
Method Article
Ein Ansatz zur Untersuchung formabhängiger Transkriptomik auf einer Ebene einer einzelzelligen Ebene
In diesem Artikel
Zusammenfassung
Dieses Papier stellt Methoden für den Anbau von Herzmyozyten mit verschiedenen Formen, die verschiedene Pathologien darstellen, und das Sortieren dieser anhaftenden Herzmyozyten basierend auf ihrer Morphologie auf einer einzelnen Zellebene. Die vorgeschlagene Plattform bietet einen neuartigen Ansatz für hohe Durchsatz- und Arzneimittelscreenings für verschiedene Arten von Herzinsuffizienz.
Zusammenfassung
Verschiedene Arten von Herzhypertrophie wurden mit einem erhöhten Volumen von Herzmyozyten (CMs) verbunden, zusammen mit Veränderungen in der CM-Morphologie. Während die Auswirkungen des Zellvolumens auf die Genexpression bekannt sind, sind die Auswirkungen der Zellform nicht gut verstanden. In diesem Artikel wird eine Methode beschrieben, die entwickelt wurde, um die Auswirkungen der CM-Morphologie auf die Genexpression systematisch zu analysieren. Es beschreibt die Entwicklung einer neuartigen einzelzelligen Fangstrategie, der dann eine einzellige mRNA-Sequenzierung folgt. Es wurde auch ein mikrogemusterter Chip entwickelt, der 3000 rechteckige Fibronectin-Mikromuster enthält. Dadurch ist es möglich, CMs in unterschiedlichen Längen-Breiten-Seitenverhältnissen (AR) anzubauen, die verschiedenen Arten von Herzinsuffizienz (HF) entsprechen. Das Papier beschreibt auch ein Protokoll, das entwickelt wurde, um einzelne Zellen aus ihrem Muster zu holen, indem sie eine halbautomatische Mikropipetting-Zellauswahl verwenden und sie einzeln in einen separaten Lysepuffer injizieren. Dies hat es ermöglicht, die Transkriptome einzelner CMs mit definierten geometrischen Morphotypen zu profilieren und sie nach einer Reihe normaler oder pathologischer Bedingungen zu charakterisieren: hypertrophe Kardiomyopathie (HCM) oder Nachlast/Konzentrat versus dilatierte Kardiomyopathie (DCM) oder Vorspannung/Exzentrik. Zusammenfassend stellt dieses Papier Methoden zum Anbau von CMs mit unterschiedlichen Formen dar, die verschiedene Pathologien darstellen, und diese anhaftenden CMs basierend auf ihrer Morphologie auf einer Einzelzellebene zu sortieren. Die vorgeschlagene Plattform bietet einen neuartigen Ansatz für hohe Durchsatz- und Arzneimittelscreenings für verschiedene Arten von HF.
Einleitung
Nach Angaben der Weltgesundheitsorganisation ist Herz-Kreislauf-Erkrankungen (CVD) weltweit eine der Hauptursachen für Morbidität und Mortalität. CVD wirkt sich dramatisch auf die Lebensqualität der Menschen aus und hat enorme sozioökonomische Auswirkungen. Kardiomyopathien, wie HCM und DCM, sind primäre Erkrankungen des Herzmuskels und Die Hauptursachen für HF wurden mit hoher Morbidität und Mortalität in Verbindung gebracht. Es gibt viele Ursachen für HF, einschließlich Umweltauswirkungen, wie Infektionen und Exposition gegenüber Toxinen oder bestimmten Medikamenten8. HF kann auch durch genetische Veranlagung verursacht werden, nämlich Mutationen9. Es wird angenommen, dass die Veränderungen in der genetischen Zusammensetzung, die extrazelluläre Matrix (ECM) Moleküle beeinflussen, Integrine oder zytoskelettale Proteine für beeinträchtigte Mechanosensation und verschiedene Arten von Herzerkrankungen verantwortlich sein könnte10.
Das Hauptmerkmal von HCM ist unerklärliche Hypertrophie der linken Ventrikle11, und manchmal der rechten Ventrikle12, und dies stellt häufig mit überwiegender Beteiligung des interventrikulären Septums. HCM ist auch durch diastolische Dysfunktion und Myozyten-Disarray und Fibrose13gekennzeichnet. In den meisten Fällen wird der kontraktile Apparat des Herzens durch Mutationen in sarkomerischen Proteinen beeinflusst, was zu einer erhöhten Kontraktilität der Myozyten14führt. Im Gegensatz dazu ist DCM durch Diedilatation einer oder beider Ventrikel gekennzeichnet und hat eine familiäre Ätiologie in 30% bis 50% der Fälle15. DCM betrifft eine breite Palette von zellulären Funktionen, was zu einer beeinträchtigten Kontraktion der Myozyten, Zelltod und fibrotische Reparatur16.
Genetik hat gezeigt, dass bestimmte Arten von Mutationen einzelne CMs zwingen, während HCM3spezifische Formmerkmale anzunehmen, nämlich quadratische Zellen mit einer Länge:Breite AR, die fast gleich 1:14 (AR1) ist. Dasselbe gilt für DCM, mit länglichen Zellen mit einem AR, der fast gleich 11:1 (AR11) ist. Darüber hinaus kann HF durch erhöhte Nachbelastung (z.B. bei Bluthochdruck) verursacht werden. In diesen Fällen zwingen hämodynamische Anforderungen CMs, quadratische Formen gemäß dem Laplace-Gesetz zu übernehmen, und die AR ändert sich von 7:15 (AR7) auf 1:16,7. HF kann auch durch eine Erhöhung der Vorspannung verursacht werden (z.B. bei Bedingungen, die zu einer Volumenüberlastung führen). In diesem Fall erzwingen die biophysikalischen Einschränkungen die Dehnung der CMs, und die AR ändert sich von 7:1 auf 11:1.
Die Signalaktivität an Membranen hängt von globalen Zellgeometrieparametern wie der zellulären AR, der Größe, der Membranoberfläche und der Membrankrümmung18ab. Wenn neonatale Ratten-CMs auf Substraten plattiert wurden, die gemustert wurden, um die Zellen in einer bestimmten Länge zu beschränken: Breite AR, zeigten sie die beste kontraktile Funktion, wenn die Verhältnisse ähnlich wie die Zellen in einem gesunden Erwachsenenherz waren. Im Gegensatz dazu schnitten sie schlecht ab, wenn die Verhältnisse denen von Myozyten in versagenden Herzen ähnlich waren19. In den frühen Stadien der Hypertrophie werden die Zellen breiter, was sich in einer Zunahme des Querschnittsbereichs widerspiegelt. HF tritt in den späteren Stadien der Hypertrophie auf und Zellen erscheinen in der Regel ländiert. Daher ist es nicht verwunderlich, dass in vivo Rattenmodelle der chronischen Hypertrophie eine Zunahme der linken ventrikulären Myozytenlänge von etwa 30%20berichtet haben, aber erwachsene CMs aus transgenem Mausmodell, die akut mit hypertrophen Reizen in vitro behandelt wurden, zeigten ähnliche Erhöhungen der Zellbreite statt21.
Die einzellige RNA-Sequenzierung, die eine präzise Analyse des Transkriptoms einzelner Zellen ermöglicht, revolutioniert derzeit das Verständnis der Zellbiologie. Diese Technologie war die bevorzugte Methode, wenn es darum ging, die Frage zu beantworten, wie einzelne Zellformen die Genexpression beeinflussen. Wir verglichen einzelne Zellen mit verschiedenen Formen, insbesondere mit ARs von 1:1, 7:1 oder 11:1. Dies geschah durch Aussaat der neonatalen ventrikulären CMs der Neonatalen Ratte auf einen speziell entwickelten Chip, der mit den fibronectinbeschichteten Mikromustern2 mit definierten ARs von 1:1, 7:1 oder 11:1 gefüllt wurde. Die Mikromuster wurden mit Photolithographie-Technologie hergestellt. Die Mikromuster wurden mit Fibronectin beschichtet, umgeben von zytophober Oberfläche. Daher werden CMs die definierte AR von Mikromustern anbringen, verteilen und erfassen, indem sie ausschließlich auf dem Fibronectin-Substrat wachsen und gleichzeitig den zytophoben Bereich vermeiden. Die Mikromuster sind nicht in einem gut geformten Format. Stattdessen liegt der Fibronectinspiegel genau auf der gleichen Höhe des umgebenden zytophoben Bereichs. Dies bot ähnliche Bedingungen wie wachsende Zellen in einer Petrischale, da es keinen Stress von den umliegenden Wänden gibt. Darüber hinaus ist die Oberfläche von Mikromustern mit unterschiedlichen ARs gleich.
Es gab zwei besonders wichtige Aspekte des experimentellen Designs, die zur Verwendung von einzelliger RNA-Sequenzierung anstelle von Bulk-RNA-Sequenzierung führten. Erstens können nur wenige Prozente der Mikromuster von einer einzelnen Zelle belegt werden. Zweitens, manchmal besetzt eine einzelne Zelle die Mikromusteroberfläche nicht vollständig. Einzelne Zellen, die eine Mikromusteroberfläche vollständig abdecken, müssen für die einzellige RNA-Analyse ausgewählt werden. Da nur eine Untergruppe der plattierten Zellen auf einem Chip beide Kriterien erfüllte, war es nicht möglich, einfach den gesamten Chip zu versuchen und alle Zellen für die Massen-RNA-Sequenzierung zu sammeln. Qualifizierte Zellen mussten einzeln mit einem halbautomatischen Zellenwähler ausgewählt werden.
Es bleibt derzeit nicht bekannt, ob DIE CM-Form allein einen intrafunktionalen Einfluss auf das myokarde Syncytium hat. Der Hauptzweck der in diesem Papier vorgeschlagenen Methoden war die Entwicklung einer neuartigen Plattform, um zu untersuchen, ob die Zellform an sich einen Einfluss auf das Transkriptom17hatte. Obwohl sich In-vitro-Studien von In-vivo-Studien unterscheiden, bestand der Zweck dieser Studie darin, die Wirkung verschiedener Zellformen auf die Genexpression zu untersuchen, wobei zu berücksichtigen war, dass der Vergleich von Zellen mit verschiedenen Formen in vivo äußerst anspruchsvoll ist. Diese Experimente wurden von Kuo et al.19inspiriert, die einen ähnlichen Ansatz verfolgten und berichteten, dass sie Veränderungen der physiologischen Parameter aufgrund von Veränderungen in der Zellform beobachteten.
Protokoll
Alle Verfahren, an denen Tiere beteiligt waren, entsprachen den Vorschriften der Tierethikkommission des Karolinska Institutet, Stockholm, Schweden.
1. Mikrogemusterte Chip-Layout
- Verwenden Sie einen kundenspezifischen Chip (Tabelle der Materialien) ( Abbildung1A): ein 19,5 mm x 19,5 mm Deckblatt mit aktivierten Mikromustern, gedruckt durch Photolithographie auf Borosilikatglas.
HINWEIS: Diese Mikromuster sind von einem zytophoben Bereich umgeben. Daher kann eine gesäte Zelle nur an einem dieser Mikromuster anheften und wachsen und die AR dieses Mikromusters erfassen. Der Chip ist in drei Zonen unterteilt und jede Zone besteht aus Mikromustern mit einem bestimmten AR. Das Chip-Layout ist in Abbildung 1Bdargestellt. Die Geometrie der definierten ARs ist in Tabelle 1dargestellt. Vergrößerte fluoreszierende Bilder der verschiedenen Formen der Fibronectin-Mikromuster sind in den unteren Bildern in Abbildung 1Bdargestellt.
2. Beschichtung mikrogemusterter Chips
- Bereiten Sie 2x Beschichtungsproteinlösung für jeden Chip vor, indem Sie 80 g Fibronectin zu 2 ml Phosphat-gepufferter Saline (PBS-/-) hinzufügen.
- Einen Chip auf eine 35 mm Greiner Petrischale übertragen und sofort 2 ml PBS-/-hinzufügen. Dann 2 ml der 2x Beschichtungsproteinlösung hinzufügen.
- Inkubieren Sie die Chips bei Raumtemperatur für 2 h.
- Waschen Sie die Beschichtungslösung durch aufeinanderfolgende Verdünnungsschritte mit PBS-/-. Die Spanoberfläche sollte immer nass sein. Dann ersetzen Sie die PBS mit 2 ml Beschichtungsmedium und inkubieren bei 37 °C bis zur Aussaat zellen.
3. Isolierung von CMs
- Bereiten Sie das Beschichtungsmedium vor, indem Sie DMEM:M199 (4:1) mit 10% Pferdeserum, 4% fetalem Rinderserum, 2% HEPES (1 M) und 1% Penicillin/Streptomycin (10.000 U/ml)22ergänzen.
- Sezieren Sie das Gewebe aus dem linken Ventrikel von 2 Tage alten neonatalen Rattenherzen und übertragen Sie es auf eine 10 cm große Schale, die PBS enthält. Schneiden Sie das Gewebe in ca. 1 mm3 Stück.
- Übertragen Sie das geerntete Gewebe in ein Rotorkappenrohr, ausgestattet mit einem Rotor in der Kappe für die Gewebetrennung (Tabelle der Materialien). Lassen Sie das Gewebe absetzen und entfernen Sie dann vorsichtig den Überstand.
- 2,5 ml der Mischung aus Enzymmischung 1 und 2, mit dem Neonatal Heart Dissociation Kit (Materialtabelle) hergestellt, in die C-Röhre geben und die Kappe fest schließen.
- Legen Sie das Rotorkappenrohr auf die Hülse des Dissoziators, ausgestattet mit Heizungen (Abbildung 2A und B) (Materialtabelle). Führen Sie das Inkubationsprogramm 37C_mr_NHDK_1 (Abbildung 2C) aus, das etwa eine Stunde dauert.
- Während das Inkubationsprogramm läuft, bereiten Sie den PEB-Puffer mit 2 mM EDTA und 0,5% Rinderserumalbumin (BSA) in PBS, pH 7,2, vor und halten Sie ihn bei 4 °C.
- Trennen Sie nach Beendigung des Inkubationsprogramms das Rotorkappenrohr (Abbildung 2D) und fügen Sie 7,5 ml vorgewärmtes Beschichtungsmedium hinzu.
- Setzen Sie die Probe wieder auf, und filtern Sie die Zellsuspension mit einem 70-m-Sieb.
- Waschen Sie das Sieb mit weiteren 3 ml Beschichtungsmedium.
- Zentrifugieren Sie die Zellsuspension bei 600 x g für 5 min. Aspirieren Sie den Überstand vollständig.
- Setzen Sie das Zellpellet in 60 l kalten PEB-Puffer aus.
- Fügen Sie 20 L Neonatal Cardiomyozyten Isolation Cocktail(Materialtabelle) hinzu, die Mikron-Perlen enthalten, die auf Nicht-CMs abzielen.
- Fügen Sie 20 L Anti-Rot-Blut-Zell-Perlen (Tabelle der Materialien).
- Die Suspension mischen und bei 4 °C 15 min inkubieren.
- Fügen Sie 400 L PEB-Puffer hinzu.
- Tragen Sie die Zellsuspension auf die LD-Säule (Materialtabelle) auf, die vertikal in einen Magnetständer eingeführt und mit dem PEB-Puffer gut gewaschen wurde.
- Sammeln Sie unbeschriftete Zellen und waschen Sie die Säule mit 0,5 ml PEB-Puffer.
- 8 ml Beschichtungsmedium hinzufügen und die Zellsuspension in einen 75 cm2 unbeschichteten Kulturkolben geben und bei 37 °C für 1,5 h brüten. Die verbleibenden Nicht-CMs werden beginnen, der unbeschichteten Zellkultur zu entsprechen, und die Zellsuspension wird durch die CM-Population angereichert.
4. Zerpattern CMs
HINWEIS: Wir haben einzelne CMs mit ARs von 1:1, 7:1 oder 11:1 verglichen. Dies geschieht durch Aussaat der isolierten neonatalen Ratten-CMs auf einen speziell entwickelten Chip, der mit fibronectinbeschichteten Mikromustern mit definierten ARs von 1:1, 7:1 oder 11:1 gefüllt ist. Die Mikromuster wurden mit Fibronectin beschichtet, umgeben von zytophober Oberfläche. Daher werden CMs die definierte AR von Mikromustern anbringen, verteilen und erfassen, indem sie ausschließlich auf dem Fibronectin-Substrat wachsen. Mustern Sie die isolierten CMs nach den folgenden Schritten.
- Übertragen Sie die Zellsuspension in ein 15 ml-Rohr, zählen Sie die Zellen und verdünnen Sie sie auf eine Konzentration von 100.000 Zellen pro ml, indem Sie geeignetes Beschichtungsmedium hinzufügen.
- Fügen Sie 2 ml Zellsuspension auf den Chip, der bereits in 2 ml warmes Beschichtungsmedium in einer 35 mm Greiner Petrischale untergetaucht ist.
- Inkubieren Sie die Schale bei 37 °C mit 5%CO2, damit die CMs an den fibronectinbeschichteten Mikromustern befestigt werden und jeder CM die AR seines Substratmikromusters erwerben kann.
- Nach 18 h, überprüfen Sie den Chip. Wenn die meisten Zellen angebunden sind, lösen und entfernen Sie die Trümmer und abgestorbenen Zellen, die an die gemusterte Zelle angehängt sind. Nehmen Sie dies vor, indem Sie das Beschichtungsmedium entfernen und sanft PBS-/- tropfenweise hinzufügen, beginnend von der Mitte des Chips und sich dann zu den Seiten bewegen. Wiederholen Sie dies 2 Mal.
- Aspirieren Sie das PBS mit frischem Pflegemedium, indem Sie DMEM:M199 (4:1) mit 4% Pferdeserum, 4% fetalem Rinderserum, 2% HEPES (1 M) und 1% Penicillin/Streptomycin (10.000 U/ml) ergänzen.
5. Kommissionierung von klebenden CMs
ANMERKUNG: Nach einer Kultivierungsphase von 72 Stunden werden gemusterte Einzel-CMs aus ihren Fibronectin-Mikromustern mit einem halbautomatischen Zellwähler (Materialtabelle) (Abbildung 3) ausgewählt. Der Zellenwähler verwendet eine Software23, um die motorisierte Stufe zu steuern (Abbildung 3A). Zur Auswahl und Injektion der gemusterten neonatalen Ratten-CMs wird eine Mikrokapillare aus 70 m Glas(Abbildung 3B)verwendet. Der Zellwähler sortiert anhimierte Zellen, indem er ein Vakuum erzeugt und die Zellen durch Druck injiziert. Das Vakuum in Spritze Nummer 1 wird durch Ziehen der Spritze mit der Spritzenpumpe aufgebracht (Abbildung 3C). Der hydrostatische Druck basiert auf der Schwerkraft und wird durch Platzierung der Spritze Nummer 2 in einem Abstand von 87 cm über dem Mikroskoptisch induziert. Die Spritzen 1 und 2 werden jeweils über PTFE-Rohre mit ventilen 1 und 2 verbunden, die in die Steuereinheit eingebettet sind (Abbildung 3D). Die PTFE-Rohre sind komplett mit RNase-freiem Wasser gefüllt. Die entnommene Einzelzelle wird dann in das Polymerase-Kettenreaktionsrohr (PCR) injiziert, das 3,55 l Lysepuffer enthält.
- Nach einer Kultivierungszeit von 72 Stunden das alte Medium entfernen und die Spanoberfläche vorsichtig mit warmem DPBS-/-spülen.
- Halten Sie die Schale flach und aspirieren Sie das alte Medium vorsichtig mit einer 1000-L-Pipette von einer Seite der Schale. Stellen Sie sicher, dass der Chip zu jeder Zeit nass bleibt.
- Fügen Sie 2 ml DPBShinzu -/- entfernen Sie vorsichtig und tropfenweise die meisten abgestorbenen Zellen, die an den gemusterten Zellen befestigt sind, beginnend mit der Mitte des Chips und bewegen sich dann zu seinen Seiten.
- Aspirieren Sie die meisten DPBS, um so viele freistehende schwimmende Zellen wie möglich zu entfernen, beginnend von der Mitte des Chips und dann zu den Seiten bewegen.
- Wiederholen Sie die Schritte 5.1.2 und 5.1.3 erneut.
- Verwenden Sie abgewinkelte Zangen, um die Kante des Chips zu greifen und übertragen Sie ihn sofort auf eine neue sterile 35 mm Greiner Petri Schale, um die Anzahl der schwimmenden Zellen während der Kommissionierung zu reduzieren.
- 1,5 ml DPBS sofort hinzufügen-/- damit der Chip nicht austrocknet.
- Fügen Sie 1,5 L Vibrant Dye Cycle grün hinzu, um die Kerne der lebenden Zellen zu visualisieren.
- Legen Sie den Chip in die Mitte der Greiner Petrischale mit der Spitze der Zange.
- Legen Sie eine Kammer (Materialtabelle) über den Chip. Die Kammer fixiert den Chip an der Unterseite der Schale, ohne den Zugang zu den Zellmustern zu blockieren.
- Montieren Sie die Greiner Petrischale auf den Tellerhalter der Zellpflücker-Bühne und legen Sie die Magnetkappe ein.
- Kalibrieren Sie die automatische Injektion.
- Suchen Sie das Fadenkreuz, das auf der motorisierten Bühne in der Mitte des Bildes im Live View-Fenster eingraviert ist.
- Konzentrieren Sie sich auf das Fadenkreuz und wählen Sie im Scan- und Sortierfenster die Taste Kalibrierung für die automatisierte Injektion aus.
- Ersetzen Sie die DPBS-/- mit 1,5 ml DPBS/Trypsin-/- (1:1), während die Schale auf der Bühne ist, um die Zellen aus dem Fibronectin zu lösen, so dass ein flüssiges Vakuum verwendet werden kann, um die Zellen zu wählen.
- Scannen Sie den gesamten Chip mit der Registerkarte Scannen im Scan- und Sortierfenster. Suchen Sie die obere linke Ecke des Chips im Sichtfeld und klicken Sie auf Aktuelle Mikroskopposition in der oberen linken Eckreihe.
- Bewegen Sie dann die motorisierte Bühne in die untere rechte Ecke des Chips. Fokussieren Sie das Mikroskop und klicken Sie auf Aktuelle Mikroskopposition in der unteren rechten Eckreihe abrufen.
- Klicken Sie auf die Schaltfläche Die schärfste Ebene einstellen und im geöffneten Fenster auf den Weg in die obere rechte Ecke. Fokussieren Sie das Mikroskop und klicken Sie auf Gehe zur unteren linken Ecke und stellen Sie den Fokus ein. Wenn Sie fertig sind, klicken Sie auf die Schaltfläche Fertig stellen und starten Sie den Scanvorgang.
- Wenn der Scanvorgang beendet wird, wechseln Sie zur Registerkarte Analysieren, und wählen Sie die einzelnen Zellen aus, die die Studienkriterien bestehen.
- Stellen Sie sicher, dass sich die Mikrokapillare aus Glas in der Mitte der Live-Ansicht des Mikroskops befindet.
- Steuern Sie die Spritzenpumpe über das Pumpenfenster der Software. Erstellen Sie ein Vakuum, indem Sie 4 ml aus einer 50 ml Spritze Nummer 1 mit einem Durchmesser von 27 mm abziehen.
- In der Registerkarte Sortieren
- Stellen Sie die Einspritzparameter der Ventile ein. Es wurde berechnet, dass das Injektionsvolumen, das eine kommissionierte Einzelzelle lieferte, 1 l betrug, wenn Ventil 2 für 120 Millisekunden geöffnet und dann Ventil 1 nach einem Zeitraffer von 200 Millisekunden für 20 Millisekunden geöffnet wurde. Aufgrund der Elastizität der Rohre, öffnen Sie Ventil 1, um die Injektion des Durchflusses zu stoppen.
- Stellen Sie die Aufnahmeparameter der Ventile ein. Es wurde berechnet, dass, wenn Ventil 1 für 20 Millisekunden geöffnet wurde und dann Ventil 2 für 10 Millisekunden geöffnet wurde, nach einem Zeitraffer von 10 Millisekunden, die meisten gemusterten Zellen aufgenommen werden können. Dies liegt daran, dass ihre Fibronectin-Bindungen nach der Behandlung durch Trypsin gelockert wurden.
- Klicken Sie auf die Schaltfläche Pfad berechnen. Die Software berechnet den schnellsten Pfad von Zelle zu Zelle, um die ausgewählten Zellen im gesamten Chip aufzunehmen und zu injizieren.
- Konzentrieren Sie das Mikroskop auf eine gemusterte Zelle auf der Spanoberfläche.
- Mit dem Joystick die Mikrokapillare vorsichtig nach unten bewegen, so dass das schärfste Bild der Spitze der Mikrokapillare erhalten werden kann, ohne die Zelle zu berühren.
- Klicken Sie im Abschnitt Micropipette-Offset auf die Schaltfläche Festlegen. Es wird ein neues Fenster mit dem Mikrokapillarquerschnitt angezeigt. Klicken Sie auf die genaue Mitte der Kapillare. Die Software zeichnet dann den Spitzenversatz der Kapillare in den x-, y- und z-Koordinaten auf.
- Starten Sie die Sortierung mit der Schaltfläche Sortieren starten.
Ergebnisse
Gewebe wurde aus dem linken Ventrikel der 2 Tage alten neonatalen Rattenherzen seziert und in einzelne Zellen geteilt. Dann wurden die angereicherten CMs auf einem Chip mit Fibronectin-Mustern mit deutlichen ARs gesät. Nach 72 Stunden Kultivierung wurde das Medium durch 1:1000 Vibrant Dye Cycle green in DPBS-/- für 2 min ersetzt, um die Kerne der lebenden Zellen zu visualisieren. Als nächstes wurden die Zellen mit DPBS-/-/trypsin (1:1) behandelt, um die Zellen aus dem Fibronectin zu lösen, so dass ein flüssiges Vakuum verwendet werden konnte, um die Zellentnahme zu erleichtern. In der Zwischenzeit wurde der gesamte Chip mit einer Vergrößerung von 10x gescannt, mit einem invertierten Mikroskop, das mit dem Zellpflücker verbunden ist. Dies wurde durchgeführt, bevor die Zellen durch Trypsin-Behandlung gerundet wurden. Die qualifizierten Zellen wurden basierend auf dem gescannten Bild ausgewählt, und ihre Koordinaten wurden in der Zellenauswahlsoftware gespeichert. Die Mikromuster wurden nur ausgewählt, wenn sie eine mononukleierte Einzelzelle enthielten und nur, wenn die Zelle ihr Fibronectin-Mikromuster vollständig bedeckte. Der Zellsortierer wählte die ausgewählten Zellen nacheinander aus und jede einzelne Zelle, die erfolgreich entnommen wurde, wurde sofort in eine einzelne PCR-Röhre injiziert und auf die Mikroskopstufe gelegt. Jede PCR-Röhre enthielt 3,55 l Lysepuffer (Tabelle 2). Der Sortiervorgang, der mit dem Entfernen der Medien begann, wurde innerhalb von 40 Minuten abgeschlossen. Die komplementäre Desoxyribonukleinsäuresynthese (cDNA), PCR-Vorverstärkung und -Reinigung wurden auf Basis des Smart-Seq2-Protokolls24 (Tabelle 3) an den lysierten Einzelzellen durchgeführt. Die Qualität der gereinigten cDNA wurde durch einen automatisierten Elektrophorese-Analysator überprüft. Das Elektropherogramm der vorverstärkten cDNA einer ausgewählten Einzelzelle ist in Abbildung 4dargestellt. Die RNA-Seq-Bibliotheken wurden nach dem Smart-Seq2-Protokoll24erstellt.
Um die sarcomere Struktur der gemusterten CMs zu beobachten, wurden die gemusterten CMs mit sarkomischem α-Actinin-Antikörper niert. Die Zellen wurden mit Esel Anti-Maus IgG Alexa Fluor 488 1:800 für 1 Stunde bei Raumtemperatur für die sekundäre Färbung inkubiert. Die Kerne wurden mit 1 g/ml DAPI gefärbt. Immunfluoreszenzbilder wurden mit einem invertierten konfokalen Mikroskop mit einem 63-fachen Öl-Immersion (NA 1.4) Objektiv(Abbildung 5)aufgenommen.

Abbildung 1: Layout des Chips mit Fibronectin-Mikromustern.
(A) Bild des kundenspezifischen Chips. Der Chip ist ein 19,5 mm x 19,5 mm Coverslip mit Fibronectin-Mikromustern, gedruckt durch Photolithographie auf einem Borosilikatglas. (B) Chip-Layout. Der Chip ist in drei Zonen unterteilt und jede Zone besteht aus Fibronectin-Mikromustern mit spezifischem AR. Fluoreszierende Bilder verschiedener Formen von Fibronectin-Mikromustern werden in vergrößerter Ansicht für jede Zone dargestellt. Diese Zahl wurde von Haftbaradaran Esfahani et al.2von "ergänzendem Material 1" geändert, das unter http://creativecommons.org/licenses/by/4.0/ verwendet wurde. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 2: Dissoziator mit Heizungsgerät.
(A) Das gesamte Dissoziatorinstrument, das für die vollautomatische Dissoziation von 2 Tage alten neonatalen Ratte links Ventrikeln verwendet wird. (B) Heizeinheit. (C) Rotorkappenrohr. (D) Gebrauchsfertige Programme für einen vollautomatischen Workflow der Gewebetrennung. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 3: Zellpflückergerät.
(A) Vergrößerte Ansicht der motorisierten Stufe des Zellenpflückers. Ein Petri-Schale-Halter und 80 Löcher für 10 PCR-Streifen und ein Loch zur Kalibrierung simnetend ist auf der Bühne eingebettet. (B) Vergrößerte Ansicht einer Glasmikrokapillare. (C) Die Spritzenpumpe. (D) Das Steuergerät, das das Öffnungs- und Schließzeitfenster der Ventile 1 und 2 steuert, ist im Steuergerät montiert. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 4: Das Elektropherogramm der vorverstärkten cDNA einer entnommenen Einzelzelle.
19 PCR-Zyklen der Vorverstärkung wurden verwendet, um 15 l von 1 ng/l gereinigte cDNA-Ausbeute zu erhalten. Es wird ein klares Band in gelähnlicher Densitometrie-Plot beobachtet, das dem Peak bei 1852 bp im Elektropherogramm entspricht. Die durchschnittliche Größe der Fragmente beträgt 1588 bp. Darüber hinaus weist die geringe Anzahl von Fragmenten, die kürzer als 300 bp sind, auf eine gute cDNA-Bibliothek hin. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 5: Immunfluoreszenzfärbung von α-Actinin-Sarkomischer Struktur (grün) und Kern (blau) von gemusterten CMs mit unterschiedlichen ARs.
Das Chromatin wurde von DAPI gefärbt. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
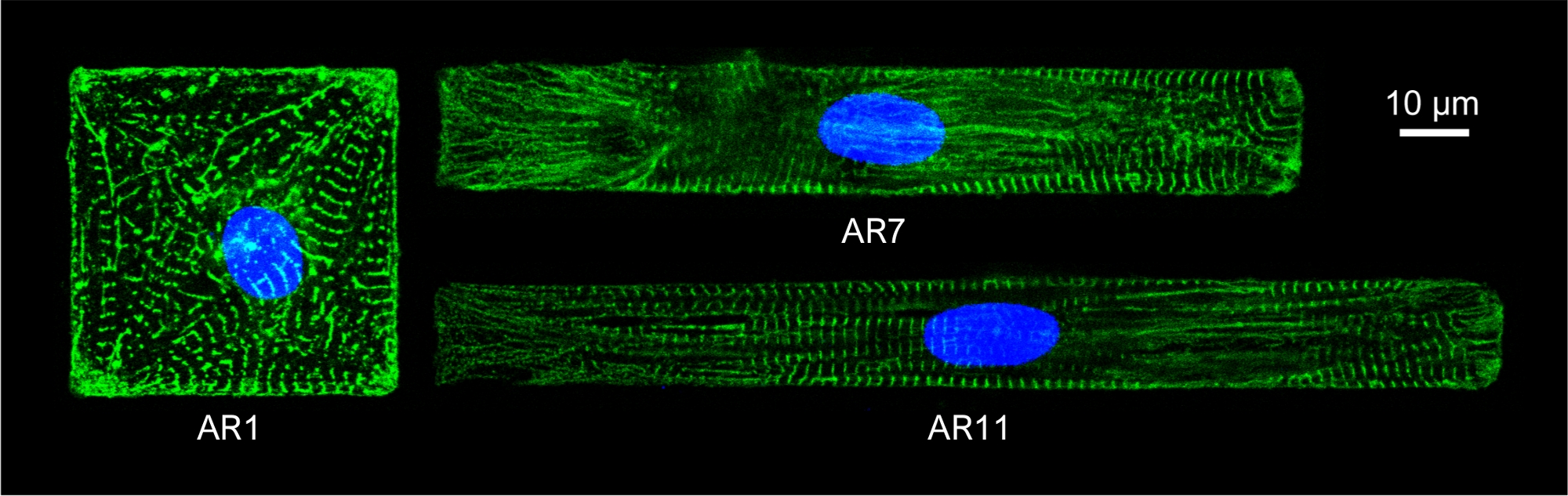{kind=link}
| Morphotyp | Ar | Länge (m) | Breite (m) | Fibronectin-Bereich (m2) |
| AR1 | 1:1 | 47 | 47 | 2209 |
| AR7 | 7:1 | 126 | 18 | 2268 |
| AR11 | 11:1 | 155 | 14 | 2170 |
Tabelle 1: Geometrie gemusterter CMs.
| Komponente | Volumen (L) |
| Nukleasfreies Wasser | 0.65 |
| (0,4% Vol./Vol) Triton X-100 | 1.8 |
| dNTP-Mischung (25 mM) | 0.8 |
| RNase-Inhibitor (40 U l-1) | 0.1 |
| Oligo-dT30VN-Oligonukleotide (100 m) | 0.1 |
| ERCC RNA Spike-In Mix (2,5 x 105 Verdünnung) | 0.1 |
| Injizierte Einzelzelle | 1 |
| Gesamtvolumen | 4.55 |
Tabelle 2: Benutzerdefinierter Lysepuffer für einzelne Zellen.
| Komponente | Volumen (L) |
| Superscript II First-Strang-Puffer (5x) | 2 |
| DTT (100 mM) | 0.5 |
| Betain (5 M) | 2 |
| Mgcl2 (1 M) | 0.1 |
| RNase-Inhibitor (40 Ul -1) | 0.25 |
| Hochschrift II Reverse Transkriptase (200 U-L -1) | 0.5 |
| TSO (100 m) | 0.1 |
| Gesamtvolumen | 5.45 |
Tabelle 3: Reverse Transkriptions-Mix (RT) für eine RT-Reaktion zur Synthese von cDNA mit erstem Strang aus dem Lysat eines einzelnen CM.
Diskussion
Diese Studie verwendete eine einzellige RNA-Sequenzierung, eine neuartige und leistungsstarke Technologie, die das Transkriptom einzelner Zellen detektieren kann. Es wurde mit einem innovativen Ansatz zur Kultivierung einzelner CMs kombiniert, so dass sie unterschiedliche ARs anwandten, die andernfalls nur in vivo hätten beobachtet werden können.
Die Studie hatte einige Einschränkungen. Zum Beispiel mussten neonatale CMs verwendet werden, um verschiedene Morphotypen zu erzeugen, da es außerordentlich schwierig ist, genügend vitale Erwachsenen-CMs für 72 Stunden in definierten Formen zu kultitogen. Darüber hinaus wurden CMs 72 Stunden ex vivo kultiviert, was sich auf das Genexpressionsmuster ausgewirkt haben könnte. Diese Kultivierung war jedoch notwendig, damit die Zellen spezifische Morphotypen bilden konnten. Darüber hinaus wurden nur einzelne Zellen, die mononukleiert waren und das Fibronectin-Mikromuster vollständig bedeckten, für die Sortierung ausgewählt. Gemusterte Zellen auf jedem Chip müssen nur in einer Sortierungsrunde sortiert werden. Schließlich wurden etwa 50 Zellen, das ist etwa ein Drittel der ausgewählten Zellen erfolgreich von jedem Chip aufgenommen. Es gibt zwei Gründe, die die Anzahl der erfolgreich ausgewählten Zellen einschränken. Erstens waren einige Zellen zu eng am Fibronectin-Muster befestigt und der Pickup-Flow war nicht gewaltsam genug, um sie erfolgreich aufzunehmen. Zweitens, durch die Trypsin-Behandlung, die Anhaftung zwischen einigen Zellen und Fibronectin wurde zu locker. Folglich wurden diese Zellen von ihren Fibronectin-Mikromustern weggedrückt, als sich die Mikrokapillare ihnen näherte, und sie wurden nicht aufgenommen. Die Autoren behaupten nicht, dass dieses Setup das gleiche wie eine in vivo-Umgebung ist, aber es erwies sich als ein praktikabler Ansatz, um die Forschungsfrage zu beantworten.
Die vorgeschlagene Methode ist auf verschiedene Zelltypen anwendbar (z. B. für hiPS-CMs). Die folgenden Faktoren sollten jedoch optimiert werden, um andere Zelltypen zu untersuchen. Geeignete ECM-Klebstoffmoleküle zur Befestigung des spezifischen Zelltyps sollten zur Beschichtung der Mikromuster verwendet werden. Die Geometrie der Mikromuster sollte entsprechend der Studienfrage und dem Zelltyp geändert werden. Die Kultivierungszeit kann auf der Grundlage der Studienfrage geändert werden. Das Ablösungsreagenz und seine Inkubationszeit sollten exakt für den Studienzelltyp optimiert werden. Zum Beispiel kann Accutase anstelle von TryplE zur Ablösung embryonaler und neuronaler Stammzellen verwendet werden. Die Öffnungszeitparameter der Ventile sollten überprüft werden, um Zellen erfolgreich, aber schonend zu pflücken. Zusammenfassend haben wir eine neuartige Plattform entwickelt, um die Zellform zu untersuchen, die eine wertvolle Ressource für Forscher auf diesem Gebiet darstellen kann. In diesem Zusammenhang haben wir einen experimentellen Ansatz entwickelt, der in vitro charakteristische Formen nachahmte, die CM in vivo durch hämodynamische Einschränkungen auferlegt wurden, um das Zusammenspiel zwischen zellulärer Architektur und Genexpression zu identifizieren. Wir berichten auch über die Entwicklung einer neuartigen Plattform zur Untersuchung von HF in vitro und die Identifizierung der Zellform als einen starken Determinanten der Genexpression. Dies ist eine neuartige Beobachtung mit weitreichenden Auswirkungen auf Biologie und Medizin.
Offenlegungen
nichts.
Danksagungen
nichts.
Materialien
| Name | Company | Catalog Number | Comments |
| 2100 Bioanalyzer Instrument | Agilent Technologies | G2939BA | Automated electrophoresis analyzer |
| Anti-Red Blood Cell MicroBeads | Miltenyi Biotec | 130-109-681 | |
| Axio Observer microscope | Zeiss | Z1 | Inverted microscope |
| Betaine solution (5 M) | Sigma-Aldrich, MERCK | B0300 | |
| CellSorter | CELLSORTER | https://www.singlecellpicker.com/ | Cell picker |
| CYTOOchamber | CYTOO | 30-010 | Custom-designed chip |
| CYTOOchip | CYTOO | 10-950-00-18 | Chamber |
| DMEM | Thermo Fisher Scientific | 31966-021 | high glucose, GlutaMAX Supplement |
| dNTP mix (25 mM) | Thermo Fisher Scientific | R1122 | |
| Donkey Anti-Mouse IgG Alexa Fluor 488 | Abcam PLC | ab150105 | |
| DTT (100 mM) | Thermo Fisher Scientific | 18064071 | |
| ERCC RNA Spike-In Mix | Thermo Fisher Scientific | 4456740 | |
| Fetal Bovine Serum | Thermo Fisher Scientific | 10082-147 | |
| Fibronectin | Sigma-Aldrich, MERCK | F4759 | |
| gentleMACS C Tube | Miltenyi Biotec | 130-093-237 | Rotor-cap tube |
| gentleMACS Octo Dissociator with Heaters | Miltenyi Biotec | 130-096-427 | Dissociator with heater |
| Greiner CELLSTAR Petri dish | Sigma-Aldrich, MERCK | P6987 | |
| HEPES (1 M) | Thermo Fisher Scientific | 15630-056 | |
| Horse Serum | Sigma-Aldrich, MERCK | H0146 | |
| LD column | Miltenyi Biotec | 130-042-901 | |
| Medium 199 | Thermo Fisher Scientific | 31150-022 | |
| NE-1000 syringe pump | New Era Pump Systems | NE-1000 | Syringe pump |
| Neonatal Cardiomyocyte Isolation Cocktail, rat | Miltenyi Biotec | 130-105-420 | |
| Neonatal Heart Dissociation Kit, mouse and rat | Miltenyi Biotec | 130-098-373 | |
| Oligo-dT30VN oligonucleotides | IDT Technology | 5′–AAGCAGTGGTATCAACGCAGAGTACT30VN-3′ | |
| RNAse inhibitor (40 U µL-1) | Clontech | 2313A | |
| sarcomeric α-actinin | Sigma-Aldrich, MERCK | EA-53 | |
| SP8 confocal microscope | Leica Microsystems | SP8 | Confocal microscope |
| Superscript II first-strand buffer (5x) | Thermo Fisher Scientific | 18064071 | |
| Superscript II reverse transcriptase (200 U µL-1) | Thermo Fisher Scientific | 18064071 | |
| Triton X-100 | Sigma-Aldrich, MERCK | T9284 | |
| TryplE Express enzyme, no phenol red | Thermo Fisher Scientific | 12604013 | |
| TSO (100 µM) | QIAGEN | 5′-AAGCAGTGGTATCAACGCAGAGTACATrGrG+G-3′ | |
| Vibrant Dye Cycle green | Thermo Fisher Scientific | V35004 |
Referenzen
- Heineke, J., Molkentin, J. D. Regulation of cardiac hypertrophy by intracellular signalling pathways. Nature Reviews Molecular Cell Biology. 7 (8), 589-600 (2006).
- Haftbaradaran Esfahani, P., et al. Cell shape determines gene expression: cardiomyocyte morphotypic transcriptomes. Basic Research in Cardiology. 115 (1), 7(2019).
- Kontrogianni-Konstantopoulos, A., Benian, G., Granzier, H. Advances in Muscle Physiology and Pathophysiology 2011. Journal of Biomedicine and Biotechnology. 2012, 930836(2012).
- Hill, J. A., Olson, E. N. Cardiac plasticity. New England Journal of Medicine. 358 (13), 1370-1380 (2008).
- Bray, M. A., Sheehy, S. P., Parker, K. K. Sarcomere alignment is regulated by myocyte shape. Cell Motility and the Cytoskeleton. 65 (8), 641-651 (2008).
- Benjamin, I. J., Schneider, M. D. Learning from failure: congestive heart failure in the postgenomic age. Journal of Clinical Investigation. 115 (3), 495-499 (2005).
- Opie, L. H., Commerford, P. J., Gersh, B. J., Pfeffer, M. A. Controversies in ventricular remodelling. Lancet. 367 (9507), 356-367 (2006).
- Braunwald, E. Cardiomyopathies: An Overview. Circulation Research. 121 (7), 711-721 (2017).
- Knoll, R., et al. The cardiac mechanical stretch sensor machinery involves a Z disc complex that is defective in a subset of human dilated cardiomyopathy. Cell. 111 (7), 943-955 (2002).
- Frangogiannis, N. G. The Extracellular Matrix in Ischemic and Nonischemic Heart Failure. Circulation Research. 125 (1), 117-146 (2019).
- Marian, A. J., Braunwald, E. Hypertrophic Cardiomyopathy: Genetics, Pathogenesis, Clinical Manifestations, Diagnosis, and Therapy. Circulation Research. 121 (7), 749-770 (2017).
- Mozaffarian, D., Caldwell, J. H. Right ventricular involvement in hypertrophic cardiomyopathy: a case report and literature review. Clinical Cardiology. 24 (1), 2-8 (2001).
- Olsson, M. C., Palmer, B. M., Stauffer, B. L., Leinwand, L. A., Moore, R. L. Morphological and functional alterations in ventricular myocytes from male transgenic mice with hypertrophic cardiomyopathy. Circulation Research. 94 (2), 201-207 (2004).
- Toepfer, C. N., et al. Hypertrophic cardiomyopathy mutations in MYBPC3 dysregulate myosin. Science Translational Medicine. 11 (476), (2019).
- McKenna, W. J., Maron, B. J., Thiene, G. Classification, Epidemiology, and Global Burden of Cardiomyopathies. Circulation Research. 121 (7), 722-730 (2017).
- Schultheiss, H. P., et al. Dilated cardiomyopathy. Nature Reviews Disease Primers. 5 (1), 32(2019).
- Knoll, R. A role for membrane shape and information processing in cardiac physiology. Pflugers Arch. 467 (1), 167-173 (2015).
- Rangamani, P., et al. Decoding information in cell shape. Cell. 154 (6), 1356-1369 (2013).
- Kuo, P. L., et al. Myocyte shape regulates lateral registry of sarcomeres and contractility. American Journal of Pathology. 181 (6), 2030-2037 (2012).
- Gerdes, A. M., Onodera, T., Wang, X., McCune, S. A. Myocyte remodeling during the progression to failure in rats with hypertension. Hypertension. 28 (4), 609-614 (1996).
- Kehat, I., et al. Extracellular signal-regulated kinases 1 and 2 regulate the balance between eccentric and concentric cardiac growth. Circulation Research. 108 (2), 176-183 (2011).
- Sreejit, P., Kumar, S., Verma, R. S. An improved protocol for primary culture of cardiomyocyte from neonatal mice. In Vitro Cellular & Developmental Biology - Animal. 44 (3-4), 45-50 (2008).
- Kornyei, Z., et al. Cell sorting in a Petri dish controlled by computer vision. Scientific Reports. 3, 1088(2013).
- Picelli, S., et al. Full-length RNA-seq from single cells using Smart-seq2. Nature Protocols. 9 (1), 171-181 (2014).
- Schmick, M., Bastiaens, P. I. H. The interdependence of membrane shape and cellular signal processing. Cell. 156 (6), 1132-1138 (2014).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten