
Method Article
היבטים מעשיים של הכנת מדגם והגדרת 1H R1ρ הרפיה ניסויים של RNA
In This Article
Summary
אנו מציגים פרוטוקול למדידת דינמיקה מיקרו-אלפית השנייה על ספקטרוסקופיית RNA עם תווית N של 13C/15עם תווית N עם ספקטרוסקופיית פיזור הרפיה של 1H R1ρ. המוקד של פרוטוקול זה טמון הכנה מדגם טוהר גבוהה והגדרת ניסויי NMR.
Abstract
RNA הוא ביומולקולה גמישה מאוד, שבה שינויים במבנים ממלאים תפקידים מכריעים בפונקציות שמולקולות RNA מבצעות כשליחים ומאפננים תאיים. בעוד מצבים דינמיים אלה נשארים מוסתרים לרוב השיטות המבניות, ספקטרוסקופיית פיזור הרפיה (RD) R 1ρ מאפשרת לחקור דינמיקה קונפורמציה במשטר מיקרו-אלפיות השנייה ברזולוציה אטומית. השימוש ב- 1H כגרעין הנצפה מרחיב עוד יותר את משטר הזמן המכוסה ונותן גישה ישירה לקשרי מימן ולזוג בסיסים.
הצעדים המאתגרים במחקר כזה הם טוהר גבוה והכנת מדגם בתפוקה גבוהה, פוטנציאל 13C- ו 15N-labeled, כמו גם הגדרת ניסויים והתאמה של נתונים כדי לחלץ אוכלוסייה, שער חליפין, ומבנה משני של המצב הבלתי נראה בעבר. פרוטוקול זה מספק צעדים מעשיים קריטיים בהכנת מדגם כדי להבטיח הכנת דגימת RNA מתאימה והגדרת ניסויי 1 H R1ρ עם דגימותRNA איזוטופיות ולא מתויגות.
Introduction
RNAs לבצע שפע שלרגולציה 1, קטליטי2, ופונקציות מבניות3 בתא, שרבים מהם מתואמים למבנה מולקולרי גמיש ושינויים מורכבים של מבנים אלה4,5,6,7. מדינות מאוכלסות נמוכות נשארות בלתי נראות לרוב השיטות לקביעת מבנים או אינן מאפשרות את חקר מצבים נסתרים אלה ברזולוציה אטומית גבוהה. ספקטרוסקופיית תהודה מגנטית גרעינית (NMR) של מצב פתרון משלבת את שני ההיבטים על ידי מתן גישה לגרעיני אטומים בודדים, כמו גם הצעת ארגז כלים גדול של ניסויים המתמקדים בדינמיקה באמצעות משטרי כל הזמנים8. ניסויי RD NMR מספקים גישה לחילופי קונפורמציה בציר הזמן הבינוני, שבו צפויים שינויים בדפוסי זיווג הבסיס ובארגון מחדש מבני מקומי5,9,10,11,12,13,14. ניסויי מו"פ מבוצעים כמדידות R2 ארוכות בצורת רכבת פולס קאר-פרסל-מייבום-גיל15 או כמדידות הרפיה במסגרת המסתובבת, הנקראת ניסויי R1ρ RD16.
למרות ששניהם יכולים לשמש לחילוץ אוכלוסייה ושער חליפין והפרש תזוזה כימי למצב המשני, ניסויי R1ρ RD גם נותנים את הסימן להבדל המשמרת הכימית של המצב הנרגש. זה מאפשר מסקנה על מבנה משני, אשר מתאם מאוד לשינוי כימי במבני RNA17. השינוי הכימי הוא אינדיקטור טוב של helicity במקרה של פרוטונים ארומטיים ופחמנים על nucleobases, של שותפי זיווג בסיס עבור פרוטונים imino, ושל דסקי סוכר על C4 ' ו C1 ' אטומים18,19. יש לציין כי לאחרונה ניסוי העברת רוויית חילופי כימיקלים (CEST) באמצעות כוח נעילת ספין גבוהה יותר (SL), ובכך הסטת הישימות של ניסוי CEST כדי להחליף לוחות זמנים מהירים יותר, פורסם כחלופה לניסוי R1ρ RD למערכות עם מצב נרגש אחד.
למרות 13C ו 15N איזוטופים שימשו לעתים קרובות כדי לגשת חילופי מבנים, עבודה האחרונה ממעבדה זו השתמש פרוטונים ארומטיים imino כמו בדיקות עבור חילופי קונפורמציה9,10. השימוש ב- 1H כגרעין הנצפה מביא מספר יתרונות, לדוגמה, גישה להחלפה בלוחות זמנים מהירים ואיטיים יותר, רגישות גבוהה יותר זמני מדידה קצרים יותר. זה מתאפשר עוד יותר על ידי גישת ניסוי פרוטון ממוטב SELective (SELOPE), מתן גישה פרוטונים ארומטיים באמצעות decrowding של ספקטרום חד ממדי (1D) באמצעות צימודים סקלריים הומו-גרעיניים, במקום העברת מגנטיזציה הטרונוקלרית, וביטול הצורך בתוויות איזוטופ20. פרוטוקול זה מטפל במדידה בניסויי RD של 1H R1ρ של דגימות אחידות של 13C/15N עם תווית N ולא מתויגות. לכן, מאמר זה מציג שיטת הכנה לדוגמה שנמצאה כרב-תכליתית ביותר לצרכי הכנת מדגם שונים21 ודן בחלופות בחלק האחרון של מאמר זה (איור 1).
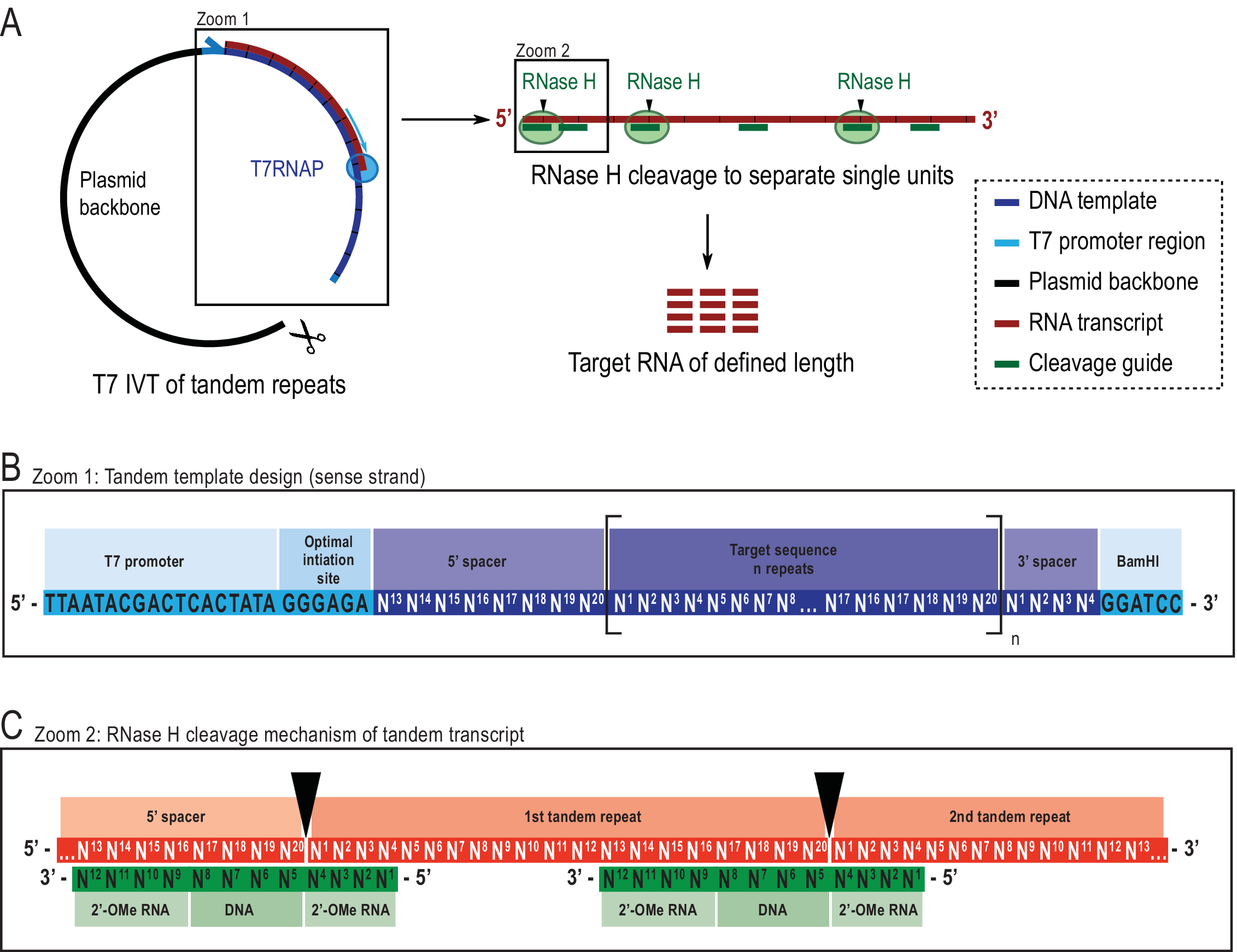
בשלב זה, הקורא צריך לציין כי טכניקות הכנה מדגם אחרות מקובלות עבור 1H R1ρ RD ניסויים, וכי שיטות אחרות של ניתוח מבני ופונקציונלי יכול להתבצע עם הדגימות מסונתז עם הטכניקה המוצגת. 1 ניסויי H R1ρ RD דורשים ריכוזי RNA גבוהים (באופן אידיאלי >1 mM) כמו גם הומוגניות גבוהה, הן באורך RNA והן בקונפורמציה מבנית כדי להבטיח אפיון אמין של דינמיקה מולקולרית. תמלול במבחנה (IVT) היא שיטת הבחירה עבור חוקרים רבים לייצר 13 C/15 דגימותRNA עם תווית N בשל הזמינות של טריפוספטים נוקלאוזידים מסומנים (NTPs) והתאגדות facile בתגובה אנזימטית22. עם זאת, T7 RNA פולימראז בשימוש נרחב (T7RNAP)23,24,25 סובל הומוגניות נמוכה 5 'במקרה של רצפי חניכה מסוימים26,27 ולעתים קרובות גם 3 ' הומוגניות במהלך נגר תמלול28. טיהור מינים RNA היעד הופך יקר יותר ומייגע בשל הצורך בכמויות גדולות של ~ 200 נמול. השיטה המשמשת כאן הוצגה בעבר שבה נדונו היתרונותבכללותו 21. בקצרה, הוא פותר בעיות המתוארות על ידי תעתיק דו-מושבי גדול יותר, שאז הוא נבקע במיוחד על ידי Escherichia coli RNase H, בהדרכת אוליגונוקלאוטיד כימרי29,30 (ראה איור 2 לפרטים).
שילוב של רצף מרווחים בקצוות 5' ו-3' של תמליל הטנדם מאפשר שימוש ברצף ייזום בתפוקה גבוהה והסרת אובר-התייבשות סופניות קרוב לאתר הליניאריזציה של תבנית הפלסמיד, בהתאמה (איור 2B). השיטה הוצגה כדי לשפר את התשואות באופן משמעותי, תוך הפחתת עלות ועבודה, עם אזהרה של סינתזת תבנית מורכבת יותר ואת הצורך אנזים נוסף אוליגונוקלאוטיד. הספציפיות הגבוהה של מחשוף RNase H מקלה על הטיהור בשל היעדר מינים RNA בטווח גודל דומה. הפרוטוקול הנוכחי משתמש בשלב כרומטוגרפיה נוזלית (HPLC) של החלפת יונים שפורסם על ידי מעבדה זו לאחרונה31, אם כי שיטות אחרות הן חלופות אפשריות. 1 H R1ρ RD יכול, באופן כללי, להיות נרכש על דגימות מסומנות או ללא תווית עם שני רצפי פעימה בהתאמה, "מסומן" 1H R1ρ הטרונוקליר יחיד מתאם קוונטי (HSQC) מבוסס ניסוי עם 13 C עקיף ממד 10ואת "לא תווית" 1H R1ρ SELOPE מבוסס על ממד עקיף 1H20.
ניסויים דו-ממדיים אלה (דו-ממדיים) יכולים לשמש בדיקה ראשונה, ללא קשר לשאלה אם הדינמיקה בציר הזמן R1ρ קיימת במדגם. סקירה כללית של מו"פ עבור כל הפסגות שנפתרו בספקטרום ניתן להשיג, ופסגות עניין לניתוח מו"פ יסודי יותר ניתן לזהות. משמעות הדבר היא כי אפילו דגימות ללא תווית ניתן לבדוק לפני החלטה לייצר מדגם יקר יותר, מתויג מתקבל. לאחר שיא עם תרומת חילופי קונפורמציה נבחר להיחקר ביסודיות רבה יותר, עדיף לעבור לגרסאות 1D של הניסויים לעיל (אם השיא עדיין ניתן לפתור) כדי לבצע מה שנקרא ניסויים מחוץ תהודה. עבור הגירסה המסומנת, העברת HSQC ל- 13C מוחלפת בשלב סלקטיבי של קיטוב הטרונוקלרי (HCP) כפי שנעשה שימוש בניסויים 32,33,34,35, בעוד שבמקרה של ניסוי SELOPE, הניסוי פשוט מנוהל כ- 1D, אשר שימושי במיוחד עבור אותות H8 ו- H2 השוכבים באלכסון ב- 2D בכל מקרה. קריטריון אחד לגבי איזה רצף להשתמש, בתנאי ששניהם, מסומנים כמו גם מדגם ללא תווית זמינים, הוא עד כמה מבודד שיא העניין בשני הניסויים.
באופן כללי, הניסוי SELOPE מומלץ עבור דגימות RNA של עד 50 נוקלאוטידים. עבור RNAs גדולים יותר, החפיפה תהיה גדולה יותר; עם זאת, נוקלאוטידים מעניינים מבחינה מבנית מופיעים לעתים קרובות באזורי שינוי כימיים שהם פחות חופפים ועדיין עשויים להיות נגישים ב- RNAs גדולים עוד יותר. טיעון נוסף יהיה כי בדגימות ללא תווית, אין J-צימוד מתרחש בין 1H ו 12C. עם זאת, מכיוון שכוח נעילת הסיבוב המינימלי מוגדר על ידי הכוח המינימלי המשמש לניתוק שני הסיבובים הללו (~ 1 kHz) בניסוי המסומן, הניסוי הלא מתויג מאפשר שימוש בטווח רחב יותר של עוצמות נעילת ספין (SL) ולכן, גישה לטווח זמן רחב יותר של חליפין. ניסויים אלה מחוץ תהודה לספק מידע נוסף kex, כגון אוכלוסיית המצב הנרגש (קונפורמייר חלופי), pES, כמו גם מידע משמרת כימית יקר מאוד בצורה של Δω (ההבדל שינוי כימי של מצב הקרקע ואת המצב הנרגש).

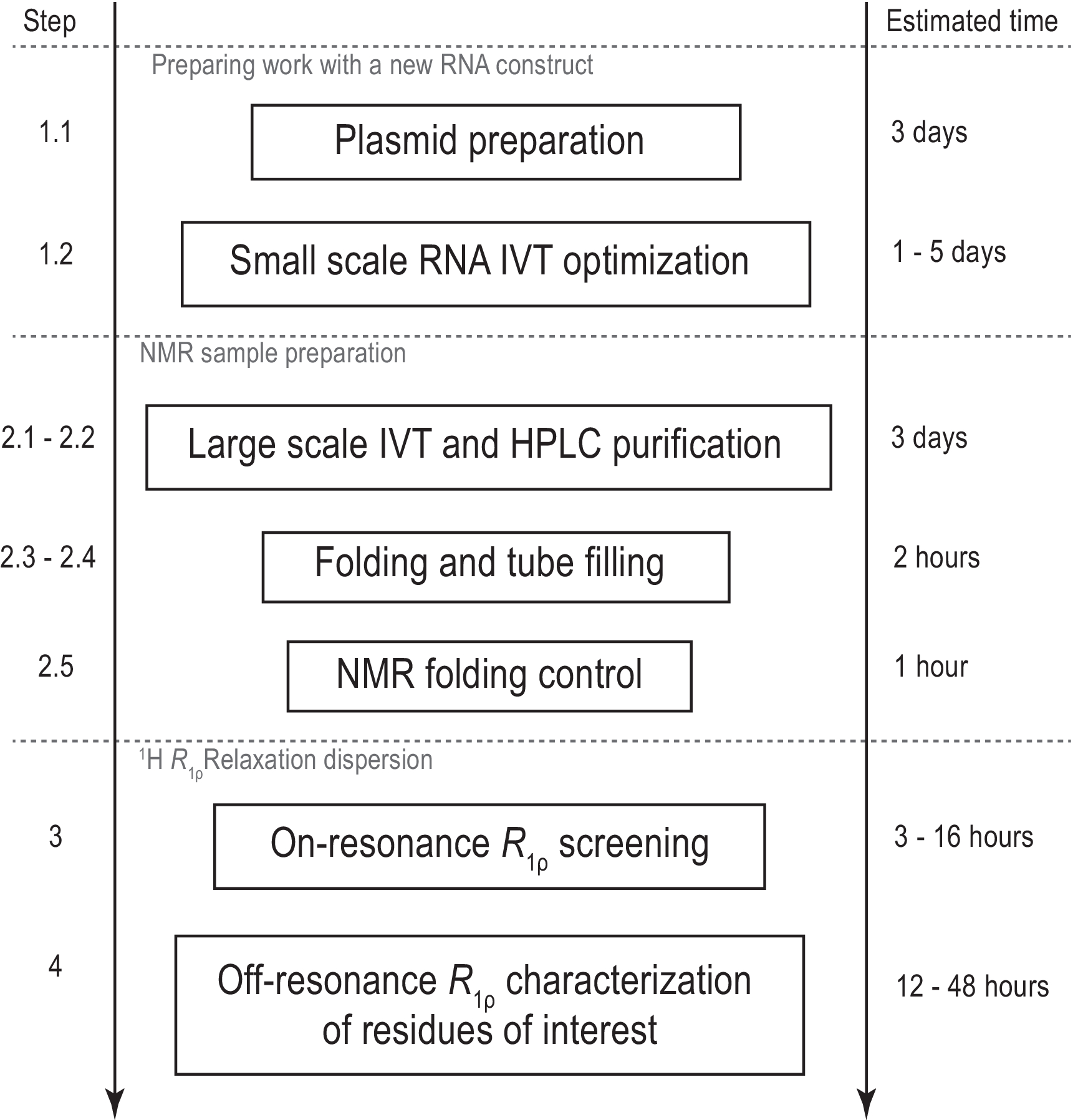
איור 1: זרימת עבודה של הפרוטוקול המוצג. הכנה לפני ייצור מדגם בקנה מידה גדול בפועל, המורכב הכנת תבנית ואישור מוצלח של תמלול במבחנה ומחשוף RNase H. ייצור בקנה מידה גדול כולל טיהור HPLC, מילוי צינור NMR ואישור של קיפול RNA. במקרה של סינתזה עם תווית איזוטופ, יש לבצע טיהור ללא תווית לאופטימיזציה הדרגתית באותו יום. אפיון NMR של דינמיקה קונפורמציה עם ניסויי R1ρ. כל שלב יכול להתבצע באופן עצמאי, למשל., ניתוח 1H R1ρ RD ניתן להחיל על כל דגימת RNA מתאימה המיוצרת בשיטה אחרת. קיצורים: IVT = תמלול במבחנה; HPLC = כרומטוגרפיה נוזלית בעלת ביצועים גבוהים; NMR = תהודה מגנטית גרעינית; RD = פיזור הרפיה. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
מטרת פרוטוקול זה היא לספק פרטיםמעשיים ופרמטרים קריטיים לחקר הדינמיקה הקונפורמציה עם פיזור הרפיה 1 H R1ρ במולקולות סיכת ראש RNA. לאחר מתן פרוטוקול מפורט של טיהור HPLC של תכנון, סינתזה והחלפת יונים של RNA יעד שניתן לבצע באמצעות כל, חלקם, או אף אחד NTPs כגירסאות 13C /15N-labeled, זרימת העבודה של השלמת מדגם NMR ואישור חילופי הקונפורמציה עם ספקטרוסקופיית NMR תוארה. לבסוף, הפרטים על ההתקנה של 1H R1ρ RD ניסויים על ספקטרומטר NMR Bruker מתוארים (איור 1). הפרוטוקול נותן כל שלב כדי להגדיר את גירסת 1D עבור דוגמאות מסומנות בתווית והערות נוספות וטבלה להתאמה עבור ההגדרה של גירסת SELOPE (טבלה 2). לאחר הפרוטוקול, נדונים צעדים קריטיים ודרכים חלופיות להכנת מדגם והגדרת RD 1H R1ρ.
Protocol

איור 2: ייצוג סכמטי של פרוטוקול IVT הדו-מושבי המדווח. (A)תמלול טנדם מתבנית פלסמיד ליניארית עם T7RNAP (משמאל) ומחשוף עוקב של RNAse H של התמליל להשגת RNA באורך היעד, בהנחיית מדריך DNA כימרי (מימין). (B)סכמטי מפורט של תבנית הטנדם החל ממקדם T7RNAP ויראלי, רצף אתחול. רצף היעד (כחול כהה, דוגמה כאן הוא באורך 20 nt) חוזר על עצמו "n" פעמים. החזרות מאגפות רצפי מרווח של 5′ ו- 3′ המורכבים משמונה וארבעת הנוקלאוטידים הראשונים האחרונים, בהתאמה, כדי לאפשר הסרה של רצפי החניכה וההגבלה מהיחידה החוזרת הראשונה והאחרונה. (C)הכלאה של תעתיק הדו-מושבי (אדום) ומדריכי המחשוף הכימרי (ירוק). RNase H מבקע את הרנ"א שממול לסוף ה- DNA 5′. אגפי RNA 2′-OMe מגבירים את הייחודיות על-ידי שיפור הזיקה המחייבת של מדריך המחשוף לרנ"א היעד. נתון זה שונה מ- 21. קיצורים: T7RNAP = T7 RNA פולימראז. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
1. הכנת עבודה למבנה RNA חדש
- עיצוב והכנה של פלסמיד
- כתוב את רצף התבניות בכלי שכפול, למשל., משכפל טורי.
- קח את רצף הקדם T7 והוסף רצף ייזום בתשואה גבוהה (T7: 5'-TAATACGACTCACTATA ^GGGAGA-3').
הערה: התמלול יתחיל בנוקלאוטיד המצוין בתותק (^). רצף החניכה GGGAGA משתנה, אך תלוי רצף מאוד; לכן, השימוש ברצף זה מומלץ. - הוסף את 8 הנוקלאוטידים האחרונים (nt) של רצף היעד כ- spacer בגודל 5 ' (5 S).
- הוסף חזרות של רצף היעד (TS).
- הוסף את ארבעת הנוקלאוטידים הראשונים כמרווח בגודל 3' לאחר החזרות (5 S).
- הוסף אתר הגבלת BamHI (RS) או אתר הגבלה ייחודי דומה.
הערה: הרצף הכולל כפי שמוצג ישוכפל או יוזמן בקלות בפלסטיד בעל עותק גבוה חיידקי (לדוגמה.., pUC19): 5'-T7-5 S-(TS)n-3 S-RS-3' (איור 2B). מספר החזרות צריך להיות גבוה ככל המותר על ידי סינתזת גנים ( מקסימום של 600 nt בפרוטוקול זה). - להגביר את plasmid ב E. coli באמצעות ערכה מסחרית.
- ליניארי את plasmid מטוהר ב 20 ng / μL באמצעות אתר ההגבלה המתאים. הגבלת קנה מידה מתעכלת עם BamHI על ידי עד 1 מ"ל.
- לטהר את plasmid מתעכל, ולאשר ליניאריזציה מוצלחת על ג'ל אגרוז 1%. לאחסן את plasmid ליניארי ב -20 °C (50 °F) במשך כמה חודשים.
- עיצוב מדריך מחשוף (איור 2C)
- כתוב את שמונת הנוקלאוטידים האחרונים של רצף הרנ"א היעד בכיוון 5'-3', והוסף את ארבעת הנוקלאוטידים הראשונים של רצף הרנ"א של המטרה בקצה 3', גם בכיוון של 5'-3.5.
- צור את המשלים ה- DNA ההפוך של רצף זה
- שנה את ארבעת הנוקלאוטידים הראשונים והאחרונים לשינויי 2'-OMe שלהם על-ידי הוספת 'm' לפני אות הנוקלאוטיד.
הערה: עבור סינתזה, mU משמש במקום mT. - הזמינו את האוליגו עם טיהור התפלה סטנדרטי.
הערה: בדוק אם אוליגו שנוצר יכול לאגד במקום אחר מלבד החיבור של שני רצפי RNA. נדרשת השלמה מלאה בארבעת הנוקלאוטידים המרכזיים של הדנ"א, בעוד שאזורי האגף עלולים לאפשר אי התאמה. במידת הצורך, הרחב את האגפים עד 18 nt כדי ליצור רצף איגוד ייחודי36.
- IVT בקנה מידה קטן
הערה: לעבודה ללא RNase, הכינו את כל הריאגנטים בתנאים סטריליים וללא RNase. השתמש RNase טיהור reagent (ראה את טבלת החומרים) ו 95% v / v אתנול כדי לנקות משטחי עבודה ופיפטות לפני השימוש. יש לשטוף כפפות עם 95% אתנול וללבוש בגדים עם שרוולים ארוכים ללא מוך. כדי למזער את זיהום RNase, אין לנשום מעל צינורות פתוחים.- הכן פתרונות מלאי של Tris-Cl (pH 8.0), dithiothreitol, MgCl2, spermidine, ו NTPs / GMP (ללא מנוצל). ערבבו ריאגנטים כפי שמוצג בטבלה 1. הכן תערובת מאסטר של ריאגנטים אלה מראש, לפני תוספת של אנזימים או חומצות גרעין.
הערה: אם אתם משתמשים בריגנטים קפואים, מערבבים אותם ביסודיות לאחר ההפשרה. ריאגנטים עשויים לזרז אם מעורבים בריכוזים גבוהים מדי, ולכן מומלץ מאוד לעקוב אחר הסדר בטבלה 1. - הוסף את הסדר הבא: plasmid, מדריך מחשוף, פוספטאז אנאורגני (IPPase), RNase H, T7RNAP. כמו פעילות אנזימים עשוי להשתנות עבור אנזימים המיוצרים בתוך הבית, לבדוק כמה ריכוזים לפני בחירת הטוב ביותר.
הערה: כלול שליטה שלילית עבור תגובת המחשוף, למשל, ללא RNase H, כדי לייחס רצועת יעד חסרה למחשוף RNase H פגום ולא לתמלול לא מוצלח. - לדגור על התגובה ב 37 °C במשך 1 שעות ולאשר תגובה על אלקטרופורזה ג'ל polyacrylamide denaturing (PAGE) (איור 3A). לדלל את פתרון הטעינה של הדגימה פי 10, ולהעמיס 1 μL על הג'ל.
הערה: תערובת ג'ל: 8 M אוריאה, 20% אקרילאמיד (19:1 אקרילאמיד:ביסאקרילמיד) ב 1x TBE. פתרון טעינה: 5 mM אתילנדיאמין חומצה טטראצטית (EDTA), 300 μM ברומופנול כחול פורממיד. תגובות מחשוף RNase H לא ניתן לצפות להיות שלם לאחר 1 שעה, כמו RNA חדש מיוצר כל הזמן. בשלב זה, חפשו רצועת יעד ברורה והיעדר מין בעל משקל מולקולרי דומה (למשל,±3 מוצרי נוקלאוטיד (nt).
- הכן פתרונות מלאי של Tris-Cl (pH 8.0), dithiothreitol, MgCl2, spermidine, ו NTPs / GMP (ללא מנוצל). ערבבו ריאגנטים כפי שמוצג בטבלה 1. הכן תערובת מאסטר של ריאגנטים אלה מראש, לפני תוספת של אנזימים או חומצות גרעין.
| מגיב | ריכוז מלאי | כמות בקנה מידה קטן (μL) |
| ח2O | - | 24 |
| טריס | 1 מטר | 5 |
| MgCl2 | 1 מטר | 0.5 |
| DTT | 1 מטר | 0.5 |
| זרעון | 250 מ"ר | 5 |
| GMP | 100 מ"ר | 2.5 |
| ATP | 100 מ"ר | 1.5 |
| GTP | 100 מ"ר | 1.5 |
| UTP | 100 מ"ר | 1.5 |
| CTP | 100 מ"ר | 1.5 |
| פלסמיד | 20 ננוגרם/μL | 5 |
| מדריך מחשוף | 100 מיקרומטר | 10 |
| iPPase | 10 מ"ג/מ"ל | 0.5 |
| RNase H | 10 מיקרוגרם/מ"ל | 2 |
| T7 RNA פולימראז | 5 מ"ג/מ"ל | 2 |
טבלה 1: שולחן ריאגנט עבור IVT דו-מושבי ומחשוף RNase H בו זמנית. ניתן להתאים את ריכוזיהמלאילנוחיות המשתמש. אם מחשוף RNase H חייב להתבצע לאחר T7 IVT, הוסף מדריך מחשוף ו- RNase H לאחר אי-החלמת חום של T7RNAP. כמויות המשמשות בקנה מידה ליניארי עם סולם תגובה. קיצורים: T7RNAP = T7 RNA פולימראז; IVT = תמלול במבחנה.
2. הכנת מדגם NMR
- שפר את התגובה לנפח הרצוי (בדרך כלל 10 מ"ל), והפעל את התגובה בן לילה. בדיקה להשלמת תגובה למחרת עם ג'ל PAGE denaturing(איור 3A).
הערה: תגובת מחשוף לא שלמה מוצגת על ידי מינים במשקל מולקולרי גבוה יותר מעל רצועת היעד.- אם המחשוף לא הצליח או הושלם, רנ"א reanneal ואת מדריך המחשוף בכלי התגובה על ידי חימום הפתרון במיקרוגל קונבנציונאלי ב 450 W עבור 15 s.
- מצננים את הפתרון לאט ל-37 מעלות צלזיוס למשך 40 דקות. השתמש בלוק חימום עבור אמצעי אחסון מתחת 1 מ"ל. שים לב להיווצרות של משקעים חדשים.
- הוסף עוד IPPase ו- RNase H, ודגרה לעוד 1-3 שעות ב 37 °C (70 °F). אשר את השלמת תגובת המחשוף עם דף denaturing.
- כאשר תגובת מחשוף RNase H הושלמה, להרוות את התגובה על ידי הוספת EDTA לריכוז הסופי 50 mM מערבולת ביסודיות.
הערה: משקעים פירופוספט פוטנציאליים יתמוססו, וצורות מזרזות חלבון חדשות. - סנן את הפתרון באמצעות מסנן מזרק של 0.2 מיקרומטר והתרכז באמצעי אחסון להזרקה במערכת HPLC, בהתאם לגודל לולאת ההזרקה.
הערה: הפרוטוקול יכול להיות מושהה כאן על ידי הקפאת דגימה ב -20 °C (50 °F).
- טיהור HPLC בקנה מידה גדול
- הכן מאגרי החלפת יונים A ו- B תוך שבוע מהשימוש. סנן ודגה את המאגרים.
הערה: חוצץ A: 20 mM נתרן אצטט; 20 מ"מ נתרן פרכלוראט, pH 6.5. חוצץ B: 20 mM נתרן אצטט; 600 מ"מ נתרן פרכלוראט, pH 6.5. - שווה את העמודה עם 100% מאגר B ואחריו 100% מאגר A עבור לפחות 2 אמצעי אחסון עמודות ב 75 °C (75 °F).
- הכן את רצף HPLC(איור 3B)בקצב זרימה של 5.5 מ"ל/דקה. השתמש ברצף הבא לטיהור של RNA בגודל בין 20 ל 30 nt: 0-7 דקות: 0% B; 7-16 דקות: הדרגתי 0-20% B; 16-46 דקות: elution, בדרך כלל עם שיפוע של 20-30% B (לייעל לפי הצרכים); 46-62 דקות: 100% B; 62-73 דקות: 0% B.
הערה: שינוי בקצב הזרימה מ- 5.5 ל- 8 מ"ל / דקה לא השפיע על ההפרדה בפרוטוקול זה. - מטב את שיפוע ההזרקה על-ידי הזרקה של מ"ל אחד של תגובת שעתוק (ללא תווית) בכל פעם.
הערה: לפרטים נוספים ולדיון, עיין בקרלסון ואח '31 ופיירר ואח '21. - בדוק את השברים שנאספו בדף denaturing. אם פסגת ההעפלה הראשית מבודדת היטב ומכילה את הרנ"א המטרה הטהור, גדילו את הטיהור לשווי של 10 מ"ל של תגובת שעתוק.
- לאסוף את שברי העניין, להתרכז ולהחליף את המאגר עם מאגר NMR. השתמש ביחידת סינון אולטרה-צנטריפוגלית (עיין בטבלת החומרים) עבור אמצעי אחסון מעל 50 מ"ל.
הערה: חוצץ NMR: 15 mM נתרן פוספט; 25 מ"מ נתרן כלורי; 0.1 מ"מ EDTA, pH 6.5. כדי למזער את האובדן מ- RNA ההצמדה לקירות צינור פלסטיק, יש לשטוף את כל צינורות האיסוף עם 1 מ"ל של מים, מערבולת וצנטריפוגה כדי לאסוף את כל הנוזלים. - לקבוע את הריכוז באמצעות ספקטרוסקופיה אולטרה סגולה. חשב את תשואת התגובה על פי פיירר ואח' 21.
הערה: הריכוז של מדגם NMR לניסויי RD לא צריך להיות מתחת 130 נמול, אשר מתאים 500 μM בנפח מדגם של 250 μL באמצעות צינורות NMR(טבלת חומרים).
- הכן מאגרי החלפת יונים A ו- B תוך שבוע מהשימוש. סנן ודגה את המאגרים.
- קיפול של דגימת RNA
- לדלל aliquot המדגם של נפח של ~ 10 מ"ל לתוך 1 מ"ל לכל צינור.
- מחממים את עליקות הרנ"א ל-95 מעלות צלזיוס למשך 5 דקות.
- מצננים את הדגימות על ידי הנחתן על קרח או בתערובת מלח-קרח במים ודגרה במשך 30 דקות.
- דגימות בריכה ולהתרכז ~ 250 μL ביחידת מסנן צנטריפוגלי 2 מ"ל.
- מילוי צינור NMR
- נקה את צינור NMR בניקוי הצינור NMR על ידי שטיפה עם מים בשפע, ריאגנט טיהור RNase, מים, 95% אתנול (EtOH), ומים שוב. השאירו לייבוש.
- נקו את הבוכנה על ידי שטיפה במים וניגוב עם ריאגנט טיהור RNase ו-95% EtOH באמצעות מגבון ללא מוך. השאירו לייבוש.
- הוסף 10% (v/v) של D2O למדגם NMR.
- מלא את דגימת הרנ"א לתוך צינור NMR באמצעות קצה פיפטה גדול. תן לנוזל לזרום לאורך הצד של קיר הצינור.
- הכנס את הבוכנה והסר בועות אוויר על ידי דחיפת הבוכנה כלפי מטה יחד עם תנועת פיתול מהירה.
- משוך את הבוכנה למעלה לאט מבלי ליצור בועות אוויר חדשות ולתקן אותו עם סרט שעווה פרפין.
- אשר קיפול על ידי NMR.
הערה: בשלב זה, יש צורך לבצע לפחות משימת תהודה חלקית כדי לאשר את המבנה המשני של מדגם RNA ולזהות אזורים מעניינים לחקר הדינמיקה הקונפורמיאלית. תיאור ממצה על הקצאת תהודה RNA יעלה על פרוטוקול זה, ולכן אנו מתייחסים לספרות מבוססת היטב בשלב זה19,37,38. משמרות ניידות אלקטרופורטיות (EMSA) יכולות להיות אינדיקטור מועיל של קיפול RNA ולשמש כנתונים משלימים לניסויי NMR.- השווה את הספקטרום הבא של המדגם שעבורו מבוצעים ניסויי RD של 1H R1ρ עם מדגם הייחוס המקופל כראוי (איור 4): 1H 1D, במיוחד אזור האימינו 10-15 ppm; ארומטי 1H,13C-HSQC; 1 H,1H-SELOPE (אופציונלי).
הערה: טביעת אצבע ארומטית נחוצה גם, גם במקרה של הסכמה בין אותות imino, כי היווצרות dimer לעתים קרובות מראה אותו או דומה אותו אותות imino כמו סיכת ראש RNA. ניסוי SELOPE יכול להחליף 1 H,13C-HSQC עבור טביעת אצבע ארומטית, כמו ניסויים heteronuclear על דגימותללא תווית הם מאוד זמן רב. - השתמש בלולאת UUCG כהפניית טביעת אצבע (אם קיימת).
- בצע השוואהזו בכל פעם לפני 1 H R1ρ RD ניסויים נרשמים.
- השווה את הספקטרום הבא של המדגם שעבורו מבוצעים ניסויי RD של 1H R1ρ עם מדגם הייחוס המקופל כראוי (איור 4): 1H 1D, במיוחד אזור האימינו 10-15 ppm; ארומטי 1H,13C-HSQC; 1 H,1H-SELOPE (אופציונלי).
3. 1H R1ρ פיזור הרפיה-עלתהודה (מסומן 1D גרסה)
הערה: השלבים שלהלן מתארים את ההתקנה של ניסויי RD עבור מדגם עם תווית באמצעות הגירסה 1D של רצף דופק RD מבוסס HSQC. בצע את אותם שלבים עבור רצף 1D מבוסס SELOPE עבור דגימות ללא תווית. מבט כולל על שמות פרמטרים והגדרות עבור שני האירועים מוצג בטבלה 2. ההתמקדות בגרסאות 1D נובעת מכך שהן מעשיות יותר למדידות מחוץ לתהודה, והגדרת גרסאות הדו-ממד של הניסויים מבוססי SELOPE ו- HSQC נדונו בפירוט על ידי Schlagnitweit ואח'20 ושטיינר ואח'10, בהתאמה.
- קבעו הספק של 1שעות לדופק קשה של 90° (P1).
- אפשרות א': השתמש בפקודה דופק ברוקר.
- אפשרות ב': בניסוי zg, קבעו את הדופק של 360מעלות על ידי מדידת עקומת אגוזים ברמת הכוח של הפרוטון בדופק הקשה על פסגת המים.
הערה: אורך הדופק של 90° הוא רבע משך הזמן שבו אות אפס נמדד (אם נמדדת עקומת אגוז מלאה, אז זה האפס השני; עם זאת, בפועל, רק האזור סביב הערך הצפוי עבור 360° נדגם).
- הפעל ספקטרום 1 H 1D zgesgp.f2f3dec באמצעות אורךהדופק שנקבע בשלב 3.1 כדי לאשר את קיפול ה- RNA לפני כל מדידת R1ρ.
הערה: אם 1ניסויי H SL מופעלים בפעם הראשונה, בדוק אם כוח ה- SL המחושב תואם לחשמל המועבר לדגימה על-ידי כיול כוח SL עבור כל רוחב פס רצוי. שלבי כיול מפורטים מתוארים בשטיינר ואח '10. - צור 1H R1ρ עבור ערכת נתונים עם תווית והגדר פרמטרי מפתח.
- צור ערכת נתונים חדשה; מבוסס באופן אידיאלי על ערכת נתונים ארומטית 1H-13C HSQC כפי שנעשה בה שימוש בדגימות RNA בעלות תווית מלאה להקצאת RNA.
הערה: פעולה זו תבטיח כי 13C, כמו גם כוח 15N וכוח ניתוק כבר מוגדרים. - הגדר את הפרמטרים הכלליים בהתאם לחלק הראשון של טבלה 2.
- הגדר פרמטרים ספציפיים RD בהתאם לחלק השני של טבלה 2.
- הגדר הספק SL של 1H לערך הנמוך ביותר (1.2 קילו-הרץ) לבדיקה.
- צור רשימת VD לבדיקה עם ערך אחד בלבד, 0 ms, (כדי למטב את רשימת vd, כמתואר בשלב 3.4), הגדר את TDF1 ל- 1 ועדכן D30.
- הפעל ספקטרום בדיקה עם הגדרות אלה.
- צור ערכת נתונים חדשה; מבוסס באופן אידיאלי על ערכת נתונים ארומטית 1H-13C HSQC כפי שנעשה בה שימוש בדגימות RNA בעלות תווית מלאה להקצאת RNA.
- מטב את רשימת ה- VD (רשימה של אורכי SL לשימוש).
- הפעל את הניסוי עם רשימת VD מבחן (למשל., שישה ערכים: 0 מ ', 5 מ ', 10 מ ', 20 מ ', 30 מ ', 40 מ '; לטרוף ערכים אלה כדי למנוע שגיאות שיטתיות עקב חימום).
- עדכון D30 ו- TDF1 בהתאם (בדוגמה זו, D30 = 42m ו- TDF1 = 6).
- עוצמת התוויית השיא לעומת אורך SL. זהה את אורך ה- SL שבו עוצמת הפסגה המקורית יורדת ל- 1/3.
- צור את רשימת VD הסופית לשימוש בניסוי, תוך התחשבות באפשרויות הבאות: לקבוע את אורך ה- SL הארוך ביותר כמתואר בשלב הקודם; הימנע משימוש ברשימה בסדר יורד או עולה; ולהוסיף כמה כפילויות למחקרים סטטיסטיים. זכור לעדכן D30 ו- TDF1 בכל פעם שיש שינויים ברשימת vd.
הערה: הניסוי מנוהל עם אורכי SL שונים כפי שניתן ברשימת VD בצורה פסאודו-2D. - בחר את מספר הסריקות כך שהשיא החלש ביותר ברשימה יהיה בעל יחס אות לרעש (SINO) של לפחות 10.
הערה: למרות שרשימת vd הייתה ממוטבת עבור הספק SL נמוך (1.2 kHz), רשימת vd זו צריכה להיבדק גם בהספק ה- SL הגבוה ביותר לשימוש (לדוגמה.., 15 kHz). הסיבה לכך היא שהריקבון יהיה הרבה יותר איטי בעוצמה גבוהה של SL לפסגות עם תרומה משמעותית של kEX. לכן, ריקבון מספיק צריך להיות מאומת גם בעוצמה SL גבוהה.
| תיאור פרמטר | שם פרמטר ברצף דופק | |
| 1D עם תווית | סלופה 1D | |
| תוכנית דופק עבור 1Ds בתהודה | 1HR1rho_HCP_onres1D.es | 1HR1r_HH_onres1D.js |
| 1 תדרי ספק H (ppm) | O1P = תהודה במים ב- ppm | O1P = שינוי כימי של שיא העניין (ppm) |
| CNST28 = שינוי כימי של שיא עניין (ppm) | CNST29 = תהודה במים ב-ppm | |
| 1 דופק 90 מעלות קשה | P1 @ PL1 (כפי שכויל ב- 3.1.1) | P1 @ PL1 (כפי שכויל ב- 3.1.1) |
| פולסים מעוצבים וכוחות לדיכוי מים | P25 = 1000 us @ sp3 | P12 = 2000 us @ sp1 |
| (ווטרגייט) | (פיסול עירור) | |
| 13 תדר נושא C, על תהודה עם 13C שינוי כימי של שיא עניין | O2P | – |
| 15 תדר נושא N, שינוי כימי ממוצע של 15N לפירוק (כפי שנעשה שימוש ב- HSQC ארומטי) | O3P | – |
| 13 C/15N ניתוק (מוגדר כמו ב- HSQC) | pcpd2, cpd2 | – |
| pcpd3, cpd3 | ||
| העברת HCP (למשל, p = 1/ J @ 100 הרץ) | – | |
| ניתן להשתמש בפקודות pulse ו- pulsef2 כדי לקבוע כוחות מפולסים קשים | ||
| משך הזמן (מוגדר ל- 1/J(1H-13C) של שיא עניין) | P11 | |
| הספק 1H ו-Power ב-13C | SP1, SP12 | |
| העברת סלופה (ד = 1/4J(H5-H6)) | – | D5 |
| דופק סלקטיבי (למשל, אזור ארומטי) עבור SELOPE (4000 לנו, Eburp) | – | P13 ו- SP4 |
| פרמטרים ספציפיים ל- SL / RD: | ||
| 1 כוח H SL, המתקבל מדופק קשיח מכויל (למשל, באמצעות פקודת הדופק). | Pl25 ו- CNST12 (1.2 – 15 קילו-הרץ) | Pl24 (50 הרץ – 15 קילו-הרץ) |
| רשימת השהיית משתנה עבור משך SL (בתחילה ערך 1, 0, מיטוב המתואר תחת 3.1.3) | vdlist (~ 0 – 40 ms) | vdlist (~ 0 – 150 ms בשל Rנמוך 2 בדגימות ללא תווית) |
| TDF1 מספר הערכים ברשימת vd (בתחילה 1) | TDF1 | TDF1 |
| פיצוי חום: | ||
| D30 = הערך הגדול ביותר ברשימת vd + 2ms | D30 | D30 |
| פיצוי חום נוסף עבור מגוון רחב מאוד של SLs | PL25 | |
| פרמטרים ספציפיים מחוץ לתהודה: | ||
| תוכנית דופק עבור 1Ds מחוץ תהודה | 1HR1rho_HCP_offres1D.es | 1HR1r_HH_offres1D.js |
| היסט לניסויי מחוץ לתהודה | CNST30 | CNST30 |
טבלה 2: מבט כולל על פרמטרים כדי להגדיר ניסויים מבוססי HCP מבוססי 1D ו- 1D-SELOPE 1H R1ρ. קיצורים: 1D = חד ממדי; HCP = קיטוב צולב הטרונונוקלרי; SELOPE = ניסוי פרוטון ממוטב סלקטיבי; ppm = חלקים למיליון; HSQC= מתאם קוונטי ספין הטרונוקלארי; SL = נעילת ספין; RD = פיזור הרפיה
- הקמה ורכישה של ניסויי H R1ρ בתהודה
- העתק את הניסוי מסעיף 3.4 לתיקיה חדשה בטופספין.
- בתיקיה זו, הגדר ניסויים בעוצמות SL שונות, בכל פעם שינוי PL25 ו- CNST12. קבע את רמת ההספק הנכונה עבור כל עוצמת SL באמצעות הפקודה pulse. השתמשו בעוצמות SL הנעות בין 1.2 ל-15 קילו-הרץ, עם דגימה צפופה יותר לעוצמות SL נמוכות יותר (ראו איור 5G לעוצמות SL נבחרות). הוסף עותקים של חלק מהניסויים כדי שיהיו כפילויות עבור חלק מכוחות ה- SL.
- תריץ את הניסויים האלה.
- ניתוח של ניסויים בתהודה 1H R1ρ
- ב- TopSpin, עבד כל פרוסה של כל ערכת נתונים מדומה-דו-פעמית באמצעות אותם פרמטרי עיבוד (למשל, הרחבת קו, שלב) באמצעות הפקודה xf2,ופצל את ערכת הנתונים ל- 1D באמצעות תוכנית Bruker AU split2D.
- השגו עוצמות ואמצעי אחסון של אותות עבור כל פרוסה 1D.
הערה: בפועל, עדיף לפרק את הספקטרום כדי להיפטר תרומות פסגות חופפות פוטנציאלית ומאפשר את השימוש של Bruker AU תוכנית multidcon, אשר מסכם בנוחות את האינטנסיביות או האזורים של הפסגות של כל הפרוסות בניסוי אחד בקובץ הטקסט decall.txt, אשר לאחר מכן ניתן לקרוא בקלות עם תוכניות אחרות (סקריפטים פייתון שנכתבו בתוך הבית שימשו כאן, כפי שתואר על ידי שטיינר ואח'10) בשלבים 3.6.3 ו 3.6.4. - התאם ריקבון מעריכי מונו עבור כל כוח SL כדי להשיג את הערך R1ρ (או על תהודה, R2+REX).
- התווה ערכי R2+REX (y) לעומת. חוזק SL (x) (איור 5F, G).
הערה: אם הערכים גבוהים משמעותית עבור עוצמות SL נמוכות ויורדות עם כוח SL גבוה יותר (כפי שמוצג באיור 5G), אז השיא הנחקר מראה פיזור, וזה עשוי להיות מעניין לבצע ניסויים נוספים (מחוץ תהודה) כדי להשיג מידע על האוכלוסייה הבדל שינוי כימי של המצב הנרגש לעומת. מצב הקרקע.
4. 1H R1ρ פיזור הרפיה – מחוץ לתהודה (מסומן 1D גרסה)
- הקמה ורכישה של ניסויים מחוץ לתהודה 1H R1ρ
- בתיקיה חדשה topspin, הגדר ניסויים בעוצמה SL מסוימת (בדרך כלל תחילה בעוצמה SL הנמוכה ביותר כמו תרומת REX הוא הגבוה ביותר שם, ראה איור 5G עבור מבחר מייצג של SLs מחוץ תהודה), אבל עם קיזוזים שונים, בכל פעם שינוי CNST30.
- השתמש בהיסט של עד ± (3 או 4)*SL, עם דגימה צפופה יותר סביב היסט 0, כפי שניתן לראות באיור 5H,I.
- תריץ את הניסויים האלה.
- ניתוח של ניסויים מחוץ לתהודה 1H R1ρ
- השתמש באותה אסטרטגיית עיבוד, כמו ב- 3.6.1– 3.6.3, כדי לקבוע ערך R1ρ עבור כל היסט.
- התווה ערכים אלה לעומת היסט (איור 5G).
הערה: אסימטריה בעקומה זו כבר יכולה להצביע על כך שניתן להשיג מידע על שינויים כימיים עבור המצב הנרגש. התאמה וניתוח יסודיים באמצעות משוואות בלוך-מקונל או לגואר צריכים להתבצע כדי לקבל מידע על kEX, pES, כמו גם Δω10,20 (איור 5G). ניתן למצוא ערכות נתונים לדוגמה, תוכניות דופק ופקודות מאקרו עבור שני הניסויים ה-1D במאגר Github במעבדת Petzold (https://github.com/PetzoldLab). מבט כולל על פרמטרים ניתן בטבלה 2.
תוצאות
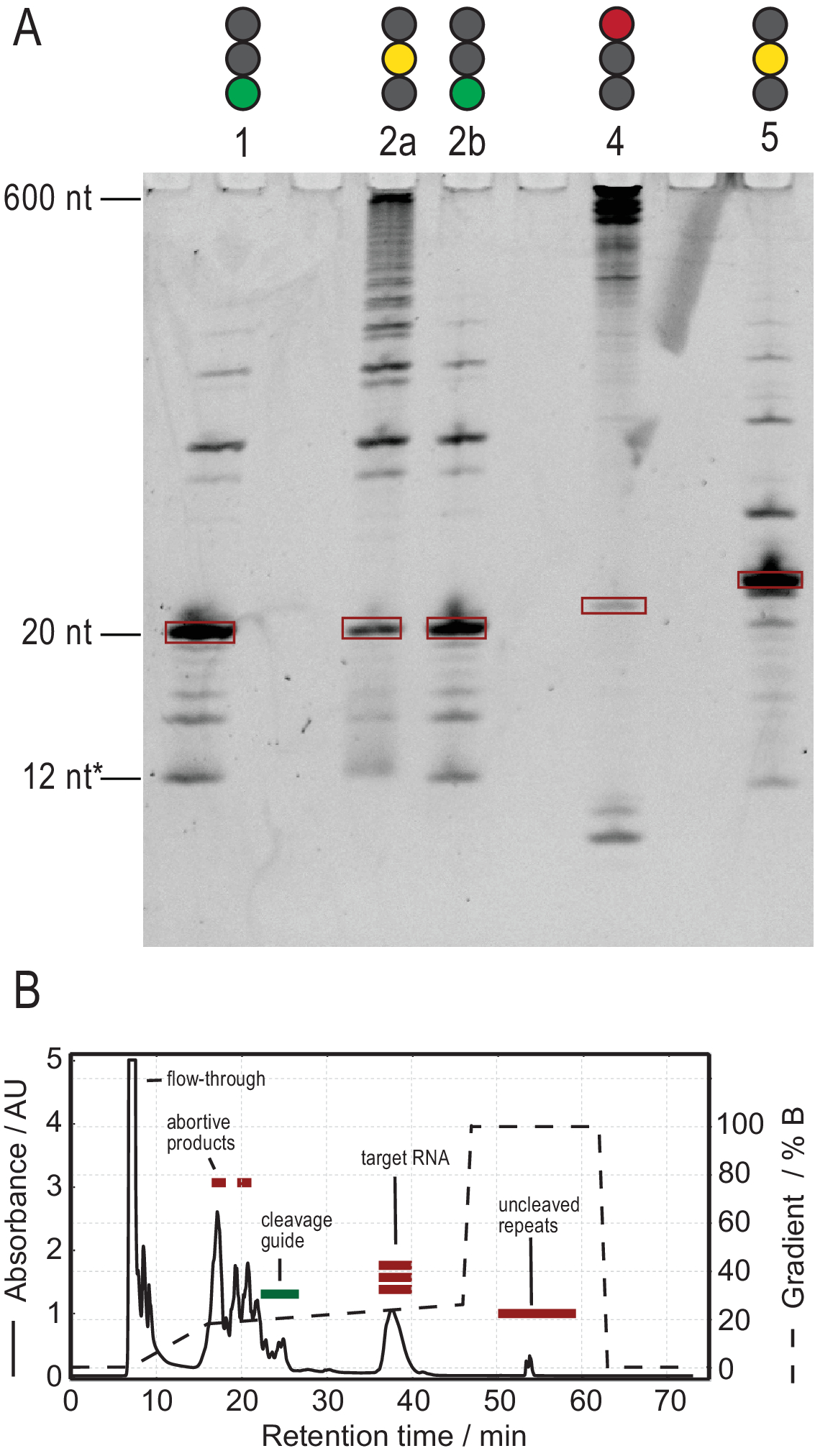
הפרוטוקול לייצור RNA מאפשר טיהור באמצעות יצירת תמלילי טוהר גבוה. איור 3A מציג את התוצאות של מספר תגובות מחשוף של תמלילים דו-מושביים, ומספק תגובות מוצלחות ולא מוצלחות. ליין 1 מציג את המקרה האופטימלי של תעתיק מחשוף לחלוטין עם עקבות קלושים בלבד של מוצרי צד. ליין 2a מציג מחשוף לא שלם, אשר ניתן לפתור על ידי חישול מחדש ותוספת של RNase H יותר (נתיב 2b, שלב 2.1.2). מבני הרנ"א של נתיבים 1, 2a ו- 2b זהים. המדגם בנתיב 3 מראה מחשוף לא מוצלח. פתרון בעיות תגובה זו יכלול בדיקה של רצף מדריך המחשוף, טוהר תבנית ה- DNA וטמפרטורות חישול. באופן פוטנציאלי, מחשוף RNase H יצטרך להתבצע לאחר T7 IVT כפי שמוצג עבור מדגם 2.
המדגם בנתיב 4 מראה כמות משמעותית של מוצרים בצד המחשוף, שקשה להסירם באמצעות HPLC חילופי יונים. פתרון בעיות מדגם כזה יכול לכלול (א) הורדת טמפרטורה, כמות RNase H או זמן תגובה, (ב) הפחתת שיפוע elution ונפח הזרקה וניסיון להפריד את שברי היעד ממוצרי הצד. מידע נוסף על אופן הגדלת הרזולוציה בטיהור HPLC חילופי יונים נדון על ידי קרלסון ואח'31. HPLC מפרידה את הרנ"א היעד מחומצות גרעין ארוכות או קצרות יותר ומחלבונים או מזהמים של מולקולות קטנות. איור 3B מציג את התוצאה האופטימלית לטיהור HPLC של חילופי היונים. יש לבחור את שיפוע ההעמקה כך שמין ה-RNA היעד חומק לפחות מנפח עמודה אחד (בדוגמה זו: 35 מ"ל) לאחר המינים הקטנים הבאים ונפח עמודה אחד לפני המין הגדול הבא.
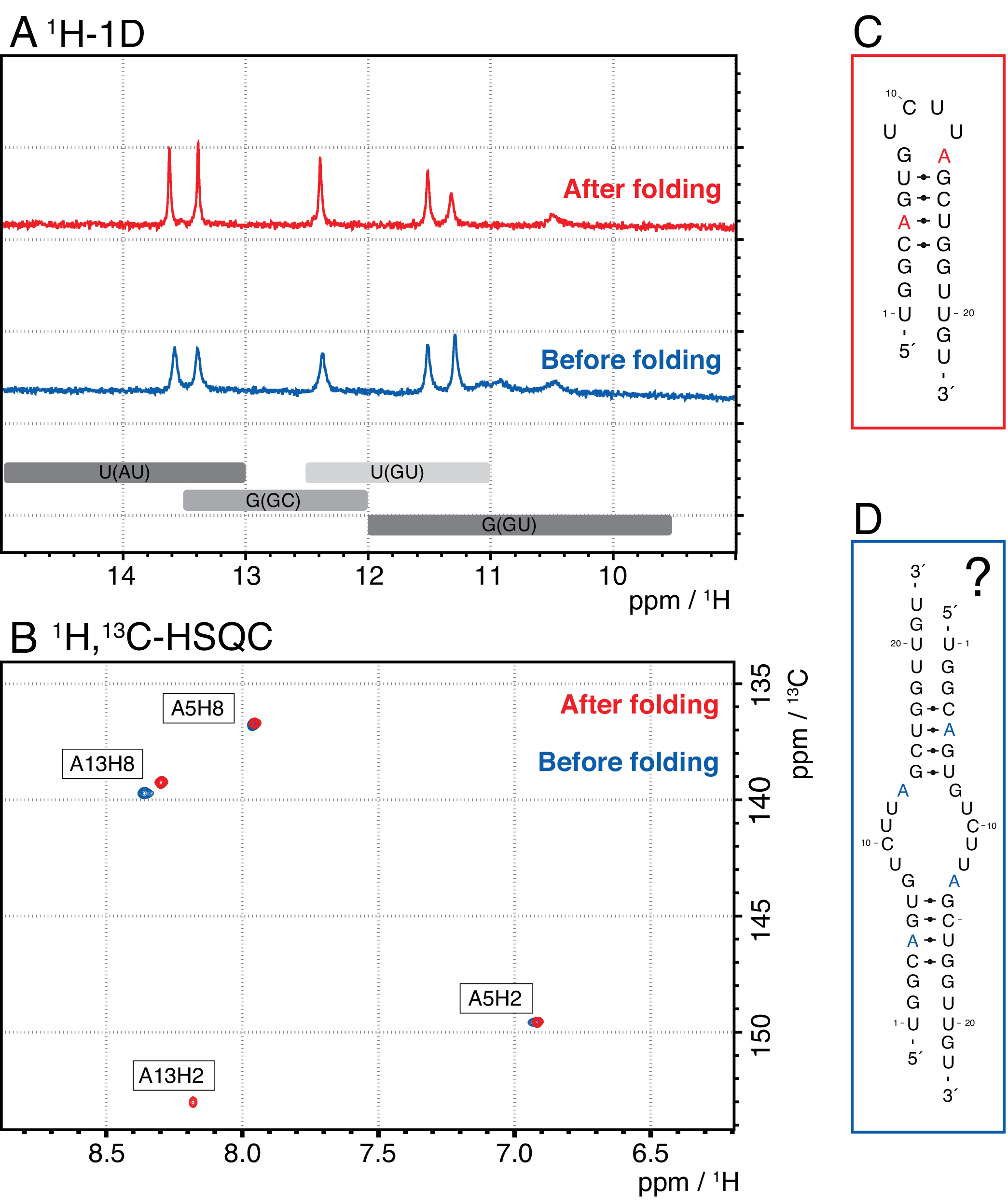
מינים קטנים יותר בשיטה זו כוללים נוקלאוטידים בודדים, מוצרים מפלים (8-12 nt), רצפי spacer 3' ו-5' (5-14 nt) ומדריך מחשוף (12 חומצת גרעין כימרית nt), בעוד רצפים ארוכים יותר הם חוזרים טנדם טמאים פוטנציאליים ואת plasmid. כאשר מושגת פסגת אלוטיון מופרדת היטב, ניתן להרחיב את הטיהור עד למקבילה של ~ 20 מ"ל של תגובת IVT לכל זריקה. הקיפול הנכון של דגימת RNA חיוני לניסויי מו"פ ויש לאשרו לפני כל מדידה. איור 4 מציג RNA של 22 מרים עם תווית A לפני החלת פרוטוקול הקיפול בשלב 2.4 (כחול), ואותה דגימה לאחר הקיפול הנכון הושגה (אדום). תחזית מבנה משני Mc-Fold(איור 4C)מציעה את מבנה סיכת הראש המוצג עם 4 זוגות בסיסים וכתוצאה מכך 5 אותות imino.
שתי הספקטרום באיור 4A מאשרות את האותות החזויים הללו, אם כי בעוצמות יחסיות מעט שונות, מה שמצביע על כך שמבנה מסוים (כאן, דימר) יכול להיות בעייתי להערכה עם ספקטרום 1H 1D בלבד. ספקטרום ארומטי של 1H,13C-HSQC(איור 4B),עם זאת, מראה רק 3 מהאותות הארומטיים לדגימה לפני פרוטוקול הקיפול (כחול), אך כל 4 האותות לדגימה שקופלה לפי שלב 2.4 (אדום). המדגם המוצג בכחול יצר ככל הנראה הומודימר (מבנה שהוצע באיור 4D)שיגרום לאותות אימון זהים כמו סיכת הראש. האות של A13H2 נראה מורחב חילופי. תוצאות אלה מסייעות להדגיש את החשיבות של קיפול אישור עם ניסויי טביעות אצבע אימוניים וארומטיים לפני כל ניסוי מו"פ. רצפי פעימות 1H R1ρ המתוארים בפרוטוקול זה מאפשרים זיהוי דינמיקה במשטר חילופי הביניים. בתחילה נרשמת עקומת תהודה, ואם קיימת דינמיקה עבור שאריות ספציפיות, ניתן לראות פיזור בערכי R2+REX שהושגו, בעוד שעקומה זו שטוחה עבור שאריות ללא החלפה.
איור 5 מציג עקומות מייצגות על תהודה המתקבלות עבור שני אטומי H8 שונים בסיכת ראש סינתטית של RNA (איור 5A),שבה G6H8 חווה חילופים (איור 5C), בעוד A4H8 אינו (איור 5B). מכיוון שההחלפה איטית יחסית במדגם זה (kEX = 292 ± 40 הרץ), היתרון של ניסוי SELOPE להשגת עוצמות SL נמוכות נוצל, ושתי עקומות התהודה נרשמו באמצעות גרסת 1D של רצף הדופק. אותו רצף דופק שימש אז להשגת נתוני תהודה עבור השאריות המציגות פיזור בפרופיל התהודה. איור 5D מציג את ערכי R1ρ המתקבלים לעומת היסט שבו א-סימטריה קלה של העקומה כבר מציינת את הסימן של Δω.
זה הופך להיות אפילו יותר ברור בעלילת R2+ REX שבה מוסרת תרומת R1 (איור 5E). העמודה הימנית של אותה דמות מציגה עקומות ייצוגיות על תהודה המתקבלות עבור שני אטומי H8 שונים בסיכת ראש סינטטית מעט שונה עם חילופי RNA מהירים יותר, כאשר G6H8 חווה חילופים (איור 5G), ואילו A4H8 לא (איור 5F). שער החליפין המהיר יותר (kEX = 43,502 ± 38,478 הרץ) אפשר את הקלטת המו"פ של כל הפרוטונים הארומטיים בבת אחת באמצעות גרסת SELOPE 2D כדי להשיג נתונים, על ומחוץ לתהודה (נתוני G6H8 המוצגים באיור 5H,I).
מזהים כלליים עבור תוצאות חיוביות ושליליות
תוצאות חיוביות מחשוף ה- IVT וה- RNase H המנדם ניתן לזהות כדלקמן: 1) רצועת היעד היא הרצועה החזקה ביותר בג'ל הדף המנוגד. 2) אין או רק להקות חלשות סביב הלהקה הראשית. 3) אין או רק מינים חלשים במשקל מולקולרי גבוה יותר. 4) הכרומטוגרמה של HPLC מציגה שיא מופרד היטב של הרנ"א היעד. 5) כאשר נדגם הפסגה הראשית, רק רצועה אחת מופיעה על ג'ל PAGE denaturing.
תוצאות שליליות במחשוף ה- IVT וה- RNase H הנדכם נוכחים כדלקמן: 1) לא או רק רצועה ראשית חלשה נראית על ג'ל PAGE denaturing. 2) ניתן לראות דפוס של מינים במשקל מולקולרי גבוה מחזרות טנדם RNA. 3) למרות הלהקה הראשית נוכחת, להקות בעוצמה דומה הם מעל או מתחת ללהקה הראשית בתוך ± 3 nt.
מדגם מקופל היטב ניתן לזהות כדלקמן: 1) מספר פרוטונים imino שנצפו תואם את מספר פרוטונים imino הצפוי מסימולציה מבנה משנית (למשל. 2) זוג הבסיסים של Syn G-U בלולאת UUCG (אם קיים) נראה ב- ~ 9.5 ppm, לפעמים נראה רק בטמפרטורה נמוכה יותר. טביעת אצבע נוספת של לולאת UUCG תוארה על ידי Fürtig ועמיתיו40. 3) טביעת האצבע הארומטית מסכימה עם מדגם שהוקצה בעבר שאושר להתקפל כראוי (איור 4C).
מדגם שגוי או מושפל ניתן לזהות כדלקמן: 1) ישנם אותות imino יותר מאשר סימולציית מבנה משני מנבא (הערה: פחות אותות imino לא בהכרח מרמזים על misfolding, כמו זוגות בסיס סגירה לעתים קרובות אינם גלויים, חילופי קונפורמציה מרחיב קווים). 2) היעדר אותות אימונו. 3) אותות צרים בעוצמה גבוהה באזור הארומטי, המצביעים על מוצרי השפלה נוקלאוטיד יחיד. 4) סטייה בין אותות אימונו או ארומטיים לדגימת התייחסות לקיפול מאושר(איור 4C).
ניתן לזהות אטום שאינו מראה החלפה בציר הזמן הניתן לזיהוי כדלקמן: 1) מפרופיל מו"פ שטוח (בשל תרומת REX החסרה המשתנה עם כוח ה- SL החל) (איור 5B ואיור 5F). 2) יש לנקוט זהירות במקרה של חילופי ביניים איטיים כאשר kEX ו Δω הם באותו סדר גודל. במקרה זה, תרומת התהודה יכולה להיות קטנה מאוד כפי שניתן לראות באיור 5C (במקרה זה הפרמטרים המותאמים הם kEX = 292 ± 40 הרץ ו- Δω = 112 ± 4 הרץ). אם יש ספק, ניתן להקליט עבור אימות עקומת SL נמוכה מחוץ לתהודה.
ניתן לזהות אטום המציג חילופים בסולם הזמן הבינוני 1) מפרופיל פיזור הרפיה לא שטוח בניסוי מו"פ על התהודה (איור 5B ואיור 5F); 2) קו רחב יותר בניסוי HSQC או SELOPE יכול גם להיות אינדיקטור להחלפה.
לערכי צריכת החשמל שנבחרו היטב עבור עקומות מחוץ לתהודה(איור 5E,F): 1) יש תרומה משמעותית של kEX בעקומת התהודה (ערכי צריכת SL נבחרים מסומנים באיור 5C ובאיור 5G). 2) כאשר עקומות מחוץ לתהודה נמדדות עבור לפחות 3 ערכי הספק SL, ערכי צריכת החשמל הנבחרים של SL צריכים להתפרס על פני האזור של עקומת התהודה עם תרומת kEX. 3) להוביל R 2+REX לא שטוח עקומות לאחר Fit Laguerre (למשל, איור 5D:עוצמות SL 25, 50 ו 75 הרץ; איור 5E).
ערכי צריכת חשמל SL שנבחרו בצורה גרועה עבור עקומות מחוץ לתהודה(איור 5E,F) מובילים לעקומות Rשטוחות 2+REX לאחר התאמת Laguerre. דוגמה לכך מוצגת באיור 5E, שבו עקומת 100 הרץ מחוץ לתהודה שטוחה מאוד ולכן אינה מספקת מידע משמעותי על Δω.
אינדיקציות עבור רוטציה מסגרת גרעינית Overhauser אפקט (ROE) חפצים: 1) Δω המתקבל עקומות תהודה כבויה להתאים משמרות כימיות של פרוטונים בסביבה מרחבית / פרוטונים, אשר מראים פסגה צולבת עם שיא העניין בספקטרום אפקט האוברהאוזר הגרעיני (NOESY). (לדוגמה, איור 5I מציג עקומות רחבות מחוץ לתהודה כצפוי להחלפה מהירה-בינונית, אך לעקומות יש גם תכונות חדות יותר, למשל., ב- -3000 הרץ ו- +1500 הרץ. אלה הם סביר מאוד בשל חפץ ROE ולא שינוי כימי עבור H8 זה בקונפורמייר אחר). 2) Fit Laguerre אכן עובד, אך אינו פועל היטב (נותן שגיאות גבוהות או ערכים בלתי אפשריים פיזית) עבור תהודה ולפחות 3 עקומות מחוץ לתהודה, למרות שמעריכים הושגו מניסויים עם SINO גבוה (>20) (למשל, kEX = 43,502 ± 38,478 הרץ). לעתים קרובות כל SL מתאים בנפרד, אבל התאמתם יחד נותן שגיאה גבוהה בהרבה; ההתנהגות ההפוכה צפויה למצב נרגש אמיתי.
אינדיקציות להחלפה "אמיתית" Δω: 1) Δω המתקבל מעקומות מחוץ לתהודה אינן תואמות משמרות כימיות של פרוטונים בסביבה מרחבית / פרוטונים, המציגים שיא צולב עם שיא העניין בספקטרום NOESY (למשל. 2) Fit Laguerre נותן שגיאות נמוכות עבור תהודה ולפחות 3 עקומות מחוץ לתהודה (למשל. איור 5I, ראו כיתוב לקבלת תוצאות מתאימות).

איור 3: ייצור לדוגמה על ידי T7 טנדם IVT ו RNase H תגובת מחשוף. (A)דף denaturing של תוצאות חיוביות ושליליות של מחשוף IVT ו- RNase H. גובה הסולם מתייחס להפניות לרנ"א, 12* מתייחס למדריך המחשוף הכימרי. נתיב 1: דור מוצלח של RNA יעד 20 nt. קיימים מעט מוצרים קצרים וארוךים יותר. ליין 2א: מחשוף לא שלם של תעתיק טנדם. למרות שטיהור HPLC אפשרי, הרבה חומר יתבזבז. נתיב 2b: מחשוף RNase H מתמשך של ליין 2 מייצר מדגם נקי מוכן להזרקת HPLC (זהה לנתיב 1). ליין 4: מחשוף RNase H לא הצליח ברובו, ולא הופקה רצועת יעד. תעתיק טנדם באורך מלא עדיין גלוי ב 600 nt. ליין 5: להקת יעד הופקה, אבל להקה חזקה -1 נוכחת. למרות שניתן לבצע HPLC, יש צורך בהסרה זהירה של המוצר הצדדי. (B)דוגמה להזרקת HPLC מוצלחת. השיא של 38 דקות מכיל RNA טהור של אורך היעד, בעוד מוצרים ארוכים וקצרים יותר מופרדים היטב מהרנ"א היעד. לוח ב' שונה מ- 21. קיצורים: IVT = תמלול במבחנה; HPLC = כרומטוגרפיה נוזלית בעלת ביצועים גבוהים; nt = נוקלאוטידים; AU = יחידות שרירותיות. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 4: דוגמה לתקרת ראש של RNA לפני (כחול) ואחרי (אדום) שלב הקיפול 2.4 (ראה פרוטוקול) ב- NMR. (A) אזור Imino בספקטרום 1H-1D של RNA 22-mer עם תווית A. אזורים צפויים עבור זהות זוג הבסיס של אותות imino מסומנים באפור להלן. (B) 1H,13ספקטרום C-HSQC של התהודה הארומטית של ה- RNA מפאנל A. המדגם לאחר קיפול (אדום) מציג 4 אותות כצפוי, בעוד המדגם לפני קיפול (כחול) מראה רק 3 אותות. (C)תחזית מק-קיפול של RNA 22-mer כמו סיכת ראש. חמישה אותות imino צפויים ממבנה משני זה, אשר ניתן למצוא בשתי הדגימות בלוח A. (D) מבנה מוצע של homodimer שנוצר על ידי RNA 22-mer, וכתוצאה מכך אותם 5 זוגות בסיס כמו מבנה סיכת הראש. קיצורים: NMR = תהודה מגנטית גרעינית; 1D = חד ממדי; HSQC = מתאם קוונטי יחיד הטרונוקלרי; ppm = חלקים למיליון. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 5: 1H R1ρ RD תוצאות מייצגות עבור שני מבנים שונים המבוססים על סיכת ראש RNA. (A)העמודה השמאלית מציגה תוצאות שהושגו ב- RNA עם זוג בסיס C-G מעל U הבולט, בעוד העמודה הימנית מציגה תוצאות שהושגו במדגם שבו זוג הבסיס הוחלף ל- G-C במקום זאת. (B) ו -( F) מציגים פרופילי פיזור שטוחים כפי שהושגו עבור A4H8 עבור שני המבנים, המצביעים על כך שאין החלפה קונפורמיאלית. (C–E) הצג תהודה, מחוץ לתהודה ונתונים מותאמים שהושגו עבור G6 במבנה (G-C). ההתאמה של Laguerre מובילה לתוצאה הבאה: R1 = 2.87 ± 0.01 הרץ, R2 = 7.76 ± 0.03 הרץ, kEX = 292 ± 40 הרץ, pES = 0.31 ± 0.03 %, Δω = 112 ± 4 הרץ. (G–I) להציג על תהודה, מחוץ תהודה, ונתונים מותאמים שהושגו עבור G6 במבנה (G-C). ההתאמה של Laguerre מובילה לתוצאה הבאה: R1 = 1.93 ± 0.02 הרץ, R2 = 6.71 ± 0.86 הרץ, k EX = 43,502 ± 38,478 הרץ, p ES = 27 ± 16 %, Δω = 203 ± 166 הרץ. נתון זה שונה מ- 20. קיצור: SL = נעילת סיבוב. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
Discussion
הפרוטוקול המוצג כאן הוא סינתזה של מספר פרוטוקולים שפורסמו בעבר בצורה של מאמרי מחקר10,20,21,31. לפיכך, ניתן להחיל מקטעים של הפרוטוקול, בעוד שאחרים יכולים להיות מוחלפים להעדפת הקורא. לדוגמה, ניתן לבצע את מדידות R1ρ על דגימת RNA המיוצרת בכל שיטה, בהתחשב בכך שקיפול והומוגניות של אורך מניחים. יתר על כן, הפרוטוקול אינו מכיל מידע על הקצאת תהודה של רצף RNA - צעד הנדרש לניסויי RD - שכן זה כוסה בהרחבה בספרותהקודמת 19,37,38. תוכניות תיוג חלקיות, מגזריות או ספציפיות לאתר36,41,42,43,44 הן גישות כדי להקל על הקצאת תהודה או להפחית את החפיפה של תהודה כי הם עניין בניסויי מו"פ תוארו באריכות בספרות. שיטה זו מאפשרת שימוש בתיוג אחיד של כל זהות נוקלאוטיד, אשר כבר יכול לפשט את הקצאת התהודה באופן משמעותי.
שיטת ה- IVT המוצגת כאן מתגברת על בעיות ידועות עם רצפים ותיוג, מגדילה את התשואה ומקטינה את העלות ואת זמן העבודה בהשוואה לשיטות אחרות. השימוש ברצף החניכה הנגיפי מפחית את הצורך באופטימיזציה של תגובה, שהיא בעיה ידועה בתחום שיכולה לגזול זמן רב לביצועים ומניבה רק עותקים מעטים של התמליל במקרה של חניכה שאינה G. מחשוף T7 IVT ו- RNase H של תעתיק הטנדם יכול להתבצע בו זמנית באותו כלי. דפוס של חזרות דו-מושביות רב-מרתיות ניתן לראות על ג'ל PAGE מתמתק במהלך התגובה, המתמזג לפס אחד על הרנ"א היעד עם השלמת תגובת RNase H(איור 3A,נתיבים 1 ו- 2b). תשואות אופייניות בשיטה זו נעות בין RNA של 30 ל- 70 ננומטר לכל 1 מ"ל IVT. עם זאת, השיטה המבוססת על מחשוף RNase H של חזרות טנדם אינה באה ללא בעיות מסוימות משלה. תגובת מחשוף RNase H לעתים קרובות אינה יורדת להשלמה כאשר היא פועלת בו זמנית עם תמלול T7(איור 3A,נתיב 2a).
ניתן להשלים את ההפרדה בין יחידות טנדם על ידי חישול מדריך המחשוף לתעתיק והוספת RNase H(איור 3A,נתיב 2b, שלב 2.1.2). כמו חימום של כמויות גדולות הוא איטי ומוביל Mg2 +- מזורז הידרוליזה של RNA, מיקרוגל קונבנציונאלי שימש, אשר מחמם את המדגם כדי >95 °C (55 °F) ב 10-15 s. תופעות לוואי על הדגימות המיוצרות לא נצפו עד כה. מבנים מסוימים מראים רצועה שנייה קטנה שלא ניתן היה לבטלה על ידי אופטימיזציה של תנאי התגובה(איור 3A, נתיב 4). בדרך כלל אלה נראים בבירור ככתף בכרומטוגרמה של HPLC, אם נעשה שימוש בשיפוע אלוטציה ממוטב היטב, וניתן להסיר אותם (שלב 2.2.5). הדיון הבא נועד להדגיש צעדים קריטיים בפרוטוקול, במיוחד בכל הנוגע להשגת נתונים באיכות גבוהה המאפשרים פרשנות של דינמיקה קונפורמיאלית.
זיהום RNase
RNases חוץ תאיים נמצאים בכל מקום, יציב מאוד, ומהווים את האיום הגדול ביותר ליציבות ארוכת טווח של דגימות NMR. לכן, זה חיוני לעבוד בסביבה ללא RNase ולשמור על כל ריאגנטים ותוכנות פלסטיק RNase ללא. מומלץ להשתמש בטיפים לסינון ואולי אפילו במסות פנים. הדבר חשוב במיוחד לאחר טיהור HPLC. דגימות NMR מזוהמות עם RNases בדרך כלל להציג פסגות צרות גלויות ספקטרום 1H-1D לאחר ימים או שבועות עקב מוצרי השפלה נוקלאוטיד יחיד. מדגם כזה אינו מתאים למדידות R1ρ.
דגימת NMR
בשל אופיו הטעון מאוד, ניתן להשתמש ב- RNA בריכוזים גבוהים ללא משקעים בהשוואה לרוב החלבונים. השימוש בצינורות שיגמי NMR (ראה טבלת החומרים) הוא יתרון שכן הם מאפשרים ריכוז המדגם המרוכז ביותר במרכז סליל תוך מתן תנאי שיימינג ונעילה אידיאליים בשל תחתית הזכוכית הבוכנה תואמת הרגישות ואת הבוכנה. בדרך זו, B1-inhomogeneity מצטמצם, מה שמוליד קווים צרים יותר. נפח המדגם הטיפוסי בצינור NMR הוא 250 μL, וריכוז טיפוסי הוא 1-2 mM. דגימות מתחת 500 מיקרומטר אינן מומלצות לניסויי מו"פ מכיוון שהניסוי ייקח יותר מדי זמן ו- shim טוב. באופן דומה, נפח מדגם מתחת 200 μL אינו מומלץ כי שיים טוב ויציבות שדה (מנעול) נדרש. בעת החדרת הבוכנה, חשוב להימנע מהיווצרות בועות במדגם (שלב 2.4.5). אם לא קבוע כראוי, הבוכנה יכולה להחליק למטה לתוך המדגם, הפחתת עוצמת הקול הניתן לזיהוי. יתר על כן, שינויים מהירים בטמפרטורה יכולים להוביל להיווצרות של בועות חדשות במדגם. לכן, יש לנקוט זהירות בעת הובלת המדגם וכאשר משנים את טמפרטורת הבדיקה בספקטרומטר NMR. בדוק את המדגם עבור בועות בעת מדידה שוב לאחר תקופה ארוכה יותר.
קיפול RNA
מולקולות RNA דינמיות יכולות להתקיים בקונפורציות מרובות כאשר הן אינן מקופלות כראוי. למרות שטמפרטורות ההיתוך של מבנים משניים יכולות להיות רק מעט מעל טמפרטורת החדר, מומלץ הליך חימום וקירור יסודי לפני המדידה. דגימות סיכת ראש מרוכזות במיוחד המתקפלות תחת שליטה קינטית (חימום וקירור הצמדה) יכולות ליצור הומודימרים לאורך זמן, מה שמחייב שליטה קפדנית בקיפול RNA לפני כל מדידת NMR. אם הרנ"א הנמדד אינו מבנה סיכת ראש אלא דופלקס RNA, יש להחיל קיפול איטי תחת שליטה תרמודינמית.
במקרה זה, תהליך הקירור לאחר החימום צריך להיות בטווח של שעות, בעוד RNA משמש בנפח הסופי שלה וריכוז במדגם NMR. ספירה ראשונית של תהודה אימונית וארומטית צפויה יכולה לספק תובנה על ההומוגניות של המדגם. אם המדגם לא נראה כמו צפוי, זה צריך להיות מקופל מחדש. מ"ג2+ (נוסף כמלח כלוריד) יכול לעזור עם קיפול מבני RNA45. בפועל, בקרת הקיפול משמשת כהשוואה לדגימה ששימשה לפחות באופן חלקי להקצאת תהודה של NMR ולפתור את המבנה המשני באופן ניסיוני.
ספין נעילת שיקולי חשמל וחימום
במקרה של הפעלת ניסויי 1H R1ρ RD כניסויים סקירה 2D, כוח SL צריך להיות לא פחות מ 1.2 kHz. תדר משדר גלי הרדיו צריך להיות ממוקם באמצע אזור ppm של פסגות עניין (למשל .,7.5 ppm עבור פרוטונים ארומטיים). רוחב הפס של 1.2 קילו-הרץ יהיה גדול מספיק כדי לנעול פרוטונים אלה ללא אפקטים גדולים מחוץ לתהודה. אפקטים כאלה ניתן לזהות בפרופיל RD. אם הם מתרחשים, ערכי R2+REX גדלים במקום להקטין עם הגדלת ערכי צריכת החשמל של SL, במיוחד עבור צריכת SL נמוכה. בדוק אם ערכי צריכת החשמל המחושב של SL תואמים לצרימת החשמל המועברת לדגימה. בפועל, כוח SL מחושב יכול לשמש אם הדופק הקשה 1H 90° היה מכויל בקפידה על ספקטרומטרים חדשים יותר; עם זאת, ניתן לבדוק זאת על-ידי כיול עוצמת SL עבור כל רוחב פס רצוי.
טווח ההספק של SL, אשר ניתן להשתמש בניסויי RD H R1ρ הוא רחב מאוד, המוביל לחימום מדגם משתנה (1.2 kHz עד 15 kHz עבור HSQC עבור רצפים מבוססי HCP ו 50 הרץ עד 15 kHz לניסויי SELOPE). חימום מדגם לא שוויוני ניתן לזהות כשינוי קל בשינוי כימי בעת השוואת 1Ds המתקבל עבור SLs בהספק נמוך לעומת. אס.אל. השפעה זו בדרך כלל אינה נחשבת פיצויי חום בניסויי R1ρ על heteronuclei. פיצוי חום בניסויים אלה מוגדר בדרך כלל כדי לתקן עבור חימום שונה בשל משכי נעילת ספין שונים שצוינו ברשימת vd של כל סדרת כוח נעילת ספין. במיוחד עבור ניסוי SELOPE, פיצוי חום שני צריך לשמש על פני כל נקודות החוזק SL החלות כפי שתואר ב20.
שיקולי רשימת vd
כפי שהוזכר קודם לכן, רשימת VD צריך להכיל נקודת זמן מספיק זמן כדי להשיג ריקבון משמעותי של אינטנסיביות (באופן אידיאלי עד 30% של האות הראשוני, או נמוך ככל האפשר אם לא ניתן להגיע 70% ריקבון בתוך המפרט של הבדיקה). למרות שרשימת VD הייתה ממוטבת עבור הספק SL נמוך (1.2 kHz), רשימת vd זו צריכה להיבדק גם בהספק SL הגבוה ביותר לשימוש (למשל., 15 kHz). זאת בשל העובדה, כי עבור פסגות עם תרומת REX משמעותית, הריקבון יהיה הרבה יותר איטי בעוצמה SL גבוהה. אז ריקבון מספיק צריך להיות מאומת גם בעוצמה SL גבוהה. אותו הדבר יש לשקול עבור ריקבון ב קיזוזים גבוהים בניסויים מחוץ תהודה. נקודת הזמן המרבית האידיאלית של רשימת VD יכולה להיות שונה באופן משמעותי עבור האזורים השונים של ניסוי הפיזור. במקרה זה, ניתן לכלול יותר נקודות ברשימת VD, ונקודות רשימת VD הארוכות יותר עבור כוח SL גבוה יותר או היכודים גבוהים יותר במהלך הניתוח, בהתבסס על SINO נמוך הם יובילו, ניתן לבטל. באופן כללי, יש לשקול 5-8 נקודות רשימה vd להיות מסוגל לזהות חפצים פוטנציאליים המובילים ריקבון לא מעריכי כגון J-צימוד (ראה להלן).
1D-שיקולי סלקטיביות של HCP
יש לנקוט זהירות מיוחדת בעת הפעלת גירסת 1D מבוססת HCP אם יש שיא נוסף החופף לשיא העניין בממד 1H של הניסוי מבוסס HSQC 2D. העברות מבוססות HCP הן מאוד, אך אף פעם לא סלקטיביות ב-100%, ולכן יכול לקרות ששיא נוסף תורם לאינטנסיביות ולריקבון של שיא העניין ב-1D. אינדיקציה לכך תהיה הבדל בערכי R 1ρ בתהודה המתקבלים באמצעות גרסאות 1D ו- 2D של הניסוי שכותרתו.
שיקולי ROE:
עבור עקומות מחוץ לתהודה של אטומים עם חילופי ביניים איטיים, ניתן לזהות חפצי ROE בהתבסס על השוואה של Δω המתקבל עם ספקטרום NOESY או ROESY. אם ניתן לזהות פסגת חוצה בהפרש תזוזה כימי המתאים ל- Δω, אז המצב הנרגש הנצפה עשוי למעשה להיות חפץ ROE (למשל. מניסיון, זה תמיד הוביל גם להתקפים גרועים עם שגיאות גדולות, אולי בגלל ROE לא בעקבות אותה תבנית כמו REX עם הגדלת כוח SL. המצב הופך להיות קשה יותר עבור חילופי ביניים מהירים. בעוד עקומת התהודה היא (בהשוואה לנתוני 13C המתקבלים בגרעין השכן) עדיין מייצגת את תהליך החליפין בין GS ו- ES, עקומת התהודה מושפעת מחפצי ROE מרובים.
במקרה זה, כוח SL כדי לזהות את תהליך החליפין הוא גדול יותר (>1.5 kHz) ולכן משתרע על פני מספר גדול יותר של פרוטונים כמו עקומות מחוץ תהודה משתרע על הבדלי משמרות כימיות של מועמדים ROE שונים (עבור H8 אלה יהיו: פרוטוני אמינו ב ca. ±1000 הרץ, H5 / H1 של ב ca. -1200 הרץ, פרוטונים imino ב ca. 3500 הרץ). עד כה, לא נמצאה שיטה לדיכוי חפצי ROE אלה (מלבד שימוש בנוקלאוטידים מפורקים חלקית46), ולא ניתן לכלול נתוני תהודה להחלפה מהירה, שכן לא ניתן לחלץ מידע אמין על Δω בפועל בשיטה זו, אם לא ניתן לשלול תרומת NOE /ROE באמצעות ספקטרום NOESY.
J-צימוד (הרטמן-האן) שיקולים
למרות עקומות על תהודה עבור פרוטונים J-מצמיד הומונוקלאר, כגון H6, נרשמו בהצלחה10,20, יש לנקוט טיפול מיוחד עבור מדידות מחוץ תהודה, במיוחד עבור כוח SL נמוך כמו התנאים התואמים הרטמן-האן יכול להתפרש על מגוון רחב של קיזוזים נחקרים. ניתן לזהות את חפצי הרטמן-האן כתנודות על הריקבון המעריכי או להגדיל את ערכי R2+REX עם עוצמות SL הולכות וגדלות בחלקות מו"פ על התהודה20.
Disclosures
K.P. הוא יועץ Arrakis Therapeutics, חברה המגלה מולקולות קטנות מיקוד RNA.
Acknowledgements
אנו מודים למתקן למדע החלבון (PSF) במכון קרולינסקה על ביטוי וטיהור של T7 RNA פולימראז ו- E. coli RNase H, מרטין האלברג על המתנה הנדיבה של הפוספטאז האנאורגני, ועל פטזולאב כולו לדיונים חשובים. אנו מודים ללוקה רטינו על הכנת מבני U-בליטה ואמילי שטיינר וקרוליינה פונטנה על תרומתם לפקודות מאקרו ותסריטים מתאימים. אנו מכירים במכון קרולינסקה ובחטיבה לביוכימיה רפואית וביופיזיקה לתמיכה ברכישת ספקטרומטר 600 מגה-הרץ ומימון פוזיציות (KI FoAss ו-KID 2-3707/2013). אנו אסירי תודה על התרומה הכספית של Vetenskapsrådet (#2014-4303), Stiftelsen för strategisk Forskning (ICA14-0023 ו- FFL15-0178) ורגנאר סודרברג סטיפלסה (M91-14), הארלד אוק גרטה ג'ינסון סטיפלסה ((באנגלית: Ragnar Söderberg Stiftelse) JS20140009), קרל טריגרס סטיפלסה (CTS14-383 ו-15-383), אווה אוק אוסקר ארנס סטיפלסה, אקה ויברג סטיפלסה (467080968 ו-M14-0109), Cancerfonden (CAN 2015/388), J.S. מכיר במימון באמצעות מארי Skłodowska-Curie אם (האיחוד האירופי H2020, פרויקט מס' 747446 של MSCA-IF).
Materials
| Name | Company | Catalog Number | Comments |
| 40% Acrylamide/Bis Solution | Bio-Rad | 161-0144 | |
| 5-alpha Competent E. coli | NEB | C2987I | |
| Acetic Acid | Sigma-Aldrich | 49199 | |
| Acetonitrile | Sigma-Aldrich | 34851 | |
| AFC-3000, HPLC Fraction collector | Thermo Scientific | 5702.1 | |
| Agarose | Sigma-Aldrich | A9414 | |
| Amersham ImageQuant 800 UV | GE Healthcare | 29399482 | Replacing LAS-4000 or equivalent |
| Amicon ultra centrifugal filter unit | Sigma-Aldrich | UFC900324 | |
| Ammonium persulfate | Sigma-Aldrich | A3678 | |
| Ampicillin | Sigma-Aldrich | A9518 | |
| ATP | Sigma-Aldrich | A2383 | |
| ATP-13C10/15N5 | Sigma-Aldrich | 645702 | |
| BamHI restriction enzyme | NEB | R0136L | |
| Bottle top filter | VWR | 514-1019 | |
| Bromophenol Blue | Sigma-Aldrich | 1081220005 | |
| Cleavage guide | IDT | N/A | or equivalent |
| CTP | Sigma-Aldrich | C1506 | |
| CTP-13C10/15N5 | Sigma-Aldrich | 645699 | |
| D2O | Sigma-Aldrich | 151882 | |
| Dionex Ultimate 3000 UHPLC system | Thermo Scientific | N/A | |
| DL-Dithiotreitol | Sigma-Aldrich | 43815 | |
| DMSO | Sigma-Aldrich | D8418 | |
| DNAPac PA200 22x250 Semi-Prep column | Thermo Scientific | SP6734 | |
| DNAPac PA200 22x50 guard column | Thermo Scientific | SP6731 | |
| E.coli RNase H | NEB | M0297L | or made in-house uniprot ref. P0A7Y4 |
| EDTA | Sigma-Aldrich | E6758 | |
| Eppendorf centrifuge, rotor: A-4-44 | Eppendorf | 5804R | |
| Ethanol 95% | Fisher scientific | 11574139 | |
| Ethanol 95% denatured | VWR | 85829.29 | |
| Formamide | Sigma-Aldrich | 47671 | |
| GelRed | VWR | 41003 | |
| GeneRuler 1kbp Plus | Fisher Scientific | SM1333 | Optional |
| GMP | Sigma-Aldrich | G8377 | |
| GMP-13C10/15N5 | Sigma-Aldrich | 650684 | |
| GTP | Sigma-Aldrich | G8877 | |
| GTP-13C10/15N5 | Sigma-Aldrich | 645680 | |
| Hydrochloric Acid | Sigma-Aldrich | H1758 | |
| Inorganic pyrophosphatase | Sigma-Aldrich | I1643-100UN | or made in-house uniprot ref. P0A7A9 |
| Invitrogen UltraPure 10X TBE-buffer | Sigma-Aldrich | T4415 | |
| Julabo TW8 Water bath | VWR | 461-3117 | |
| kuroGEL Midi 13 Horizontal gel electrophoresis | VWR | 700-0056 | or comparable |
| LB broth (Lennox) | Sigma-Aldrich | L3022 | |
| LB broth with agar (Lennox) | Sigma-Aldrich | L2897 | |
| Low Range ssRNA Ladder | NEB | N0364S | Optional |
| LPG-3400RS Pump | Thermo Scientific | 5040.0036 | |
| Magnesium chloride hexahydrate | Sigma-Aldrich | 63068 | |
| microRNA Marker | NEB | N2102S | |
| Microwave oven | Samsung | MS23F301EAW | |
| Mini-PROTEAN electrophoresis equipment | Bio-Rad | 1658004 | |
| NucleoBond Xtra Maxi | Machinery-Nagel | 740414.10M | |
| pUC19 plasmid containing tandem insert | Genscript | N/A | or equivalent |
| RNaseZAP | Sigma-Aldrich | R2020 | |
| Shigemi tube 5mm | Sigma-Aldrich | Z529427 | |
| Single-use syringe, Luer lock tip | VWR | 613-2008 | |
| Sodium acetate | Sigma-Aldrich | S2889 | |
| Sodium chloride | Sigma-Aldrich | 730-1470 | |
| Sodium perchlorate | Sigma-Aldrich | 71853 | |
| Sodium phosphate dibasic | Sigma-Aldrich | S3264 | |
| Sodium phosphate monobasic | Sigma-Aldrich | S3139 | |
| Spermidine trihydrochloride | Sigma-Aldrich | 85578 | |
| SYBR Gold | ThermoFisher | S11494 | |
| Syringe filters | VWR | 514-0061 | |
| T7 RNA polymerase | Sigma-Aldrich | 10881767001 | or made in-house uniprot ref. P00573 |
| TCC-3000RS Column thermostat | Thermo Scientific | 5730 | |
| Tetramethylethylenediamine | Sigma-Aldrich | T9281 | |
| Tris Base | Fisher Scientific | 10103203 | |
| UMP | Sigma-Aldrich | U6375 | |
| UMP-13C9/15N2 | Sigma-Aldrich | 651370 | |
| Urea | Sigma-Aldrich | U5378 | |
| UTP | Sigma-Aldrich | U6625 | |
| UTP-13C10/15N5 | Sigma-Aldrich | 645672 | |
| VWD-3100 Detector | Thermo Scientific | 5074.0005 |
References
- Djebali, S., et al. Landscape of transcription in human cells. Nature. 489 (7414), 101-108 (2012).
- Doudna, J. A., Cech, T. R. The chemical repertoire of natural ribozymes. Nature. 418 (6894), 222-228 (2002).
- Sehgal, P. B., Westley, J., Lerea, K. M., DiSenso-Browne, S., Etlinger, J. D. Biomolecular condensates in cell biology and virology: phase-separated membraneless organelles (MLOs). Analytical Biochemistry. , 597 (2020).
- Herschlag, D., Allred, B. E., Gowrishankar, S. From static to dynamic: the need for structural ensembles and a predictive model of RNA folding and function. Current Opinion Structural Biology. 30, 125-133 (2015).
- Kimsey, I. J., Petzold, K., Sathyamoorthy, B., Stein, Z. W., Al-Hashimi, H. M. Visualizing transient Watson-Crick-like mispairs in DNA and RNA duplexes. Nature. 519 (7543), 315-320 (2015).
- Dethoff, E. A., Petzold, K., Chugh, J., Casiano-Negroni, A., Al-Hashimi, H. M. Visualizing transient low-populated structures of RNA. Nature. 491 (7426), 724-728 (2012).
- Baisden, J. T., Boyer, J. A., Zhao, B., Hammond, S. M., Zhang, Q. Visualizing a protonated RNA state that modulates microRNA-21 maturation. Nature Chemical Biology. 17 (1), 80-88 (2021).
- Marušič, M., Schlagnitweit, J., Petzold, K. RNA dynamics by NMR spectroscopy. Chembiochem. 20 (21), 2685-2710 (2019).
- Baronti, L., et al. Base-pair conformational switch modulates miR-34a targeting of Sirt1 mRNA. Nature. 583 (7814), 139-144 (2020).
- Steiner, E., Schlagnitweit, J., Lundström, P., Petzold, K. Capturing excited states in the fast-intermediate exchange limit in biological systems using 1H spectroscopy. Angewandte Chemie International Edition. 55 (51), 15869-15872 (2016).
- Moschen, T., et al. Ligand-detected relaxation dispersion NMR spectroscopy: dynamics of preQ1-RNA binding. Angewandte Chemie International Edition. 54 (2), 560-563 (2015).
- LeBlanc, R. M., Longhini, A. P., Tugarinov, V., Dayie, T. K. NMR probing of invisible excited states using selectively labeled RNAs. Journal of Biomolecular NMR. 71 (3), 165-172 (2018).
- Strebitzer, E., Nußbaumer, F., Kremser, J., Tollinger, M., Kreutz, C. Studying sparsely populated conformational states in RNA combining chemical synthesis and solution NMR spectroscopy. Methods. 1148, 39-47 (2018).
- Rangadurai, A., Shi, H., Al-Hashimi, H. M. Extending the sensitivity of CEST NMR spectroscopy to micro-to-millisecond dynamics in nucleic acids using high-power radio-frequency fields. Angewandte Chemie International Edition. 59 (28), 11262-11266 (2020).
- Hansen, D. F., Vallurupalli, P., Kay, L. E. Using relaxation dispersion NMR spectroscopy to determine structures of excited, invisible protein states. Journal of Biomolecular NMR. 41 (3), 113-120 (2008).
- Lundström, P., Akke, M. Off-resonance rotating-frame amide proton spin relaxation experiments measuring microsecond chemical exchange in proteins. Journal of Biomolecular NMR. 32 (2), 163-173 (2005).
- Lee, J., Dethoff, E. A., Al-Hashimi, H. M. Invisible RNA state dynamically couples distant motifs. Proceedings of the National Academy of Sciences of the United States of America. 111 (26), 9485-9490 (2014).
- Schnieders, R., Keyhani, S., Schwalbe, H., Fürtig, B. More than proton detection- new avenues for NMR spectroscopy of RNA. Chemistry. 26 (1), 102-113 (2020).
- Fürtig, B., Richter, C., Wöhnert, J., Schwalbe, H. NMR spectroscopy of RNA. Chembiochem. 4 (10), 936-962 (2003).
- Schlagnitweit, J., Steiner, E., Karlsson, H., Petzold, K. Efficient detection of structure and dynamics in unlabeled RNAs: The SELOPE approach. Chemistry. 24 (23), 6067-6070 (2018).
- Feyrer, H., Munteanu, R., Baronti, L., Petzold, K. One-pot production of RNA in high yield and purity through cleaving tandem transcripts. Molecules. 25 (5), 1142 (2020).
- Baronti, L., Karlsson, H., Marušič, M., Petzold, K. A guide to large-scale RNA sample preparation. Analytical and Bioanalytical Chemistry. 410 (14), 3239-3252 (2018).
- Brunelle, J. L., Green, R. In vitro transcription from plasmid or PCR-amplified DNA. Methods in Enzymology. 530, 101-114 (2013).
- Borkotoky, S., Murali, A. The highly efficient T7 RNA polymerase: A wonder macromolecule in biological realm. International Journal of Biological Macromolecules. 118, 49-56 (2018).
- Arnaud-Barbe, N., Cheynet-Sauvion, V., Oriol, G., Mandrand, B., Mallet, F. Transcription of RNA templates by T7 RNA polymerase. Nucleic Acids Research. 26 (15), 3550-3554 (1998).
- Guillerez, J., Lopez, P. J., Proux, F., Launay, H., Dreyfus, M. A mutation in T7 RNA polymerase that facilitates promoter clearance. Proceedings of the National Academy of Sciences of the United States of America. 102 (17), 5958-5963 (2005).
- Kuzmine, I., Gottlieb, P. A., Martin, C. T. Binding of the priming nucleotide in the initiation of transcription by T7 RNA polymerase. Journal of Biological Chemistry. 278 (5), 2819-2823 (2003).
- Gholamalipour, Y., Karunanayake Mudiyanselage, A., Martin, C. T. 3' end additions by T7 RNA polymerase are RNA self-templated, distributive and diverse in character - RNA-Seq analyses. Nucleic Acids Research. 46 (18), 9253-9263 (2018).
- Inoue, H., Hayase, Y., Iwai, S., Ohtsuka, E. Sequence-dependent hydrolysis of RNA using modified oligonucleotide splints and RNase H. FEBS Letters. 215 (2), 327-330 (1987).
- Wang, X., Li, C., Gao, X., Wang, J., Liang, X. Preparation of small RNAs using rolling circle transcription and site-specific RNA disconnection. Molecular Therapy - Nucleic Acids. 4, 215 (2015).
- Karlsson, H., Baronti, L., Petzold, K. A robust and versatile method for production and purification of large-scale RNA samples for structural biology. RNA. 26 (8), 1023-1037 (2020).
- Hartmann, S. R., Hahn, E. L. Nuclear double resonance in the rotating frame. Physical Review. 128 (5), 2042-2053 (1962).
- Chiarparin, E., Pelupessy, I., Bodenhausen, G. Selective cross-polarization in solution state NMR. Molecular Physics. 95 (5), 759-767 (1998).
- Korzhnev, D. M., Orekhov, V. Y., Kay, L. E. Off-resonance R 1ρ NMR studies of exchange dynamics in proteins with low spin-lock fields: an application to a Fyn SH3 domain. Journal of the American Chemical Society. 127 (2), 713-721 (2005).
- Hansen, A. L., Nikolova, E. N., Casiano-Negroni, A., Al-Hashimi, H. M. Extending the range of microsecond-to-millisecond chemical exchange detected in labeled and unlabeled nucleic acids by selective carbon R 1ρ NMR spectroscopy. Journal of the American Chemical Society. 131 (11), 3818-3819 (2009).
- Duss, O., Maris, C., von Schroetter, C., Allain, F. H. -. T. A fast, efficient and sequence-independent method for flexible multiple segmental isotope labeling of RNA using ribozyme and RNase H cleavage. Nucleic Acids Research. 38 (20), 188 (2010).
- Krähenbühl, B., Lukavsky, P., Wider, G. Strategy for automated NMR resonance assignment of RNA: application to 48-nucleotide K10. Journal of Biomolecular NMR. 59 (4), 231-240 (2014).
- LeBlanc, R. M., Longhini, A. P., Le Grice, S. F. J., Johnson, B. A., Dayie, T. K. Combining asymmetric 13C-labeling and isotopic filter/edit NOESY: a novel strategy for rapid and logical RNA resonance assignment. Nucleic Acids Research. 45 (16), 146 (2017).
- Parisien, M., Major, F. The MC-Fold and MC-Sym pipeline infers RNA structure from sequence data. Nature. 452 (7183), 51-55 (2008).
- Fürtig, B., Richter, C., Bermel, W., Schwalbe, H. New NMR experiments for RNA nucleobase resonance assignment and chemical shift analysis of an RNA UUCG tetraloop. Journal of Biomolecular NMR. 28 (1), 69-79 (2004).
- Keyhani, S., Goldau, T., Blümler, A., Heckel, A., Schwalbe, H. Chemo-enzymatic synthesis of position-specifically modified RNA for biophysical studies including light control and NMR spectroscopy. Angewandte Chemie International Edition. 57 (37), 12017-12021 (2018).
- Marchanka, A., Kreutz, C., Carlomagno, T. Isotope labeling for studying RNA by solid-state NMR spectroscopy. Journal of Biomolecular NMR. 71, 151-164 (2018).
- Becette, O., Olenginski, L. T., Dayie, T. K. Solid-phase chemical synthesis of stable isotope-labeled RNA to aid structure and dynamics studies by NMR spectroscopy. Molecules. 24 (19), 3476 (2019).
- Zhang, X., Li, M., Liu, Y. Optimization and characterization of position-selective labelling of RNA (PLOR) for diverse RNA and DNA sequences. RNA Biology. 17 (7), 1009-1017 (2020).
- Roh, J. H., et al. Effects of preferential counterion interactions on the specificity of RNA folding. The Journal of Physical Chemistry Letters. 9 (19), 5726-5732 (2018).
- Juen, M. A., et al. Excited states of nucleic acids probed by proton relaxation dispersion NMR spectroscopy. Angewandte Chemie German Edition. 55 (39), 12008-12012 (2016).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved