
Method Article
ספקטרוסקופיית מתאם פלואורסצנטית של וריאציה נקודתית לניתוח דיפוזיה מולקולרית בקרום הפלזמה של תאים חיים
* These authors contributed equally
In This Article
Summary
מאמר זה נועד להציג פרוטוקול כיצד לבנות מיקרוסקופ ספקטרוסקופיה של מתאם פלואורסצנטי (svFCS) של וריאציה נקודתית כדי למדוד דיפוזיה מולקולרית בקרום הפלזמה של תאים חיים.
Abstract
תהליכים ביולוגיים דינמיים בתאים חיים, כולל אלה הקשורים לארגון קרום הפלזמה, מתרחשים בקני מידה מרחביים וטמפורליים שונים, החל מננומטרים ועד מיקרומטרים ומיקרו-שניות ועד דקות, בהתאמה. מגוון כה רחב של תהליכים ביולוגיים מאתגר גישות מיקרוסקופיות קונבנציונליות. כאן אנו מפרטים את הנוהל ליישום מדידות ספקטרוסקופיה של מתאם פלואורסצנטי (svFCS) של וריאציות נקודתיות באמצעות מיקרוסקופ פלואורסצנטי קלאסי שהותאם אישית. הפרוטוקול כולל בדיקת ביצועים ספציפית של מערך svFCS ואת ההנחיות למדידת דיפוזיה מולקולרית על ידי svFCS על קרום הפלזמה של תאים חיים בתנאים פיזיולוגיים. בנוסף, אנו מספקים הליך לשיבוש ננו-דומיינים של רפסודות ממברנת פלזמה על ידי טיפול בכולסטרול אוקסידאז ומדגימים כיצד שינויים אלה בארגון הלטרלי של קרום הפלזמה עשויים להתגלות על ידי ניתוח svFCS. לסיכום, שיטה פלואורסצנטית זו יכולה לספק פרטים חסרי תקדים על הארגון הלטרלי של קרום הפלזמה עם הרזולוציה המרחבית והזמנית המתאימה.
Introduction
המורכבות של ארגון ממברנת הפלזמה
ההבנה הנוכחית של ארגון קרום התא צריכה לקחת בחשבון מספר היבטים1. ראשית, הרכב שומנים מורכב משתנה לא רק בין סוגי תאים, אלא גם בתוך תא בודד (אברוני ממברנה/קרום פלזמה). חוץ מזה, חלבוני ממברנה קשורים או פנימיים מאורגנים ברובם בקומפלקסים דינמיים רב-ימיים, עם תחומים גדולים המשתרעים מחוץ לממברנה, ומהווים שטח גדול משמעותית מזה של תחומי הטרנס-ממברנה בלבד. יתר על כן, חלבונים הקשורים לממברנה מפגינים יכולות ספציפיות של קשירת שומנים או אינטראקציה עם שומנים, הממלאות תפקידים בוויסות תפקוד החלבון. אלה תלויים ישירות בהרכב המקומי ובנגישות של השומנים2.
לבסוף, רמה משמעותית של אסימטריה נצפתה בין שני עלוני ממברנה בשל המבנה האסימטרי הפנימי של חלבוני הממברנה והתפלגות השומנים. ואכן, איזון מטבולי של שומנים בין סינתזה להידרוליזה, בשילוב עם כפכף שומנים בין העלונים, יוצר התפלגות אסימטרית כגון. מכיוון שכל הובלה על פני הדו-שכבתית מוגבלת על ידי האנרגיה החופשית הנדרשת כדי להזיז את קבוצת ראש הקוטב דרך הפנים ההידרופוביות של הממברנות, היא בדרך כלל נעזרת במובילים סלקטיביים. עבור כל סוג תא, אסימטריה נוטה להישמר היטב. בסך הכל, גורמים אלה תורמים לאי-הומוגניות צידית או למידור של קרום הפלזמה 3,4.
אנו מעשירים את הייצוג הזה של קרום הפלזמה על ידי התחשבות בדיפוזיה המולקולרית הפנימית בתוך ולרוחב הדו-צדדי, מה שתורם להטרוגניות הצידית הדינמית בסולם של עשיריות עד מאות ננומטרים ומיקרו-שניות לשניות. לדוגמה, ננו-דומיינים של ממברנה התלויה בשומנים – מה שמכונה רפסודות ליפידים, המוגדרות ככולסטרול, ופלטפורמות איתות עשירות בספינגוליפידים – תורמות למידור קרום הפלזמה 5,6. עם זאת, ההשקפה הנוכחית של ארגון הממברנה אינה מוגבלת לרפסודות שומנים בלבד. ננו-דומיינים של ממברנה הם מורכבים יותר והטרוגניים יותר בהרכב, במקור ובתפקוד. ובכל זאת, נוכחותם בקרום הפלזמה חייבת להיות מתואמת היטב, ונראה כי אינטראקציות דינמיות בין חלבונים ושומנים חשובות בחלוקה המרחבית ובשינוי הכימי של ננו-דומיינים של ממברנה 1,3,7,8.
עקרון svFCS ויישומו כדי לחקור את הארגון של קרום הפלזמה
למרות התקדמות רבה נעשתה בניתוח של תחומי ממברנה, בעיקר באמצעות טכניקות ביופיזיות, הדטרמיננטות המכתיבות את הארגון המקומי של קרום הפלזמה צריכות להיות מעודנות עם רזולוציה מרחבית וטמפורלית מתאימה. דטרמיננטות המבוססות על מעקב אחר מולקולות בודדות מספקות דיוק מרחבי מצוין ומאפשרות אפיון של מצבי תנועה שונים 9,10,11,12, אך יש להן רזולוציה טמפורלית מוגבלת עם קצבי פריימים נמוכים קלאסיים של מצלמה ודורשות מאמץ ניסיוני רב יותר כדי לתעד מספר משמעותי של מסלולים. לחלופין, ניתן להעריך את מקדם הדיפוזיה של רכיבי הממברנה על ידי התאוששות פלואורסצנטית לאחר פוטו-הלבנה (FRAP)13 או ספקטרוסקופיה של מתאם פלואורסצנטי (FCS)14. האחרון זכה לתשומת לב רבה יותר, בעיקר בגלל הרגישות הגבוהה והסלקטיביות שלו, נפח הגילוי המיקרוסקופי, הפולשנות הנמוכה והטווח הדינמי הרחב15.
הבסיס הרעיוני של FCS הוצג על ידי מגד ועמיתיו לפני כ-50 שנה, לפני 16,17 שנה. הוא מבוסס על רישום התנודתיות של פליטת פלואורסצנציה ברזולוציה טמפורלית גבוהה (מ-μs ל-s)18. בגרסתו המודרנית, מדידות בתאים חיים מבוצעות על ידי נפח עירור קונפוקלי קטן (~0.3 פמטוליטרים) הממוקם בתוך אזור עניין (למשל, בקרום הפלזמה); האות הפלואורסצנטי שנוצר על ידי פיזור מולקולות פלואורסצנטיות הנכנסות ויוצאות מנפח התצפית נאסף ברזולוציה טמפורלית גבוהה מאוד (כלומר, זמן הגעתו של כל פוטון לגלאי). לאחר מכן, האות מחושב כדי ליצור את פונקציית התיקון האוטומטי (ACF), שממנה מופק הזמן הממוצע td (זמן דיפוזיה) שעבורו מולקולה נשארת בתוך נפח המוקד, יחד עם המספר הממוצע של חלקיקים, (N), הנמצא בנפח התצפית, שהוא ביחס הפוך למשעת של ACF. פרמטר אחרון זה עשוי להיות מידע שימושי על ריכוז המולקולה בתוך נפח התצפית.
מאז, מספר גדל והולך של שיטות FCS יושמו הודות למכשור המתפתח במהירות בביופוטוניקה, המאפשר תיאור של תופעות דינמיות המתרחשות במערכות חיות. ובכל זאת, מין מולקולרי יחווה התפלגות חופפת יותר של ערכי מקדם הדיפוזיה, אשר משתקפת בדרך כלל על ידי מאפיין דיפוזיה חריג, שבו מולקולות מתפזרות עם קשר לא ליניארי בזמן19, וקושי לזהות את המשמעות הביולוגית של תת-דיפוזיה חריגה זו. בעבר, קושי זה התגבר במידה מסוימת על ידי תיעוד הדיפוזיה המולקולרית על ידי FRAP מאזורים בגדלים שונים, ולא מאזור אחד בלבד, ובכך סיפק מידע מרחבי נוסף. זה איפשר, למשל, את ההמשגה של מיקרו-דומיינים של ממברנות 20,21,22.
תרגום של אסטרטגיה זו למדידות FCS (כלומר, מה שמכונה וריאציה נקודתית ספקטרוסקופיה של מתאם פלואורסצנטי (svFCS)) נקבע על ידי שינוי גודל נפח המוקד של התצפית, מה שאפשר לתנודתיות בפלואורסצנציה להירשם בקני מידה מרחביים שונים23. לפיכך, גישת svFCS מספקת מידע מרחבי עקיף המאפשר זיהוי וקביעה של מצבי דיפוזיה מולקולרית וסוג של חלוקת ממברנות (תחומים מבודדים לעומת רציפים24) של מולקולות שנחקרו. על ידי התוויית זמן הדיפוזיה td כפונקציה של הסולמות המרחביים השונים המוגדרים על ידי ערך המותניים (ω), התואם את גודל רדיוס קרן הגילוי במקרה זה23,25, ניתן לאפיין את חוק הדיפוזיה של מולקולה נתונה במצב פיזיולוגי נתון. ה-svFCS הוא, אם כן, אנלוגי מושלם למעקב אחר חלקיקים בודדים בתחום הזמן26. תחת אילוץ הדיפוזיה הבראונית, יש לצפות לקשר ליניארי לחלוטין בין זמן הדיפוזיה td לבין המותניים ω (איור 1)23,25. ניתן לייחס את מקור הסטייה של חוק הדיפוזיה מתוכנית זו לסיבות לא בלעדיות, כגון עבודת רשת שלד ציטוסקלטון, צפיפות מולקולרית, חלוקה דינמית בננו-דומיינים, או כל שילוב של השפעות אלה ואחרות (איור 1), ויש לבחון אותה באופן ניסיוני25.
כאן אנו מספקים את כל מחסומי הבקרה הדרושים לשימוש יומיומי במערכת אופטית svFCS מותאמת אישית הבנויה מאפס, המשלימה את סקירות הפרוטוקול הקודמות שלנו 27,28 על גישה ניסיונית זו. יתר על כן, כהוכחת היתכנות, אנו נותנים קווים מנחים לגבי כיול ההתקנה, הכנת תאים, איסוף נתונים וניתוח להקמת חוק הדיפוזיה svFCS (DL) עבור Thy1-GFP, חלבון מעוגן גליקוסילפוספטידילינוזיטול עם קרום פלזמה, אשר ידוע כמיקום בננו-דומיינים של רפסודת שומנים29. לבסוף, אנו מדגימים כיצד התערערות חלקית של ננו-דומיינים של רפסודות שומנים על ידי טיפול באוקסידאז כולסטרול משפיעה על תכונות הדיפוזיה של Thy1-GFP. בנוסף, תיאור מפורט של בניית מערך svFCS מאפס מסופק בחומר משלים.
Protocol
1. הגדרת מפרט להרכבת הגדרת svFCS מותאמת אישית
הערה: הפשטות של הגדרת svFCS המוצעת מאפשרת התקנה, הפעלה ותחזוקה קלות בעלות נמוכה תוך הבטחת יעילות בשחזור הפוטונים. לפרטים נוספים, ראו חומר משלים.
- חדר ניסויים ובטיחות
- התקן את המערכת בחדר התייצב סביב 21 °C (76 °F).
- הימנע מזרימת אוויר ישירה על הטבלה האופטית הפסיבית (או האקטיבית) ופעל לפי כללי בטיחות הלייזר ליישור אופטי.
- חומרה ותוכנה
הערה: חומר משלים מפרט את שלבי ההתקנה המתוארים באיור 2.- כתוב את תוכנת הרכישה והבקרה העיקרית ב- LabVIEW באמצעות מכונת מצבים וארכיטקטורת מבנה אירועים שבה לוח רכישה רב-תכליתי מניע את רוב הבקרים.
הערה: המתאם, הלייזר ומד ההספק נשלטים או מנוטרים על ידי התוכנה שלהם. - התאם את נהלי התקנת החומרה והתוכנה בהתאם לחומרה שבה נעשה שימוש.
- כתוב את תוכנת הרכישה והבקרה העיקרית ב- LabVIEW באמצעות מכונת מצבים וארכיטקטורת מבנה אירועים שבה לוח רכישה רב-תכליתי מניע את רוב הבקרים.
- הגדרה אופטית
הערה: איור 3 ממחיש את מודולי הספסל האופטי המשמשים בסעיפים הבאים כדי לשלוט באיכות היישורים האופטיים. כל מפרטי האלמנטים האופטיים מפורטים בטבלת החומרים. הנוהל לבניית ההתקנה מפורט באופן נרחב בחומר משלים. מערכת זו כוללת לייזר גלים רציף, מיקרוסקופ הפוך ממונע המצויד במטרה של מי טבילה, גלאי פוטודיודה של מפולת שהוצמד למודול ספירת פוטונים יחיד, ומתאם חומרה. תא דגירה של מיקרוסקופ עם תנורי חימום נטולי רעידות תוכנן במיוחד כדי לשלוט בטמפרטורה לניסויים על תאים חיים. על פי המוסכמה, ציר XY מתאים למישור הניצב של הנתיב האופטי, וציר ה-Z מתאים לנתיב האופטי.
2. מחסום יומי לפני ביצוע הניסוי
- שלוט בנתיב העירור (איור 3,
 &
&  ).
).- פתחו את כל הסרעפת של הקשתית.
- מדוד את עוצמת הלייזר באמצעות מד הכוח, תוך שמירה על הקשתית הראשונה פתוחה לחלוטין.
- סובב את לוחית חצי הגל (HWP) כדי למצוא את ההספק המרבי.
- בדוק את היישור באמצעות הקשתיות אם כוח הלייזר נמוך מהרגיל, והזז L1 ו- M1 לסירוגין, במידת הצורך.
- שים לב לערך ההספק במחברת מעבדת הניסויים.
- שלוט בנתיב הזיהוי (איור 3,
 ו
ו  ).
).- מקם את המים, כיסוי, וטיפה של תמיסת רודמין 6G (Rh6G) של 2 ננומטר על המטרה.
- אם האות הפלואורסצנטי (מספר הספירה ב-APD, שהוקלט באמצעות תוכנת LabVIEW) נמוך מהרגיל, צור מחדש את פתרון ה-Rh6G, בדוק את המיקום ואת מספר הכיסוי על העדשה האובייקטיבית, או בטל בועות, אם בכלל.
- אם האות הפלואורסצנטי עדיין נמוך מהרגיל, מקם את מד הכוח בתוך הנתיב האופטי כדי לחסום את הקרן.
- כבו את ה-APD (להלן, APD מתייחס ל-APD ולמודול ספירת הפוטונים הבודד).
- הסר את המדגם.
- נקו והחליפו את העדשה האובייקטיבית במטרה רפלקטיבית.
- בדוק את קרן הלייזר על המטרה הרפלקטיבית על ידי הסרת מד הכוח מנתיב האור. ודאו שהקרן של המטרה ממורכזת, וההשתקפות האחורית מגיעה לקשת הראשונה בקו (איור 3).
- אם לא, התאם את מיקום המרכז עם M2 או את ההשתקפות האחורית עם המראה הדיכרואית.
- אם צימוד המיקרוסקופ נכון, דחפו את העדשה האובייקטיבית לאחור, הוסיפו טיפת מים, כיסוי טיפה וטיפה של תמיסת Rh6G מרוכזת יותר (כלומר, 200 ננומטר), וקבעו הספק לייזר נמוך יותר מאשר עבור המדידות הקלאסיות (כמה μW).
- הפעילו את ה-APD ומטבו את יישור ה-APD וה-pinhole, לסירוגין, עם ברגי כוונון XYZ המתאימים להם תוך ניטור אות העוצמה (תוכנת LabVIEW).
- שנה את הכיסוי והוסף ריכוז נמוך יותר של Rh6G (2 ננומטר). הזיזו את חור הסיכה לאורך ציר ה-Z כדי למצוא מיקום שבו יחס הבהירות המולקולרית עולה, והמותניים הם מינימליים.
- סגור את הקשתית עד שהאות יורד למטה: גודל קרן הלייזר מגיע לגודל הצמצם האחורי של המטרה (כלומר, גודל המותניים המינימלי, ראה חומר משלים).
- הפעל את תוכנת המתאם ותקליט נתונים (ראה סעיף 7 להקלטת נתונים).
- בדוק את ה- ACF, שאמור להציג כמות נמוכה של רעש, תן גודל מותניים קטן, וקצב ספירה גבוה למולקולה לשנייה (ראה סעיף 7 לניתוח נתונים והערכת גודל המותניים).
3. שיקולים כלליים להקלטה וניתוח של נתוני svFCS
- תעד ותנתח את נתוני הפלואורסצנציה בעקבות סכימה כללית זו (ראה סעיפים 7, 8 ו-9): (1) הקלטה פלואורסצנטית ויצירת ACF (תוכנת מתאם), (2) השלכה בלתי צפויה של נתונים, ממוצע של נתונים שמורים, המתאימים למודל המתאים (עם תוכנת איגור פרו תוצרת בית), (3) עלילת חוק הדיפוזיה (תוכנת MATLAB תוצרת בית 1) ו-(4) השוואת חוקי דיפוזיה אופציונלית (תוכנת MATLAB תוצרת בית 2). התוכנות השונות זמינות על פי בקשה.
הערה: למתאם החומרה יש זמן דגימה מינימלי של 12.5 ns (כלומר, תדר דגימה של 80 MHz). הוא מספק רזולוציה טמפורלית הנמוכה לפחות ב-1,000 מזמן התושבים הטיפוסי של פיזור חופשי של מולקולה קטנה בתמיסה ו-106 קטנה מזמן הדיפוזיה של חלבוני הממברנה בתוך נפח תצפית קונפוקלי.
4. תרבית תאים וטרנספקציה
- זרעו את תאי ה-Cos7 בכיסוי של 8 תאים עם תחתית זכוכית בורוסיליקט מס' 1.0 בצפיפות של 10,000 תאים/באר באמצעות מדיום הנשרים המהונדסים (DMEM) שלם של Dulbecco בתוספת 5% סרום בקר עוברי, פניצילין (100 U/mL), סטרפטומיצין (100 U/mL) ו-L-גלוטמין (1 mM).
- תרבית את התאים בטמפרטורה של 37 מעלות צלזיוס באטמוספרה לחה המכילה 5% CO2 למשך 24 שעות.
- הסר את המדיום, הוסף 300 μL של מדיום התרבית השלם הטרי לכל באר, והקדים את התאים במשך 30 דקות בטמפרטורה של 37 מעלות צלזיוס.
- דילול 0.5 מיקרוגרם של הדנ"א הפלסמידי המקודד Thy-1 חלבון התמזג עם eGFP25 ב-50 μL של DMEM ללא סרום. מערבולת לזמן קצר כדי לערבב.
- דיללו 1.5 μL של מגיב טרנספקציית הדנ"א ב-50 μL של DMEM ללא סרום, וערבבו היטב את התמיסה.
- מוסיפים את מגיב הטרנספקציה המדולל ישירות לתמיסת הדנ"א המוכנה, ומערבבים את התרכובות באופן מיידי.
- דגירה של התערובת המוכנה למשך 10 עד 15 דקות בטמפרטורת החדר.
- הוסיפו 10 μL של קומפלקסים משולבים של ריאגנטים מסוג DNA/transfection שנופלים על המדיום בכל באר, והומוגניים על ידי סיבוב עדין של הצלחת.
- הדגירה של התאים בטמפרטורה של 37 מעלות צלזיוס עם 5% CO2 למשך 3 שעות.
- לאחר הדגירה, החליפו את המדיום המכיל קומפלקסים של ריאגנט דנ"א/טרנספקציה ב-400 μL של DMEM שלם טרי, ותרבו את התאים במשך 16 שעות לפני ניסוי svFCS.
5. הכנת תאים למדידות svFCS
- הסר את המדיום התרבותי.
- שטפו את התאים בעדינות פעמיים עד שלוש עם תמיסת המלח המאוזנת (HBSS) של האנק ללא סרום המכילה Ca2+ ו-Mg2+ בתוספת 10 mM (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid) (HEPES), pH 7.4 (HBSS/HEPES).
- שמור על התאים במאגר HBSS/HEPES במהלך כל רכישות svFCS.
6. טיפול תרופתי
- הסירו את מדיום התרבית ושטפו את התאים פעמיים עד שלוש פעמים עם HBSS ללא סרום בתוספת 10 mM HEPES, pH 7.4 (HBSS/HEPES).
- הדגירה של התאים עם תמיסת 1 U/mL של כולסטרול אוקסידאז (COase) ב-HBSS/HEPES buffer למשך שעה אחת ב-37 מעלות צלזיוס.
- הסר את התמיסה ושמור על התאים בנוכחות 0.1 U/mL של COase במאגר HBSS/HEPES תוך ביצוע מדידות svFCS.
7. כיול גודל ספוט
- מחממים מראש את תא המיקרוסקופ בטמפרטורה של 37 מעלות צלזיוס.
- הכן פתרון סטנדרטי של 2 ננומטר של Rh6G על ידי דילול סדרתי.
- טיפה 200 μL של תמיסת 2 ננומטר Rh6G על מכסה זכוכית המוצב על מטרת טבילת המים.
- הפעל את כל החומרה והתוכנה.
- מדידה והתאמת הספק קרן הלייזר של 488 ננומטר ל-300 μW. בהתאם לבהירות וליציבות הצילום של הגשושית הפלואורסצנטית שבה נעשה שימוש, התאימו את ההספק הזה בהתאם ל-(1) עוצמת הפלואורסצנציה (בתוכנת LabVIEW), שאמורה להיות יציבה, (2) צורת ה-ACF (על תוכנת המתאם), שאמורה להיות בעלת צורה קבועה לאורך זמן, ו-(3) הפרמטרים המתאימים המעניקים גודל מותניים קטן וקצב ספירה גבוה למולקולה (פוטונים למולקולה לשנייה, בדרך כלל כמה עשרות עד מאות פוטונים למולקולה לשנייה).
הערה: המשרעת של ה-ACF (הנקראת G(0)) היא ביחס הפוך למספר המולקולה (כלומר, ריכוז הגשושית הפלואורסצנטית). עבור כיול גודל המותניים, זהו פרמטר מועמד טוב לבקרת איכות. לכן, G(0) צריך להיות דומה עבור אותו ריכוז מיום ליום כפי שהוא מקשר את גודל המותניים ואת הריכוז. עבור מדידות תאים, מכיוון ש- FCS מדויק יותר עבור ריכוז נמוך, G(0) צריך להיות גבוה עבור מיצוי התאמת הפרמטרים המתאימה. - הגדר את יציאת מיקרוסקופ התאורה/זיהוי של svFCS עם תוכנת LabVIEW.
- הפעל את ה- APD.
- סגור את הקשתית עד שהאות יורד למטה כדי לקבל את גודל המותניים המינימלי, או סגור אותה לגודל מותניים גדול יותר.
- הקלט מספר ACFs של משך הזמן שנבחר (כלומר ריצה) כדי לשפר את יכולת השכפול הסטטיסטי, בדרך כלל 10 ריצות שנמשכות 20 שניות כל אחת עם תוכנת המתאם.
- כבה את ה-APD.
- השתמש בתוכנת Igor Pro כדי לבדוק ולהשליך את הריצות עם תנודות חזקות עקב אגרגטים מולקולריים. בצע שלב זה באופן ידני - הוא צריך להיות בלתי תלוי למשתמש לאחר שהמשתמשים הוכשרו.
- התאם את הממוצע של ה- ACFs שנשמרו עם מודל דיפוזיה תלת-ממדי.
- חלץ מהפרמטרים המתאימים את זמן
 הדיפוזיה הממוצע ושמור אותו בקובץ ".txt" (פורמט הקובץ מוכתב על ידי תוכנת Igor Pro).
הדיפוזיה הממוצע ושמור אותו בקובץ ".txt" (פורמט הקובץ מוכתב על ידי תוכנת Igor Pro). - בדוק את קצב הספירה למולקולה לשנייה (אינדיקטור לביצועים טובים) על ידי חלוקת העוצמה הממוצעת (המופקת מהמעקב הפלואורסצנטי) במספר המולקולות (המופקות מה-ACF).
הערה: ודא שערך זה גבוה ויציב מיום ליום עבור אותם פרמטרים של רכישה. - לדעת את מקדם הדיפוזיה של Rh6G בתמיסה מימית ב 37 °C (D) ו (ראה 7.13), לחשב את גודל המותניים הניסויי ω על פי:
 .
. - החל את ההליך עבור כל שינוי בגודל המותניים הנדרש כדי להתוות את חוק הדיפוזיה של FCS ולפני כל סדרה ניסיונית חדשה של רכישת נתוני svFCS.
8. רכישות נתונים של svFCS על תאים
- מדוד והתאם את הספק הקרן של 488 ננומטר בין 2 ל-4 μW. בהתאם לבהירות וליציבות הצילום של הגשושית הפלואורסצנטית שבה נעשה שימוש, התאימו את העוצמה הזו כדי לאפשר קצב ספירה גבוה לכל מולקולה (בדרך כלל כמה אלפי פוטונים למולקולה בשנייה), תוך שמירה על הפוטו-הלבנה נמוכה (כלומר, עקבות בעוצמה יציבה על תוכנת LabVIEW).
- שיווי משקל דגימות במשך 10 דקות ב 37 מעלות צלזיוס לפני תחילת המדידות.
- הגדר את מיקרוסקופ התאורה האפי-פלואורסצנטי עם תוכנת LabVIEW.
- בחר תא עם מיקום בדיקה פלואורסצנטית מתאים ועוצמת אות פלואורסצנטית (נמוכה).
הערה: ככל שהפלואורסצנציה נמוכה יותר, כך מדידות ה- FCS טובות יותר (ראה שלב 8.1). - הגדר את יציאת מיקרוסקופ התאורה/זיהוי של svFCS עם תוכנת LabVIEW.
- הפעל את ה- APD.
- בצע סריקת xy של התא שנבחר באמצעות תוכנת LabVIEW.
- בצע סריקת z ואתר את הנקודה הקונפוקלית בעוצמת הפלואורסצנציה המקסימלית על ידי בחירת קרום הפלזמה בחלק העליון והתחל ברכישת הנתונים. כדי למקסם את ההפרדה בין שתי הממברנות, רצוי לבצע את הסריקה באזור הגרעיני של התא.
- הקלט סדרה אחת של 20 ריצות שנמשכות 5 שניות, כל אחת עם תוכנת המתאם.
הערה: ודא שמשך הזמן של כל ריצה ארוך מספיק כדי להשיג ACFs עם רעש מופחת. רכישות ארוכות רגישות להלבנת תמונות או לשינויים משמעותיים בלתי צפויים (למשל, אגרגטים). התאימו את מספר הריצות, את משך הזמן שלהן ואת מספר הסדרות לדגימות, אך ודאו שהן נשארות קבועות בתוך אותו עיקר הניסויים לצורך שכפול. - כבה את ה-APD.
- בטל ריצות בלתי צפויות עם תוכנת Igor Pro.
- התאימו ל-ACF הממוצע מודל דיפוזיה דו-ממדי של 2 מינים. התאם מודל זה לסוג התנהגות הדיפוזיה של מולקולת המטרה.
- שמור את הפרמטרים המתאימים בקובץ הקודם (ראה שלב 7.13).
- בצע 10 עד 15 סדרות של הקלטות בלפחות 10 תאים שונים, ושחזר את שלבים 8.3 עד 8.13. בדוק שהקובץ היחיד שהתקבל מכיל את פרטי גודל המותניים ואת הפרמטרים המתאימים של 10-15 ההקלטות.
- כדי לקבוע חוק דיפוזיה יחיד, נתחו לפחות ארבעה גדלי מותניים הנעים בין 200 ל-400 ננומטר. טווח זה מוגדר על ידי הגבול האופטי של עקיפה, אך הוא תלוי אובייקטיבי - (מפתח צמצם מספרי) ולייזר (אורך גל).
הערה: מכיוון שכיול גודל המותניים אינו מוחלט ויש לו מידה מסוימת של אי-ודאות, תוכנת MATLAB ייעודית28 הרואה בחשבון את השגיאה x ו-y (כלומר ω2 ו-td) נבנתה כך שתתאים לחוק הדיפוזיה. - הפעל את תוכנת MATLAB 1 ובחר תיקיה המכילה את כל קבצי ".txt" המתאימים לארבעה ניסויים בגודל המותניים לפחות.
- העלילה <td> לעומת <ω2>, כלומר חוק הדיפוזיה. ניתן לחלץ שני פרמטרים עיקריים: יירוט ציר y (t0) ומקדם הדיפוזיה האפקטיבי (Deff, ביחס הפוך לשיפוע).
9. חוקי דיפוזיה של השוואת מצבים ניסיוניים שונים
הערה: במידת הצורך, שחזרו את סעיפים 7 ו-8 עבור תנאי ניסוי שונים. תוכנה ייעודית (תוכנת MATLAB 2) פותחה כדי לקבוע אם חוקי דיפוזיה אלה דומים או לא על פי ערכי t0 ו - Deff 28. הוא בוחן שתי השערות: שני הערכים שונים, או ששני הערכים אינם שונים בסף שנקבע מעל הסתברות לאזעקת שווא (PFA). ערך PFA שרירותי של 5% (T = 3.8) נחשב לגבול העליון של מובהקות בין שני פרמטרים (t0 או Deff), מה שמצביע על כך שיש סיכוי של 5% בלבד ששני הערכים זהים.
- צור קובץ ".xls" המכיל את ערכי חוק הדיפוזיה האופייניים של כל תנאי להשוואה (כלומר, קובץ המכיל את השגיאה t0, t0 , Deff ו- Deff עבור התנאים הלא מטופלים (NT) והטיפול (COase) כטבלה).
- הפעל את תוכנת MATLAB 2.
- בחר את הקובץ ".xls".
- נתחו את העלילה הדו-ממדית המקודדת בצבע שנוצרה, שבה יש לשרטט את המבחנים הסטטיסטיים t0 ו-Deff בצירי x ו-y, בהתאמה (איור 4). ככל ש-T גבוה יותר, כך ההפרש בין הערכים המשווים גדול יותר.
10. מדידות ריכוז כולסטרול
- טיפול בתאים וליזיס
- זרעו את תאי ה-Cos7 במשולש בצלחות של 6 בארות ב-4 × 105 תאים/באר ודגרו ב-2 מ"ל של DMEM שלם ב-37 מעלות צלזיוס עם 5% CO2 למשך הלילה כדי לאפשר לתאים להתחבר לצלחת.
- הסר את מדיום התרבית ושטף את התאים שלוש פעמים עם מלח בעל מאגר פוספט (PBS).
- הוסף 1 מ"ל של מאגר HBSS/HEPES המכיל (או לא, עבור בקרות) 1 U / mL של Coase, ודגירה למשך שעה אחת ב - 37 ° C עם 5% CO2.
- החלף את המדיום ב-1 מ"ל של HBSS/HEPES המכיל 0.1 U/mL של Coase, ודגירה למשך שעה אחת ב-37 °C עם 5% CO2.
- הסר את התמיסה וקצר את התאים.
- לשטוף את התאים שלוש פעמים עם PBS, וצנטריפוגה ב 400 × גרם במשך 5 דקות בטמפרטורת החדר.
- Lyse התאים עם חיץ בדיקת רדיו-אימונופרציפציה (25 mM HEPES, pH 7.4, 150 mM NaCl, 1% NP40, 10 mM, MgCl2, 1 mM ethylenediamine חומצה טטראצטית, 2% גליצרול, פרוטאז וקוקטייל מעכב פוספטאז) במשך 30 דקות על קרח.
- צנטריפוגה את lysates ב 10000 × גרם במשך 10 דקות ב 4 ° C ולאסוף את supernatant.
- לכמת את ריכוז החלבון הכולל עבור כל דגימה על ידי בדיקת החלבון של ברדפורד שעברה שינוי באמצעות תמיסת העבודה בהתאם להמלצות היצרן.
- מדידת ריכוז כולסטרול
- כדי לקבוע את רמת הכולסטרול בתאים הכוללת באופן אנזימטי, השתמש בערכה המתאימה (למשל, ערכת בדיקת הכולסטרול האדום של Amplex) בהתאם להמלצות היצרן.
- עבור כל תגובה, ערבבו את הדגימה המכילה 5 מיקרוגרם של חלבון עם ריאגנט אדום Amplex / חזרת פרוקסידאז / כולסטרול אוקסידאז / כולסטרול אסטראז תמיסת עבודה, ודגירה במשך 30 דקות ב 37 ° C בחושך.
- מדוד את הפלואורסצנציה באמצעות עירור של 520 ננומטר, וזהה את הפליטה ב-560–590 ננומטר באמצעות קורא מיקרו-לוחיות.
- הפחת את הרקע מהערך הסופי, וקבע את ריכוז הכולסטרול באמצעות עקומה סטנדרטית.
- חשב את תכולת הכולסטרול הסופית ב- ng של כולסטרול לכל מיקרוגרם של חלבון.
תוצאות
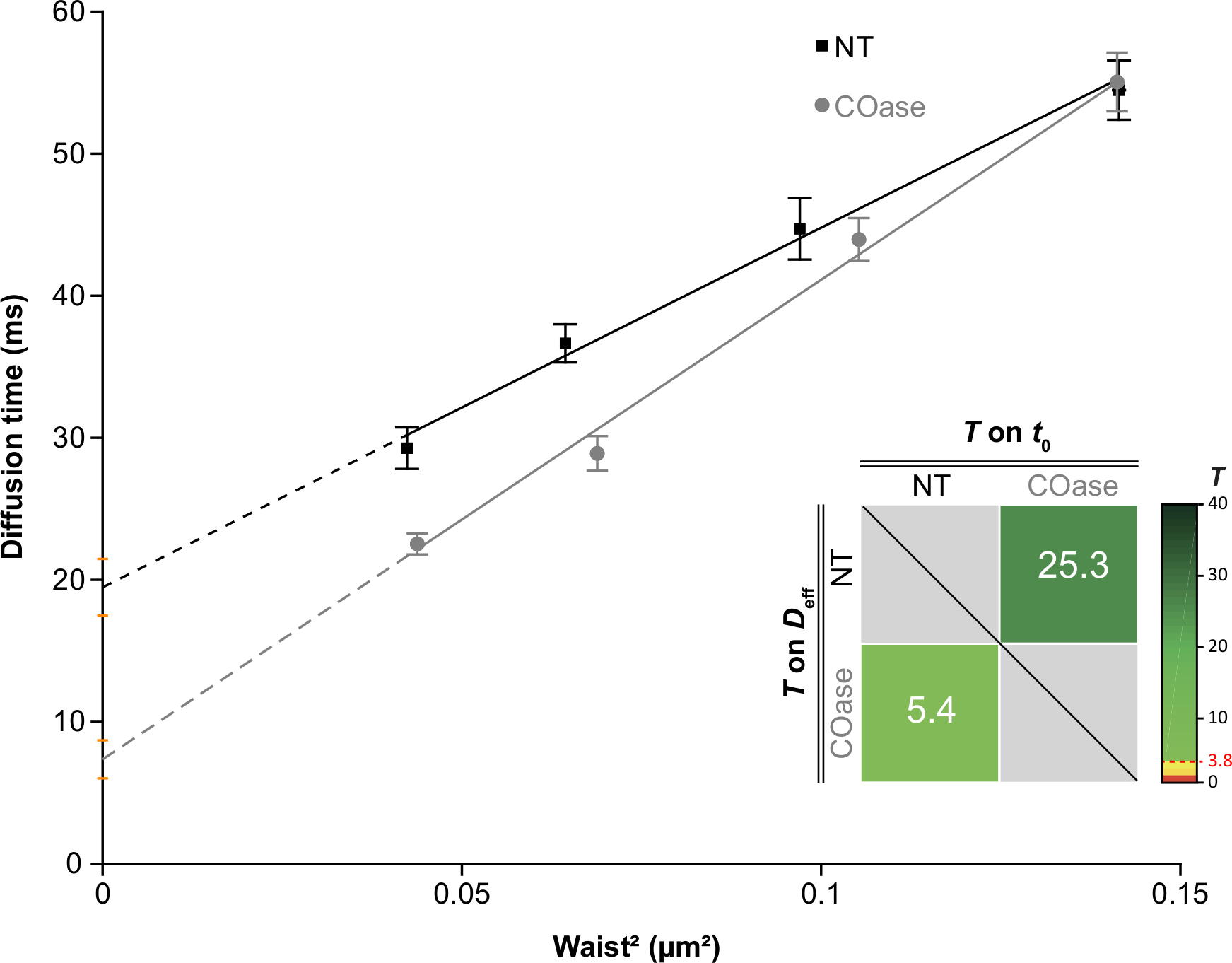
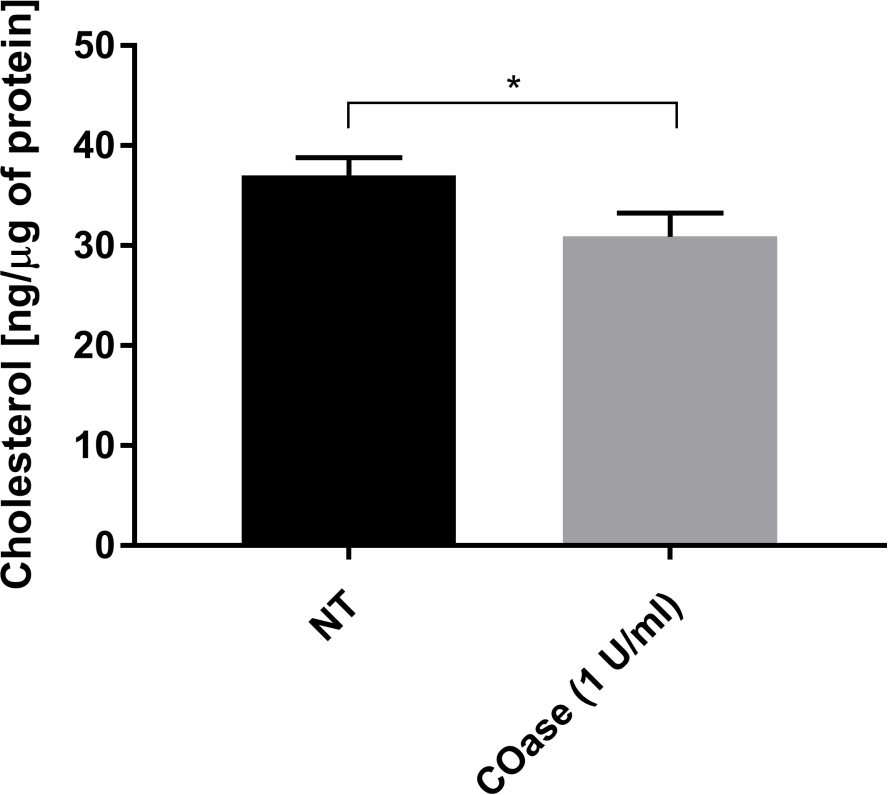
יצרנו DL עבור Thy1-GFP המבוטא בתאי Cos-7 (איור 4, ריבועים שחורים). לחוק הדיפוזיה יש ערך t0 חיובי (19.47 אלפיות השנייה ± 2 אלפיות השנייה), המציין כי Thy1-GFP מוגבל במבנים ננו-דומייניים של קרום הפלזמה. הטיפול בכולסטרול אוקסידאז בתאים המבטאים Thy1-GFP הביא להסטת ערך DL t0 ל-7.36 ± 1.34 אלפיות השנייה (איור 4, ריבועים אפורים). תצפית זו מאשרת כי אופי הכליאה של Thy1-GFP תלוי בתכולת הכולסטרול וקשור לננו-דומיינים של רפסודות שומנים. שני חוקי דיפוזיה אלה מוצגים כשונים על פי המבחן הסטטיסטי שתואר לעיל (ראה שלב 9.1.3) במונחים של ערכי t0 ו - Deff . בנוסף, הערכנו את הריכוז של הכולסטרול התאי הכולל בתאי Cos-7 שאינם מטופלים לעומת התאים שטופלו ב-COase. ירידה קטנה, אך משמעותית, בתכולת הכולסטרול הכללי נצפתה בטיפול ב-COase (איור 5). מכיוון שאנזים זה פועל רק במאגר הכולסטרול הנגיש בעלון החיצוני של קרום הפלזמה, אנו מניחים שהירידה הנצפית בכולסטרול קשורה רק לקרום הפלזמה וגורמת לערעור היציבות של ננו-דומיינים של רפסודות שומנים.

איור 1: מדומה של ספקטרוסקופיית מתאם פלואורסצנטי (FCS) חוקי דיפוזיה שנקבעו על ידי FCS בווריאציה נקודתית עבור צורות שונות של ארגון ממברנה. (לוחות עליונים) ייצוג סכמטי של ארגון ממברנה – (A) דיפוזיה חופשית, (B) מחסומי רשת, ו-(C) כליאה של מלכודות/תחומים – כאשר המסלול נמשך עבור מולקולה בודדת (אדום). עיגולים כחולים מציינים את ההצטלבות של הממברנה וקרן הלייזר של המותניים ω. (לוחות תחתונים) חוקי הדיפוזיה של FCS המיוצגים על ידי התוויית זמן הדיפוזיה td כפונקציה של הרדיוס בריבוע ω2. היטל חוק הדיפוזיה (קו מקווקו ירוק) מיירט את ציר הזמן ב- (A) המקור (t0 = 0) במקרה של דיפוזיה חופשית; (B) בציר שלילי (t0 < 0) כאשר יש מחסומי רשת, או (C) בציר חיובי (t0 > 0) כאשר יש מלכודות ותחומים (רפסודות שומנים). D הוא מקדם הדיפוזיה הלטרלי לתנועה בראונית; Deff, מקדם הדיפוזיה האפקטיבי; Dmicro, מקדם הדיפוזיה המיקרוסקופי בתוך מלכודות הרשת; Din, מקדם הדיפוזיה בתוך תחומים; Dout, מקדם הדיפוזיה מחוץ לתחומים; L, גודל הצד של תחום מרובע; ו-rD, הרדיוס של תחום מעגלי. נתון זה שונה מ-He ו-Marguet6. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 2: תצוגה סכמטית של בקרת החומרה של svFCS. המחשב שולט בכל המכשירים באמצעות פרוטוקולי תקשורת שונים: טורי (מיקרוסקופ, תריס חיצוני), USB (שלב פיאזואלקטרי XYZ, מתאם) ו- PCI (לוח רכישה). DAQ: לוח איסוף נתונים, APD: פוטודיודה מפולת שלגים, SPCM: מודול ספירת פוטונים בודדים, DO: פלט דיגיטלי. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
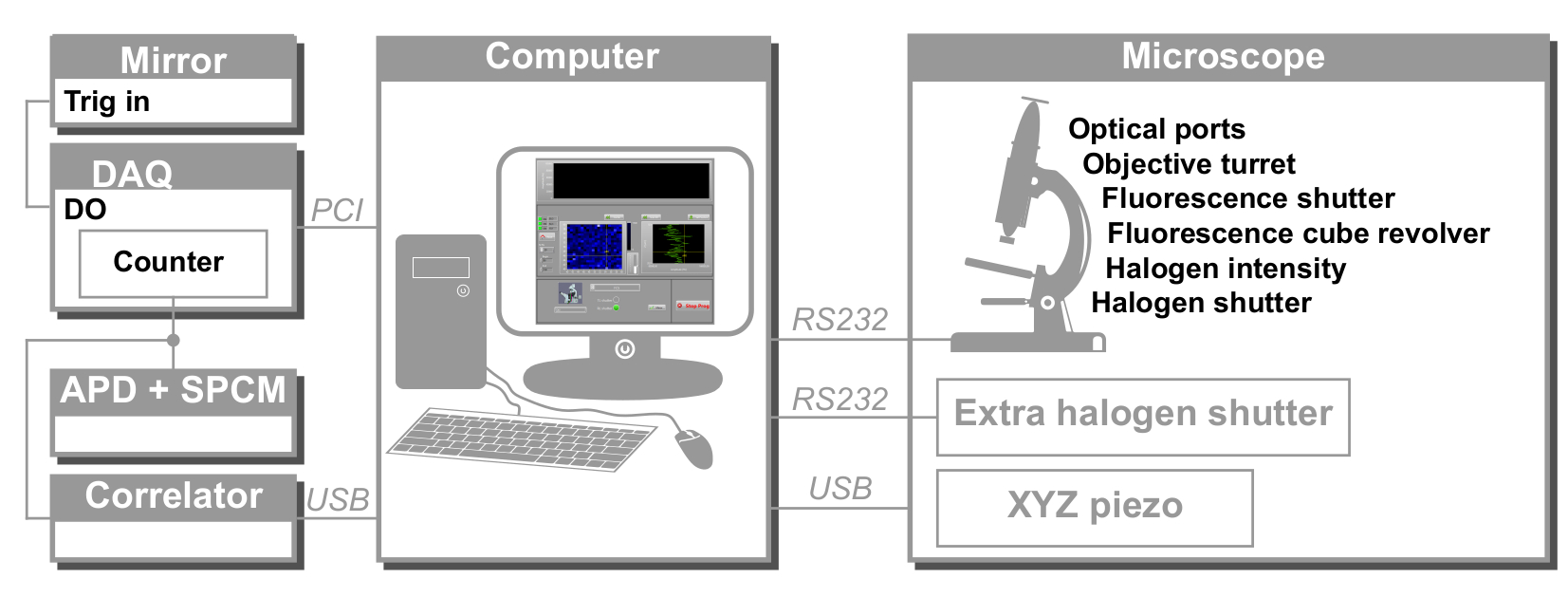{kind=link}

איור 3: תצוגה סכמטית של נתיבים אופטיים של עירור ופליטה של הגדרת svFCS. מערך ה-svFCS מכיל ארבעה מודולים: (1) התפוקה של לייזר 488 ננומטר עם סיבים מתנגשת, (2) שילוב של צלחת חצי גל ומפצל קרן מקטב קובע את העוצמה האופטית, (3) קרן הלייזר המתמקדת בדגימה לאחר שעברה דרך מיקרוסקופ ממונע ללא עדשת צינור, ו-(4) הפלואורסצנטיות מזוהה דרך נתיב זיהוי דמוי קונפוקל על פוטודיודה של מפולת שהוצמדה למודול ספירת פוטונים יחיד, המספק אות למתאם חומרה. הפשטות מעניקה למערכת את הרגישות, החוסן וקלות השימוש שלה (צוין בהרחבה בחומר משלים). אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
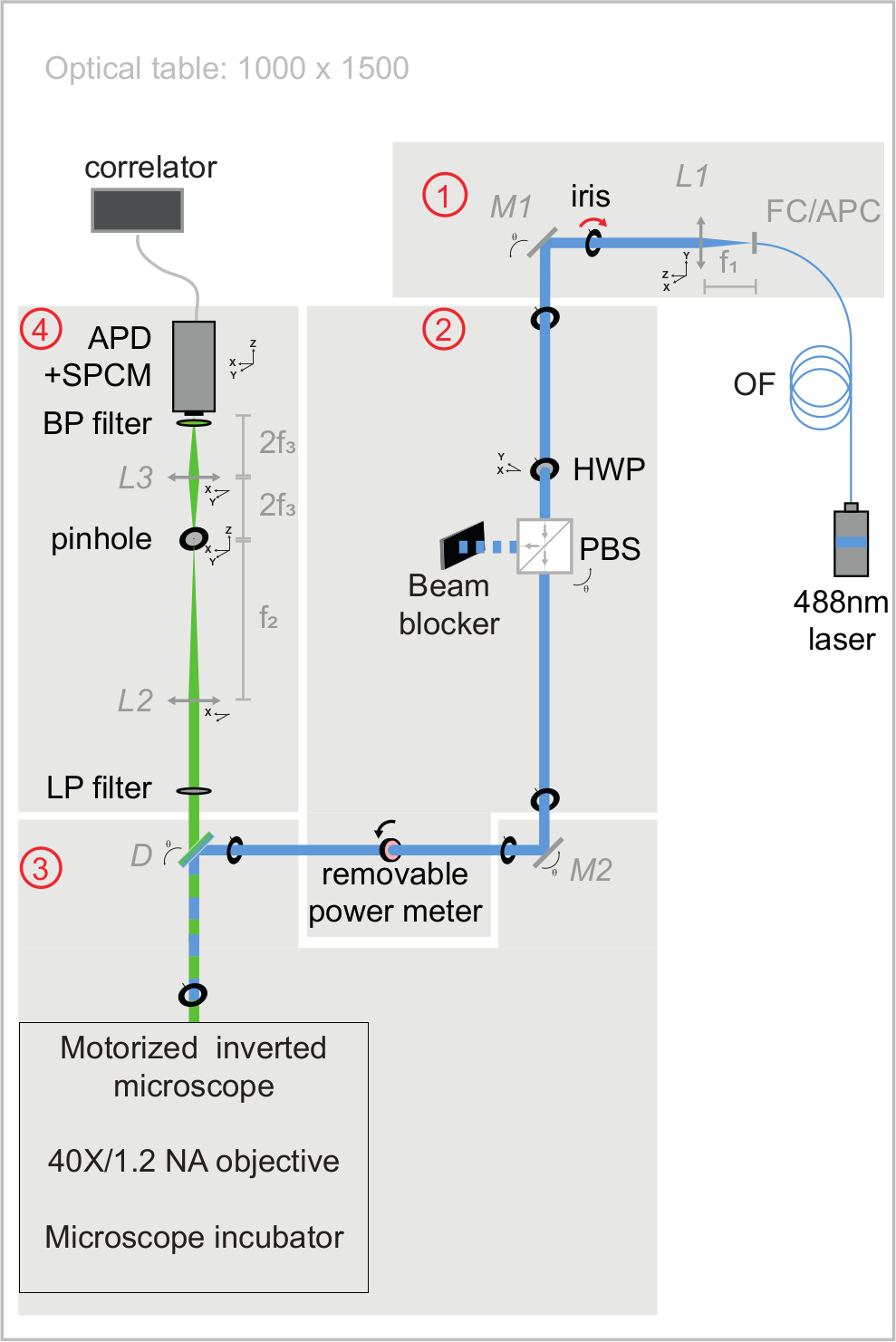{kind=link}

איור 4: חוקי הדיפוזיה של svFCS שנוצרו מניתוח דיפוזיה של Thy1-GFP שבאו לידי ביטוי ב-Cos-7. חוקי הדיפוזיה של svFCS של תאי Cos-7 ללא טיפול (NT, ריבועים שחורים) ולאחר טיפול באוקסידאז כולסטרול (COase, עיגולים אפורים). התוספת בגרף מייצגת בדיקה סטטיסטית של הבדל משמעותי בין שני חוקי הדיפוזיה של svFCS שהוצגו (על פי Mailfert et al.28). ערך הבדיקה (T) צריך להיות מעל הסף שנקבע ב- 3.8 כאשר שני חוקי הדיפוזיה שונים. ככל שהוא גבוה יותר, כך גדל ההבדל בין חוקי הדיפוזיה. הערך של T מקודד בצבע. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 5: השוואה של תכולת הכולסטרול הכוללת בתאי Cos-7. תאי Cos-7 לא טופלו (NT) או טופלו ב-1 U/mL של כולסטרול אוקסידאז (COase) במשך שעה אחת. הנתונים מייצגים דוגמה לניסוי אחד במשולש. מבחן t דו-זנב ולא מזווג שימש להערכת ההפרש הסטטיסטי (α=0.05). אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}
טבלת חומרים: רשימת האלמנטים האופטיים הנדרשים להגדרת svFCS.
חומר משלים: מסמך זה מתאר את בנייתה של הגדרת svFCS מאפס. אנא לחץ כאן כדי להוריד קובץ זה.
Discussion
כאן תיארנו את היישום של מודול svFCS על מיקרוסקופ פלואורסצנטי סטנדרטי, גישה ניסיונית רבת עוצמה לפענוח הדינמיקה של ארגון קרום הפלזמה בתאים חיים הודות לניתוח חוק הדיפוזיה של FCS. מבחינה מושגית, ה- svFCS מבוסס על עיקרון פשוט: מדידות קורלציה של פלואורסצנציה בתחום הזמן תוך שינוי גודל אזור ההארה23. אסטרטגיה זו סייעה להסיק מידע ננוסקופי ממדידות מיקרוסקופיות, המסייעות לפענח את היסודות הפיזיקוכימיים העיקריים התורמים לארגון קרום הפלזמה במצב יציב25 ובתהליכים פיזיולוגיים 30,31,32,33. בסך הכל, ניתוחי svFCS אלה מדגימים באופן חד משמעי את קיומם של ננו-דומיינים תלויי שומנים בסוגים שונים של תאים ואת ההשלכה הישירה שלהם בכוונון אירועי איתות שונים.
במסגרת זו, ישנם כמה היבטים אופטיים שיש לקחת בחשבון בעת בניית הגדרת svFCS כדי לייעל את תקציב הפוטונים ולמזער סטיות אופטיות. לפיכך, אנו ממליצים להשתמש במיקרוסקופ שממנו ניתן להסיר את עדשת הצינור כאשר מתבצעת מדידת svFCS. יתר על כן, קשתית בודדת ממלאת תפקיד מפתח במערך svFCS: היא משנה את גודל הקרן בפתח האחורי של המטרה, ובכך משנה ישירות את גודל המותניים האפקטיבי (כלומר, נפח העירור האפקטיבי). קוטר הקורה צריך להתאים לאישון האחורי האובייקטיבי כדי לקבל את גודל המותניים הקטן ביותר34. אפשרות זו, המסייעת לכוונן את גודל המותניים, מבטיחה אופטימיזציה של תקציב הפוטונים וקלה ליישום. לבסוף, מספר מינימלי של חלקים אופטיים משמשים לאורך נתיב האור; ככל שהמערכת פחות מורכבת, כך הפוטונים שאבדו פחות. כל האפשרויות הללו משפרות באופן משמעותי את החוסן של ניסויי svFCS.
לגבי הפרוטוקול עצמו, יש לקחת בחשבון כמה צעדים קריטיים. החשוב ביותר הוא יישור מתאים של הנתיבים האופטיים החיוני למדידות svFCS מוצלחות (פרוטוקול, סעיף 2). קל לבדוק זאת על ידי ניתוח האות הפלואורסצנטי מפתרון 2 ננומטר Rh6G, שאמור להיות ~ 200 קילוהרץ תחת תאורת לייזר של 300 μW. יש לפתוח את כל הקשתיות, ול-ACFs צריכה להיות משרעת חשובה (בדרך כלל G0~1.5-2.0). נקודה קריטית נוספת נוגעת לתאים ולהכנתם לניתוח svFCS (פרוטוקול, סעיפים 4-8). הצפיפות שלהם צריכה להיות מותאמת כך שתאים מבודדים לצפייה יהיו זמינים לניתוח. תאים שאינם דבקים צריכים להיות משותקים על כוסית כיסוי קאמרית באמצעות תמיסת פולי-L-ליזין. האות הפלואורסצנטי מתיוג תאים לא צריך להיות חזק מדי, או שהוא יגרום ל-ACFs שטוחים מאוד שקשה להתאים, ופרמטרים של התאמה עמוסים בשגיאה חשובה. בנוסף, תיוג לא-הומוגני ואגרגטים פלואורסצנטיים בתאים הופכים את מדידות ה-svFCS לקשות מאוד לפענוח. לבסוף, טיפול בכולסטרול אוקסידאז משפיע על כדאיות התאים, וניתוח svFCS לא יעלה על שעה אחת לאחר הטיפול. כמו כן, עדיף לרשום את התנודות הפלואורסצנטיות מממברנת הפלזמה העליונה מכיוון שהיא אינה מחוברת לתמיכה, ואין סיכון להפרעה לדיפוזיה של מולקולות בשל האינטראקציות הפיזיקליות עם התמיכה.
חלה התקדמות מספקת בטכניקת svFCS לשימוש בה בגישות שונות בשל מגוון האופנים להתאמת נפח הגילוי, מה שמאפשר לחקור תהליכים ביולוגיים שונים בתאים חיים. חלופה להתאמת גודל נפח העירור היא שימוש במרחיב קרן משתנה35. ניתן גם פשוט לווסת את גודל אזור ההארה על ידי הקלטת האות הפלואורסצנטי מהיירוט של קרום הפלזמה לאורך כיוון z36. ניתן לעשות זאת על מיקרוסקופ קונפוקלי סטנדרטי שעבורו פותחה מסגרת תיאורטית לגזירת חוק הדיפוזיה37,38.
למרות ששיטת svFCS מציעה רזולוציה מרחבית-טמפורלית, הנחוצה לאפיון הארגון הלטרלי האי-הומוגני של קרום הפלזמה, אופני הכליאה הגיאומטריים אינם בלעדיים זה לזה. סטייה של t0 בכיוון זה או אחר חושפת באופן בלעדי מצב דומיננטי של כליאה25. יתר על כן, מגבלה חשובה נוספת של שיטת svFCS הנוכחית נובעת ממגבלת עקיפה אופטית קלאסית (~ 200 ננומטר). זה ללא ספק גדול יותר מהתחומים המגבילים את המולקולות בתוך קרום הפלזמה של התא. לכן, ניתוח הכליאה מסיק מהערך t0 , אקסטרפולציה מחוק הדיפוזיה.
חיסרון זה התגבר על ידי יישום שיטות חלופיות. בתחילה, שימוש בסרטים מתכתיים שנקדחו עם ננו-צמצם הציע את האפשרות להאיר שטח ממברנה קטן מאוד (כלומר, מתחת לגבול עקיפה אופטית של מפתחי צמצם ננומטריים בודדים של רדי הנעים בין 75 ל-250 ננומטר)39. לפיכך דווח על משטר המעבר שנחזה מחוק הדיפוזיה התיאורטי לארגון תחום מבודד, והוא איפשר חידוד של הגודל האופייני של הטרוגניות הממברנה הננומטרית ואומדן כמותי של שטח הפנים שנכבש על ידי ננו-דומיינים תלויי ליפידים39. לחלופין, תאורה ננומטרית פותחה גם באמצעות מיקרוסקופיה אופטית לסריקת שדה קרוב40 או ננו-אנטנות אופטיות מישוריות41. לאחרונה, שילוב של דלדול פליטה מגורה (STED) ו-FCS סיפק כלי רב עוצמה ורגיש לתיעוד חוק הדיפוזיה ברזולוציה מרחבית גבוהה מאוד. STED-FCS זה נותן גישה למאפייני דיפוזיה מולקולרית בקנה מידה ננומטרי המתרחשים תוך פרק זמן קצר, ומאפשרים לחקור את הארגון הדינמי של בדיקות השומנים בקרום הפלזמה42,43. עם זאת, הדיכוי הלא שלם של פלואורסצנציה בתהליך STED מאתגר את הניתוח של עקומות המתאם האוטומטי ב- FCS.
פותח מודל מתאים חדש כדי להתגבר על קושי זה, ולשפר את הדיוק של זמני הדיפוזיה ומדידות מספרי המולקולות הממוצעות44. לבסוף, עבור דיפוזיה מולקולרית איטית בקרום הפלזמה, ניתן ליישם את עקרון svFCS על נתונים שנרשמו על ידי ספקטרוסקופיית מתאם תמונה45. לאחרונה, הוכח כי שילוב מיקרוסקופיה של כוח אטומי (AFM) עם הדמיה של השתקפות פנימית כוללת-FCS (ITIR-FCS) תורם לעידון אופי המנגנון המעכב דיפוזיה מולקולרית בקרום הפלזמה, במיוחד בסמוך לתצורת ממברנת סף החלחול בגלל צפיפות גבוהה של ננו-דומיינים46.
לסיכום, קביעת חוק הדיפוזיה על ידי svFCS סיפקה את הראיות הניסיוניות להסקת הטרוגניות מקומית שנוצרה על ידי שומנים קולקטיביים דינמיים וקשרים של חלבוני ממברנה. כפי שנאמר על ידי Wohland ועמיתיו46, "ניתוח חוק הדיפוזיה של FCS נותר כלי רב ערך להסקת תכונות מבניות וארגוניות מתחת למגבלת הרזולוציה ממידע דינמי". ובכל זאת, עלינו לפתח מודלים חדשים כדי לחדד את הפרשנות של חוק הדיפוזיה שאמורים לאפשר הבנה טובה יותר של הדינמיקה של האירועים המולקולריים המתרחשים בקרום הפלזמה.
Disclosures
למחברים אין מה לחשוף.
Acknowledgements
SB, SM ו- DM נתמכו על ידי מימון מוסדי מ- CNRS, Inserm ואוניברסיטת Aix-Marseille ומענקי תוכנית מסוכנות המחקר הלאומית הצרפתית (ANR-17-CE15-0032-01 ו- ANR-18-CE15-0021-02) ומענקי התוכנית הצרפתיים "Investissement d'Avenir" (ANR-10-INBS-04 France-BioImaging, ANR-11-LABX-054 labex INFORM). KW מכיר ב-"BioTechNan", תוכנית ללימודי דוקטורט סביבתיים בין-תחומיים בתחום הביוטכנולוגיה והננוטכנולוגיה. EB מכיר בתמיכה הכספית של מרכז המדע הלאומי של פולין (NCN) במסגרת פרויקט מס '2016/21/D/NZ1/00285, כמו גם ממשלת צרפת ושגרירות צרפת בפולין. MŁ מכיר בתמיכה הכספית של משרד הפיתוח הפולני (CBR POIR.02.01.00-00-0159/15-00/19) והמרכז הלאומי למחקר ופיתוח (Innochem POIR.01.02.00-00-0064/17). TT מכיר בתמיכה כספית ממרכז המדע הלאומי של פולין (NCN) במסגרת פרויקט מס '2016/21/B/NZ3/00343 ומהמרכז הביוטכנולוגיה של ורוצלב (KNOW).
Materials
| Name | Company | Catalog Number | Comments |
| Aligment tool | Spanner Wrench for SM1-Threaded Retaining Rings | Thorlabs | SPW602 |
| Avalanche Photodiode and Single Photon Counting Module (SPCM) | Single-Photon Counting Module, Avalanche Photodiode | Excelitas | SPCM-AQRH-15 |
| BNC 50 Ω plug to 50 Ω plug lead 2 m | RS Components | 742-4315 | |
| Coaxial cable 415 Cinch Connectors, RG-316, 50 Ω With connector, 1.22 m, RoHS2 | RS Components | 885-8172 | |
| Tee 50Ω RF Adapter BNC Plug to BNC Socket 0 → 1GHz | RS Components | 546-4948 | |
| Brennenstuhl 2.5 m, 8 Socket Type E – French Extension Lead, 230 V | RS Components | 768-5500 | |
| Mascot, 6W Plug In Power Supply 5V dc, 1.2A, 1 Output Switched Mode Power Supply, Type C | RS Components | 452-8394 | |
| Crystek CCSMACL-MC-24 Reference Oscillator Power Cable RF Adapter | RS Components | 792-4079 | |
| Fluorescence filtering | 535/70 ET Bandpass, AOI 0° Chroma Diameter 25 mm | AHF filter | F47-539 |
| Laser Beamsplitter zt488 RDC, AOI 45° Chroma 25.5 x 36 x 1 mm | AHF filter | F43-088 | |
| 496/LP BrightLine HC Longpass Filter, AOI 0° Chroma Diameter 25 | AHF filter | F37-496 | |
| Hardware correlator | 80 MHz Digital Correlator | Correlator.com | Flex02-12D |
| Laser | LASER LASOS LDM-XT fiber coupled, 488 nm, 65 mW | Lasos | BLD-XT 488100 |
| Laser safety | High-Performance Black Masking Tape, 1" x 180' (25 mm x 55 m) Roll | Thorlabs | T743-2.0 |
| Lens Tissues, 25 Sheets per Booklet, 5 Booklets | Thorlabs | MC-5 | |
| Laser Safety Glasses, Light Orange Lenses, 48% Visible Light Transmission | Thorlabs | LG3B | |
| Microscope | Zeiss Axiovert 200M Motorized Inverted Fluorescence Microscope Fine and coarse focusing, reflector turret rotation, objective nosepiece rotation, switching camera ports, and internal light shutters | Carl Zeiss | |
| C-Apochromat 40x/1,2 W Korr.selected for FCS (D=0.14-0.19 mm) (WD=0.28 mm at D=0.17 mm), UV-VIS-IR | Carl Zeiss | 421767-9971-711 | |
| Adapter W0.8 / M27x0.75 H "5" | Carl Zeiss | 000000-1698-345 | |
| Middle ring W0.8 - W0.8 H "5" | Carl Zeiss | 000000-1698-347 | |
| Optical path | D25.4mm Mirror, Protected Silver | Thorlabs | PF10-03-P01 |
| D25.4mm, F=60.0.mm, Visible Achromat | Thorlabs | AC254-060-A | |
| D25.4mm, F=35.0.mm, Visible Achromat | Thorlabs | AC254-035-A | |
| 25 µm mounted pinhole | Thorlabs | P25S I | |
| 25.4mm Mounted Zero, Order 1/2 Waveplate 488 nm | Thorlabs | WPH10M-488 (HWP) | |
| 20mm Polarizing Beamsplitter Cube 420-680 nm | Thorlabs | PBS201 | |
| Rotation Stage 56 mm x 26 mm Threaded ID | Thorlabs | RSP1/M | |
| 52 mm x 52 mm Kinematic Platform Mount | Thorlabs | KM100B/M | |
| Adjustable Prism Clamp | Thorlabs | PM3/M | |
| Beam block - active area 19 mm x 38 mm | Thorlabs | LB1/M | |
| Iris Diaphragm 1 mm to 25 mm Aperture | Thorlabs | ID25/M | |
| Left-Handed Kinematic Cylindrical Lens Mount | Thorlabs | KM100CL | |
| 1" Optic Holder, M4 Tap | Thorlabs | MFF101/M | |
| 1" Stackable Lens Tube | Thorlabs | SM1L03 | |
| Stackable Lens Mount for 1" optic-usable depth ½ | Thorlabs | SM1L05 | |
| Stackable Lens Mount For 1"Optic-usable Depth 2" | Thorlabs | SM1L20 | |
| Small Optical Rails 600mm, metric | Thorlabs | RLA600/M | |
| Small Optical Rails 75mm, metric | Thorlabs | RLA075/M | |
| Small Optical Rails 150mm, metric | Thorlabs | RLA150/M | |
| Rail Carrier, Counterbored Hole 1"x 1" | Thorlabs | RC1 | |
| Rail Carrier, Perpendicular Dovetail | Thorlabs | RC3 | |
| High Precision Translating Lens Mount for 1 inch | Thorlabs | LM1XY/M | |
| ½ " (12mm) Dovetail Translation Stage | Thorlabs | DT12/M | |
| Rail Clamps | Thorlabs | CL6 | |
| Metric XYZ Translation Stage (Includes PT102) | Thorlabs | PT3/M | |
| Black Rubberized Fabric | Thorlabs | BK5 | |
| Ball Driver kit/ 6 tools | Thorlabs | BD-KIT/M | |
| Adapter with External M6 x 1.0 Threads and External M4 x 0.7 Threads | Thorlabs | AP6M4M | |
| Mounting Base, 25 mm x 58 mm x 10 mm, 5 Pack | Thorlabs | BA1S/M-P5 | |
| Lens Mount for 25.4mm optic | Thorlabs | LMR1/M | |
| SM1 FC/APC Adapter | Thorlabs | SM1FCA | |
| Kinematic Mirror Mount For 1 inch Optics | Thorlabs | KM100 | |
| Silicon Power Head, 400-1100nm, 50mW | Thorlabs | S120C | |
| 12.7 mm Post Holders, Spring-Loaded Hex-Locking Thumbscrew, L=50 mm, 5 Pack | Thorlabs | PH50/M-P5 | |
| Post Holder with Spring-Loaded Hex-Locking Thumbscrew, L=20 mm | Thorlabs | PH20/M | |
| 12.7 mm x 50 mm Stainless Steel Optical Post, M4 Stud, M6-Tapped Hole, 5 Pack | Thorlabs | TR50/M-P5 | |
| 12.7 mm x 75 mm Stainless Steel Optical Post, M4 Stud, M6-Tapped Hole, 5 Pack | Thorlabs | TR75/M-P5 | |
| USB Power and Energy Meter Interface | Thorlabs | PM100USB | |
| 12.7 mm x 30 mm Stainless Steel Optical Post, M4 Stud, M6-Tapped Hole | Thorlabs | TR30/M | |
| 12.7 mm x 20 mm Stainless Steel Optical Post, M4 Stud, M6-Tapped Hole | Thorlabs | TR20/M | |
| 750 mm long Structural Rail (detection box) | Thorlabs | XE25L750/M | |
| 350 mm long Structural Rail (detection box) | Thorlabs | XE25L350/M | |
| Quick Corner Cube for 25 mm Rails | Thorlabs | XE25W3 | |
| Right-Angle Bracket for 25 mm Rails | Thorlabs | XE25A90 | |
| Black posterboard 20" x 30" (508 mm x 762 mm), 1/16" (1.6 mm) Thick, 5 Sheets | Thorlabs | TB5 | |
| Hinge for 25 mm Rail Enclosures | Thorlabs | XE25H | |
| Lid Stop for 25 mm Rail Enclosures | Thorlabs | XE25LS | |
| M4 Cap Screw Kit | Thorlabs | HW-KIT1/M | |
| M6 Cap Screw Kit and Hardware kit | Thorlabs | HW-KIT2/M | |
| Table Clamp, L-Shape, 5 Pack | Thorlabs | CL5-P5 | |
| SM1 Ring-Actuated Iris Diaphragm (Ø0.8 - Ø12 mm) | Thorlabs | SM1D12D | |
| Ø1" SM1-Mounted Frosted Glass Alignment Disk w/Ø1 mm Hole | Thorlabs | DG10-1500-H1-MD | |
| Honeycomb Optical Table Top, Standa | Standa | 1HB10-15-12 | |
| Optical Table support, Standa | Standa | 1TS05-12-06-AR | |
| Sample nano-positionning | Precision XYZ Nanopositioning | Physik Instrumente | PI P527-3.CD |
| Digital Multi-Channel Piezo Co, 3 Channels, -30 to 130 V Sub- D Connector(s), Capacitive Sensors, | Physik Instrumente | PI E727-3.CD | |
| Temperature chamber | Zeiss 200M Inverted Microscope Incubator System MATT BLK | Digital Pixel | |
| Dual Channel Microprocessor Temperature Controller | Digital Pixel | DP_MTC_2000_DUO | |
| Two Vibration Free Heater Modules | Digital Pixel | DP_150_VF | |
| PT100 Temperature Sensor | Digital Pixel | DP_P100_TS | |
| Biological Reagents and Materials | |||
| Cell culture and transfection | Cos7 cells | ATCC® | CRL-1651™ |
| 8- well Lab-Tek chambers | Thermo Fisher Scientific | 155411PK | |
| Dulbecco's Modified Eagle Medium (DMEM) | Thermo Fisher Scientific | 11965092 | |
| Fetal bovine serum | Thermo Fisher Scientific | 16000044 | |
| L-glutamine | Thermo Fisher Scientific | 25030081 | |
| PBS buffer | Thermo Fisher Scientific | 14190144 | |
| PenStrep | Thermo Fisher Scientific | 15140122 | |
| PolyJet Transfection Reagent | SignaGen Laboratories | SL100688 | |
| Cholesterol content measurement | Amplex Red Cholesterol Assay Kit | Thermo Fisher Scientific | A12216 |
| Protease Inhibitor Cocktail | Thermo Fisher Scientific | 87786 | |
| Phosphatase Inhibitor Cocktail | Thermo Fisher Scientific | 78420 | |
| ROTI Nanoquant Working Solution | Roth | K880 | |
| GloMax Discover Microplate Reader | Promega | GM3000 | |
| svFCS measurements | HBSS buffer | Thermo Fisher Scientific | 14025092 |
| Hepes buffer | Thermo Fisher Scientific | 15630080 | |
| Cholesterol oxidase | Sigma-Aldrich | C8868 | |
| Rhodamine 6G | Sigma-Aldrich | 83697-1G |
References
- Engelman, D. M. Membranes are more mosaic than fluid. Nature. 438 (7068), 578-580 (2005).
- Newton, A. C. Regulation of the ABC kinases by phosphorylation: protein kinase C as a paradigm. The Biochemical Journal. 370, Pt 2 361-371 (2003).
- Marguet, D., Lenne, P. F., Rigneault, H., He, H. T. Dynamics in the plasma membrane: how to combine fluidity and order. The EMBO Journal. 25 (15), 3446-3457 (2006).
- Edidin, M. The state of lipid rafts: From model membranes to cells. Annual Review of Biophysics and Biomolecular Structure. , (2003).
- Lingwood, D., Simons, K. Lipid rafts as a membrane-organizing principle. Science. , (2010).
- He, H. T., Marguet, D. Detecting nanodomains in living cell membrane by fluorescence correlation spectroscopy. Annual Reviews of Physical Chemistry. 62, 417-436 (2011).
- Rossy, J., Ma, Y., Gaus, K. The organisation of the cell membrane: Do proteins rule lipids. Current Opinion in Chemical Biology. 20 (1), 54-59 (2014).
- Nicolson, G. L. The Fluid - Mosaic Model of Membrane Structure: Still relevant to understanding the structure, function and dynamics of biological membranes after more than 40 years. Biochimica et Biophysica Acta - Biomembranes. 1838 (6), 1451-1466 (2014).
- Fujiwara, T., Ritchie, K., Murakoshi, H., Jacobson, K., Kusumi, A. Phospholipids undergo hop diffusion in compartmentalized cell membrane. The Journal of Cell Biology. 157 (6), 1071-1081 (2002).
- Kusumi, A., et al. Paradigm shift of the plasma membrane concept from the two-dimensional continuum fluid to the partitioned fluid: high-speed single-molecule tracking of membrane molecules. Annual Review of Biophysics and Biomolecular Structure. 34, 351-378 (2005).
- Destainville, N., Salomé, L. Quantification and correction of systematic errors due to detector time-averaging in single-molecule tracking experiments. Biophysical Journal. 90 (2), 17-19 (2006).
- Wieser, S., Moertelmaier, M., Fuertbauer, E., Stockinger, H., Schütz, G. J. Un)confined diffusion of CD59 in the plasma membrane determined by high-resolution single molecule microscopy. Biophysical Journal. 92 (10), 3719-3728 (2007).
- Axelrod, D., Koppel, D. E., Schlessinger, J., Elson, E., Webb, W. W. Mobility measurement by analysis of fluorescence photobleaching recovery kinetics. Biophysical Journal. , (1976).
- Petrášek, Z., Schwille, P. Precise measurement of diffusion coefficients using scanning fluorescence correlation spectroscopy. Biophysical Journal. , (2008).
- Elson, E. L. 40 Years of FCS: How it all began. Methods in Enzymology. , (2013).
- Elson, E. L., Magde, D. Fluorescence correlation spectroscopy. I. Conceptual basis and theory. Biopolymers. , (1974).
- Magde, D., Elson, E. L., Webb, W. W. Fluorescence correlation spectroscopy. II. An experimental realization. Biopolymers. , (1974).
- Schwille, P., Haupts, U., Maiti, S., Webb, W. W. Molecular dynamics in living cells observed by fluorescence correlation spectroscopy with one- and two-photon excitation. Biophysical Journal. , (1999).
- Bouchaud, J. P., Georges, A. Anomalous diffusion in disordered media: Statistical mechanisms, models and physical applications. Physics Reports. , (1990).
- Yechiel, E., Edidin, M. Micrometer-scale domains in fibroblast plasma membranes. Journal of Cell Biology. 105 (2), 755-760 (1987).
- Salomé, L., Cazeils, J. L., Lopez, A., Tocanne, J. F. Characterization of membrane domains by FRAP experiments at variable observation areas. European Biophysics Journal. 27 (4), 391-402 (1998).
- Niv, H., Gutman, O., Kloog, Y., Henis, Y. I. Activated K-Ras and H-Ras display different interactions with saturable nonraft sites at the surface of live cells. The Journal of Cell Biology. 157 (5), 865-872 (2002).
- Wawrezinieck, L., Rigneault, H., Marguet, D., Lenne, P. F. Fluorescence correlation spectroscopy diffusion laws to probe the submicron cell membrane organization. Biophysical Journal. 89 (6), 4029-4042 (2005).
- Saxton, M. J. Fluorescence corralation spectroscopy. Biophysical Journal. , (2005).
- Lenne, P. F., et al. Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork. EMBO Journal. 25 (14), 3245-3256 (2006).
- Ruprecht, V., et al. Cortical contractility triggers a stochastic switch to fast amoeboid cell motility. Cell. , (2015).
- Billaudeau, C., et al. Probing the plasma membrane organization in living cells by spot variation fluorescence correlation spectroscopy. Methods in Enzymology. 519, 277-302 (2013).
- Mailfert, S., Hamon, Y., Bertaux, N., He, H. T., Marguet, D. A user's guide for characterizing plasma membrane subdomains in living cells by spot variation fluorescence correlation spectroscopy. Methods in Cell Biology. 139, 1-22 (2017).
- Rege, T. A., Hagood, J. S. Thy-1, a versatile modulator of signaling affecting cellular adhesion, proliferation, survival, and cytokine/growth factor responses. Biochimica et Biophysica Acta - Molecular Cell Research. , (2006).
- Cahuzac, N., et al. Fas ligand is localized to membrane rafts, where it displays increased cell death-inducing activity. Blood. , (2006).
- Guia, S., et al. Confinement of activating receptors at the plasma membrane controls natural killer cell tolerance. Science Signaling. 4 (167), 21(2011).
- Blouin, C. M., et al. Glycosylation-dependent IFN-γR partitioning in lipid and actin nanodomains is critical for JAK activation. Cell. 166 (4), 920-934 (2016).
- Chouaki-Benmansour, N., et al. Phosphoinositides regulate the TCR/CD3 complex membrane dynamics and activation. Scientific Reports. , (2018).
- Wawrezinieck, L., Lenne, P. F., Marguet, D., Rigneault, H. Fluorescence correlation spectroscopy to determine diffusion laws: application to live cell membranes. Biophotonics Micro- and Nano-Imaging. , (2004).
- Masuda, A., Ushida, K., Okamoto, T. New fluorescence correlation spectroscopy enabling direct observation of spatiotemporal dependence of diffusion constants as an evidence of anomalous transport in extracellular matrices. Biophysical Journal. , (2005).
- Humpolíčková, J., et al. Probing diffusion laws within cellular membranes by Z-scan fluorescence correlation spectroscopy. Biophysical Journal. , (2006).
- Benda, A., et al. How to determine diffusion coefficients in planar phospholipid systems by confocal fluorescence correlation spectroscopy. Langmuir. , (2003).
- Ganguly, S., Chattopadhyay, A. Cholesterol depletion mimics the effect of cytoskeletal destabilization on membrane dynamics of the serotonin1A receptor: A zFCS study. Biophysical Journal. , (2010).
- Wenger, J., et al. Diffusion analysis within single nanometric apertures reveals the ultrafine cell membrane organization. Biophysical Journal. 92 (3), 913-919 (2007).
- Manzo, C., Van Zanten, T. S., Garcia-Parajo, M. F. Nanoscale fluorescence correlation spectroscopy on intact living cell membranes with NSOM probes. Biophysical Journal. , (2011).
- Regmi, R., et al. Planar optical nanoantennas resolve cholesterol-dependent nanoscale heterogeneities in the plasma membrane of living cells. Nano Letters. , (2017).
- Mueller, V., et al. FCS in STED microscopy: Studying the nanoscale of lipid membrane dynamics. Methods in Enzymology. , (2013).
- Sezgin, E., et al. Measuring nanoscale diffusion dynamics in cellular membranes with super-resolution STED-FCS. Nature Protocols. , (2019).
- Wang, R., et al. A straightforward STED-background corrected fitting model for unbiased STED-FCS analyses. Methods. , (2018).
- Veerapathiran, S., Wohland, T. The imaging FCS diffusion law in the presence of multiple diffusive modes. Methods. , (2018).
- Gupta, A., Phang, I. Y., Wohland, T. To hop or not to hop: exceptions in the FCS diffusion law. Biophysical Journal. , (2020).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved