
Method Article
Une approche pour étudier la transcriptomics dépendante de la forme à un seul niveau cellulaire
Dans cet article
Résumé
Cet article présente des méthodes pour la croissance des myocytes cardiaques de différentes formes, qui représentent différentes pathologies, et le tri de ces myocytes cardiaques adhérents basés sur leur morphologie à un seul niveau cellulaire. La plate-forme proposée offre une nouvelle approche au débit élevé et au dépistage des médicaments pour différents types d’insuffisance cardiaque.
Résumé
Différents types d’hypertrophie cardiaque ont été associés à un volume accru de myocytes cardiaques (MC), ainsi qu’à des changements dans la morphologie cm. Bien que les effets du volume cellulaire sur l’expression des gènes soient bien connus, les effets de la forme cellulaire ne sont pas bien compris. Cet article décrit une méthode qui a été conçue pour analyser systématiquement les effets de la morphologie cm sur l’expression des gènes. Il détaille le développement d’une nouvelle stratégie de piégeage à cellules simples qui est ensuite suivie d’un séquençage de l’ARNm unicelliste. Une puce micropatrée a également été conçue, qui contient 3000 micropatterns de fibronectine en forme de rectangulaire. Cela permet de faire croître les MC en longueur distincte : rapports d’aspect largeur (AR), correspondant à différents types d’insuffisance cardiaque (HF). Le document décrit également un protocole qui a été conçu pour ramasser les cellules individuelles de leur modèle, à l’aide d’un micro-pipetting semi-pipetting picker cellulaire, et les injecter individuellement dans un tampon de lyse séparée. Cela a permis de profiler les transcriptomes des MC simples avec des morphotypes géométriques définis et de les caractériser en fonction d’une gamme de conditions normales ou pathologiques : cardiomyopathie hypertrophique (HCM) ou après charge/concentrique versus cardiomyopathie dilatée (DCM) ou préchargement/excentrique. En résumé, cet article présente des méthodes pour cultiver des MC de formes différentes, qui représentent différentes pathologies, et le tri de ces MC adhérents en fonction de leur morphologie à un niveau unicellulaire. La plate-forme proposée offre une nouvelle approche du dépistage du débit élevé et des médicaments pour différents types de HF.
Introduction
Selon l’Organisation mondiale de la santé, les maladies cardiovasculaires (MCV) sont une cause majeure de morbidité et de mortalité dans le monde. La MCV affecte considérablement la qualité de vie des gens et a un impact socioéconomique énorme. Les cardiomyopathies, telles que HCM et DCM, sont des désordres primaires du muscle cardiaque et les causes principales de HF ont été associées à la morbidité et à la mortalité élevées. Il existe de nombreuses causes de HF, y compris les effets environnementaux, tels que les infections et l’exposition aux toxines ou à certainsmédicaments 8. La HF peut également être causée par une prédisposition génétique, à savoir les mutations9. On croit que les changements dans la composition génétique qui affectent les molécules, les intégines ou les protéines cytosquelettiques de matrice extracellulaire (ECM) pourraient être responsables de la mécanosensation altérée et de divers types de maladiecardiaque 10.
La caractéristique principale de HCM est hypertrophie inexpliquée du ventriculegauche 11,et parfois du ventricule droit12,et ceci se présente fréquemment avec la participation prédominante du septum intertriculaire. HCM est également caractérisé par le dysfonctionnement diastolique et le désarroi de myocyte et lafibrose 13. Dans la plupart des cas, l’appareil contractile du coeur est affecté par des mutations dans les protéines sarcomeric, menant à la contractilité accrue des myocytes14. En revanche, DCM est caractérisé par la dilatation d’un, ou les deux, ventricules et a une étiologie familiale dans 30% à 50% des cas15. DCM affecte un large éventail de fonctions cellulaires, menant à la contraction altérée des myocytes, à la mort cellulaire et à la réparation fibrotique16.
La génétique a montré que certains types de mutations forcent les MC uniques à adopter des caractéristiques de forme spécifiques pendant hcm3, à savoir les cellules de forme carrée avec une longueur: largeur AR qui est presque égale à 1:14 (AR1). Il en va de même pour le DCM, avec des cellules allongées avec un AR qui est presque égal à 11:1 (AR11). De plus, la HF peut être causée par une augmentation de la charge après chargement (p. ex., dans l’hypertension). Dans ces cas, les exigences hémodynamiques obligent les MC à prendre des formes carrées, selon la loi de Laplace, et l’AR passe de 7:15 (AR7) à 1:16,7. Hf peut également être causée par une augmentation du préchargement (par exemple, dans des conditions qui conduisent à une surcharge de volume). Lorsque cela se produit, les contraintes biophysiques forcent les MC à s’allonger et l’AR passe de 7:1 à 11:1.
L’activité de signalisation aux membranes dépend des paramètres mondiaux de géométrie cellulaire, tels que l’AR cellulaire, la taille, la surface de la membrane et la courbure membranaire18. Quand les MC néonatals de rat ont été plaqués sur des substrats qui ont été modelés pour limiter les cellules dans une longueur spécifique : AR de largeur, ils ont démontré la meilleure fonction contractile quand les rapports étaient semblables aux cellules dans un coeur adulte sain. En revanche, ils ont mal performé quand les rapports étaient semblables à ceux des myocytes dans les coeurs défaillants19. Dans les premiers stades de l’hypertrophie, les cellules deviennent plus larges, comme en témoigne une augmentation de la zone transversale. Hf se produit dans les derniers stades de l’hypertrophie et les cellules semblent généralement allongées. Par conséquent, il n’est pas surprenant que les modèles in vivo de rat de l’hypertrophie chronique aient rapporté une augmentation de la longueur ventriculaire gauche de myocyte d’environ 30%20,mais les MC adultes du modèle transgénique de souris qui ont été aiguëment traités avec des stimulus hypertrophiques in vitro ont démontré des augmentations semblables de largeur cellulaireau lieu 21.
Le séquençage de l’ARN à cellules individuelles, qui permet une analyse précise du transcriptome des cellules individuelles, révolutionne actuellement la compréhension de la biologie cellulaire. Cette technologie était la méthode préférée lorsqu’il s’agissait de répondre à la question de savoir comment les formes cellulaires individuelles affectaient l’expression des gènes. Nous avons comparé des cellules individuelles avec différentes formes, en particulier avec des AR de 1:1, 7:1 ou 11:1. Ceci a été fait en ensemencant les MC ventriculaires néonatals de rat sur une puce spécialement conçue remplie des micropatterns fibronectin-enduits2 avec les ARs définis de 1:1, 7:1 ou 11:1. Les micropatternes ont été fabriqués à l’aide de la technologie de photolithographie. Les micropatterns ont été enduits par la fibronectine, entourées par la surface cytophobe. Par conséquent, les MC fixent, propagent et capturent l’AR défini des micropatrins en se cultivant uniquement sur le substrat de fibronectine, tout en évitant la zone cytophobe. Les micropatternes ne sont pas dans un format bien formé. Au lieu de cela, le niveau de fibronectine est exactement à la même hauteur de la zone cytophobe environnante. Ceci a fourni des conditions semblables aux cellules croissantes dans une boîte de Petri, car il n’y a aucun effort des murs environnants. En outre, la surface des micropatrins avec différents AR sont égaux.
Il y avait deux aspects particulièrement importants de la conception expérimentale, qui ont mené à l’utilisation du séquençage d’ARN unicelliste au lieu du séquençage en vrac d’ARN. Premièrement, seuls quelques pourcentages des micropatrins peuvent être occupés par une seule cellule. Deuxièmement, parfois, une seule cellule n’occupe pas entièrement la surface du micropatrin. Les cellules individuelles qui recouvrent complètement une surface micropatreuse doivent être cueillies pour l’analyse de l’ARN unicellule. Étant donné que seul un sous-groupe de cellules plaquées sur une puce satisfaisait aux deux critères, il n’était pas possible de simplement trypsiniser la puce entière et de recueillir toutes les cellules pour le séquençage en vrac de l’ARN. Les cellules qualifiées devaient être cueillies individuellement à l’aide d’un cueilleur de cellules semi-automatisé.
On ne sait pas encore si la forme du CM, en soi, a un impact intrafonctionnel sur le syncytium myocardique. L’objectif principal des méthodes proposées dans cet article était de développer une nouvelle plate-forme pour étudier si la forme cellulaire en soi avait un impact sur le transcriptome17. Bien que les études in vitro soient différentes des études in vivo, le but de cette étude était d’étudier l’effet des différentes formes cellulaires sur l’expression des gènes, en gardant à l’esprit que la comparaison des cellules avec différentes formes in vivo est extrêmement exigeante. Ces expériences ont été inspirées par Kuo et coll.19, qui ont utilisé une approche similaire et ont signalé qu’elles observaient des changements dans les paramètres physiologiques en raison de changements dans la forme des cellules.
Protocole
Toutes les procédures concernant les animaux étaient conformes aux règlements du comité d’éthique animale de l’Institut Karolinska de Stockholm, en Suède.
1. Mise en page des puces à micro-motifs
- Utilisez une puce sur mesure (Table of Materials) ( Figure1A): un coverslip de 19,5 mm x 19,5 mm avec micropatrins activés, imprimé par photolithographie sur verre borosilicate.
REMARQUE : Ces micropatrins sont entourés d’une zone cytophobe. Par conséquent, une cellule ensemencée ne peut s’attacher et se développer sur l’un de ces micropatrins et capturer l’AR de ce micropatrin. La puce est divisée en trois zones et chaque zone se compose de micropatrins avec un AR spécifique. La disposition des copeaux est indiquée dans la figure 1B. La géométrie des AR définis est présentée dans le tableau 1. Des images fluorescentes amplifiées des différentes formes des micropatrins de fibronectine sont montrées dans les images inférieures de la figure 1B.
2. Revêtement de copeaux micropatternés
- Préparer 2x solution de protéines de revêtement pour chaque puce en ajoutant 80 μg de fibronectine à 2 mL de solution saline tamponnée de phosphate (PBS-/-).
- Transférer une puce dans une boîte de Pétri Greiner de 35 mm et ajouter immédiatement 2 mL de PBS-/-. Ajouter ensuite 2 mL de la solution protéique 2x coating.
- Incuber les copeaux à température ambiante pendant 2 h.
- Lavez la solution de revêtement par des étapes successives de dilution avec PBS-/-. La surface de la puce doit toujours être mouillée. Ensuite, remplacez le PBS par 2 mL de placage moyen et incubez à 37 °C jusqu’à ce que les cellules d’ensemencement.
3. Isolement des MC
- Préparer le milieu de placage en complétant le DMEM:M199 (4:1) avec 10% de sérum chevalin, 4% sérum bovin fœtal, 2% HEPES (1 M) et 1% pénicilline/streptomycine (10 000 U/mL)22.
- Disséquer le tissu du ventricule gauche des cœurs néonatals de rat de 2 jours et transférer dans un plat de 10 cm contenant du PBS. Couper le tissu en environ 1 mm3 morceaux.
- Transférer le tissu récolté dans un tube rotor-capuchon, équipé d’un rotor dans le bouchon pour la dissociation des tissus (Tableau des matériaux). Laissez le tissu s’installer, puis retirez soigneusement le surnatant.
- Ajouter 2,5 mL du mélange d’enzymes 1 et 2, préparé à l’aide de la trousse néonatale de dissociation cardiaque(tableau des matériaux),au tube C et fermer le bouchon hermétiquement.
- Insérez le tube rotor-capuchon sur le manchon du dissociateur, équipé de radiateurs (Figure 2A et B) ( Tableau desmatériaux). Exécutez le programme d’incubation 37C_mr_NHDK_1( Figure 2C), qui dure environ une heure.
- Pendant que le programme d’incubation est en cours d’exécution, préparez un tampon PEB contenant 2 mM EDTA et 0,5 % d’albumine de sérum bovin (BSA) dans PBS, pH 7.2, et gardez-le à 4 °C.
- Après la fin du programme d’incubation, détachez le tube rotor-capuchon(figure 2D)et ajoutez 7,5 mL de milieu de placage préchauffé.
- Resuspendez l’échantillon et filtrez la suspension cellulaire à l’aide d’une passoire de 70 μm.
- Laver la passoire avec un autre 3 mL de placage moyen.
- Centrifugeuse de la suspension cellulaire à 600 x g pendant 5 min. Aspirez complètement le surnatant.
- Resuspendez la pastille cellulaire dans 60 μL de tampon PEB froid.
- Ajouter 20 μL de cocktail d’isolation néonatale cardiomyocyte (Table of Materials), contenant des perles de la taille d’un micron qui ciblent les non-MC.
- Ajouter 20 μL de perles anti-globules rouges (Tableau des matériaux).
- Mélanger la suspension et incuber à 4 °C pendant 15 min.
- Ajouter 400 μL de tampon PEB.
- Appliquez la suspension cellulaire sur la colonne LD (Tableau des matériaux), qui a été inséré verticalement dans un support magnétique et bien lavé avec tampon PEB.
- Recueillir les cellules non étiquetées et laver la colonne avec 0,5 mL de tampon PEB.
- Ajouter 8 mL de milieu de placage et transférer la suspension cellulaire dans un flacon de culture non encoated de 75 cm2 et incuber à 37 °C pendant 1,5 h. Les autres non-MC commenceront à adhérer à la culture cellulaire non encoated et la suspension cellulaire sera enrichie par la population de CM.
4. Mc patterning
REMARQUE : Nous avons comparé les MC simples avec les AR de 1:1, 7:1 ou 11:1. Ceci est fait en ensemencer les MC de rat néonatal isolés sur une puce spécialement conçue remplie de micropatternes enduits de fibronectine avec des AR définis de 1:1, 7:1 ou 11:1. Les micropatterns ont été enduits par la fibronectine, entourées par la surface cytophobe. Par conséquent, les MC fixent, propagent et capturent l’AR défini des micropatrins en se cultivant uniquement sur le substrat de fibronectine. Modeler les MC isolés en fonction des étapes suivantes.
- Transférer la suspension cellulaire dans un tube de 15 mL, compter les cellules et les diluer à une concentration de 100 000 cellules par mL en ajoutant un support de placage approprié.
- Ajouter 2 mL de suspension cellulaire sur la puce, qui a déjà été immergée dans 2 mL de plat chaud moyen à l’intérieur d’une boîte greiner petri de 35 mm.
- Incuber le plat à 37 °C avec 5 % de CO2 pour laisser les MC s’attacher aux micropatternes recouverts de fibronectine et permettre à chaque CM d’acquérir l’AR de son micropattern de substrat.
- Après 18 h, vérifiez la puce. Si la plupart des cellules se sont attachées, détachez et enlevez les débris et les cellules mortes qui sont attachés à la cellule à motifs. Faites ceci en enlevant le milieu de placage et en ajoutant doucement PBS-/- dropwise, commençant du centre de la puce puis se déplaçant vers les côtés. Répétez 2 fois.
- Aspirez le PBS avec le milieu frais d’entretien en complétant DMEM:M199 (4:1) avec le sérum de cheval de 4%, le sérum bovin foetal de 4%, 2% HEPES (1 M) et 1% la pénicilline/streptomycine (10.000 U/mL).
5. Sélection des MC adhérents
REMARQUE : Après une période de culture de 72 heures, les MC simples à motifs sont choisis à partir de leurs micropatrins de fibronectine à l’aide d’un cueilleur de cellules semi-automatisé (Tableau des matériaux) ( Figure3). Le ramasseur de cellules utilise unlogiciel 23 pour contrôler l’étape motorisée (Figure 3A). Un microcapillaire en verre de 70 μm (figure 3B) est utilisé pour cueillir et injecter les MC à rat néonatal à motifs. Le cueilleur de cellules trie les cellules adhérentes en générant un vide et en injectant les cellules en appliquant une pression. Le vide dans la seringue numéro 1 est appliqué en tirant la seringue à l’aide de la pompe à seringues (figure 3C). La pression hydrostatique est basée sur la gravité et induite en plaçant la seringue numéro 2 à une distance de 87 cm au-dessus du bureau du microscope. Les seringues 1 et 2 sont reliées respectivement par tubes PTFE aux vannes 1 et 2, qui sont intégrées dans l’unité de commande (figure 3D). Les tubes PTFE sont entièrement remplis d’eau sans RNase. La cellule unique choisie est ensuite injectée au tube de réaction en chaîne de polymésase (PCR), contenant 3,55 μL de tampon de lyse.
- Après une période de culture de 72 heures, retirer l’ancien milieu et rincer doucement la surface de la puce avec du DPBSchaud -/-.
- Garder le plat plat et aspirer doucement l’ancien milieu avec une pipette de 1000 μL d’un côté du plat. Assurez-vous que la puce reste mouillée en tout temps.
- Ajouter 2 mL de DPBS-/- détacher doucement et goutte-sage la plupart des cellules mortes qui sont attachées aux cellules à motifs, à partir du centre de la puce, puis se déplaçant sur ses côtés.
- Aspirez la plupart du DPBS à enlever autant de cellules flottantes détachées que possible, en commençant par le centre de la puce, puis en se déplaçant sur les côtés.
- Répétez les étapes 5.1.2 et 5.1.3 une fois de plus.
- Utilisez des forceps inclinés pour saisir le bord de la puce et transférez-le immédiatement dans une nouvelle boîte greiner petri stérile de 35 mm pour réduire le nombre de cellules flottantes lors de la cueillette.
- Ajouter immédiatement 1,5 mL de DPBS-/- de sorte que la puce ne se dessèche pas.
- Ajouter 1,5 μL de vert vibrant cycle de teinture pour visualiser les noyaux des cellules vivantes.
- Placez la puce au centre de la boîte de Greiner Petri en utilisant la pointe des forceps.
- Mettez une chambre (Table of Materials) sur la puce. La chambre fixera la puce au fond du plat, sans bloquer l’accès aux modèles cellulaires.
- Montez la boîte greiner petri sur le porte-vaisselle de l’étape du cueilleur de cellules et insérez le bouchon magnétique.
- Calibrez l’injection automatisée.
- Localisez le réticule, qui est gravé sur la scène motorisée au milieu de l’image dans la fenêtre Live View.
- Concentrez-vous sur le réticule et sélectionnez l’étalonnage pour le bouton d’injection automatisé dans la fenêtre de balayage et de tri.
- Remplacer le DPBS-/- par 1,5 mL de DPBS/trypsine-/- (1:1) pendant que le plat est sur scène, pour desserrer les cellules de la fibronectine afin qu’un vide fluidique puisse être utilisé pour ramasser les cellules.
- Scannez l’ensemble de la puce à l’aide de l’onglet Numérisation dans la fenêtre de balayage et de tri. Localisez le coin supérieur gauche de la puce dans le champ de vision et cliquez sur Obtenir la position actuelle du microscope dans la rangée d’angle supérieur gauche.
- Ensuite, déplacez la scène motorisée vers le coin inférieur droit de la puce. Concentrez le microscope et cliquez sur Obtenir la position actuelle du microscope dans la rangée d’angle inférieur droit.
- Cliquez sur le bouton set plan le plus pointu et sur la fenêtre surgi cliquez sur le Go vers le coin supérieur droit. Concentrez le microscope et cliquez sur Allez dans le coin inférieur gauche et réglez la mise au point. Une fois terminé, cliquez sur le bouton Finition et commencez à numériser.
- Lorsque la numérisation est terminée, allez à l’onglet Analyse et sélectionnez les cellules individuelles qui passent les critères d’étude.
- Assurez-vous que le microcapillaire en verre est au milieu de la vue en direct du microscope.
- Contrôlez la pompe à seringues à l’aide de la fenêtre Pompe du logiciel. Créer un vide en retirant 4 mL d’une seringue numéro 1 de 50 mL, d’un diamètre de 27 mm.
- Dans l’onglet Tri
- Définissez les paramètres d’injection des valves. On a calculé que le volume d’injection qui a livré une seule cellule choisie était de 1 μl, si la valve 2 a été ouverte pendant 120 millisecondes, puis la valve 1 ouverte pendant 20 millisecondes après un laps de temps de 200 millisecondes. En raison de l’élasticité des tubes, ouvrez la valve 1 pour arrêter d’injecter le flux.
- Définissez les paramètres de ramasseuse des vannes. Il a été calculé que si la valve 1 a été ouverte pendant 20 millisecondes, puis la valve 2 a été ouverte pendant 10 millisecondes, après un laps de temps de 10 millisecondes, la plupart des cellules modelées peuvent être ramassées. C’est parce que leurs fixations de fibronectine ont été desserrées après avoir été traitées par trypsine.
- Cliquez sur le bouton Calculer le chemin. Le logiciel calcule le chemin le plus rapide d’une cellule à l’autre, pour ramasser et injecter les cellules sélectionnées dans toute la puce.
- Concentrez le microscope sur une cellule à motifs sur la surface de la puce.
- À l’aide du joystick, déplacez soigneusement le microcapillaire vers le bas, de sorte que l’image la plus nette de la pointe du microcapillaire puisse être obtenue sans toucher la cellule.
- Cliquez sur le bouton Définir dans la section offset Micropipette. Une nouvelle fenêtre apparaîtra, montrant la section transversale microcapillaire. Cliquez sur le centre exact du capillaire. Le logiciel enregistrera ensuite le décalage de pointe du capillaire dans les coordonnées x, y et z.
- Lancer le tri à l’aide du bouton de tri Démarrer.
Résultats
Le tissu a été disséqué du ventricule gauche des coeurs néonatals de rat de 2 jours et divisé en cellules simples. Ensuite, les MC enrichis ont été ensemencés sur une puce contenant des modèles de fibronectine avec des AR distincts. Après 72 heures de culture, le milieu a été remplacé par 1:1000 Vibrant Dye Cycle vert en DPBS-/- pendant 2 min pour visualiser les noyaux des cellules vivantes. Ensuite, les cellules ont été traitées avec DPBS-/-/trypsine (1:1) pour desserrer les cellules de la fibronectine, de sorte qu’un vide fluidique pourrait être utilisé pour faciliter la cueillette cellulaire. Pendant ce temps, la puce entière a été scannée à un grossissement de 10x, à l’aide d’un microscope inversé relié au cueilleur de cellules. Ceci a été effectué avant que les cellules soient arrondies dues au traitement de trypsine. Les cellules qualifiées ont été sélectionnées, en fonction de l’image numérisée, et leurs coordonnées ont été enregistrées dans le logiciel de ramasseur de cellules. Les micropatrins n’ont été sélectionnés que s’ils contenaient une cellule unique mononucléée et seulement lorsque la cellule couvrait entièrement son micropatrin fibronectine. Le trieur de cellules a choisi les cellules sélectionnées une par une et chaque cellule qui a été cueillie avec succès a été immédiatement injectée dans un tube PCR individuel et placée au stade du microscope. Chaque tube PCR contenait 3,55 μL de tampon de lyse (tableau 2). Le processus de tri, qui a commencé par la suppression des médias, a été achevé dans les 40 minutes. La synthèse complémentaire de l’acide désoxyribonucléique (ADN), la pré-amplification et la purification du PCR ont été effectuées sur les cellules individuelles lysées, basées sur le protocole Smart-Seq224 (tableau 3). La qualité de la CDNA purifiée a été vérifiée par un analyseur automatisé d’électrophoresis. L’électrophéogramme de l’ADND pré-amplifié d’une seule cellule choisie est présenté à la figure 4. Les bibliothèques RNA-Seq ont été préparées selon le protocole Smart-Seq224.
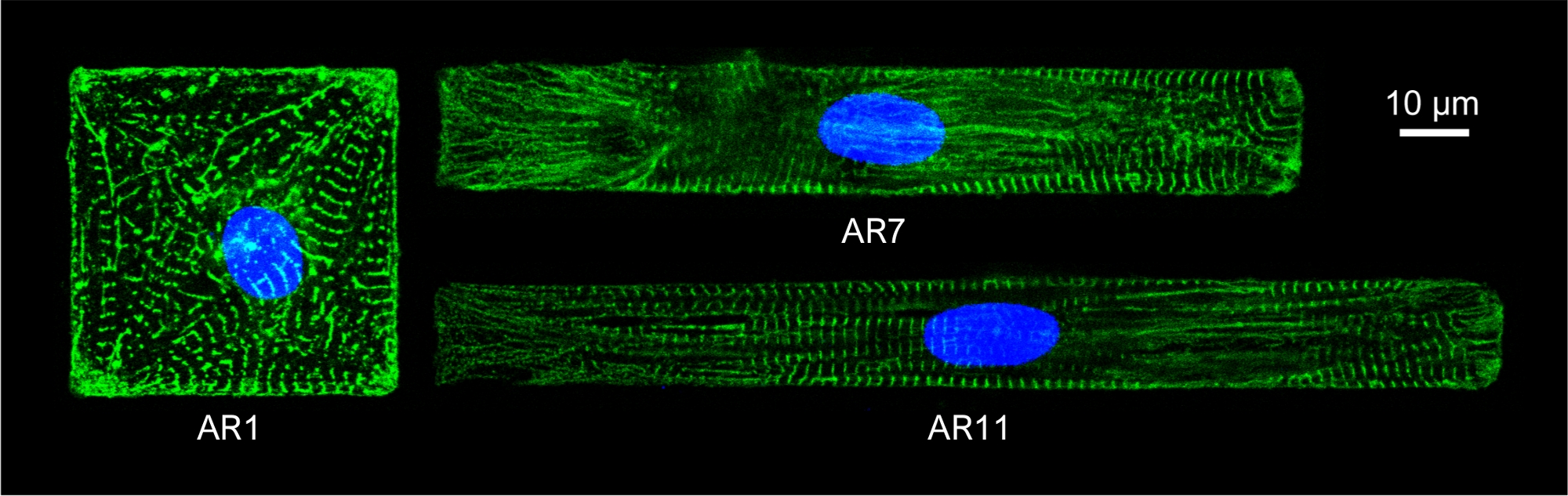
Pour observer la structure sarcomere des MC à motifs, les MC à motifs ont été tachés d’anticorps sarcomeric α-actine. Les cellules ont été incubées avec Donkey Anti-Mouse IgG Alexa Fluor 488 1:800 pendant 1 heure à température ambiante pour la coloration secondaire. Les noyaux ont été tachés avec 1 μg/mL DAPI. Des images immunofluorescentes ont été acquises avec un microscope confocal inversé, à l’aide d’un objectif d’immersion d’huile de 63 x (NA 1.4) (figure 5).

Figure 1 : Disposition de la puce avec des micropatrins de fibronectine.
(A) Image de la puce sur mesure. La puce est un coverslip de 19,5 mm x 19,5 mm avec micropatterns fibronectin, imprimé par photolithographie sur un verre borosilicate. (B) Mise en page des copeaux. La puce est divisée en trois zones et chaque zone se compose de micropatrins de fibronectine avec ar spécifique. Des images fluorescentes de différentes formes de micropatrins de fibronectine sont affichées en vue agrandie pour chaque zone. Ce chiffre a été modifié à partir du «matériau supplémentaire 1» par Haftbaradaran Esfahani et coll.2, utilisé sous http://creativecommons.org/licenses/by/4.0/ . S’il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 2 : Dissociateur équipé d’appareils de chauffage.
(A) L’instrument de dissociateur entier utilisé pour la dissociation entièrement automatisée des ventricules gauches néonatals de rat de 2 jours. (B) Unité de chauffage. (C) Tube rotor-capuchon. (D) Programmes prêts à l’emploi pour un flux de travail entièrement automatisé de dissociation des tissus. S’il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 3 : Appareil de ramasseur de cellules.
(A) Vue agrandie de l’étape motorisée du cueilleur de cellules. Un porte-boîte Petri et 80 trous pour 10 bandes PCR et un trou pour étalonnage crosshair est intégré sur la scène. (B) Vue agrandie d’un microcapillaire en verre. (C) La pompe à seringues. (D) L’unité de commande, qui contrôle la fenêtre d’ouverture et de fermeture des vannes 1 et 2, montée à l’intérieur de l’unité de commande. S’il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 4 : L’électrophéogramme de l’ADND pré-amplifié d’une seule cellule choisie.
19 cycles pcr de pré-amplification ont été utilisés pour obtenir 15 μL de rendement cDNA purifié de 1 ng/μL. On observe une bande claire dans la parcelle de densitométrie gel-like qui correspond au pic à 1852 bp dans l’électrophéogramme. La taille moyenne des fragments est de 1588 pb. En outre, la petite quantité de fragments qui sont plus courts que 300 bp indique une bonne bibliothèque cDNA. S’il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 5 : Coloration immunofluorescente de la structure sarcomerique α-actinine (verte) et du noyau (bleu) des MC à motifs avec différents AR.
La chromatine a été tachée par DAPI. S’il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
{kind=link}
| Morphotype | Ar | Longueur (μm) | Largeur (μm) | Superficie fibronectine (μm2) |
| AR1 (AR1) | 1:1 | 47 | 47 | 2209 |
| AR7 (ar7) | 7:1 | 126 | 18 | 2268 |
| AR11 (AR11) | 11:1 | 155 | 14 | 2170 |
Tableau 1 : Géométrie des MC à motifs.
| Composant | Volume (μL) |
| Eau sans noyaux | 0.65 |
| (0,4% vol/vol) Triton X-100 | 1.8 |
| mélange dNTP (25 mM) | 0.8 |
| Inhibiteur de la RNase (40 U μL-1) | 0.1 |
| Oligo-dT30Oligonucleotides VN (100 μM) | 0.1 |
| ERCC ARN Spike-In Mix (2,5 x 105 dilution) | 0.1 |
| Cellule unique injectée | 1 |
| Volume total | 4.55 |
Tableau 2 : Tampon de lyse personnalisé à cellule unique.
| Composant | Volume (μL) |
| Tampon de premier brin Superscript II (5x) | 2 |
| TNT (100 mM) | 0.5 |
| Bétaïne (5 M) | 2 |
| Mgcl2 (1 M) | 0.1 |
| Inhibiteur de la RNase (40 U μL-1) | 0.25 |
| Transcriptase inverse Superscript II (200 U μL-1) | 0.5 |
| TSO (100 μM) | 0.1 |
| Volume total | 5.45 |
Tableau 3 : Mélange de transcription inverse (RT) pour une réaction rt pour synthétiser l’ADND du premier brin à partir du lysate d’un seul CM.
Discussion
Cette étude a utilisé le séquençage de l’ARN à cellules simples, qui est une technologie nouvelle et puissante qui peut détecter le transcriptome des cellules individuelles. Il a été combiné avec une approche novatrice de la culture des MC unique, de sorte qu’ils ont pris différents ARs qui, autrement, n’auraient pu être observés in vivo.
L’étude avait certaines limites. Par exemple, les MC néonatals ont dû être utilisés pour générer différents morphotypes, car il est exceptionnellement difficile de culture suffisamment vitale mc adultes pendant 72 heures dans des formes définies. En outre, les MC ont été cultivés pendant 72 heures ex vivo, ce qui aurait pu avoir un impact sur le modèle d’expression des gènes. Cependant, cette culture était nécessaire, de sorte que les cellules puissent former des morphotypes spécifiques. En outre, seules les cellules simples qui ont été mononucléées et entièrement couvert le micropattern fibronectine ont été sélectionnés pour le tri. Les cellules à motifs sur chaque puce doivent être triées simplement en un seul tour de tri. Enfin, environ 50 cellules, soit environ un tiers des cellules sélectionnées, ont été captées avec succès à partir de chaque puce. Il y a deux raisons qui ont limité le nombre de cellules choisies avec succès. Tout d’abord, certaines cellules étaient trop étroitement attachées au modèle de fibronectine et le flux de ramassage n’était pas assez forcé pour réussir à les ramasser. Deuxièmement, en raison du traitement à la trypsine, l’attachement entre certaines cellules et la fibronectine est devenu trop lâche. Par conséquent, ces cellules ont été repoussées loin de leurs micropatterns de fibronectine, quand le microcapillaire s’est approché d’eux, et elles n’ont pas été ramassées. Les auteurs ne prétendent pas que cette configuration est la même qu’un environnement in vivo, mais il s’est avéré être une approche viable pour répondre à la question de recherche.
La méthode proposée s’applique aux différents types de cellules (p. ex., pour les mcs hiPS). Toutefois, les facteurs suivants devraient être optimisés pour étudier d’autres types de cellules. Des molécules adhésives ECM appropriées pour l’attachement du type cellulaire spécifique doivent être utilisées pour enduire les micropatrins. La géométrie des micropatrins doit être modifiée en fonction de la question de l’étude et du type cellulaire. La période de culture peut être modifiée en fonction de la question de l’étude. Le réaccente de détachement et son temps d’incubation doivent être optimisés précisément pour le type de cellule d’étude. Par exemple, Accutase peut être utilisé au lieu de TryplE pour le détachement de cellules souches embryonnaires et neuronales. Les paramètres de temps d’ouverture des valves doivent être scrutés à la loupe pour choisir les cellules avec succès, mais doucement. En résumé, nous avons conçu une nouvelle plate-forme pour étudier la forme cellulaire qui peut fournir une ressource précieuse pour les chercheurs dans le domaine. Dans ce contexte, nous avons conçu une approche expérimentale qui imitait les formes caractéristiques in vitro imposées au CM in vivo par des contraintes hémodynamiques pour identifier l’interaction entre l’architecture cellulaire et l’expression génique. Nous rapportons également le développement d’une nouvelle plate-forme pour étudier hf in vitro et l’identification de la forme cellulaire comme un déterminant puissant de l’expression des gènes. Il s’agit d’une nouvelle observation qui a des implications profondes pour la biologie et la médecine.
Déclarations de divulgation
Aucun.
Remerciements
Aucun.
matériels
| Name | Company | Catalog Number | Comments |
| 2100 Bioanalyzer Instrument | Agilent Technologies | G2939BA | Automated electrophoresis analyzer |
| Anti-Red Blood Cell MicroBeads | Miltenyi Biotec | 130-109-681 | |
| Axio Observer microscope | Zeiss | Z1 | Inverted microscope |
| Betaine solution (5 M) | Sigma-Aldrich, MERCK | B0300 | |
| CellSorter | CELLSORTER | https://www.singlecellpicker.com/ | Cell picker |
| CYTOOchamber | CYTOO | 30-010 | Custom-designed chip |
| CYTOOchip | CYTOO | 10-950-00-18 | Chamber |
| DMEM | Thermo Fisher Scientific | 31966-021 | high glucose, GlutaMAX Supplement |
| dNTP mix (25 mM) | Thermo Fisher Scientific | R1122 | |
| Donkey Anti-Mouse IgG Alexa Fluor 488 | Abcam PLC | ab150105 | |
| DTT (100 mM) | Thermo Fisher Scientific | 18064071 | |
| ERCC RNA Spike-In Mix | Thermo Fisher Scientific | 4456740 | |
| Fetal Bovine Serum | Thermo Fisher Scientific | 10082-147 | |
| Fibronectin | Sigma-Aldrich, MERCK | F4759 | |
| gentleMACS C Tube | Miltenyi Biotec | 130-093-237 | Rotor-cap tube |
| gentleMACS Octo Dissociator with Heaters | Miltenyi Biotec | 130-096-427 | Dissociator with heater |
| Greiner CELLSTAR Petri dish | Sigma-Aldrich, MERCK | P6987 | |
| HEPES (1 M) | Thermo Fisher Scientific | 15630-056 | |
| Horse Serum | Sigma-Aldrich, MERCK | H0146 | |
| LD column | Miltenyi Biotec | 130-042-901 | |
| Medium 199 | Thermo Fisher Scientific | 31150-022 | |
| NE-1000 syringe pump | New Era Pump Systems | NE-1000 | Syringe pump |
| Neonatal Cardiomyocyte Isolation Cocktail, rat | Miltenyi Biotec | 130-105-420 | |
| Neonatal Heart Dissociation Kit, mouse and rat | Miltenyi Biotec | 130-098-373 | |
| Oligo-dT30VN oligonucleotides | IDT Technology | 5′–AAGCAGTGGTATCAACGCAGAGTACT30VN-3′ | |
| RNAse inhibitor (40 U µL-1) | Clontech | 2313A | |
| sarcomeric α-actinin | Sigma-Aldrich, MERCK | EA-53 | |
| SP8 confocal microscope | Leica Microsystems | SP8 | Confocal microscope |
| Superscript II first-strand buffer (5x) | Thermo Fisher Scientific | 18064071 | |
| Superscript II reverse transcriptase (200 U µL-1) | Thermo Fisher Scientific | 18064071 | |
| Triton X-100 | Sigma-Aldrich, MERCK | T9284 | |
| TryplE Express enzyme, no phenol red | Thermo Fisher Scientific | 12604013 | |
| TSO (100 µM) | QIAGEN | 5′-AAGCAGTGGTATCAACGCAGAGTACATrGrG+G-3′ | |
| Vibrant Dye Cycle green | Thermo Fisher Scientific | V35004 |
Références
- Heineke, J., Molkentin, J. D. Regulation of cardiac hypertrophy by intracellular signalling pathways. Nature Reviews Molecular Cell Biology. 7 (8), 589-600 (2006).
- Haftbaradaran Esfahani, P., et al. Cell shape determines gene expression: cardiomyocyte morphotypic transcriptomes. Basic Research in Cardiology. 115 (1), 7(2019).
- Kontrogianni-Konstantopoulos, A., Benian, G., Granzier, H. Advances in Muscle Physiology and Pathophysiology 2011. Journal of Biomedicine and Biotechnology. 2012, 930836(2012).
- Hill, J. A., Olson, E. N. Cardiac plasticity. New England Journal of Medicine. 358 (13), 1370-1380 (2008).
- Bray, M. A., Sheehy, S. P., Parker, K. K. Sarcomere alignment is regulated by myocyte shape. Cell Motility and the Cytoskeleton. 65 (8), 641-651 (2008).
- Benjamin, I. J., Schneider, M. D. Learning from failure: congestive heart failure in the postgenomic age. Journal of Clinical Investigation. 115 (3), 495-499 (2005).
- Opie, L. H., Commerford, P. J., Gersh, B. J., Pfeffer, M. A. Controversies in ventricular remodelling. Lancet. 367 (9507), 356-367 (2006).
- Braunwald, E. Cardiomyopathies: An Overview. Circulation Research. 121 (7), 711-721 (2017).
- Knoll, R., et al. The cardiac mechanical stretch sensor machinery involves a Z disc complex that is defective in a subset of human dilated cardiomyopathy. Cell. 111 (7), 943-955 (2002).
- Frangogiannis, N. G. The Extracellular Matrix in Ischemic and Nonischemic Heart Failure. Circulation Research. 125 (1), 117-146 (2019).
- Marian, A. J., Braunwald, E. Hypertrophic Cardiomyopathy: Genetics, Pathogenesis, Clinical Manifestations, Diagnosis, and Therapy. Circulation Research. 121 (7), 749-770 (2017).
- Mozaffarian, D., Caldwell, J. H. Right ventricular involvement in hypertrophic cardiomyopathy: a case report and literature review. Clinical Cardiology. 24 (1), 2-8 (2001).
- Olsson, M. C., Palmer, B. M., Stauffer, B. L., Leinwand, L. A., Moore, R. L. Morphological and functional alterations in ventricular myocytes from male transgenic mice with hypertrophic cardiomyopathy. Circulation Research. 94 (2), 201-207 (2004).
- Toepfer, C. N., et al. Hypertrophic cardiomyopathy mutations in MYBPC3 dysregulate myosin. Science Translational Medicine. 11 (476), (2019).
- McKenna, W. J., Maron, B. J., Thiene, G. Classification, Epidemiology, and Global Burden of Cardiomyopathies. Circulation Research. 121 (7), 722-730 (2017).
- Schultheiss, H. P., et al. Dilated cardiomyopathy. Nature Reviews Disease Primers. 5 (1), 32(2019).
- Knoll, R. A role for membrane shape and information processing in cardiac physiology. Pflugers Arch. 467 (1), 167-173 (2015).
- Rangamani, P., et al. Decoding information in cell shape. Cell. 154 (6), 1356-1369 (2013).
- Kuo, P. L., et al. Myocyte shape regulates lateral registry of sarcomeres and contractility. American Journal of Pathology. 181 (6), 2030-2037 (2012).
- Gerdes, A. M., Onodera, T., Wang, X., McCune, S. A. Myocyte remodeling during the progression to failure in rats with hypertension. Hypertension. 28 (4), 609-614 (1996).
- Kehat, I., et al. Extracellular signal-regulated kinases 1 and 2 regulate the balance between eccentric and concentric cardiac growth. Circulation Research. 108 (2), 176-183 (2011).
- Sreejit, P., Kumar, S., Verma, R. S. An improved protocol for primary culture of cardiomyocyte from neonatal mice. In Vitro Cellular & Developmental Biology - Animal. 44 (3-4), 45-50 (2008).
- Kornyei, Z., et al. Cell sorting in a Petri dish controlled by computer vision. Scientific Reports. 3, 1088(2013).
- Picelli, S., et al. Full-length RNA-seq from single cells using Smart-seq2. Nature Protocols. 9 (1), 171-181 (2014).
- Schmick, M., Bastiaens, P. I. H. The interdependence of membrane shape and cellular signal processing. Cell. 156 (6), 1132-1138 (2014).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.