
Method Article
ŞekilBağımlı Transkripsiyonları Tek Hücre Düzeyinde Çalışma Yaklaşımı
Bu Makalede
Özet
Bu makalede, farklı patolojileri temsil eden farklı şekillerde kardiyak miyositlerin yetiştirilmesi ve morfolojilerine göre bu yapışık kardiyak miyositlerin tek hücre düzeyinde sıralanması için yöntemler yer alıyor. Önerilen platform kalp yetmezliği farklı türleri için yüksek iş ve ilaç tarama için yeni bir yaklaşım sağlar.
Özet
Farklı tipkardiyak hipertrofi, artmış kardiyak miyosit hacmi (CM) ile ve CM morfolojisindeki değişikliklerle ilişkilendirilmiştir. Hücre hacminin gen ekspresyonu üzerindeki etkileri iyi bilinmekle birlikte, hücre şeklinin etkileri iyi anlaşılamamıştır. Bu makalede, CM morfolojisinin gen ekspresyonu üzerindeki etkilerini sistematik olarak analiz etmek üzere tasarlanmış bir yöntem açıklanmaktadır. Daha sonra tek hücreli mRNA sıralama takip yeni bir tek hücreli yakalama stratejisinin geliştirilmesi ayrıntıları. Mikro desenli çip de 3000 dikdörtgen şekilli fibronektin mikro desenleriçeren tasarlanmıştır. Bu, farklı kalp yetmezliği türlerine (HF) karşılık gelen genişlik en boy oranları (AR) şeklinde CM'lerin büyümesini mümkün kılar. Kağıt da yarı otomatik mikro-pipet hücre toplayıcı kullanarak, kendi desen tek hücreleri almak ve ayrı bir lysis tampon içine enjekte etmek için tasarlanmış bir protokol açıklar. Bu, tanımlanmış geometrik morfotiplerle tek CM'lerin transkripsiyonlarının profilini çıkarmak ve bunları normal veya patolojik koşullara göre karakterize etmeyi mümkün kılmıştır: hipertrofik kardiyomiyopati (HCM) veya sonradan yük/konsantrik karşı dilate kardiyomiyopati (DCM) veya preload/eksantrik. Özetle, bu makalede, farklı patolojileri temsil eden farklı şekillerde CM'ler yetiştirmek ve bu yapışkan CM'leri morfolojilerine göre tek hücreli düzeyde sıralamak için yöntemler sunmaktadır. Önerilen platform HF farklı türleri için yüksek iş ve ilaç tarama için yeni bir yaklaşım sağlar.
Giriş
Dünya Sağlık Örgütü'ne göre, kardiyovasküler hastalık (KVD) dünya çapında morbidite ve mortalitenin önemli bir nedenidir. CVD önemli ölçüde insanların yaşam kalitesini etkiler ve büyük bir sosyoekonomik etkiye sahiptir. HCM ve DCM gibi kardiyomiyopatiler kalp kasının primer bozukluklarıdır ve HF'nin başlıca nedenleri yüksek morbidite ve mortalite ile ilişkilidir. Enfeksiyonlar ve toksinler veya bazı ilaçlara maruz kalma gibi çevresel etkileri de dahil olmak üzere HF birçok nedeni vardır8. HF de genetik yatkınlık neden olabilir, yani mutasyonlar9. Bu ekstrasellüler matriks etkileyen genetik bileşiminde değişiklikler (ECM) molekülleri, integrins veya sitaiskelet proteinleri bozulmuş mekanosensation ve kardiyak hastalığın çeşitli sorumlu olabileceğine inanılmaktadır10.
HCM ana özelliği sol ventrikül açıklanamayan hipertrofi olduğunu11, ve bazen sağ ventrikül12, ve bu sık interventriküler septum baskın tutulum ile ortaya çıkar. HCM ayrıca diyastolik disfonksiyon ve miyosit düzensizliği ve fibrozis13ile karakterizedir. Çoğu durumda, kalbin kontraktil cihaz sarkoerik proteinlerde mutasyonlar etkilenir, miyositlerin kontrility yol14. Buna karşılık, DCM bir dilatasyon ile karakterizedir, veya her ikisi, ventriküller ve olguların% 30 ila% 50 ailesel etiyolojisi vardır15. DCM hücresel fonksiyonlarıgeniş bir yelpazede etkiler, miyositlerin bozulmuş kasılması yol açan, hücre ölümü ve fibrotik onarım16.
Genetik mutasyonların belirli türleri hcm3sırasında belirli şekil özellikleri benimsemeye tek CMs zorlamak göstermiştir , yani bir uzunluk ile kare şekilli hücreler: neredeyse eşit genişlik AR 1:14 (AR1). Aynı dcm için de geçerlidir, neredeyse 11:1 (AR11) eşit bir AR ile uzatılmış hücreleri ile. Buna ek olarak, HF artan afterload neden olabilir (örneğin, hipertansiyon). Bu gibi durumlarda, hemodinamik talepler CMs kare şekiller almaya zorlamak, Laplace yasasına göre, ve AR değişiklikleri 7:15 (AR7) için 1:16,7. HF ayrıca ön yükteki artışa da neden olabilir (örn. hacim aşırı yüklenmesine yol açan koşullarda). Bu durumda, biyofiziksel kısıtlamalar CM'leri elongate'e zorlar ve AR 7:1'den 11:1'e değişir.
Membranlarda sinyalleme aktivitesi hücresel AR, boyut, membran yüzey alanı ve membran eğriliği18gibi küresel hücre geometrisi parametrelerine bağlıdır. Yenidoğan sıçan CM'leri hücreleri belirli bir uzunlukta sınırlandırmak için desenli substratlara kaplandığında: genişlik AR, oranlar sağlıklı bir erişkin kalpteki hücrelere benzer olduğunda en iyi kontraktil fonksiyonu gösterdiler. Buna karşılık, oranları başarısız kalplerde miyositler benzer olduğunda kötü performans19. Hipertrofisin erken evrelerinde, kesit seli ndeki artıştan da yansıyan hücreler genişler. HF hipertrofisin ilerleyen aşamalarında ortaya çıkar ve hücreler genellikle uzamış görünür. Bu nedenle, kronik hipertrofi in vivo sıçan modelleri yaklaşık% 3020sol ventriküler miyosit uzunluğu bir artış rapor etmiş olması şaşırtıcı değildir , ama akut in vitro hipertrofik uyaranlarla tedavi edildi transgenik fare modelinden yetişkin CMs hücre genişliği yerine benzer artışlar gösterdi21.
Tek hücreli RNA dizilimi, tek hücreli transkripsiyon kesin analizisağlar, şu anda hücre biyolojisi anlayışı devrim. Bu teknoloji, bireysel hücre şekillerinin gen ekspresyonunu nasıl etkilediği sorusunu yanıtlamak için tercih edilen bir yöntemdi. Tek hücreleri farklı şekillerde, özellikle 1:1, 7:1 veya 11:1'lik AR'larla karşılaştırdık. Bu, yenidoğan sıçan ventriküler CM'lerinin 1:1, 7:1 veya 11:1 tanımlı ar'leri ile fibronektin kaplı mikro desenler2 ile doldurulmuş özel olarak tasarlanmış bir çipe tohumlanması yla yapıldı. Mikrodesenler fotolitografi teknolojisi kullanılarak üretildi. Mikro desenler fibronektin ile kaplandı, sitofobik yüzeyle çevriliydi. Bu nedenle, CM'ler sitobik alan kaçınArak, sadece fibronektin substrat üzerinde büyüyen mikrodesenler tanımlanan AR eklemek, yaymak ve yakalamak. Mikro desenler iyi şekilli bir biçimde değildir. Bunun yerine, fibronektin düzeyi tam olarak çevreleyen sitobik alanın aynı yükseklikte. Bu, çevredeki duvarlardan herhangi bir stres olmadığı için Petri kabındaki büyüyen hücrelere benzer koşullar sağladı. Buna ek olarak, farklı AR ile mikro desenlerin yüzey alanı eşittir.
Deneysel tasarımın, toplu RNA dizilimi yerine tek hücreli RNA dizilimi nin kullanılmasına yol açan iki önemli yönü vardı. İlk olarak, mikro desenlerin sadece birkaç yüzdesi tek bir hücre tarafından işgal edilebilir. İkinci olarak, bazen tek bir hücre mikro desen yüzeyini tam olarak kaplamaz. Tek hücreli RNA analizi için mikro desen yüzeyini tamamen kaplayan tek hücreler seçilmelidir. Bir çip üzerindeki kaplama hücrelerinin sadece bir alt grubu her iki kriteri de karşıladığı için, sadece tüm çipi denemek ve toplu RNA sıralaması için tüm hücreleri toplamak mümkün değildi. Nitelikli hücrelerin yarı otomatik hücre seçici kullanılarak tek tek seçilmesi gerekiyordu.
Şu anda CM şekli, kendisi tarafından, miyokardiyal senkron üzerinde bir intra-fonksiyonel etkisi olup olmadığı bilinmemektedir. Bu yazıda önerilen yöntemlerin temel amacı, hücre şeklinin transkripsiyon17üzerinde bir etkisi olup olmadığını incelemek için yeni bir platform geliştirmekti. In vitro çalışmalar in vivo çalışmalardan farklı olmasına rağmen, bu çalışmanın amacı farklı hücre şekillerinin gen ekspresyonu üzerindeki etkisini araştırmak, in vivo'da farklı şekillerle hücreleri karşılaştırmanın son derece zahmetli olduğunu göz önünde bulundurarak. Bu deneyler, benzer bir yaklaşım kullanan ve hücre şeklindeki değişiklikler nedeniyle fizyolojik parametrelerde değişiklikler gözlemlediklerini bildiren Kuo ve ark.19'danesinlenerek yapıldı.
Protokol
Hayvanlarla ilgili tüm prosedürler, İsveç'in Stockholm'deki Karolinska Enstitüsü hayvan etik komitesinin yönetmeliklerine uygundu.
1. Mikro desenli talaş düzeni
- Borosilikat camüzerine fotolitografi ile basılmış, aktif mikrodesenli 19,5 mm x 19,5 mm'lik bir kapak fişidir.
NOT: Bu mikro desenler sitofobik bir alanla çevrilidir. Bu nedenle, bir tohumlu hücre sadece bu mikro desenlerden birine bağlanabilir ve büyüyebilir ve bu mikro desenin AR'ını yakalayabilir. Çip üç bölgeye ayrılır ve her bölge belirli bir AR ile mikro desenleroluşur. Fiş düzeni Şekil 1B'degösterilmiştir. Tanımlanan AR'lerin geometrisi Tablo 1'desunulmuştur. Fibronektin mikro desenlerinin farklı şekillerinin büyütülmüş floresan görüntüleri Şekil 1B'dekialt görüntülerde gösterilmiştir.
2. Kaplama mikro desenli yongaları
- Her talaş için 2 mL fosfat tamponlu saline 80 μg fibronektin ekleyerek 2x kaplama protein çözeltisi hazırlayın (PBS-/-).
- Bir çipi 35 mm'lik Greiner Petri kabına aktarın ve hemen 2 mL PBSekleyin -/-. Daha sonra 2x kaplama protein çözeltisinin 2 mL'sini ekleyin.
- 2 saat oda sıcaklığında cips kuluçka.
- Kaplama çözeltisini art arda seyreltme adımlarıyla PBS-/-ile yıkayın. Talaş yüzeyi her zaman ıslak olmalıdır. Daha sonra PBS'yi 2 mL kaplama orta sı ve inkübünle 37 °C'de tohumlama hücrelerine kadar değiştirin.
3. CM'lerin İzolasyon
- DMEM:M199 (4:1) %10 at serumu, %4 fetal sığır serumu, %2 HEPES (1 M) ve %1 penisilin/streptomisin (10.000 U/mL)22ile kaplama ortamını hazırlayın.
- 2 günlük yenidoğan sıçan kalplerinin sol ventrikülünden dokuyu inceleyin ve PBS içeren 10 cm'lik bir tabağa aktarın. Yaklaşık 1 mm3 adet doku kesin.
- Hasat edilen dokuyu, doku dissosiyamı için kapakta bir rotor la donatılmış bir rotor-kapak tüpüne aktarın(Malzeme Tablosu). Doku yerleşmek ve daha sonra dikkatle supernatant çıkarın.
- Neonatal Kalp Ayrıştırma Kiti(Malzeme Tablosu)kullanılarak hazırlanan 1 ve 2 enzim karışımının 2,5 mL'sini C Tüpüne ekleyin ve kapağı sıkıca kapatın.
- Rotor-kapak tüpünü ısıtıcılarla donatılmış dissosiyatörün koluna takın (Şekil 2A veB) ( MalzemeTablosu). Yaklaşık bir saat süren kuluçka programını(Şekil 2C)37C_mr_NHDK_1 çalıştırın.
- Kuluçka programı çalışırken PBS,pH 7.2'de 2 mM EDTA ve %0.5 büyükbaş serum albumin (BSA) içeren PEB tamponunu hazırlayın ve 4 °C'de tutun.
- Kuluçka programı sona erdikten sonra rotor-kapak tüpünü(Şekil 2D)ayırın ve önceden ısıtılmış kaplama ortamından 7,5 mL ekleyin.
- Numuneyi yeniden askıya alın ve hücre süspansiyonuna 70 μm'lik bir süzgeç kullanarak filtre uygulayın.
- Süzgeci 3 mL daha kaplama aracıyla yıkayın.
- 5 dakika için 600 x g hücre süspansiyon santrifüj. Supernatant'ı tamamen aspire edin.
- Hücre peletini 60 μL soğuk PEB tamponunda yeniden askıya alın.
- 20 μL Neonatal Kardiyomiyosit İzolasyon Kokteyli(Malzeme Tablosu),mikron boyutlu boncuklar içeren ve CM'siz leri hedef alın.
- Anti-Kırmızı Kan Hücresi boncuk 20 μL ekleyin (Malzeme Tablosu).
- Süspansiyonu karıştırın ve 4 °C'de 15 dakika kuluçkaya yatırın.
- 400 μL PEB arabellek ekleyin.
- Hücre süspansiyonunu, mıknatıslı bir standa dikey olarak yerleştirilen ve PEB tamponu yla iyi yıkanan LD sütununa(Malzeme Tablosu)uygulayın.
- Etiketlenmemiş hücreleri toplayın ve 0,5 mL PEB arabelleği içeren sütunu yıkayın.
- 8 mL kaplama ortamı ekleyin ve hücre süspansiyonuna 75cm'lik 2 kaplamasız kültür şişesine aktarın ve 37 °C'de 1,5 saat boyunca kuluçkaya yatırın. Kalan non-CM'ler kaplamasız hücre kültürüne uymaya başlayacak ve hücre süspansiyonu CM popülasyonu tarafından zenginleştirilmiş olacaktır.
4. Desenleme CM'leri
NOT: Tek CM'leri 1:1, 7:1 veya 11:1'lik AR'larla karşılaştırdık. Bu, izole neonatal sıçan CM'lerinin 1:1, 7:1 veya 11:1 tanımlı ar'larla fibronektin kaplı mikro desenlerle dolu özel olarak tasarlanmış bir çipe tohumlanmasıyla yapılır. Mikro desenler fibronektin ile kaplandı, sitofobik yüzeyle çevriliydi. Bu nedenle, CM'ler sadece fibronektin substrat üzerinde büyüyerek mikro desenlerin tanımlanan AR'ını takacak, yayacak ve yakalayacak. İzole CM'leri aşağıdaki adımlara göre desenle.
- Hücre süspansiyonuna 15 mL'lik bir tüp aktarın, hücreleri sayın ve uygun kaplama ortamı ekleyerek mL başına 100.000 hücre konsantrasyonuna seyreltin.
- 35 mm'lik Greiner Petri kabının içine 2 mL sıcak kaplama ortamına batırılmış olan çipe 2 mL hücre süspansiyonu ekleyin.
- CM'lerin fibronektin kaplı mikro desenlere bağlanmasına izin vermek ve her CM'nin substrat mikrodemasını elde etmesini sağlamak için yemeği %5 CO2 ile 37 °C'de kuluçkaya yatırın.
- Saat 18'den sonra çipi kontrol et. Hücrelerin çoğu bağlıysa, desenli hücreye bağlı olan enkaz ve ölü hücreleri ayırın ve çıkarın. Bunu, kaplama ortamını çıkararak ve çipin merkezinden başlayarak pbs-/- damla gibi ekleyerek ve sonra yanlara doğru hareket ederek yapın. 2 kez tekrarlayın.
- DMEM:M199 (4:1) %4 at serumu, %4 fetal sığır serumu, %2 HEPES (1 M) ve %1 penisilin/streptomisin (10.000 U/mL) ile taze bakım ortamı ile PBS'yi aspire edin.
5. Yapıştırma CM'leri çekme
NOT: 72 saatlik bir kültürlenme süresinden sonra, desenli tek CM'ler fibronektin mikro desenlerinden yarı otomatik hücre toplayıcısı(Malzeme Tablosu)kullanılarak seçilir (Şekil 3). Hücre seçici motorlu sahne(Şekil 3A)kontrol etmek için bir yazılım23 kullanır. 70 μm cam mikrocapiller(Şekil 3B)desenli yenidoğan sıçan CM'lerini almak ve enjekte etmek için kullanılır. Hücre seçici bir vakum üreten ve basınç uygulayarak hücreleri enjekte ederek yapışık hücreleri sıralar. 1 numaralı şırıngadaki vakum şırınga pompası kullanılarak çekilerek uygulanır (Şekil 3C). Hidrostatik basınç yerçekimine dayanır ve mikroskop masasının üzerine 87 cm mesafede şırınga sayısı 2 yerleştirilerek indüklenir. Şırıngalar 1 ve 2 sırasıyla PTFE tüpleri ile kontrol ünitesine gömülü olan 1 ve 2 numaralı vanalara bağlanır(Şekil 3D). PTFE tüpleri tamamen RNase içermeyen su ile doldurulur. Daha sonra toplanan tek hücre, 3.55 l likiz tamponu içeren polimeraz zincir reaksiyonu (PCR) tüpüne enjekte edilir.
- 72 saatlik bir kültür den sonra, eski ortamı çıkarın ve talaş yüzeyini ılık DPBS-/-ile hafifçe yıkayın.
- Çanak düz tutun ve yemeğin bir tarafında 1000 μL pipet ile yavaşça eski orta aspire. Çipin her zaman ıslak kaldığından emin olun.
- 2 mL DPBSekleyin -/- yavaşça ve damla-bilge desenli hücrelere bağlı ölü hücrelerin çoğu ayırmak, çipin merkezinden başlayarak ve daha sonra kenarlarına hareket.
- DpBS'nin çoğunu, çipin merkezinden başlayıp kenarlara doğru hareket ederek mümkün olduğunca çok sayıda bağımsız kayan hücreyi çıkarmak için aspire edin.
- 5.1.2 ve 5.1.3 adımlarını bir kez daha tekrarlayın.
- TalaşKenarını kapmak için açılı çalgıları kullanın ve toplarken yüzen hücrelerin sayısını azaltmak için hemen yeni bir steril 35 mm Greiner Petri kabına aktarın.
- Talaşın kuruması için hemen 1,5 mL DPBS-/- ekleyin.
- Canlı hücrelerin çekirdeklerini görselleştirmek için 1,5 l Canlı Boya Döngüsü yeşili ekleyin.
- Talaşucunu kullanarak çipi Greiner Petri kabının ortasına yerleştirin.
- Çipin üzerine bir hazne(Malzeme Tablosu)koyun. Oda, hücre desenlerine erişimi engellemeden çipi yemeğin dibine sabitleyecek.
- Greiner Petri kabını hücre toplayıcı aşamasının kabının üzerine monte edin ve manyetik kapağı yerleştirin.
- Otomatik enjeksiyonu kalibre edin.
- Live View penceresinde görüntünün ortasındaki motorlu sahneye kazınmış olan artı işaretini bulun.
- Artı işaretine odaklanın ve Tarama ve sıralama penceresinde otomatik enjeksiyon düğmesi için Kalibrasyon'u seçin.
- DPBS-/- 1.5 mL DPBS/tripsin-/- (1:1) ile değiştirin, böylece hücreleri almak için sıvı bir vakum kullanılabilir fibronektin hücreleri gevşetin.
- Tarama ve sıralama penceresindeki Tarama sekmesini kullanarak tüm çipi tarayıp taramayı. Çipin sol üst köşesini görüş alanında bulun ve sol üst köşe satırındaki geçerli mikroskop pozisyonunu al'a tıklayın.
- Daha sonra, motorlu sahneyi çipin sağ alt köşesine taşıyın. Mikroskobu odaklayın ve sağ alt köşe satırında geçerli mikroskop pozisyonunu al'a tıklayın.
- En keskin düzlemi ayarla düğmesine ve açılan pencereye tıklayın sağ üst köşeye git'etıklayın. Mikroskobu odaklayın ve sol alt köşeye git'e tıklayın ve odağı ayarlayın. Bittiğinde Bitir düğmesini tıklatın ve tarama başlatın.
- Tarama sonlandırıldığında, Çözümleme sekmesine gidin ve çalışma ölçütlerini geçen tek hücreleri seçin.
- Cam mikrokapillerin mikroskobun canlı görüntüsünün ortasında olduğundan emin olun.
- Yazılımın Pompa penceresini kullanarak şırınga pompasını kontrol edin. Çapı 27 mm olan 50 mL'lik 1 şırıngadan 4 mL çekerek bir vakum oluşturun.
- Sıralama sekmesinde
- Vanaların enjeksiyon parametrelerini ayarlayın. Tek bir hücreyi teslim eden enjeksiyon hacminin 1 μl olduğu, eğer valf 2'nin 120 milisaniye, valf 1'in 200 milisaniyelik bir zaman atlamadan sonra 20 saniye açıldığı hesaplanmıştır. Tüplerin elastikiyeti nedeniyle, açık vana 1 akışı enjekte durdurmak için.
- Vanaların pick-up parametrelerini ayarlayın. Eğer valf 1 20 milisaniye ve sonra valf 2 10 milisaniye, 10 milisaniye lik bir zaman atlamadan sonra açılırsa, desenli hücrelerin çoğunun alınabileceği hesaplanmıştır. Bunun nedeni, fibronektin ciltlerinin tripsin ile tedavi edildikten sonra gevşetilmiş olmasıdır.
- Yolu Hesapla düğmesine tıklayın. Yazılım, seçilen hücreleri çip boyunca toplamak ve enjekte etmek için hücreden hücreye en hızlı yolu oluşturur.
- Mikroskobu çip yüzeyindeki desenli bir hücreye odakla.
- Joystick kullanarak, mikrokapiller dikkatle aşağı hareket ettirin, böylece mikrokapillerucun ucunun en keskin görüntüsü hücreye dokunmadan elde edilebilir.
- Micropipette ofset bölümündeki Set düğmesini tıklatın. Mikrokapiller kesiti gösteren yeni bir pencere açılır. Kılcal damarın tam merkezine tıklayın. Yazılım daha sonra x, y ve z koordinatlarında kılcal uç ofset kaydeder.
- Başlat sıralama düğmesini kullanarak sıralamayı başlatın.
Sonuçlar
Doku 2 günlük yenidoğan sıçan kalplerinin sol ventrikülden kesildi ve tek hücrelere bölündü. Daha sonra zenginleştirilmiş CM'ler farklı AR'lere sahip fibronektin desenleri içeren bir çipin üzerine tohumlandı. 72 saatlik kültürden sonra ortam, canlı hücrelerin çekirdeklerini görselleştirmek için DPBS'de 1:1000 Vibrant Dye Cycle yeşili ile değiştirildi./- 2 dakika boyunca canlı hücrelerin çekirdeklerini görselleştirmek için. Daha sonra hücreler, fibronektin'deki hücreleri gevşetmek için DPBS-/-/tripsin (1:1) ile tedavi edildi, böylece hücre toplamayı kolaylaştırmak için akışkan bir vakum kullanılabilirdi. Bu arada, tüm çip hücre toplayıcısına bağlı ters bir mikroskop kullanılarak 10x'lik bir büyütmede tarandı. Bu, tripsin tedavisi nedeniyle hücreler yuvarlanmadan önce gerçekleştirilmiştir. Kalifiye hücreler, taranmış görüntüye göre seçildi ve koordinatları hücre seçici yazılımına kaydedildi. Mikro desenler sadece mononükleli tek hücreli yse ve sadece hücre fibronektin mikrodesini tamamen kapladığında seçilmiştir. Hücre ayırıcısı seçilen hücreleri tek tek seçti ve başarılı bir şekilde seçilen her bir hücre hemen tek bir PCR tüpüne enjekte edildi ve mikroskop aşamasına yerleştirildi. Her PCR tüpü nde 3,55 μL likis tamponu bulunur(Tablo 2). Ortamın kaldırılmasıyla başlayan sıralama işlemi 40 dakika içinde tamamlandı. Tamamlayıcı deoksiribonükleik asit (cDNA) sentezi, PCR ön amplifikasyon ve arıtma, Smart-Seq2 protokolü24 'e göre lysed tek hücrelerüzerinde gerçekleştirildi (Tablo 3). Saflaştırılmış cDNA'nın kalitesi otomatik bir elektroforez analizörü tarafından kontrol edildi. Seçilmiş tek bir hücrenin önceden güçlendirilmiş cDNA'sının elektropherogramı Şekil 4'teveda edilmiştir. RNA-Seq kütüphaneleri Smart-Seq2 protokolü24'egöre hazırlanmıştır.
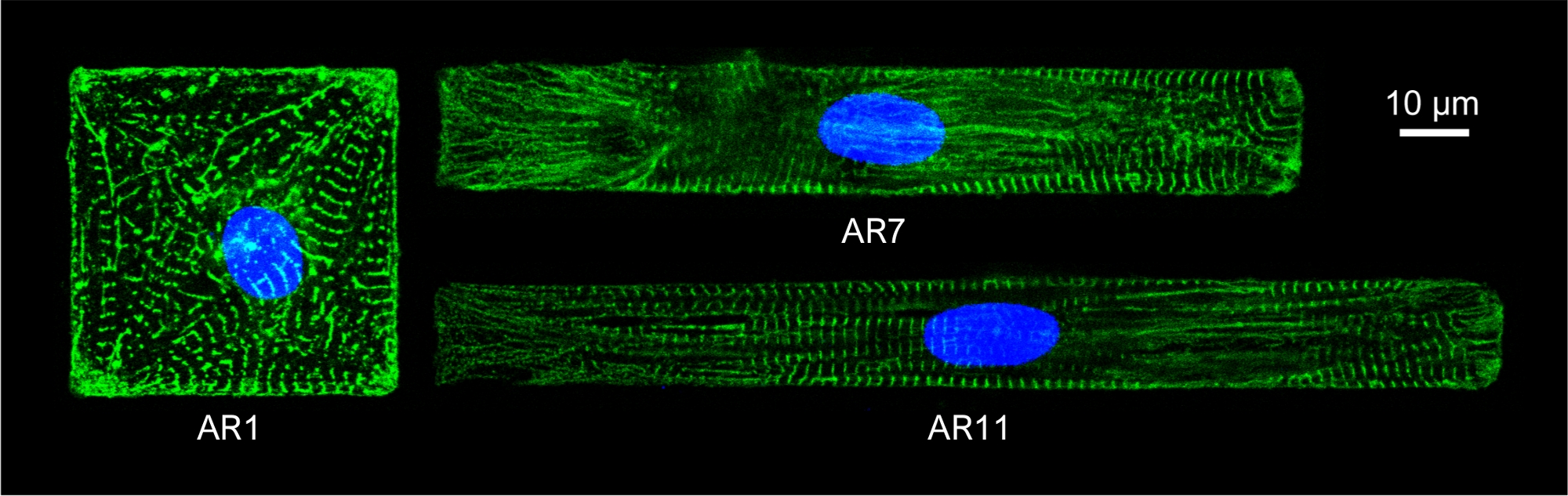
Desenli CM'lerin sarcomere yapısını gözlemlemek için desenli CM'ler sarkoerik α aktinin antikor ile boyanmıştır. Hücreler Eşek Anti-Mouse IgG Alexa Fluor 488 1:800 ile ikincil boyama için oda sıcaklığında 1 saat kuluçkaya yatırıldı. Çekirdekler 1 μg/mL DAPI ile boyandı. İmmünofloresan görüntüler ters konfokal mikroskop la elde edildi, 63x yağ daldırma (NA 1.4) hedefi kullanılarak(Şekil 5).

Şekil 1: Çipin fibronectin mikro desenli düzeni.
(A) Özel olarak tasarlanmış çipin görüntüsü. Çip, fotolitografi ile borosilikat cam üzerine basılmış fibronektin mikrodesenli 19,5 mm x 19,5 mm'lik bir kapaktır. (B) Fiş düzeni. Çip üç bölgeye ayrılır ve her bölge özel AR ile fibronektin mikro desenleroluşur. Bu şekil Haftbaradaran Esfahani ve ark. 2 tarafından "ek malzeme 1" den değiştirilmiştir,http://creativecommons.org/licenses/by/4.0/ altında kullanılan . Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: Isıtıcı aparatı ile donatılmış dissociator.
(A) 2 günlük yenidoğan sıçan sol ventriküllerin tam otomatik ayrıştırma için kullanılan tüm dissociator aracı. (B) Isıtma ünitesi. (C) Rotor-kapak tüpü. (D) Doku dissosiasyonunun tam otomatik iş akışı için kullanıma hazır programlar. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: Hücre toplayıcı aparatı.
(A) Hücre seçicinin motorlu aşamasının genişletilmiş görünümü. Bir Petri-çanak tutucu ve 10 PCR şeritler için 80 delik ve kalibrasyon crosshair için bir delik sahnede gömülüdür. (B) Bir cam mikrokapiller genişlemiş görünümü. (C) Şırınga pompası. (D) 1 ve 2 sayılı vanaların açılış ve kapanış zaman penceresini kontrol eden kontrol ünitesi, kontrol ünitesinin içine monte edilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 4: Seçilmiş tek bir hücrenin önceden güçlendirilmiş cDNA'sının elektropherogramı.
19 PCR pre-amplifikasyon döngüsü 1 ng/μL saflaştırılmış cDNA verimi elde etmek için kullanıldı. Jel benzeri densitometri çiziminde açık bir bant elektrofeogramda 1852 bp'deki zirveye karşılık gelir. Parçaların ortalama boyutu 1588 bp'dir. Ayrıca, 300 bp'den kısa olan küçük miktardaki parçalar iyi bir cDNA kitaplığını gösterir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 5: Farklı AR'li desenli CM'lerin α-aktin sarkoerik yapısının (yeşil) ve çekirdeğinin (mavi) immünororesan boyama.
Kromatin DAPI tarafından lekelendi. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
| Morfotip | Ar | Uzunluk (3m) | Genişlik (3m) | Fibronektin alanı (μm2) |
| AR1 | 1:1 | 47 | 47 | 2209 |
| AR7 | 7:1 | 126 | 18 | 2268 |
| AR11 | 11:1 | 155 | 14 | 2170 |
Tablo 1: Desenli CM'lerin geometrisi.
| Bileşen | Hacim (3L) |
| Nükleazsız su | 0.65 |
| (%0,4 vol/vol) Triton X-100 | 1.8 |
| dNTP karışımı (25 mM) | 0.8 |
| RNase inhibitörü (40 U μL-1) | 0.1 |
| Oligo-dT30VN oligonükleotid (100 μM) | 0.1 |
| ERCC RNA Spike-In Mix (2,5 x 105 seyreltme) | 0.1 |
| Enjekte edilen tek hücre | 1 |
| Toplam hacim | 4.55 |
Tablo 2: Tek hücreli özel lisis arabellek.
| Bileşen | Hacim (3L) |
| Superscript II ilk iplikçik arabellek (5x) | 2 |
| DTT (100 mM) | 0.5 |
| Betain (5 M) | 2 |
| Mgcl2 (1 M) | 0.1 |
| RNase inhibitörü (40 U μL-1) | 0.25 |
| Superscript II ters transkriptaz (200 U μL-1) | 0.5 |
| TSO (100 μM) | 0.1 |
| Toplam hacim | 5.45 |
Tablo 3: Tek bir CM'nin lysate'sinden birinci iplikçik cDNA sentezlemek için bir RT reaksiyonu için ters transkripsiyon (RT) karışımı.
Tartışmalar
Bu çalışmada tek hücreli RNA dizilimi kullanılmıştır, bu da tek hücreli transkripsiyonu tespit edebilen yeni ve güçlü bir teknolojidir. Tek CM'leri biraraya getirmek için yenilikçi bir yaklaşımla birleştirildi, böylece farklı AR'ler aldılar, aksi takdirde, sadece in vivo gözlemlenebilirdi.
Çalışmanın bazı sınırlamaları vardı. Örneğin, yenidoğan CM'leri farklı morfotipler üretmek için kullanılmak zorunda kalmak zorundaydı, çünkü tanımlanmış şekillerde 72 saat boyunca yeterince hayati yetişkin CM'leri oluşturmak son derece zor. Ayrıca, CMs 72 saat ex vivo için kültürlü edildi, hangi gen ekspresyonu deseni üzerinde bir etkisi olabilir. Ancak, bu culturing gerekli, böylece hücrelerin belirli morfotipler oluşturabiliyordu. Ayrıca, sadece mononükleatlı ve fibronektin mikrodelenini tamamen kaplayan tek hücreler sıralama için seçilmiştir. Her fiş üzerinde desenli hücreler sıralama sadece bir tur halinde sıralanmış olmalıdır. Son olarak, seçilen hücrelerin kabaca üçte biri olan yaklaşık 50 hücre her çipten başarıyla alındı. Başarıyla seçilen hücrelerin sayısını kısıtlayan iki nedeni vardır. İlk olarak, bazı hücreler fibronektin desenine çok sıkı bağlandı ve pikap akışı onları başarıyla alacak kadar zorla verilmedi. İkincisi, tripsin tedavisi nedeniyle, bazı hücreler ve fibronektin arasındaki eki çok gevşek oldu. Sonuç olarak, bu hücreler fibronektin mikro desenlerinden uzaklaştırıldılar, mikrokapiller onlara yaklaştığında, ve alınmadılar. Yazarlar bu kurulum bir in vivo ortam ile aynı olduğunu iddia yok, ama araştırma soruya cevap için uygun bir yaklaşım olduğunu kanıtladı.
Önerilen yöntem farklı hücre tipleri için geçerlidir (örneğin, hiPS-CM'ler için). Ancak, aşağıdaki faktörler diğer hücre türlerini incelemek için optimize edilmelidir. Mikro desenlerin kapılması için belirli hücre tipine bağlanmaiçin uygun ECM yapıştırıcı moleküller kullanılmalıdır. Mikro desenlerin geometrisi çalışma sorusuna ve hücre tipine göre değiştirilmelidir. Kültür süresi çalışma sorusuna göre değiştirilebilir. Müfreze reaktifi ve kuluçka süresi çalışma hücresi tipi için tam olarak optimize edilmelidir. Örneğin, Accutase yerine Embriyonik ve nöronal kök hücrelerin ayrılması için TryplE kullanılabilir. Kapakçıkların açılış süresi parametreleri hücreleri başarılı bir şekilde, ancak yavaşça seçmek için dikkatle incelenmelidir. Özetle, bu alandaki araştırmacılar için değerli bir kaynak sağlayabilecek hücre şeklini incelemek için yeni bir platform tasarladık. Bu bağlamda, hücresel mimari ile gen ekspresyonu arasındaki etkileşimi tanımlamak için hemodinamik kısıtlamalar la CM in vivo'ya dayatılan in vitro karakteristik şekilleri taklit eden deneysel bir yaklaşım tasarladık. Ayrıca HF in vitro incelemek için yeni bir platform geliştirilmesi ve gen ekspresyonunun güçlü bir belirleyicisi olarak hücre şeklinin tanımlanmasını da rapor ediyoruz. Bu biyoloji ve tıp için geniş kapsamlı etkileri ile yeni bir gözlemdir.
Açıklamalar
Hiçbiri.
Teşekkürler
Hiçbiri.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 2100 Bioanalyzer Instrument | Agilent Technologies | G2939BA | Automated electrophoresis analyzer |
| Anti-Red Blood Cell MicroBeads | Miltenyi Biotec | 130-109-681 | |
| Axio Observer microscope | Zeiss | Z1 | Inverted microscope |
| Betaine solution (5 M) | Sigma-Aldrich, MERCK | B0300 | |
| CellSorter | CELLSORTER | https://www.singlecellpicker.com/ | Cell picker |
| CYTOOchamber | CYTOO | 30-010 | Custom-designed chip |
| CYTOOchip | CYTOO | 10-950-00-18 | Chamber |
| DMEM | Thermo Fisher Scientific | 31966-021 | high glucose, GlutaMAX Supplement |
| dNTP mix (25 mM) | Thermo Fisher Scientific | R1122 | |
| Donkey Anti-Mouse IgG Alexa Fluor 488 | Abcam PLC | ab150105 | |
| DTT (100 mM) | Thermo Fisher Scientific | 18064071 | |
| ERCC RNA Spike-In Mix | Thermo Fisher Scientific | 4456740 | |
| Fetal Bovine Serum | Thermo Fisher Scientific | 10082-147 | |
| Fibronectin | Sigma-Aldrich, MERCK | F4759 | |
| gentleMACS C Tube | Miltenyi Biotec | 130-093-237 | Rotor-cap tube |
| gentleMACS Octo Dissociator with Heaters | Miltenyi Biotec | 130-096-427 | Dissociator with heater |
| Greiner CELLSTAR Petri dish | Sigma-Aldrich, MERCK | P6987 | |
| HEPES (1 M) | Thermo Fisher Scientific | 15630-056 | |
| Horse Serum | Sigma-Aldrich, MERCK | H0146 | |
| LD column | Miltenyi Biotec | 130-042-901 | |
| Medium 199 | Thermo Fisher Scientific | 31150-022 | |
| NE-1000 syringe pump | New Era Pump Systems | NE-1000 | Syringe pump |
| Neonatal Cardiomyocyte Isolation Cocktail, rat | Miltenyi Biotec | 130-105-420 | |
| Neonatal Heart Dissociation Kit, mouse and rat | Miltenyi Biotec | 130-098-373 | |
| Oligo-dT30VN oligonucleotides | IDT Technology | 5′–AAGCAGTGGTATCAACGCAGAGTACT30VN-3′ | |
| RNAse inhibitor (40 U µL-1) | Clontech | 2313A | |
| sarcomeric α-actinin | Sigma-Aldrich, MERCK | EA-53 | |
| SP8 confocal microscope | Leica Microsystems | SP8 | Confocal microscope |
| Superscript II first-strand buffer (5x) | Thermo Fisher Scientific | 18064071 | |
| Superscript II reverse transcriptase (200 U µL-1) | Thermo Fisher Scientific | 18064071 | |
| Triton X-100 | Sigma-Aldrich, MERCK | T9284 | |
| TryplE Express enzyme, no phenol red | Thermo Fisher Scientific | 12604013 | |
| TSO (100 µM) | QIAGEN | 5′-AAGCAGTGGTATCAACGCAGAGTACATrGrG+G-3′ | |
| Vibrant Dye Cycle green | Thermo Fisher Scientific | V35004 |
Referanslar
- Heineke, J., Molkentin, J. D. Regulation of cardiac hypertrophy by intracellular signalling pathways. Nature Reviews Molecular Cell Biology. 7 (8), 589-600 (2006).
- Haftbaradaran Esfahani, P., et al. Cell shape determines gene expression: cardiomyocyte morphotypic transcriptomes. Basic Research in Cardiology. 115 (1), 7(2019).
- Kontrogianni-Konstantopoulos, A., Benian, G., Granzier, H. Advances in Muscle Physiology and Pathophysiology 2011. Journal of Biomedicine and Biotechnology. 2012, 930836(2012).
- Hill, J. A., Olson, E. N. Cardiac plasticity. New England Journal of Medicine. 358 (13), 1370-1380 (2008).
- Bray, M. A., Sheehy, S. P., Parker, K. K. Sarcomere alignment is regulated by myocyte shape. Cell Motility and the Cytoskeleton. 65 (8), 641-651 (2008).
- Benjamin, I. J., Schneider, M. D. Learning from failure: congestive heart failure in the postgenomic age. Journal of Clinical Investigation. 115 (3), 495-499 (2005).
- Opie, L. H., Commerford, P. J., Gersh, B. J., Pfeffer, M. A. Controversies in ventricular remodelling. Lancet. 367 (9507), 356-367 (2006).
- Braunwald, E. Cardiomyopathies: An Overview. Circulation Research. 121 (7), 711-721 (2017).
- Knoll, R., et al. The cardiac mechanical stretch sensor machinery involves a Z disc complex that is defective in a subset of human dilated cardiomyopathy. Cell. 111 (7), 943-955 (2002).
- Frangogiannis, N. G. The Extracellular Matrix in Ischemic and Nonischemic Heart Failure. Circulation Research. 125 (1), 117-146 (2019).
- Marian, A. J., Braunwald, E. Hypertrophic Cardiomyopathy: Genetics, Pathogenesis, Clinical Manifestations, Diagnosis, and Therapy. Circulation Research. 121 (7), 749-770 (2017).
- Mozaffarian, D., Caldwell, J. H. Right ventricular involvement in hypertrophic cardiomyopathy: a case report and literature review. Clinical Cardiology. 24 (1), 2-8 (2001).
- Olsson, M. C., Palmer, B. M., Stauffer, B. L., Leinwand, L. A., Moore, R. L. Morphological and functional alterations in ventricular myocytes from male transgenic mice with hypertrophic cardiomyopathy. Circulation Research. 94 (2), 201-207 (2004).
- Toepfer, C. N., et al. Hypertrophic cardiomyopathy mutations in MYBPC3 dysregulate myosin. Science Translational Medicine. 11 (476), (2019).
- McKenna, W. J., Maron, B. J., Thiene, G. Classification, Epidemiology, and Global Burden of Cardiomyopathies. Circulation Research. 121 (7), 722-730 (2017).
- Schultheiss, H. P., et al. Dilated cardiomyopathy. Nature Reviews Disease Primers. 5 (1), 32(2019).
- Knoll, R. A role for membrane shape and information processing in cardiac physiology. Pflugers Arch. 467 (1), 167-173 (2015).
- Rangamani, P., et al. Decoding information in cell shape. Cell. 154 (6), 1356-1369 (2013).
- Kuo, P. L., et al. Myocyte shape regulates lateral registry of sarcomeres and contractility. American Journal of Pathology. 181 (6), 2030-2037 (2012).
- Gerdes, A. M., Onodera, T., Wang, X., McCune, S. A. Myocyte remodeling during the progression to failure in rats with hypertension. Hypertension. 28 (4), 609-614 (1996).
- Kehat, I., et al. Extracellular signal-regulated kinases 1 and 2 regulate the balance between eccentric and concentric cardiac growth. Circulation Research. 108 (2), 176-183 (2011).
- Sreejit, P., Kumar, S., Verma, R. S. An improved protocol for primary culture of cardiomyocyte from neonatal mice. In Vitro Cellular & Developmental Biology - Animal. 44 (3-4), 45-50 (2008).
- Kornyei, Z., et al. Cell sorting in a Petri dish controlled by computer vision. Scientific Reports. 3, 1088(2013).
- Picelli, S., et al. Full-length RNA-seq from single cells using Smart-seq2. Nature Protocols. 9 (1), 171-181 (2014).
- Schmick, M., Bastiaens, P. I. H. The interdependence of membrane shape and cellular signal processing. Cell. 156 (6), 1132-1138 (2014).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır