
Method Article
Un approccio per studiare la trascritomica dipendente dalla forma a un singolo livello cellulare
In questo articolo
Riepilogo
Questo articolo presenta metodi per coltivare miociti cardiaci con forme diverse, che rappresentano diverse patologie, e ordinare questi miociti cardiaci aderenti in base alla loro morfologia a un singolo livello cellulare. La piattaforma proposta fornisce un nuovo approccio all'elevata produttività e allo screening farmacologico per diversi tipi di insufficienza cardiaca.
Abstract
Diversi tipi di ipertrofia cardiaca sono stati associati ad un aumento del volume di miociti cardiaci (CM), insieme a cambiamenti nella morfologia cm. Mentre gli effetti del volume cellulare sull'espressione genica sono ben noti, gli effetti della forma cellulare non sono ben compresi. Questo articolo descrive un metodo che è stato progettato per analizzare sistematicamente gli effetti della morfologia CM sull'espressione genica. Descrive in dettaglio lo sviluppo di una nuova strategia di cattura a cella singola che viene poi seguita dal sequenziamento dell'mRNA a cella singola. È stato anche progettato un chip micropatterned, che contiene 3000 micropattern di fibronecina di forma rettangolare. Ciò consente di far crescere le macchine virtuali in proporzioni di lunghezza/larghezza distinte (AR), corrispondenti a diversi tipi di insufficienza cardiaca (HF). Il documento descrive anche un protocollo che è stato progettato per raccogliere singole celle dal loro modello, utilizzando un selettore di celle micro-pipettante semi-automatizzato, e iniettarle singolarmente in un buffer di lisi separato. Ciò ha permesso di profilare i trascrittami di singole MACCHINE con morfotipi geometrici definiti e caratterizzarli secondo una gamma di condizioni normali o patologiche: cardiomiopatia ipertrofica (HCM) o cardiomiopatia post-carico/concentrica rispetto alla cardiomiopatia dilatata (DCM) o precarica/eccentrica. In sintesi, questo documento presenta metodi per la coltivazione di MACCHINE con forme diverse, che rappresentano diverse patologie, e lo smistamento di queste MACCHINE aderenti in base alla loro morfologia a livello di singola cellula. La piattaforma proposta fornisce un nuovo approccio all'elevata produttività e allo screening farmacologico per diversi tipi di HF.
Introduzione
Secondo l'Organizzazione Mondiale della Sanità, le malattie cardiovascolari (CVD) sono una delle principali cause di morbilità e mortalità in tutto il mondo. La CVD influisce in modo drammatico sulla qualità della vita delle persone e ha un enorme impatto socioeconomico. Le cardiomiopatie, come HCM e DCM, sono disturbi primari del muscolo cardiaco e le principali cause di HF sono state associate ad alta morbilità e mortalità. Ci sono molte cause di HF, compresi gli effetti ambientali, come infezioni ed esposizione a tossine o alcuni farmaci8. HF può anche essere causato da predisposizione genetica, vale a dire mutazioni9. Si ritiene che i cambiamenti nella composizione genetica che influenzano molecole di matrice extracellulare (ECM), integrine o proteine citoscheletriche potrebbero essere responsabili di compromissione della meccanosensazione e vari tipi di malattie cardiache10.
La caratteristica principale dell'HCM è l'ipertrofia inspiegabile del ventricolosinistro 11, e talvolta del ventricolodestro 12, e questo spesso presenta un coinvolgimento predominante del setto interventricolare. L'HCM è anche caratterizzato da disfunzione diastolica e disordine di miocita e fibrosi13. Nella maggior parte dei casi, l'apparato contrattile del cuore è influenzato da mutazioni nelle proteine sarcomeriche, portando ad una maggiore contrattilità dei miociti14. Al contrario, il DCM è caratterizzato dalla dilatazione di uno o entrambi i ventricoli e ha un'eziologia familiare nel 30-50% dei casi15. DCM influisce su una vasta gamma di funzioni cellulari, portando a una contrazione compromessa dei miociti, morte cellulare e riparazione fibrotica16.
La genetica ha dimostrato che alcuni tipi di mutazioni costringono singole MACCHINE ad adottare caratteristiche di forma specifiche durante hcm3, vale a dire cellule a forma quadrata con un AR di lunghezza:larghezza che è quasi uguale a 1:14 (AR1). Lo stesso vale per DCM, con cellule allungate con un AR che è quasi uguale a 11:1 (AR11). Inoltre, HF può essere causato da un aumento del carico post-carico (ad esempio, in ipertensione). In questi casi, le richieste emodinamiche costringono le MACCHINE ad assumere forme quadrate, secondo la legge di Laplace, e l'AR cambia da 7:15 (AR7) a 1:16,7. HF può anche essere causato da un aumento del precarico (ad esempio, in condizioni che portano a sovraccarico di volume). Quando ciò accade, i vincoli biofisici costringono le macchine virtuali ad allungarsi e l'AR cambia da 7:1 a 11:1.
L'attività di segnalazione alle membrane dipende dai parametri globali della geometria cellulare, come l'AR cellulare, le dimensioni, la superficie della membrana e la curvatura della membrana18. Quando le RIC del ratto neonatale sono state placcate su substrati che sono stati modellati per vincolare le cellule in un AR specifico di lunghezza: larghezza, hanno dimostrato la migliore funzione contrattile quando i rapporti erano simili alle cellule in un cuore adulto sano. Al contrario, hanno funzionato male quando i rapporti erano simili a quelli dei miociti nei cuori che falliscono19. Nelle prime fasi dell'ipertrofia, le cellule diventano più ampie, come riflesso da un aumento dell'area della sezione trasversale. L'HF si verifica nelle fasi successive dell'ipertrofia e le cellule appaiono tipicamente allungate. Pertanto, non sorprende che i modelli di ratti in vivo di ipertrofia cronica abbiano riportato un aumento della lunghezza del miocita ventricolare sinistro di circa il 30%20, ma le MACCHINE adulte del modello di topo transgenico che sono state acutamente trattate con stimoli ipertrofici in vitro hanno dimostrato aumenti simili della larghezza delle celluleinvece di 21.
Il sequenziamento dell'RNA a cella singola, che consente un'analisi precisa del trascritoma di singole cellule, sta attualmente rivoluzionando la comprensione della biologia cellulare. Questa tecnologia era il metodo preferito quando si trattava di rispondere alla domanda su come le singole forme cellulari influenzavano l'espressione genica. Abbiamo confrontato singole celle con forme diverse, in particolare con AR di 1:1, 7:1 o 11:1. Questo è stato fatto seminando le MACCHINE ventricolari del ratto neonatale su un chip appositamente progettato riempito con i micropattern rivestiti di fibronectina2 con AR definiti di 1:1, 7:1 o 11:1. I micropattern sono stati fabbricati utilizzando la tecnologia della fotolitografia. I micropattern erano rivestiti di fibronectina, circondati da superficie citofobica. Pertanto, le macchine virtuali attaccheranno, diffonderanno e cattureranno l'AR definito dei micropattern crescendo esclusivamente sul substrato di fibronectina, evitando al contempo l'area citofobica. I micropattern non sono in un formato ben modellato. Invece, il livello di fibronectina è esattamente alla stessa altezza dell'area citofobica circostante. Ciò ha fornito condizioni simili alle cellule in crescita in una piastra di Petri, in quanto non c'è stress dalle pareti circostanti. Inoltre, la superficie dei micropattern con AR diversi è uguale.
C'erano due aspetti particolarmente importanti del progetto sperimentale, che portarono all'uso del sequenziamento dell'RNA a singola cella invece del sequenziamento dell'RNA sfuso. In primo luogo, solo poche percentuali dei micropattern possono essere occupate da una singola cellula. In secondo luogo, a volte una singola cella non occupa completamente la superficie del micropattern. Le singole celle che coprono completamente una superficie micropattern devono essere selezionate per l'analisi dell'RNA a singola cella. Poiché solo un sottogruppo delle cellule placcate su un chip soddisfaceva entrambi i criteri, non era possibile provare semplicemente l'intero chip e raccogliere tutte le cellule per il sequenziamento dell'RNA in blocco. Le celle qualificate dovevano essere prelevate singolarmente utilizzando una selezione celle semi-automatizzata.
Attualmente non si sa se la forma CM, di per sé, abbia un impatto intra-funzionale sul sincizio miocardico. Lo scopo principale dei metodi proposti in questo documento era quello di sviluppare una nuova piattaforma per studiare se la forma cellulare di per sé avesse un impatto sul trascrittame17. Sebbene gli studi in vitro siano diversi dagli studi in vivo, lo scopo di questo studio era quello di studiare l'effetto delle diverse forme cellulari sull'espressione genica, tenendo presente che confrontare cellule con forme diverse in vivo è estremamente impegnativo. Questi esperimenti sono stati ispirati da Kuo etal.
Protocollo
Tutte le procedure relative agli animali erano conformi alle norme del comitato etico per gli animali del Karolinska Institutet di Stoccolma, Svezia.
1. Layout di chip micro-modellato
- Utilizzare un chip progettato su misura (Tabella dei materiali) ( Figura1A): un coverslip da 19,5 mm x 19,5 mm con micropattern attivati, stampato per fotolitografia su vetro borosilicato.
NOTA: Questi micropattern sono circondati da un'area citofobica. Pertanto, una cellula seminata può attaccare e crescere solo su uno di questi micropattern e catturare l'AR di quel micropattern. Il chip è diviso in tre zone e ogni zona è costituita da micropattern con un AR specifico. Il layout del chip è illustrato nella figura 1B. La geometria degli AR definiti è presentata nella tabella 1. Immagini fluorescenti ingrandite delle diverse forme dei micropattern della fibronecina sono mostrate nelle immagini inferiori nella figura 1B.
2. Rivestimento di trucioli micropatterned
- Preparare una soluzione proteica di rivestimento 2x per ogni truciolo aggiungendo 80 μg di fibronectina a 2 mL di salina tamponata da fosfati (PBS-/-).
- Trasferire un chip in una piastra di Petri Greiner da 35 mm e aggiungere immediatamente 2 mL di PBS-/-. Quindi aggiungere 2 mL della soluzione proteica di rivestimento 2x.
- Incubare i trucioli a temperatura ambiente per 2 ore.
- Lavare la soluzione di rivestimento con successive fasi di diluizione con PBS-/-. La superficie del truciolo dovrebbe essere sempre bagnata. Quindi, sostituire il PBS con 2 mL di mezzo di placcatura e incubare a 37 °C fino alle cellule di semina.
3. Isolamento delle macchine virtuali
- Preparare il mezzo di placcatura integrando DMEM:M199 (4:1) con siero di cavallo al 10%, siero bovino fetale al 4%, 2% HEPES (1 M) e 1% penicillina/streptomicina (10.000 U/mL)22.
- Sezionare il tessuto dal ventricolo sinistro di cuori di ratto neonatale di 2 giorni e trasferirlo in un piatto di 10 cm contenente PBS. Tagliare il tessuto in circa 1 mm3 pezzi.
- Trasferire il tessuto raccolto in un tubo del tappo del rotore, dotato di un rotore nel tappo per la dissociazione dei tessuti(Tabella dei materiali). Lasciare depositare il tessuto e quindi rimuovere con cura il supernatante.
- Aggiungere 2,5 ml della miscela di enzimi 1 e 2, preparata utilizzando il kit di dissociazione cardiaca neonatale(tabella dei materiali),al tubo C e chiudere saldamente il cappuccio.
- Inserire il tubo del tappo del rotore sul manicotto del dissociatore, dotato di riscaldatori(Figura 2A e B)(Tabella dei materiali). Eseguire il programma di incubazione 37C_mr_NHDK_1 (Figura 2C), che dura circa un'ora.
- Mentre il programma di incubazione è in esecuzione, preparare tampone PEB contenente 2 mM EDTA e 0,5% di albumina di siero bovino (BSA) in PBS, pH 7.2, e mantenerlo a 4 °C.
- Dopo aver terminato il programma di incubazione, staccare il tubo del tappo delrotore (Figura 2D)e aggiungere 7,5 ml di mezzo di placcatura preri riscaldato.
- Rimorsi il campione e filtrare la sospensione cellulare utilizzando un colino da 70 μm.
- Lavare il colino con altri 3 mL di mezzo di placcatura.
- Centrifugare la sospensione cellulare a 600 x g per 5 min. Aspirare completamente il supernatante.
- Resuspend il pellet cellulare in 60 μL di tampone PEB freddo.
- Aggiungere 20 μL di Neonatal Cardiomyocyte Isolation Cocktail (Table of Materials), contenente perline di dimensioni micron che si rivolgono a non-CRM.
- Aggiungere 20 μL di perline anti-globuli rossi (Tabella dei materiali).
- Mescolare la sospensione e incubare a 4 °C per 15 min.
- Aggiungere 400 μL di tampone PEB.
- Applicare la sospensione cellulare sulla colonna LD (Table of Materials), che è stata inserita verticalmente in un supporto magnetico e lavata bene con tampone PEB.
- Raccogliere celle non etichettate e lavare la colonna con 0,5 mL di tampone PEB.
- Aggiungere 8 ml di mezzo di placcatura e trasferire la sospensione cellulare in un pallone dacoltura non rivestito da 75 cm 2 e incubare a 37 °C per 1,5 ore. I restanti non-CRM inizieranno ad aderire alla coltura cellulare non patinato e la sospensione cellulare sarà arricchita dalla popolazione CM.
4. Modelli di macchine virtuali
NOTA: abbiamo confrontato singole macchine virtuali con AR di 1:1, 7:1 o 11:1. Questo viene fatto seminando le MACCHINE di ratto neonatale isolate su un chip appositamente progettato riempito con micropattern rivestiti di fibronectina con AR definiti di 1:1, 7:1 o 11:1. I micropattern erano rivestiti di fibronectina, circondati da superficie citofobica. Pertanto, le macchine virtuali attaccheranno, diffonderanno e cattureranno l'AR definito dei micropattern crescendo esclusivamente sul substrato di fibronectina. Modellare le macchine virtuali isolate in base ai passaggi seguenti.
- Trasferire la sospensione cellulare in un tubo da 15 ml, contare le cellule e diluire a una concentrazione di 100.000 celle per ml aggiungendo un mezzo di placcatura appropriato.
- Aggiungere 2 mL di sospensione cellulare sul chip, che è già stato immerso in 2 mL di mezzo di placcatura caldo all'interno di una piastra greiner petri da 35 mm.
- Incubare il piatto a 37 °C con il 5% di CO2 per lasciare che le MACCHINE si attacchino ai micropattern rivestiti di fibronectina e consentire a ciascun CM di acquisire l'AR del suo micropattern del substrato.
- Dopo 18 ore, controllare il chip. Se la maggior parte delle cellule si è attaccata, staccare e rimuovere i detriti e le cellule morte che sono attaccati alla cella modellata. Eseguire questa operazioni rimuovendo il mezzo di placcatura e aggiungendo delicatamente PBS-/- dropwise, partendo dal centro del chip per poi spostarsi verso i lati. Ripetere 2 volte.
- Aspirare il PBS con un mezzo di manutenzione fresco integrando DMEM:M199 (4:1) con siero di cavallo al 4%, 4% siero bovino fetale, 2% HEPES (1 M) e 1% penicillina/streptomicina (10.000 U/mL).
5. Prelievo di macchine virtuali aderenti
NOTA: Dopo un periodo di recupero di 72 ore, le singole MACCHINE modellate vengono prelevate dai loro micropattern di fibronectina utilizzando un raccoglitore di celle semi-automatizzato (Tabella dei materiali) ( Figura3). La selezione celle utilizza un software23 per controllare lo stadio motorizzato (Figura 3A). Un microcapillare di vetro da 70 μm(figura 3B)viene utilizzato per raccogliere e iniettare le MACCHINE a ratto neonatale modellate. Il raccoglitore di celle ordina le cellule aderenti generando un vuoto e iniettando le cellule applicando pressione. Il vuoto nella siringa numero 1 viene applicato tirando la siringa utilizzando la pompa della siringa (Figura 3C). La pressione idrostatica si basa sulla gravità e indotta posizionando la siringa numero 2 a una distanza di 87 cm sopra la scrivania del microscopio. Le siringhe 1 e 2 sono collegate rispettivamente tramite tubi PTFE, alle valvole 1 e 2, che sono incorporate nell'unità di controllo (Figura 3D). I tubi PTFE sono completamente riempiti con acqua priva di RNasi. La singola cella prelevata viene quindi iniettata nel tubo di reazione a catena della polimerasi (PCR), contenente 3,55 μL di tampone di lisi.
- Dopo un periodo di coltura di 72 ore, rimuovere il vecchio mezzo e sciacquare delicatamente la superficie del truciolo con DPBScaldo -/-.
- Mantenere piatto il piatto e aspirare delicatamente il vecchio mezzo con una pipetta da 1000 μL da un lato del piatto. Assicurarsi che il chip rimanga sempre bagnato.
- Aggiungere 2 mL di DPBS-/- staccare delicatamente e a goccia la maggior parte delle cellule morte che sono attaccate alle celle a motivi geometrici, partendo dal centro del chip e quindi spostandosi ai suoi lati.
- Aspirare la maggior parte del DPBS a rimuovere il maggior numero possibile di celle galleggianti staccate, partendo dal centro del chip e quindi spostandosi ai lati.
- Ripetere ancora una volta i passaggi 5.1.2 e 5.1.3.
- Utilizzare forcep angolate per afferrare il bordo del chip e trasferirlo immediatamente in una nuova piastra Greiner Petri sterile da 35 mm per ridurre il numero di celle galleggianti durante la raccolta.
- Aggiungere immediatamente 1,5 mL di DPBS-/- in modo che il chip non si asciughi.
- Aggiungere 1,5 μL di vibrant dye cycle verde per visualizzare i nuclei delle cellule vive.
- Posizionare il truciolo al centro della piastra di Petri greiner usando la punta delle forcep.
- Mettere una camera(Tabella dei materiali)sopra il truciolo. La camera fissa il chip sul fondo del piatto, senza bloccare l'accesso ai modelli cellulari.
- Montare la piastra Greiner Petri sul portapiacstoviglie dello stadio di selezione celle e inserire il cappuccio magnetico.
- Calibrare l'iniezione automatica.
- Individuare il mirino, inciso sullo stage motorizzato al centro dell'immagine nella finestra Vista dal vivo.
- Concentrati sul mirino e seleziona il pulsante Calibrazione per iniezione automatica nella finestra di scansione e smistamento.
- Sostituire il DPBS-/- con 1,5 mL di DPBS/tripina-/- (1:1) mentre il piatto è sul palco, per allentare le cellule dalla fibronectina in modo che sia possibile utilizzare un vuoto fluido per raccogliere le cellule.
- Eseguire la scansione dell'intero chip utilizzando la scheda Scansione nella finestra di scansione e ordinamento. Individuare l'angolo in alto a sinistra del chip nel campo visivo e fare clic su Ottieni posizione corrente del microscopio nella riga dell'angolo in alto a sinistra.
- Quindi, spostare lo stadio motorizzato nell'angolo in basso a destra del chip. Mettere a fuoco il microscopio e fare clic su Ottieni posizione corrente del microscopio nella riga dell'angolo in basso a destra.
- Fare clic sul pulsante Imposta piano più nitido e nella finestra spuntata fare clic sull'angolo Vai in alto a destra. Mettere a fuoco il microscopio e fare clic su Vai nell'angolo in basso a sinistra e impostare lo stato attivo. Al termine, fare clic sul pulsante Fine e iniziare la scansione.
- Quando la scansione viene terminata, passare alla scheda Analisi e selezionare le singole celle che superano i criteri di studio.
- Assicurarsi che il microcapillare di vetro si trova al centro della vista dal vivo del microscopio.
- Controllare la pompa della siringa utilizzando la finestra Pompa del software. Creare un vuoto prelevando 4 ml da una siringa numero 1 da 50 ml, che ha un diametro di 27 mm.
- Nella scheda Ordinamento
- Impostare i parametri di iniezione delle valvole. È stato calcolato che il volume di iniezione che ha consegnato una singola cella prelevata era di 1 μl, se la valvola 2 è stata aperta per 120 millisecondi e quindi la valvola 1 aperta per 20 era millisecondi dopo un lapista di tempo di 200 millisecondi. A causa dell'elasticità dei tubi, aprire la valvola 1 per interrompere l'iniezione del flusso.
- Impostare i parametri di prelievo delle valvole. È stato calcolato che se la valvola 1 è stata aperta per 20 millisecondi e quindi la valvola 2 è stata aperta per 10 millisecondi, dopo un lapche di tempo di 10 millisecondi, la maggior parte delle celle modellate può essere prelevata. Questo perché i loro attacchi di fibronecina sono stati allentati dopo essere stati trattati con tripparina.
- Fare clic sul pulsante Calcola percorso. Il software calcola il percorso più veloce da cella a cella, per raccogliere e iniettare le celle selezionate in tutto il chip.
- Mettere a fuoco il microscopio su una cella a motivi geometrici sulla superficie del truciolo.
- Usando il joystick, spostare il microcapillare verso il basso con attenzione, in modo che l'immagine più nitida della punta del microcapillare possa essere ottenuta senza toccare la cellula.
- Fate clic sul pulsante Imposta (Set) nella sezione Offset Micropipette . Apparirà una nuova finestra, che mostra la sezione trasversale microcapillare. Clicca sul centro esatto del capillare. Il software registrerà quindi l'offset della punta del capillare nelle coordinate x, y e z.
- Avviare l'ordinamento utilizzando il pulsante Avvia ordinamento.
Risultati
Il tessuto è stato sezionato dal ventricolo sinistro del cuore neonatale di 2 giorni e diviso in singole cellule. Quindi le MACCHINE arricchite sono state sezionate su un chip contenente modelli di fibronecina con AR distinti. Dopo 72 ore di coltura, il mezzo è stato sostituito da 1:1000 Vibrant Dye Cycle green in DPBS-/- per 2 minuti per visualizzare i nuclei delle cellule vive. Successivamente, le cellule sono state trattate con DPBS-/-/trypsin (1:1) per allentare le cellule dalla fibronectina, in modo che un vuoto fluido potesse essere utilizzato per facilitare il prelievo cellulare. Nel frattempo, l'intero chip è stato scansionato con un ingrandimento di 10x, utilizzando un microscopio invertito collegato al raccoglitore di celle. Questo è stato effettuato prima che le cellule si arrotondassero a causa del trattamento con tripside. Le celle qualificate sono state selezionate, in base all'immagine digitalizzata, e le relative coordinate sono state salvate nel software di selezione celle. I micropattern sono stati selezionati solo se contenevano una singola cellula mononucleata e solo quando la cellula copriva completamente il suo micropattern di fibronecina. Lo smistatore di celle ha prelevato le celle selezionate una per una e ogni singola cella che è stata prelevata con successo è stata immediatamente iniettata in un singolo tubo PCR e posizionata sullo stadio del microscopio. Ogni tubo PCR conteneva 3,55 μL di tampone dilisi(tabella 2). Il processo di ordinamento, iniziato con la rimozione del supporto, è stato completato entro 40 minuti. La sintesi complementare dell'acido deossiribonucleico (cDNA), la preamplificazione e la purificazione pcr sono state eseguite sulle singole cellule liti, sulla base del protocollo Smart-Seq224 (tabella 3). La qualità del cDNA purificato è stata controllata da un analizzatore automatico di elettroforesi. L'elettroferogramma del cDNA pre-amplificato di una singola cella selezionata è presentato nella figura 4. Le librerie RNA-Seq sono state preparate secondo il protocollo Smart-Seq224.
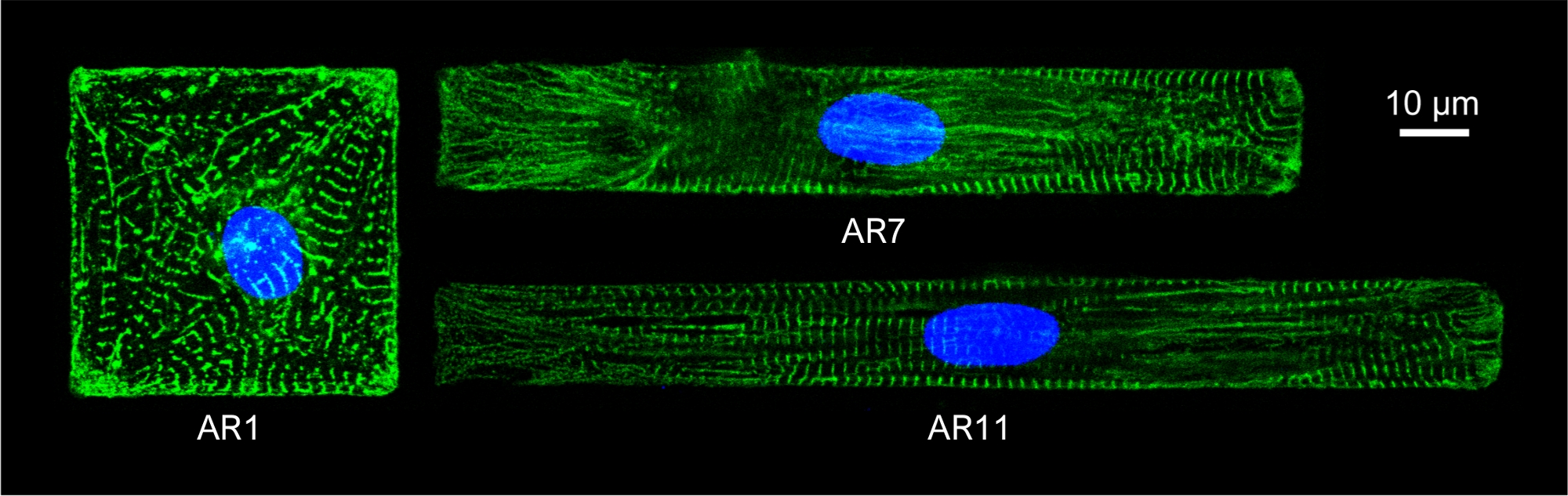
Per osservare la struttura sarcomere delle macchine virtuali modellate, le MACCHINE modellate sono state macchiate con anticorpi sarcomerici α-actina. Le cellule sono state incubate con Donkey Anti-Mouse IgG Alexa Fluor 488 1:800 per 1 ora a temperatura ambiente per la colorazione secondaria. I nuclei erano macchiati da DAPI da 1 μg/mL. Le immagini immunofluorescenti sono state acquisite con un microscopio confocale invertito, utilizzando un obiettivo 63x oil-immersion (NA 1.4)(Figura 5).

Figura 1: Disposizione del chip con micropattern di fibronecina.
(A) Immagine del chip progettato su misura. Il chip è un coverslip da 19,5 mm x 19,5 mm con micropattern di fibronectina, stampato per fotolitografia su un vetro borosilicato. (B) Layout truciolo. Il chip è diviso in tre zone e ogni zona è costituita da micropattern di fibronecina con AR specifico. Questa cifra è stata modificata da "materiale supplementare 1" di Haftbaradaran Esfahani et alhttp://creativecommons.org/licenses/by/4.0/. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Dissociatore dotato di apparecchi riscaldatori.
(A) L'intero strumento dissociatore utilizzato per la dissociazione completamente automatizzata dei ventricoli sinistro del ratto neonatale di 2 giorni. (B) Unità di riscaldamento. (C) Tubo del tappo del rotore. (D) Programmi pronti all'uso per un flusso di lavoro completamente automatizzato di dissociazione dei tessuti. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: Apparecchio di selezione cellulare.
(A) Vista ingrandita dello stadio motorizzato del raccoglitore di celle. Sul palco è incorporato un supporto per piastre di Petri e 80 fori per 10 strisce PCR e un foro per il mirino di calibrazione. (B)Vista allargata di un microcapillare di vetro. (C) La pompa della siringa. (D) L'unità di comando, che controlla l'orario di apertura e chiusura delle valvole 1 e 2, montata all'interno della centralina. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: L'elettroferogramma del cDNA preamplificato di una singola cella selezionata.
19 cicli PCR di pre-amplificazione sono stati utilizzati per ottenere 15 μL di 1 ng/μL di resa cDNA purificata. Si osserva una banda chiara nella trama densitometrica simile a un gel che corrisponde al picco a 1852 bp nell'elettroferogramma. La dimensione media dei frammenti è di 1588 bp. Inoltre, la piccola quantità di frammenti che sono più corti di 300 bp indica una buona libreria cDNA. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 5: Colorazione immunofluorescente della struttura sarcomerica α-actina (verde) e nucleo (blu) di CCI modellate con ARS diversi.
La cromatina era macchiata da DAPI. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}
| Morfotipo | Ar | Lunghezza (μm) | Larghezza (μm) | Area fibronectina (μm2) |
| Brr | 1:1 | 47 | 47 | 2209 |
| AR7 | 7:1 | 126 | 18 | 2268 |
| AR11 | 11:1 | 155 | 14 | 2170 |
Tabella 1: Geometria delle MACCHINE con serie.
| Componente | Volume (μL) |
| Acqua priva di nucleas | 0.65 |
| (0,4% vol/vol) Tritone X-100 | 1.8 |
| miscela dNTP (25 mM) | 0.8 |
| Inibitore della RNasi (40 U μL-1) | 0.1 |
| Oligo-dT30VN oligonucleotidi (100 μM) | 0.1 |
| ERCC RNA Spike-In Mix (2,5 x 105 diluizione) | 0.1 |
| Singola cella iniettata | 1 |
| Volume totale | 4.55 |
Tabella 2: Buffer di lysis personalizzato a cella singola.
| Componente | Volume (μL) |
| Buffer del primo filamento Superscript II (5x) | 2 |
| DTT (100 mM) | 0.5 |
| Betaina (5 M) | 2 |
| Mgcl2 (1 M) | 0.1 |
| Inibitore della RNasi (40 U μL-1) | 0.25 |
| Trascrizione inversa Superscript II (200 U μL-1) | 0.5 |
| TSO (100 μM) | 0.1 |
| Volume totale | 5.45 |
Tabella 3: Mix di trascrizione inversa (RT) per una reazione RT per sintetizzare il cDNA del primo filamento dal lysate di un singolo CM.
Discussione
Questo studio ha utilizzato il sequenziamento dell'RNA a singola cella, che è una tecnologia nuova e potente in grado di rilevare il trascrittame di singole cellule. È stato combinato con un approccio innovativo alla coltivazione di singole macchine virtuali, in modo che abbiano assunto diverse AR che, altrimenti, avrebbero potuto essere osservate solo in vivo.
Lo studio aveva alcuni limiti. Ad esempio, le macchine virtuali neonatale hanno dovuto essere utilizzate per generare diversi morfotipi, in quanto è eccezionalmente impegnativo per la coltura di CCI adulte abbastanza vitali per 72 ore in forme definite. Inoltre, le macchine virtuali sono state coltivate per 72 ore ex vivo, il che potrebbe aver avuto un impatto sul modello di espressione genica. Tuttavia, questa ristrutturazione era necessaria, in modo che le cellule potessero formare morfotipi specifici. Inoltre, sono state selezionate solo singole cellule mononucleate e completamente coperte dal micropattern della fibronectina per lo smistamento. Le celle modellate su ogni chip devono essere ordinate semplicemente in un ciclo di ordinamento. Infine, circa 50 celle che rappresentano circa un terzo delle cellule selezionate sono state prelevate con successo da ogni chip. Esistono due motivi che hanno limitato il numero di celle selezionate correttamente. In primo luogo, alcune cellule erano troppo strettamente attaccate al modello di fibronecina e il flusso di pickup non era abbastanza forzato da raccoglierle con successo. In secondo luogo, a causa del trattamento con tripina, l'attaccamento tra alcune cellule e fibronectina è diventato troppo sciolto. Di conseguenza, queste cellule sono state allontanate dai loro micropattern di fibronectina, quando il microcapillare si è avvicinato a loro, e non sono state raccolte. Gli autori non sostengono che questa configurazione sia la stessa di un ambiente in vivo, ma si è rivelata un approccio praticabile per rispondere alla domanda di ricerca.
Il metodo proposto è applicabile a diversi tipi di cellule (ad esempio, per le macchine hiPS-CRM). Tuttavia, i seguenti fattori dovrebbero essere ottimizzati per studiare altri tipi di cellule. Per il rivestimento dei micropattern devono essere utilizzate molecole adesive ECM adatte per l'attacco del tipo di cella specifico. La geometria dei micropattern dovrebbe essere modificata in base alla domanda di studio e al tipo di cella. Il periodo di coltivazione può essere modificato in base alla domanda di studio. Il reagente di distacco e il suo tempo di incubazione dovrebbero essere ottimizzati proprio per il tipo di cellula di studio. Ad esempio, l'accutasi può essere usato al posto di TryplE per il distacco di cellule staminali embrionali e neuronali. I parametri del tempo di apertura delle valvole devono essere esaminati per prelevare le celle con successo, ma delicatamente. In sintesi, abbiamo progettato una nuova piattaforma per studiare la forma cellulare che può fornire una risorsa preziosa per i ricercatori del settore. In questo contesto, abbiamo progettato un approccio sperimentale che imitava le forme caratteristiche in vitro imposte al CM in vivo da vincoli emodinamici per identificare l'interazione tra architettura cellulare ed espressione genica. Reportiamo anche lo sviluppo di una nuova piattaforma per studiare HF in vitro e l'identificazione della forma cellulare come potente determinante dell'espressione genica. Questa è una nuova osservazione con implicazioni di vasta portata per la biologia e la medicina.
Divulgazioni
Nessuno.
Riconoscimenti
Nessuno.
Materiali
| Name | Company | Catalog Number | Comments |
| 2100 Bioanalyzer Instrument | Agilent Technologies | G2939BA | Automated electrophoresis analyzer |
| Anti-Red Blood Cell MicroBeads | Miltenyi Biotec | 130-109-681 | |
| Axio Observer microscope | Zeiss | Z1 | Inverted microscope |
| Betaine solution (5 M) | Sigma-Aldrich, MERCK | B0300 | |
| CellSorter | CELLSORTER | https://www.singlecellpicker.com/ | Cell picker |
| CYTOOchamber | CYTOO | 30-010 | Custom-designed chip |
| CYTOOchip | CYTOO | 10-950-00-18 | Chamber |
| DMEM | Thermo Fisher Scientific | 31966-021 | high glucose, GlutaMAX Supplement |
| dNTP mix (25 mM) | Thermo Fisher Scientific | R1122 | |
| Donkey Anti-Mouse IgG Alexa Fluor 488 | Abcam PLC | ab150105 | |
| DTT (100 mM) | Thermo Fisher Scientific | 18064071 | |
| ERCC RNA Spike-In Mix | Thermo Fisher Scientific | 4456740 | |
| Fetal Bovine Serum | Thermo Fisher Scientific | 10082-147 | |
| Fibronectin | Sigma-Aldrich, MERCK | F4759 | |
| gentleMACS C Tube | Miltenyi Biotec | 130-093-237 | Rotor-cap tube |
| gentleMACS Octo Dissociator with Heaters | Miltenyi Biotec | 130-096-427 | Dissociator with heater |
| Greiner CELLSTAR Petri dish | Sigma-Aldrich, MERCK | P6987 | |
| HEPES (1 M) | Thermo Fisher Scientific | 15630-056 | |
| Horse Serum | Sigma-Aldrich, MERCK | H0146 | |
| LD column | Miltenyi Biotec | 130-042-901 | |
| Medium 199 | Thermo Fisher Scientific | 31150-022 | |
| NE-1000 syringe pump | New Era Pump Systems | NE-1000 | Syringe pump |
| Neonatal Cardiomyocyte Isolation Cocktail, rat | Miltenyi Biotec | 130-105-420 | |
| Neonatal Heart Dissociation Kit, mouse and rat | Miltenyi Biotec | 130-098-373 | |
| Oligo-dT30VN oligonucleotides | IDT Technology | 5′–AAGCAGTGGTATCAACGCAGAGTACT30VN-3′ | |
| RNAse inhibitor (40 U µL-1) | Clontech | 2313A | |
| sarcomeric α-actinin | Sigma-Aldrich, MERCK | EA-53 | |
| SP8 confocal microscope | Leica Microsystems | SP8 | Confocal microscope |
| Superscript II first-strand buffer (5x) | Thermo Fisher Scientific | 18064071 | |
| Superscript II reverse transcriptase (200 U µL-1) | Thermo Fisher Scientific | 18064071 | |
| Triton X-100 | Sigma-Aldrich, MERCK | T9284 | |
| TryplE Express enzyme, no phenol red | Thermo Fisher Scientific | 12604013 | |
| TSO (100 µM) | QIAGEN | 5′-AAGCAGTGGTATCAACGCAGAGTACATrGrG+G-3′ | |
| Vibrant Dye Cycle green | Thermo Fisher Scientific | V35004 |
Riferimenti
- Heineke, J., Molkentin, J. D. Regulation of cardiac hypertrophy by intracellular signalling pathways. Nature Reviews Molecular Cell Biology. 7 (8), 589-600 (2006).
- Haftbaradaran Esfahani, P., et al. Cell shape determines gene expression: cardiomyocyte morphotypic transcriptomes. Basic Research in Cardiology. 115 (1), 7(2019).
- Kontrogianni-Konstantopoulos, A., Benian, G., Granzier, H. Advances in Muscle Physiology and Pathophysiology 2011. Journal of Biomedicine and Biotechnology. 2012, 930836(2012).
- Hill, J. A., Olson, E. N. Cardiac plasticity. New England Journal of Medicine. 358 (13), 1370-1380 (2008).
- Bray, M. A., Sheehy, S. P., Parker, K. K. Sarcomere alignment is regulated by myocyte shape. Cell Motility and the Cytoskeleton. 65 (8), 641-651 (2008).
- Benjamin, I. J., Schneider, M. D. Learning from failure: congestive heart failure in the postgenomic age. Journal of Clinical Investigation. 115 (3), 495-499 (2005).
- Opie, L. H., Commerford, P. J., Gersh, B. J., Pfeffer, M. A. Controversies in ventricular remodelling. Lancet. 367 (9507), 356-367 (2006).
- Braunwald, E. Cardiomyopathies: An Overview. Circulation Research. 121 (7), 711-721 (2017).
- Knoll, R., et al. The cardiac mechanical stretch sensor machinery involves a Z disc complex that is defective in a subset of human dilated cardiomyopathy. Cell. 111 (7), 943-955 (2002).
- Frangogiannis, N. G. The Extracellular Matrix in Ischemic and Nonischemic Heart Failure. Circulation Research. 125 (1), 117-146 (2019).
- Marian, A. J., Braunwald, E. Hypertrophic Cardiomyopathy: Genetics, Pathogenesis, Clinical Manifestations, Diagnosis, and Therapy. Circulation Research. 121 (7), 749-770 (2017).
- Mozaffarian, D., Caldwell, J. H. Right ventricular involvement in hypertrophic cardiomyopathy: a case report and literature review. Clinical Cardiology. 24 (1), 2-8 (2001).
- Olsson, M. C., Palmer, B. M., Stauffer, B. L., Leinwand, L. A., Moore, R. L. Morphological and functional alterations in ventricular myocytes from male transgenic mice with hypertrophic cardiomyopathy. Circulation Research. 94 (2), 201-207 (2004).
- Toepfer, C. N., et al. Hypertrophic cardiomyopathy mutations in MYBPC3 dysregulate myosin. Science Translational Medicine. 11 (476), (2019).
- McKenna, W. J., Maron, B. J., Thiene, G. Classification, Epidemiology, and Global Burden of Cardiomyopathies. Circulation Research. 121 (7), 722-730 (2017).
- Schultheiss, H. P., et al. Dilated cardiomyopathy. Nature Reviews Disease Primers. 5 (1), 32(2019).
- Knoll, R. A role for membrane shape and information processing in cardiac physiology. Pflugers Arch. 467 (1), 167-173 (2015).
- Rangamani, P., et al. Decoding information in cell shape. Cell. 154 (6), 1356-1369 (2013).
- Kuo, P. L., et al. Myocyte shape regulates lateral registry of sarcomeres and contractility. American Journal of Pathology. 181 (6), 2030-2037 (2012).
- Gerdes, A. M., Onodera, T., Wang, X., McCune, S. A. Myocyte remodeling during the progression to failure in rats with hypertension. Hypertension. 28 (4), 609-614 (1996).
- Kehat, I., et al. Extracellular signal-regulated kinases 1 and 2 regulate the balance between eccentric and concentric cardiac growth. Circulation Research. 108 (2), 176-183 (2011).
- Sreejit, P., Kumar, S., Verma, R. S. An improved protocol for primary culture of cardiomyocyte from neonatal mice. In Vitro Cellular & Developmental Biology - Animal. 44 (3-4), 45-50 (2008).
- Kornyei, Z., et al. Cell sorting in a Petri dish controlled by computer vision. Scientific Reports. 3, 1088(2013).
- Picelli, S., et al. Full-length RNA-seq from single cells using Smart-seq2. Nature Protocols. 9 (1), 171-181 (2014).
- Schmick, M., Bastiaens, P. I. H. The interdependence of membrane shape and cellular signal processing. Cell. 156 (6), 1132-1138 (2014).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati