., Bérat, R., Brisson, A. R. Formation of solid-supported lipid bilayers: an integrated view. Langmuir. 22 (8), 3497-3505 (2006).
- Sauerbrey, G. Verwendung von Schwingquarzen zur Wägung dünner Schichten und zur Mikrowägung. Zeitschrift für Physik A Hadrons and Nuclei. 155 (2), 206-222 (1959).
- Rodahl, M., Höök, F., Krozer, A., Brzezinski, P., Kasemo, B. Quartz crystal microbalance setup for frequency and Q-factor measurements in gaseous and liquid environments. Review of Scientific Instruments. 66 (7), 3924-3930 (1995).
- Drücker, P., Pejic, M., Grill, D., Galla, H. J., Gerke, V. Cooperative binding of annexin A2 to cholesterol-and phosphatidylinositol-4, 5-bisphosphate-containing bilayers. Biophysical Journal. 107 (9), 2070-2081 (2014).
- Galic, M., et al. External push and internal pull forces recruit curvature-sensing N-BAR domain proteins to the plasma membrane. Nature Cell Biology. 14 (8), 874-881 (2012).
- Fehon, R. G., McClatchey, A. I., Bretscher, A. Organizing the cell cortex: the role of ERM proteins. Nature Reviews Molecular Cell Biology. 11 (4), 276-287 (2010).
- Braunger, J. A., Kramer, C., Morick, D., Steinem, C. Solid supported membranes doped with PIP2: influence of ionic strength and pH on bilayer formation and membrane organization. Langmuir. 29 (46), 14204-14213 (2013).
- Bianco, M., et al. Quartz crystal microbalance as cell-based biosensor to detect and study cytoskeletal alterations and dynamics. Biotechnology Journal. , 1700699(2018).
- Chen, J. Y., Penn, L. S., Xi, J. Quartz crystal microbalance: Sensing cell-substrate adhesion and beyond. Biosensors and Bioelectronics. 99, 593-602 (2018).
- Bragazzi, N. L., et al. Quartz-Crystal Microbalance (QCM) for Public Health: An Overview of Its Applications. Advances in Protein Chemistry and Structural Biology. Donev, R., et al. 101, Academic Press. 149-211 (2015).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved