
Method Article
גישה לחקר תעתיק תלוי צורה ברמת תא יחיד
In This Article
Summary
נייר זה מציג שיטות לגידול מיוציטים לבביים עם צורות שונות, המייצגות פתולוגיות שונות, ומיון מיוציטים לביים דבקים אלה בהתבסס על המורפולוגיה שלהם ברמת תא אחד. הפלטפורמה המוצעת מספקת גישה חדשנית לתפוקה גבוהה והקרנת סמים עבור סוגים שונים של אי ספיקת לב.
Abstract
סוגים שונים של היפרטרופיה לב היו קשורים נפח מוגבר של מיוציטים לב (CMs), יחד עם שינויים מורפולוגיה CM. בעוד ההשפעות של נפח התא על ביטוי גנים ידועים היטב, ההשפעות של צורת התא אינן מובנות היטב. מאמר זה מתאר שיטה שתוכננה לנתח באופן שיטתי את ההשפעות של מורפולוגיה CM על ביטוי גנים. הוא מפרט את הפיתוח של אסטרטגיית השמנה חד-תאית חדשנית, ולאחר מכן מלווה בה רצף mRNA חד-תאי. שבב micropatterned תוכנן גם, אשר מכיל 3000 מיקרופטרן פיברונקטן בצורת מלבני. הדבר מאפשר לגדל CMs באורך ברור:יחס גובה-רוחב (AR), המתאים לסוגים שונים של אי ספיקת לב (HF). המאמר מתאר גם פרוטוקול שתוכנן לאסוף תאים בודדים מהתבנית שלהם, באמצעות דוגם תא מיקרו-צנרת חצי אוטומטי, ולהזריק אותם בנפרד למאגר נפרד. זה אפשר פרופיל transcriptomes של CMs יחיד עם מורפוטיפים גיאומטריים מוגדרים ולאפיין אותם על פי מגוון רחב של תנאים נורמליים או פתולוגיים: קרדיומיופתיה היפרטרופית (HCM) או עומס ים / קונצנטרי לעומת cardiomyopathy מורחב (DCM) או preload/ אקסצנטרי. לסיכום, מאמר זה מציג שיטות לגידול CMs עם צורות שונות, המייצגות פתולוגיות שונות, ולמיון CMs נדבקים אלה בהתבסס על המורפולוגיה שלהם ברמה של תא אחד. הפלטפורמה המוצעת מספקת גישה חדשנית לתפוקה גבוהה והקרנת סמים עבור סוגים שונים של HF.
Introduction
על פי ארגון הבריאות העולמי, מחלות לב וכלי דם (CVD) היא גורם מרכזי לתחלואה ותמותה ברחבי העולם. CVD משפיע באופן דרמטי על איכות חייהם של אנשים ויש לו השפעה חברתית-כלכלית עצומה. Cardiomyopathies, כגון HCM ו DCM, הם הפרעות עיקריות של שריר הלב וגורמים עיקריים של HF היו קשורים עם תחום גבוה ותמותה. ישנם גורמים רבים של HF, כולל השפעות סביבתיות, כגון זיהומים וחשיפה לרעלים או תרופות מסוימות8. HF יכול להיגרם גם על ידי נטייה גנטית, כלומר מוטציות9. הוא האמין כי השינויים בהרכב הגנטי המשפיעים על מולקולות מטריצה חוץ תאית (ECM), integrins או חלבונים cytoskeletal יכול להיות אחראי mechanosensation לקוי וסוגים שונים של מחלותלב 10.
התכונה העיקרית של HCM היא היפרטרופיהבלתי מוסברת של החדר השמאלי 11, ולפעמיםשל החדר הימני 12, וזה לעתים קרובות מציג עם מעורבות דומיננטית של המחיצה התערבות. HCM מאופיין גם בתפקוד דיאסטולי אי סדר myocyte ופיברוזיס13. ברוב המקרים, מערכת ההתכווצות של הלב מושפעת מוטציות בחלבונים סרקומריים, מה שמוביל להתכווצות מוגברת של myocytes14. לעומת זאת, DCM מאופיין באופן מורחב של חדר אחד, או שניהם, ויש לו אטיולוגיה משפחתית ב 30% עד 50% מהמקרים15. DCM משפיע על מגוון רחב של פונקציות תאיות, המוביל התכווצות לקויה של myocytes, מוות תאים ותיקון פיברוטי16.
גנטיקה הראתה כי סוגים מסוימים של מוטציות לכפות CMs יחיד לאמץ מאפייני צורה ספציפיים במהלך HCM3, כל כך תאים בצורת מרובע עם אורך:רוחב AR כי הוא כמעט שווה 1:14 (AR1). אותו הדבר נכון לגבי DCM, עם תאים מוארך עם AR כי הוא כמעט שווה 11:1 (AR11). בנוסף, HF יכול להיגרם על ידי עומס יתר מוגבר (למשל, ביתר לחץ דם). במקרים אלה, המודינמי דורש לכפות על CMs לקחת על צורות מרובעות, על פי החוק של Laplace, ואת AR משתנה מ 7:15 (AR7) כדי 1:16,7. HF יכול להיגרם גם על ידי עלייה בעומס מראש (למשל, בתנאים שמובילים עומס יתר של נפח). כאשר זה קורה, האילוצים הביופיזיים מאלצים CMs להאריך וה- AR משתנה מ- 7:1 ל- 11:1.
פעילות איתות בממברנות תלויה בפרמטרים של גאומטריית תאים גלובליים, כגון AR הסלולרי, גודל, שטח פני הממברנה ועקמומיות קרום18. כאשר CMs חולדה ילודים היו מצופה על מצעים שהיו בדוגמת כדי להגביל את התאים באורך מסוים:רוחב AR, הם הפגינו את תפקוד ההתכווצות הטובה ביותר כאשר היחסים היו דומים לתאים בלב מבוגר בריא. לעומת זאת, הם ביצעו גרוע כאשר היחס היה דומה לאלה של myocytes בלב כושל19. בשלבים המוקדמים של היפרטרופיה, התאים הופכים רחבים יותר, כפי שמשתקף על ידי עלייה באזור חוצה חתך. HF מתרחשת בשלבים מאוחרים יותר של היפרטרופיה ותאים מופיעים בדרך כלל מוארך. לכן, אין זה מפתיע כי במודלים חולדת vivo של היפרטרופיה כרונית דיווחו על עלייה באורך myocyte החדר השמאלי של כ 30%20, אבל CMs למבוגרים ממודל עכבר transgenic שטופלו באופן חריף עם גירויים היסטרופיים במבחנה הפגינו עליותדומות ברוחב התא במקום 21.
רצף RNA חד-תאי, המאפשר ניתוח מדויק של התמלילים של תאים בודדים, מחולל כעת מהפכה בהבנת הביולוגיה של התאים. טכנולוגיה זו הייתה השיטה המועדפת בכל שאלה כיצד צורות תא בודדות השפיעו על ביטוי גנים. השווינו תאים בודדים עם צורות שונות, במיוחד עם ARs של 1:1, 7:1 או 11:1. זה נעשה על ידי זריעת CMs חולדת ילודים על שבב שתוכנן במיוחד מלא micropatterns מצופה פיברונקטין2 עם ARs מוגדר של 1:1, 7:1 או 11:1. המיקרו-פטרנים היו מפוברקים באמצעות טכנולוגיית פוטוליתוגרפיה. המיקרו-פטרנים היו מצופים בפיברונקטין, מוקפים במשטח ציטופובי. לכן, CMs יהיה לצרף, להפיץ וללכוד את AR המוגדר של micropatterns על ידי גידול אך ורק על השקוע פיברונקטין, תוך הימנעות האזור cytophobic. המיקרו-פטרנים אינם מעוצבים היטב. במקום זאת, רמת פיברונקטין היא בדיוק באותו גובה של האזור ציטופובי שמסביב. זה סיפק תנאים דומים לתאים גדלים בצלחת פטרי, כמו אין מתח מהקירות שמסביב. בנוסף, שטח הפנים של מיקרו-פטרנים עם שותפים שונים שווים.
היו שני היבטים חשובים במיוחד של העיצוב הניסיוני, מה שהוביל לשימוש של רצף RNA של תא יחיד במקום רצף RNA בתפזורת. ראשית, רק כמה אחוזים של micropatterns יכול להיות כבוש על ידי תא יחיד. שנית, לפעמים תא יחיד אינו תופס באופן מלא את משטח המיקרו-פטרן. תאים בודדים שמכסים לחלוטין משטח מיקרו-פטרן חייבים להיבחר לניתוח רנ"א חד-תאי. מכיוון שרק תת קבוצה של התאים המצופה על שבב עונה על שני הקריטריונים, זה לא היה אפשרי פשוט לנסות את השבב כולו ולאסוף את כל התאים עבור רצף RNA בתפזורת. יש לבחור תאים מתאימים בנפרד באמצעות דוגם תאים חצי אוטומטי.
זה כרגע נשאר לא ידוע אם צורת CM, בפני עצמו, יש השפעה פנים-פונקציונלית על סינכרון שריר הלב. המטרה העיקרית של השיטות המוצעות בעיתון זה הייתה לפתח פלטפורמה חדשנית כדי ללמוד אם צורת התא לעצמה הייתה השפעה על transcriptome17. למרות שמחקרים במבחנה שונים מחקרי vivo, מטרת מחקר זה היה לחקור את ההשפעה של צורות תאים שונים על ביטוי גנים, בהתחשב בכך שהשווא תאים עם צורות שונות ב vivo הוא תובעני מאוד. ניסויים אלה נוצרו בהשראת Kuo et al.19, שהשתמש בגישה דומה ודיווח כי הם הבחינו בשינויים בפרמטרים פיזיולוגיים עקב שינויים בצורת התא.
Protocol
כל ההליכים הנוגעים לבעלי חיים היו בהתאם לתקנות ועדת האתיקה של בעלי החיים של מכון קרולינסקה, שטוקהולם, שוודיה.
1. פריסת שבב בדוגמת מיקרו
- השתמש שבב מותאם אישית(טבלת חומרים)(איור 1A): 19.5 מ"מ x 19.5 מ"מ כיסוי עם micropatterns מופעל, מודפס על ידי פוטוליתוגרפיה על זכוכית borosilicate.
הערה: מיקרו-פטרנים אלה מוקפים באזור ציטופובי. לכן, תא זריעה יכול רק לצרף ולגדול על אחד אלה micropatterns וללכוד את AR של micropattern זה. השבב מחולק לשלושה אזורים וכל אזור מורכב מיקרו-פטרנים עם AR ספציפי. פריסת השבב מוצגת באיון 1B. הגיאומטריה של קוביות ה- ARs המוגדרות מוצגת בטבלה 1. תמונות פלורסנט מוגדלות של הצורות השונות של מיקרו-פטרנים פיברונקטין מוצגות בתמונות התחתיות באיור 1B.
2. ציפוי שבבים מיקרו-פטרן
- להכין 2x ציפוי חלבון תמיסה עבור כל שבב על ידי הוספת 80 μg של פיברונקטין 2 מ"ל של תמיסת מלח פוספט אגירה (PBS-/-).
- להעביר שבב לצלחת 35 מ"מ Greiner פטרי ומיד להוסיף 2 מ"ל של PBS-/-. לאחר מכן הוסף 2 מ"ל של תמיסת חלבון ציפוי 2x.
- דגירה את השבבים בטמפרטורת החדר במשך 2 שעות.
- לשטוף את פתרון הציפוי על ידי שלבי דילול רצופים עם PBS-/-. משטח השבב תמיד צריך להיות רטוב. לאחר מכן, החלף את PBS עם 2 מ"ל של ציפוי בינוני דגירה ב 37 מעלות צלזיוס עד תאי זריעה.
3. בידוד של הודעות
- להכין את מדיום ציפוי על ידי השלמת DMEM:M199 (4:1) עם 10% סרום סוס, 4% סרום בשר עוברי, 2% HEPES (1 M) ו 1% פניצילין / סטרפטומיצין (10,000 U/mL)22.
- לנתח את הרקמה מהבר השמאלי של לבבות חולדה יילודים בן יומיים ולהעביר לצלחת 10 ס"מ המכיל PBS. חותכים את הרקמה לכ-1 מ"מ3 חתיכות.
- מעבירים את הרקמה שנקטפו לתוך צינור רוטור-כובע, מצויד רוטור בכובע לניתיקת רקמות(טבלת חומרים). תן לרקמות להתיישב ואז בזהירות להסיר את supernatant.
- מוסיפים 2.5 מ"ל לתערובת תערובת האנזימים 1 ו-2, מוכנים באמצעות ערכת דיסוציאציה של הלב של יילודים(Table of Materials),לצינור C וסוגרים את המכסה בחוזקה.
- הכנס את צינור מכסה הרוטור על שרוול הדיסוציאטור, מצויד בתנורים(איור 2A ו- B)(טבלת חומרים). הפעל את תוכנית הדגירה 37C_mr_NHDK_1 (איור 2C), להימשך כשעה.
- בזמן שתכנית הדגירה פועלת, הכינו מאגר PEB המכיל 2 mM EDTA ו- 0.5% אלבומין סרום של סרום קובין (BSA) ב- PBS, pH 7.2, ולשמור אותו ב- 4 °C.
- לאחר סיום תוכנית הדגירה, נתקו את צינור הרוטור-קאפ(איור 2D) והוספו7.5 מ"ל של מדיום ציפוי חם מראש.
- תוסתה מחדש את הדגימה וסנן את מתלי התא באמצעות מסננת של 70 μm.
- שוטפים את מסננת עם עוד 3 מ"ל של ציפוי בינוני.
- צנטריפוגה מתלה התא ב 600 x g עבור 5 דקות. לשאוף את הסופרנטנט לחלוטין.
- תולה מחדש את כדור התא ב 60 μL של מאגר PEB קר.
- הוסף 20 μL של קוקטייל בידוד Cardiomyocyte יילודים(טבלת חומרים),המכיל חרוזים בגודל מיקרון כי היעד שאינם CMs.
- להוסיף 20 μL של חרוזי תאי דם אנטי אדום(טבלת חומרים).
- מערבבים את המתלים ומדגירה ב-4°C למשך 15 דקות.
- הוסף 400 μL של מאגר PEB.
- החל את מתלה התא על עמודת LD (טבלת חומרים), אשר הוכנס אנכית לתוך מעמד מגנט ונשטף היטב עם מאגר PEB.
- אסוף תאים ללא תווית ועמודת כביסה עם מאגר PEB של 0.5 מ"ל.
- הוסף 8 מ"ל של ציפוי בינוני והעבר את מתלי התא לבקבוק תרבות 75 ס"מ2 מצופים דגירה ב 37 °C עבור 1.5 שעות. שאר הלא-CMs יתחילו לדבוק בתרבות התאים הלא מנוססים והמתלה התא יועשר על ידי אוכלוסיית CM.
4. תבניות CMs
הערה: השווינו CMs יחיד עם ARs של 1:1, 7:1 או 11:1. הדבר נעשה על ידי זריעת CMs חולדת יילודים מבודד על שבב שתוכנן במיוחד מלא micropatterns מצופה פיברונקטין עם ARs מוגדר של 1:1, 7:1 או 11:1. המיקרו-פטרנים היו מצופים בפיברונקטין, מוקפים במשטח ציטופובי. לכן, CMs יהיה לצרף, להפיץ וללכוד את AR המוגדר של micropatterns על ידי גידול אך ורק על הצוללת פיברונקטין. תבנית ם CMs מבודד בהתאם ל השלבים הבאים.
- להעביר את מתלי התא לצינור 15 מ"ל, לספור את התאים ולדלל לריכוז של 100,000 תאים לכל מ"ל על ידי הוספת מדיום ציפוי מתאים.
- להוסיף 2 מ"ל של מתלה תא על השבב, אשר כבר שקע 2 מ"ל של ציפוי חם בינוני בתוך צלחת 35 מ"מ Greiner פטרי.
- דגירה את המנה ב 37 °C עם 5% CO2 כדי לאפשר CMs לצרף micropatterns מצופה פיברונקטין ולאפשר לכל CM לרכוש את AR של micropattern השקוע שלה.
- אחרי 18 שעות, בדוק את השבב. אם רוב התאים צירף, נתק והסר את הפסולת ואת התאים המתים המחוברים לתא עם התבנית. עשה זאת על-ידי הסרת מדיום הציפוי והוספת PBSבעדינות -/- dropwise, החל ממרכז השבב ולאחר מכן נע לכיוון הצדדים. חזור על הפעולה פעמיים.
- לשאוף PBS עם תחזוקה טרי בינוני על ידי השלמת DMEM:M199 (4:1) עם 4% סרום סוס, 4% סרום בשר עוברי, 2% HEPES (1 M) ו 1% פניצילין / סטרפטומיצין (10,000 U/mL).
5. בחירת CMs חסיד
הערה: לאחר תקופת איסוף של 72 שעות, CMs יחיד בדוגמת נאספים ממיקרופטרים פיברונקטין שלהם באמצעות קוגם תא חצי אוטומטי(טבלת חומרים) (איור 3). דוגם התא משתמש בתוכנה23 כדי לשלוט בשלב הממונע(איור 3A). מיקרו-כיליון זכוכית של 70 μm (איור 3B) משמש כדי לאסוף ולהזריק את ה-CMs של חולדת יילודים בדוגמת. דוגם התאים ממיין תאים דבקים על ידי יצירת ואקום והזרקת התאים על ידי הפעלת לחץ. הוואקום במזרק מספר 1 מוחל על ידי משיכת המזרק באמצעות משאבת המזרק(איור 3C). הלחץ ההידרוסטטי מבוסס על כוח המשיכה ומושרה על ידי הצבת מזרק מספר 2 במרחק של 87 ס"מ מעל שולחן המיקרוסקופ. מזרקים 1 ו- 2 מחוברים בהתאמה באמצעות צינורות PTFE, לשסתומים 1 ו- 2, המוטבעים ביחידתהבקרה (איור 3D). צינורות PTFE מלאים לחלוטין במים ללא RNase. התא היחיד שנבחר מוזרק לאחר מכן לצינור תגובת שרשרת פולימראז (PCR), המכיל 3.55 μL של מאגר lysis.
- לאחר תקופת תפירה של 72 שעות, הסר את המדיום הישן ושטוף בעדינות את משטח השבב עם DPBSחם -/-.
- שומרים על המנה שטוחה ושואפים בעדינות את המדיום הישן עם פיפטה של 1000 μL מצד אחד של המנה. ודא כי השבב נשאר רטוב בכל עת.
- הוסף 2 מ"ל של DPBS-/- בעדינות ובנקודת טיפה לנתק את רוב התאים המתים המחוברים לתאים בדוגמת, החל ממרכז השבב ולאחר מכן נע לצדדים שלה.
- לשאוף את רוב DPBS כדי להסיר תאים צפים מנותקים רבים ככל האפשר, החל ממרכז השבב ולאחר מכן נע לצדדים.
- חזור על שלבים 5.1.2 ו- 5.1.3 פעם נוספת.
- השתמש מקצות זווית כדי לתפוס את קצה השבב ולהעביר אותו מיד לצלחת חדשה סטרילית 35 מ"מ Greiner Petri כדי להפחית את מספר התאים הצפים בעת איסוף.
- מיד להוסיף 1.5 מ"ל של DPBS-/- כך השבב לא יתייבש.
- להוסיף 1.5 μL של מחזור צבע תוסס ירוק כדי לדמיין את הגרעין של התאים החיים.
- מניחים את השבב במרכז צלחת גריינר פטרי באמצעות קצה המפלצות.
- מניחים תאמעלהשבב. התא יתקן את השבב לתחתית הצלחת, מבלי לחסום את הגישה לדפוסי התא.
- הר את צלחת Greiner Petri על מחזיק המנה של שלב קוטף התא ולהכניס את הכובע המגנטי.
- כיול הזריקה האוטומטית.
- אתר את הכוונת, החרוטה על הבמה הממונעת באמצע התמונה בחלון Live View.
- התמקד על הכוונת ובחר בלחצן כיול להזרקה אוטומטית בחלון הסריקה והמיון.
- החלף אתה-DPBS -/- ב- 1.5 מ"ל של DPBS/trypsin-/- (1:1) בזמן שהמנה על הבמה, כדי לשחרר את התאים מהפירונקטין כך שניתן יהיה להשתמש בוואקום נוזלי כדי לבחור את התאים.
- סרוק את השבב כולו באמצעות הכרטיסיה סריקה בחלון סריקה ומיון. אתר את הפינה השמאלית העליונה של השבב בשדה התצוגה ולחץ על קבל את מיקום המיקרוסקופ הנוכחי בשורה השמאלית העליונה בפינה השמאלית.
- לאחר מכן, הזז את השלב הממונע לפינה הימנית התחתונה של השבב. למקד את המיקרוסקופ ולחץ על קבל מיקום מיקרוסקופ הנוכחי בשורה בפינה הימנית התחתונה.
- לחץ על לחצן הגדר את המטוס החד ביותר ובחלון הצץ לחץ על מעבר לפינה השמאלית העליונה. למקד את המיקרוסקופ ולחץ על עבור לפינה השמאלית התחתונה ולהגדיר את המוקד. לאחר שתסיים, לחץ על לחצן סיום והתחיל בסריקה.
- לאחר סיום הסריקה, עבור אל הכרטיסיה ניתוח ובחר את התאים הבודדים העוברים את קריטריוני המחקר.
- ודא כי מיקרוסקופ הזכוכית הוא באמצע התצוגה החיה של המיקרוסקופ.
- שלוט במשאבת המזרק באמצעות חלון המשאבה של התוכנה. צור ואקום על ידי משיכת 4 מ"ל ממזרק מספר 50 מ"ל 1, אשר בקוטר של 27 מ"מ.
- בכרטיסיה מיון
- הגדר את פרמטרי ההזרקה של השסתומים. זה היה מחושב כי נפח ההזרקה שנמסר תא יחיד הרים היה 1 μl, אם שסתום 2 נפתח עבור 120 אלפיות השנייה ולאחר מכן שסתום 1 נפתח עבור 20 היה אלפיות השנייה לאחר מעידת זמן של 200 אלפיות השנייה. בשל האלסטיות של הצינורות, שסתום פתוח 1 כדי להפסיק להזריק את הזרימה.
- הגדר את פרמטרי האיסוף של השסתומים. הוא חושב שאם שסתום 1 נפתח ל-20 אלפיות השנייה ואז שסתום 2 נפתח למשך 10 אלפיות השנייה, לאחר שגת זמן של 10 אלפיות השנייה, ניתן לאסוף את רוב התאים עם הדוגמאות. הסיבה לכך היא כריכות פיברונקטין שלהם התרופפו לאחר שטופלו על ידי טריפסין.
- לחץ על לחצן חשב את הנתיב. התוכנה מחשבת את הנתיב המהיר ביותר מתא לתא, כדי להרים ולהזריק את התאים הנבחרים לאורך השבב.
- למקד את המיקרוסקופ על תא בדוגמת על פני השטח שבב.
- באמצעות הג'ויסטיק, הזז את המיקרו-קבילארי למטה בזהירות, כך שניתן יהיה להשיג את התמונה החדה ביותר של קצה המיקרו-כיליון מבלי לגעת בתא.
- לחץ על לחצן הגדר במקטע היסט מיקרו-פיפטה. חלון חדש צץ, מראה את חתך הרוחב של המיקרו-החלון. לחץ על המרכז המדויק של נערך. לאחר מכן התוכנה תקליט את היסט הקצה של נערך בנקודות הציון x, y ו- z.
- הפעל מיון באמצעות לחצן התחל מיון.
תוצאות
הרקמה נותקה מהחלק השמאלי של לבבות חולדות יילודים בני יומיים וחולקה לתאים בודדים. לאחר מכן ה-CMs המועשר נזרעו על שבב המכיל דפוסי פיברונקטין עם ARs ברורים. לאחר 72 שעות של culturing, המדיום הוחלף על ידי 1:1000 מחזור צבע תוסס ירוק DPBS-/- במשך 2 דקות כדי לדמיין את הגרעין של התאים החיים. לאחר מכן, התאים טופלו DPBS-/-/ trypsin (1:1) כדי לשחרר את התאים מן fibronectin, כך ואקום נוזלי יכול לשמש כדי להקל על איסוף תאים. בינתיים, השבב כולו נסרק בהגדלה של פי 10, באמצעות מיקרוסקופ הפוך המחובר לקוטף התאים. זה בוצע לפני התאים הפכו מעוגל עקב טיפול נסהפסין. התאים המתאימים נבחרו, בהתבסס על התמונה הסרוקה, וקואורדינטותיהם נשמרו בתוכנה של קוטף התאים. המיקרו-פטרנים נבחרו רק אם הם הכילו תא יחיד חד-נו-קוטבי ורק כאשר התא כיסה באופן מלא את המיקרו-פטרן של פיברונקטין. סדרן התאים הרים את התאים שנבחרו אחד אחד וכל תא יחיד שנבחר בהצלחה הוזרק מיד לתוך צינור PCR בודד והוצב על שלב המיקרוסקופ. כל שפופרת PCR הכילה 3.55 μL של מאגר lysis(טבלה 2). תהליך המיון, שהחל בהסרת המדיה, הושלם תוך 40 דקות. הסינתזה של חומצה deoxyribonucleic משלימה (cDNA), PCR לפני ההגברה והטיהור בוצעו על התאים הבודדים, בהתבסס על פרוטוקול Smart-Seq224 (טבלה 3). איכות ה-CDNA המטוהר נבדקה על ידי מנתח אלקטרופורזה אוטומטי. האלקטרופרוגרמה של ה- CDNA מוגבר מראש של תא יחיד אחד שנבחר מוצג באיור 4. ספריות RNA-Seq הוכנו על פי פרוטוקול Smart-Seq224.
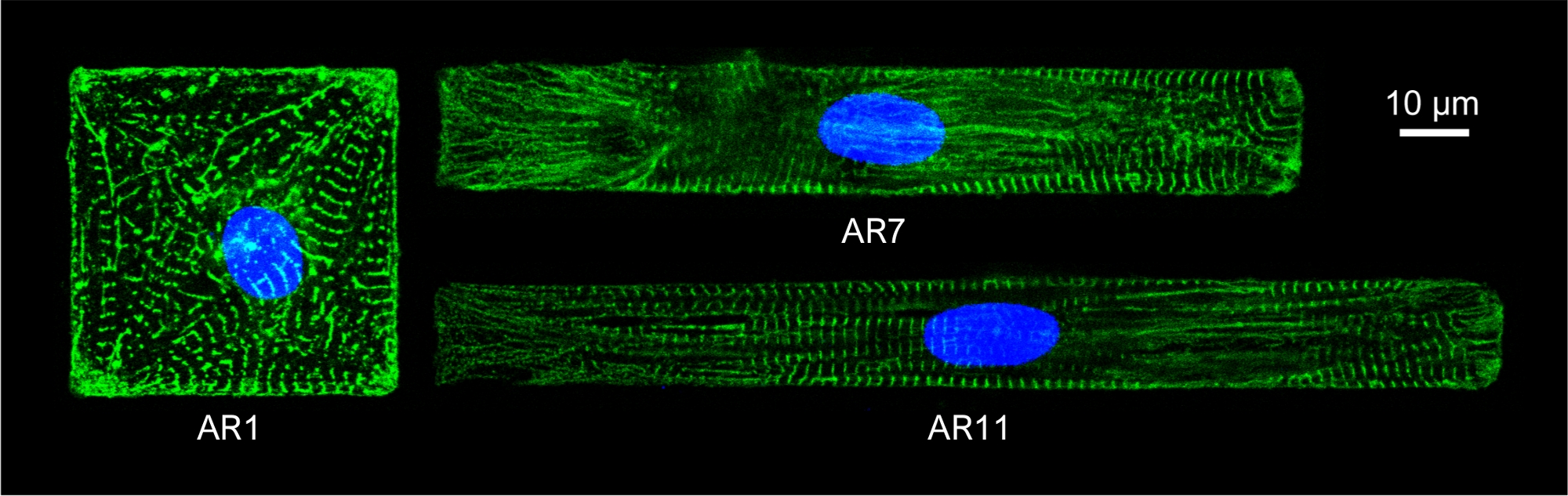
כדי לצפות במבנה הסרקומיר של ה-CMs בדוגמת, CMs בדוגמת היו מוכתמים α-actinin סרקומרי. התאים היו דגירה עם חמור נגד עכבר IgG אלקסה פלור 488 1:800 במשך שעה אחת בטמפרטורת החדר עבור הכתמים המשניים. גרעין היו מוכתם 1 μg / מ"ל DAPI. תמונות אימונופלורסנט נרכשו עם מיקרוסקופ קונפוקאלי הפוך, באמצעות 63x שמן טבילה (NA 1.4) יעד(איור 5).

איור 1: פריסת השבב עם מיקרו-פטרנים פיברונקטין.
(א)תמונה של השבב שתוכנן בהתאמה אישית. השבב הוא 19.5 ממ x 19.5 ממ כיסוי עם מיקרופטרן פיברונקטין, מודפס על ידי פוטוליתוגרפיה על borosilicate. פריסתשבב. השבב מחולק לשלושה אזורים וכל אזור מורכב מיקרופטרים פיברונקטין עם תמונות ספציפיות AR. פלורסנט של צורות שונות של מיקרופטרן פיברונקטין מוצגים בתצוגה מוגדלת עבור כל אזור. נתון זה שונה מ "חומר משלים 1" על ידי Haftbaradaran Esfahani ואח',בשימוש תחת http://creativecommons.org/licenses/by/4.0/ . לחץ כאן כדי להציג גירסה גדולה יותר של נתון זה.
{kind=link}

איור 2: דיסוציאטור מצויד בלחממים.
(א)כל מכשיר הדיסוציאטור המשמש לדיסוציאציה אוטומטית מלאה של חולדה ילודים בת יומיים בחדר השמאלי. יחידתחימום. שפופרתכובע רוטור. (D)תוכניות מוכנות לשימוש עבור זרימת עבודה אוטומטית לחלוטין של ניתקת רקמות. לחץ כאן כדי להציג גירסה גדולה יותר של נתון זה.
{kind=link}

איור 3: אפליקציית קוטף תאים.
(א)תצוגה מוגדלת של השלב הממונע של דוגם התא. מחזיק צלחת פטרי ו-80 חורים ל-10 רצועות PCR וחור לכיול על הכוונת מוטבע על הבמה. (ב)תצוגה מוגדלת של מיקרו-כיון זכוכית. משאבתהמזרק. (ד)יחידת הבקרה, השולטת בחלון זמן הפתיחה והסגירה של שסתומים 1 ו- 2, מותקנת בתוך יחידת הבקרה. לחץ כאן כדי להציג גירסה גדולה יותר של נתון זה.
{kind=link}

איור 4: האלקטרופרוגרמה של ה-cDNA מוגבר מראש של תא יחיד אחד שנבחר.
19 מחזורי PCR של טרום-ההגדלה שימש כדי להשיג 15 μL של 1 ng/μL מטוהר תפוקת cDNA. רצועה ברורה בעלילה צפיפות כמו ג'ל נצפתה אשר מתאים לשיא ב 1852 bp באלקטרופרוגרם. הגודל הממוצע של שברים הוא 1588 bp. יתר על כן, הכמות הקטנה של שברים קצרים מ 300 bp מציין ספריית cDNA טובה. לחץ כאן כדי להציג גירסה גדולה יותר של נתון זה.
{kind=link}

איור 5: כתם אימונופלורסנט של α-אקסטין (ירוק) וגרעין (כחול) של CMs בדוגמת עם ARs שונים.
הכרומטין הוכתם על ידי DAPI. לחץ כאן כדי להציג גירסה גדולה יותר של נתון זה.
{kind=link}
| מורפוטיפ (סוג של מורפוטיפ) | Ar | אורך (μm) | רוחב (μm) | אזור פיברונקטין (μm2) |
| AR1 (1) | 1:1 | 47 | 47 | 2209 |
| AR7 (100) | 7:1 | 126 | 18 | 2268 |
| אר-11 | 11:1 | 155 | 14 | 2170 |
טבלה 1: גיאומטריה של CMs בדוגמת.
| רכיב | נפח (μL) |
| מים ללא גרעין | 0.65 |
| (0.4% כרך/כ) טריטון X-100 | 1.8 |
| תערובת dNTP (25 mM) | 0.8 |
| מעכב RNase (40 U μL-1) | 0.1 |
| אוליגו-dT30VN אוליגונוקלאוטידים (100 μM) | 0.1 |
| תערובת ספייק-אין ERCC RNA (2.5 x 105 דילול) | 0.1 |
| הוזרק תא יחיד | 1 |
| נפח כולל | 4.55 |
טבלה 2: מאגר התאמה אישית של תא יחיד.
| רכיב | נפח (μL) |
| מאגר גדיל ראשון כתב עילי II (5x) | 2 |
| DTT (100 mM) | 0.5 |
| בטיין (5 מ') | 2 |
| Mgcl2 (1 M) | 0.1 |
| מעכבי RNase (40 U μL-1) | 0.25 |
| כתב עילי II תמלול הפוך (200 U μL-1) | 0.5 |
| TSO (100 μM) | 0.1 |
| נפח כולל | 5.45 |
טבלה 3: תמלול הפוך (RT) לערבב עבור תגובה אחת RT לסנתז cDNA גדיל ראשון מן lysate של ס"מ יחיד.
Discussion
מחקר זה השתמש רצף RNA חד-תאי, שהוא חדשני וטכנולוגיה רבת עוצמה שיכול לזהות את התמליל של תאים בודדים. זה היה בשילוב עם גישה חדשנית לculturing CMs יחיד, כך שהם לקחו על ARs שונים, כי, אחרת, היה יכול רק נצפתה vivo.
למחקר היו כמה מגבלות. לדוגמה, היו צריכים להשתמש ב-CMs יילודים כדי ליצור מורפוטיפים שונים, מכיוון שהיא מאתגרת במיוחד לתרבות מספיק סמ"מים בוגרים חיוניים למשך 72 שעות בצורות מוגדרות. יתר על כן, CMs היו תרבותיים במשך 72 שעות לשעבר vivo, אשר אולי הייתה השפעה על דפוס ביטוי הגן. עם זאת, culturing זה היה הכרחי, כך התאים יכולים ליצור מורפוטיפים ספציפיים. יתר על כן, רק תאים בודדים שהיו חד-נוכיים וכיסו באופן מלא את המיקרופטרן פיברונקטין נבחרו למיון. יש למיין תאים עם תבנית בכל שבב רק בסיבוב אחד של מיון. לבסוף, כ-50 תאים שהוא בערך שליש מהתאים הנבחרים נאספו בהצלחה מכל שבב. קיימות שתי סיבות שהגבילו את מספר התאים שנבחרו בהצלחה. ראשית, תאים מסוימים היו מחוברים בחוזקה מדי לתבנית פיברונקטין וזרימת האיסוף לא הייתה בכפייה מספיק כדי לאסוף אותם בהצלחה. שנית, בשל הטיפול ב trypsin, החיבור בין תאים מסוימים פיברונקטין הפך רופף מדי. כתוצאה מכך, תאים אלה נדחקו הרחק מהמיקרו-פטרונים שלהם, כאשר המיקרו-כיף התקרב אליהם, והם לא נאספו. המחברים אינם טוענים כי התקנה זו זהה לסביבת vivo, אבל זה הוכיח להיות גישה בת קיימא לענות על שאלת המחקר.
השיטה המוצעת ישימה בסוגי תאים שונים (לדוגמה, עבור hiPS-CMs). עם זאת, יש למטב את הגורמים הבאים כדי ללמוד סוגי תאים אחרים. מולקולות דבק ECM מתאימות לחיבור של סוג התא הספציפי יש להשתמש לציפוי micropatterns. הגיאומטריה של micropatterns יש לשנות על פי שאלת המחקר וסוג התא. ניתן לשנות את תקופת ההתעברות בהתבסס על שאלת המחקר. יש לייעל את ריאגנט הנתק ואת זמן הדגירה שלו בדיוק עבור סוג תא המחקר. לדוגמה, Accutase יכול לשמש במקום TryplE עבור ניתוק של תאי גזע עובריים ונוירוניים. יש לבחון את פרמטרי זמן הפתיחה של השסתומים כדי לבחור תאים בהצלחה, אך בעדינות. לסיכום, הנדסנו פלטפורמה חדשנית לחקר צורת התא שיכולה לספק משאב בעל ערך לחוקרים בתחום. בהקשר זה, עיצבנו גישה ניסיונית שמחקה במבחנה צורות אופייניות שנכפו על CM ב vivo על ידי אילוצים המודינמיים כדי לזהות את יחסי הגומלין בין אדריכלות תאית וביטוי גנים. אנו גם מדווחים על פיתוח פלטפורמה חדשנית ללמוד HF במבחנה וזיהוי של צורת התא כקביעה רבת עוצמה של ביטוי גנים. זוהי התבוננות חדשנית עם השלכות מרחיקות לכת על ביולוגיה ורפואה.
Disclosures
ללא.
Acknowledgements
ללא.
Materials
| Name | Company | Catalog Number | Comments |
| 2100 Bioanalyzer Instrument | Agilent Technologies | G2939BA | Automated electrophoresis analyzer |
| Anti-Red Blood Cell MicroBeads | Miltenyi Biotec | 130-109-681 | |
| Axio Observer microscope | Zeiss | Z1 | Inverted microscope |
| Betaine solution (5 M) | Sigma-Aldrich, MERCK | B0300 | |
| CellSorter | CELLSORTER | https://www.singlecellpicker.com/ | Cell picker |
| CYTOOchamber | CYTOO | 30-010 | Custom-designed chip |
| CYTOOchip | CYTOO | 10-950-00-18 | Chamber |
| DMEM | Thermo Fisher Scientific | 31966-021 | high glucose, GlutaMAX Supplement |
| dNTP mix (25 mM) | Thermo Fisher Scientific | R1122 | |
| Donkey Anti-Mouse IgG Alexa Fluor 488 | Abcam PLC | ab150105 | |
| DTT (100 mM) | Thermo Fisher Scientific | 18064071 | |
| ERCC RNA Spike-In Mix | Thermo Fisher Scientific | 4456740 | |
| Fetal Bovine Serum | Thermo Fisher Scientific | 10082-147 | |
| Fibronectin | Sigma-Aldrich, MERCK | F4759 | |
| gentleMACS C Tube | Miltenyi Biotec | 130-093-237 | Rotor-cap tube |
| gentleMACS Octo Dissociator with Heaters | Miltenyi Biotec | 130-096-427 | Dissociator with heater |
| Greiner CELLSTAR Petri dish | Sigma-Aldrich, MERCK | P6987 | |
| HEPES (1 M) | Thermo Fisher Scientific | 15630-056 | |
| Horse Serum | Sigma-Aldrich, MERCK | H0146 | |
| LD column | Miltenyi Biotec | 130-042-901 | |
| Medium 199 | Thermo Fisher Scientific | 31150-022 | |
| NE-1000 syringe pump | New Era Pump Systems | NE-1000 | Syringe pump |
| Neonatal Cardiomyocyte Isolation Cocktail, rat | Miltenyi Biotec | 130-105-420 | |
| Neonatal Heart Dissociation Kit, mouse and rat | Miltenyi Biotec | 130-098-373 | |
| Oligo-dT30VN oligonucleotides | IDT Technology | 5′–AAGCAGTGGTATCAACGCAGAGTACT30VN-3′ | |
| RNAse inhibitor (40 U µL-1) | Clontech | 2313A | |
| sarcomeric α-actinin | Sigma-Aldrich, MERCK | EA-53 | |
| SP8 confocal microscope | Leica Microsystems | SP8 | Confocal microscope |
| Superscript II first-strand buffer (5x) | Thermo Fisher Scientific | 18064071 | |
| Superscript II reverse transcriptase (200 U µL-1) | Thermo Fisher Scientific | 18064071 | |
| Triton X-100 | Sigma-Aldrich, MERCK | T9284 | |
| TryplE Express enzyme, no phenol red | Thermo Fisher Scientific | 12604013 | |
| TSO (100 µM) | QIAGEN | 5′-AAGCAGTGGTATCAACGCAGAGTACATrGrG+G-3′ | |
| Vibrant Dye Cycle green | Thermo Fisher Scientific | V35004 |
References
- Heineke, J., Molkentin, J. D. Regulation of cardiac hypertrophy by intracellular signalling pathways. Nature Reviews Molecular Cell Biology. 7 (8), 589-600 (2006).
- Haftbaradaran Esfahani, P., et al. Cell shape determines gene expression: cardiomyocyte morphotypic transcriptomes. Basic Research in Cardiology. 115 (1), 7(2019).
- Kontrogianni-Konstantopoulos, A., Benian, G., Granzier, H. Advances in Muscle Physiology and Pathophysiology 2011. Journal of Biomedicine and Biotechnology. 2012, 930836(2012).
- Hill, J. A., Olson, E. N. Cardiac plasticity. New England Journal of Medicine. 358 (13), 1370-1380 (2008).
- Bray, M. A., Sheehy, S. P., Parker, K. K. Sarcomere alignment is regulated by myocyte shape. Cell Motility and the Cytoskeleton. 65 (8), 641-651 (2008).
- Benjamin, I. J., Schneider, M. D. Learning from failure: congestive heart failure in the postgenomic age. Journal of Clinical Investigation. 115 (3), 495-499 (2005).
- Opie, L. H., Commerford, P. J., Gersh, B. J., Pfeffer, M. A. Controversies in ventricular remodelling. Lancet. 367 (9507), 356-367 (2006).
- Braunwald, E. Cardiomyopathies: An Overview. Circulation Research. 121 (7), 711-721 (2017).
- Knoll, R., et al. The cardiac mechanical stretch sensor machinery involves a Z disc complex that is defective in a subset of human dilated cardiomyopathy. Cell. 111 (7), 943-955 (2002).
- Frangogiannis, N. G. The Extracellular Matrix in Ischemic and Nonischemic Heart Failure. Circulation Research. 125 (1), 117-146 (2019).
- Marian, A. J., Braunwald, E. Hypertrophic Cardiomyopathy: Genetics, Pathogenesis, Clinical Manifestations, Diagnosis, and Therapy. Circulation Research. 121 (7), 749-770 (2017).
- Mozaffarian, D., Caldwell, J. H. Right ventricular involvement in hypertrophic cardiomyopathy: a case report and literature review. Clinical Cardiology. 24 (1), 2-8 (2001).
- Olsson, M. C., Palmer, B. M., Stauffer, B. L., Leinwand, L. A., Moore, R. L. Morphological and functional alterations in ventricular myocytes from male transgenic mice with hypertrophic cardiomyopathy. Circulation Research. 94 (2), 201-207 (2004).
- Toepfer, C. N., et al. Hypertrophic cardiomyopathy mutations in MYBPC3 dysregulate myosin. Science Translational Medicine. 11 (476), (2019).
- McKenna, W. J., Maron, B. J., Thiene, G. Classification, Epidemiology, and Global Burden of Cardiomyopathies. Circulation Research. 121 (7), 722-730 (2017).
- Schultheiss, H. P., et al. Dilated cardiomyopathy. Nature Reviews Disease Primers. 5 (1), 32(2019).
- Knoll, R. A role for membrane shape and information processing in cardiac physiology. Pflugers Arch. 467 (1), 167-173 (2015).
- Rangamani, P., et al. Decoding information in cell shape. Cell. 154 (6), 1356-1369 (2013).
- Kuo, P. L., et al. Myocyte shape regulates lateral registry of sarcomeres and contractility. American Journal of Pathology. 181 (6), 2030-2037 (2012).
- Gerdes, A. M., Onodera, T., Wang, X., McCune, S. A. Myocyte remodeling during the progression to failure in rats with hypertension. Hypertension. 28 (4), 609-614 (1996).
- Kehat, I., et al. Extracellular signal-regulated kinases 1 and 2 regulate the balance between eccentric and concentric cardiac growth. Circulation Research. 108 (2), 176-183 (2011).
- Sreejit, P., Kumar, S., Verma, R. S. An improved protocol for primary culture of cardiomyocyte from neonatal mice. In Vitro Cellular & Developmental Biology - Animal. 44 (3-4), 45-50 (2008).
- Kornyei, Z., et al. Cell sorting in a Petri dish controlled by computer vision. Scientific Reports. 3, 1088(2013).
- Picelli, S., et al. Full-length RNA-seq from single cells using Smart-seq2. Nature Protocols. 9 (1), 171-181 (2014).
- Schmick, M., Bastiaens, P. I. H. The interdependence of membrane shape and cellular signal processing. Cell. 156 (6), 1132-1138 (2014).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved