
Method Article
כמות מוחלטת של אינוזיטול פירופוספטים על ידי אלקטרופורזה נימית אלקטרוספריי ספקטרומטריית מסה יינון
In This Article
Summary
מתואר הליך לאלקטרופורזה נימית אלקטרופורזה ספקטרומטריית מסת יינון עבור הכמות המוחלטת של אינוזיטול פירופוספטים מתמציות תאי יונקים.
Abstract
אינוזיטול פירופוספטים (PP-InsPs) הם קבוצה חשובה של מולקולות איתות תוך-תאיות. מולקולות אלה, שמקורן באינוזיטול פוספטים (InsPs), כוללות נוכחות של לפחות פירופוספט מואטי אנרגטי אחד על טבעת מיו-אינוזיטול. הם קיימים בכל מקום באאוקריוטים ופועלים כשליחים מטבוליים הסוקרים הומאוסטזיס פוספט, רגישות לאינסולין ומטען אנרגיה תאית. בשל היעדר כרומופור במטבוליטים אלה, צפיפות מטען גבוהה מאוד ושפע נמוך, הניתוח שלהם דורש מעקב רדיואקטיבי, ולכן הוא מפותל ויקר. כאן, המחקר מציג פרוטוקול מפורט לביצוע כמות תפוקה מוחלטת וגבוהה של אינוזיטול פירופוספטים מתאי יונקים על ידי אלקטרופורזה נימית אלקטרוספריי ספקטרומטריית מסת יינון (CE-ESI-MS). שיטה זו מאפשרת פרופיל רגיש של כל מיני PP-InsPs הרלוונטיים ביולוגית בתאי יונקים, ומאפשרת הפרדה בסיסית של רג'יואיזומרים. מוצגים ריכוזים תאיים מוחלטים של PP-InsPs, כולל איזומרים קלים, וניטור השינויים הזמניים שלהם בתאי HCT116 במספר תנאי ניסוי.
Introduction
מאז הגילוי הראשוני של מיו-אינוזיטול פירופוספטים (PP-InsPs) בשנת 1993 1,2, הושגה התקדמות משמעותית כדי להבהיר את הביוסינתזה, המחזור והתפקודים שלהם3. אינוזיטול פירופוספטים מופיעים בכל מקום בתאים אאוקריוטים4 ומשמשים כמולקולות איתות מטבוליות המעורבות באופן קריטי, למשל, הומאוסטזיס פוספט5,6, רגישות לאינסולין7, תנודות סידן8,9, סחר בשלפוחית 10, אפופטוזיס 11, תיקון DNA 12, איתות חיסוני 13 ואחרים., שפע התהליכים החשובים תחת שליטתם של אינוזיטול פירופוספטים דורש הבנה עמוקה יותר של השפע התאי שלהם, תנודות ולוקליזציה.
למרות ש- InsPs ו- PP-InsPs משכו תשומת לב בין דיסציפלינות, ניתוח השפע שלהם מבוצע באופן שגרתי בשיטה שפותחה במהלך שנות ה -80, המורכבת מתיוג תאים עם אינוזיטול טריטי, פתרון PP-InsPs שחולץ על ידי כרומטוגרפיית חילופי אניונים חזקה Sax-HPLC עם ספירת scintillation שלאחר מכן. שיטות חדשות יותר המבוססות על ספקטרומטריית מסות עדיין מתמודדות עם אתגרים משמעותיים: אינוזיטול פירופוספטים עם עד שמונה יחידות פוספט מכילים אסטרי פוספט ואנהידרידים, מה שמוביל למטען שלילי משמעותי ולאובדן פוספט פוטנציאלי במהלך יינון. ישנם ארבעה סוגים עיקריים של PP-InsPs שנמצאים ביונקים (איור 1): 1,5-(PP)2-InsP 4 (או 1,5-InsP8), 5-PP-InsP 5 (או 5-InsP 7), 1-PP-InsP 5 (או 1-InsP7) ו-5-PP-Ins(1,3,4,6)P 4 (או5-PP-InsP 4)3,14. הרמות הפיזיולוגיות של PP-InsPs נמצאות בדרך כלל בטווח הננו-מיקרומולרי הנמוך, כאשר 5-PP-InsP 5 הוא הנפוץ ביותר עם ריכוזים תאיים של 0.5 - 5 מיקרומטר. 1,5-(PP)2-InsP 4 ו-1-PP-InsP 5 הם ככל הנראה עד כ-10% ממאגר 5-PP-InsP 5 ונותר קשה למעקב בתאים רבים15. 5-PP-InsP4 עם קבוצת OH חופשית הוא אפילו נמוך יותר בשפע ובדרך כלל הופך להיות ניתן לזיהוי רק כאשר הידרולזות פוספט מעוכבות עם נתרן פלואוריד (NaF)16.
צפיפות המטען הגבוהה של PP-InsPs מקשה על הפרדתם, והמופע של רג'ואיזומרים PP-InsP מסבך עוד יותר את המאמצים הללו. כתוצאה מכך, רוב הניסויים הסתמכו על כמות על ידי תיוג רדיואקטיבי מטבולי של תאים באמצעות [3H]-אינוזיטול, כאשר הרקע מהמטריצה אינו נכלל ורגישות גבוהה מושגת17,18. עם זאת, שיטה זו היא יקרה, זמן רב, ואינה מאפשרת להבחין כראוי בין regioisomers PP-InsP הקשורים. יתר על כן, [3H]-אינוזיטול תיוג אינו לוקח בחשבון סינתזה אנדוגנית של אינוזיטול מגלוקוז. שיטה מבוססת אלקטרופורזה בג'ל פוליאקרילאמיד (PAGE) היא חלופה זולה המיושמת באופן נרחב אך מוגבלת ברגישותה 19,20,21,22. פורסמו גישות אחרות הנמנעות מתיוג רדיו, כולל כרומטוגרפיית יונים ואחריה נגזרת לאחר עמודה גילוי UV23, כרומטוגרפיית אינטראקציה הידרופילית (HILIC)24, או חילופי אניונים חלשים (WAX) בשילוב עם ספקטרומטריית מסה (MS)25. עם זאת, הם אינם (עדיין) בקנה אחד עם הפרוטוקול הקלאסי [3H]-אינוזיטול SAX-HPLC.
לאחרונה, אלקטרופורזה נימית אלקטרוספריי יינון מסה ספקטרומטריה (CE-ESI-MS) הוצגה כאסטרטגיה טרנספורמטיבית לניתוח של מטבוליזם InsPs ו- PP-InsPs, העונה על כל הדרישות שנדונו לעיל16. בשילוב עם מיצוי InsP המתקדם הנוכחי על ידי חומצה פרכלורית ואחריו העשרה עם חרוזי טיטניום דו-חמצני26, CE-ESI-MS הצליח בכל אורגניזם שנבדק עד כה, משמרים ועד צמחים ויונקים. פרופיל סימולטני של InsPs ו- PP-InsPs, כולל כל הרג'ואיזומרים האפשריים, הושג בקלות. תקנים פנימיים יציבים המסומנים באיזוטופים (SIL) אפשרו כמות אבסולוטית מהירה ומדויקת, ללא קשר להשפעות המטריצה. מאחר שטרשת נפוצה יכולה ללכוד הבדלי מסה איזוטופיים, ניתן ליישם את CE-ESI-MS גם כדי לחקור מסלולי סינתזה תאית ממודרים של InsPs ו-PP-InsPs, למשל על-ידי הזנת תאים עם [13 C 6]-מיו-אינוזיטול או [13C 6]-D-גלוקוז.
המתואר כאן הוא פרוטוקול מפורט שלב אחר שלב עבור הכמות המוחלטת של PP-InsPs ו- InsPs מתאי יונקים על ידי CE-ESI-MS. מלבד האיזומר העיקרי של 5-PP-InsP 5, 1,5-(PP)2-InsP 4 ו-1-PP-InsP5 מכמתים גם הם במחקר זה, למרות השפע הנמוך שלהם. שני קווי תאי HCT116 ממעבדות שונות (NIH, UCL) נחקרים, ומאומת כי תאי HCT116UCL מכילים רמות גבוהות פי 7 של 1,5-(PP)2-InsP 4 מאשר ב- HCT116NIH, בעוד שריכוזי 5-PP-InsP5 דומים. בנוסף, סינתזת 1-PP-InsP5 ב- HCT116UCL אינה מוגברת באופן משמעותי. כמו כן, העלייה ברמות PP-InsP על ידי חסימת הדה-פוספורילציה שלהם באמצעות נתרן פלואוריד נחקרת באופן כמותי.
Protocol
1. הגדרת מערכת CE-ESI-MS
- להקים מערכת CE-ESI-MS המורכבת ממערכת CE מסחרית - וספקטרומטר מסה טנדם משולש מרובע, המצויד במקור יינון אלקטרוספריי (ESI) של זרם סילון אג'ילנט (AJS). נדרשת ערכת ריסוס CE-ESI-MS ומשאבת LC איזוקרטית (כרומטוגרפיה נוזלית).
- חבר את מפצל זרימת הנדן 1:100 (כלול בערכת הריסוס CE-MS) ואת שקע משאבת LC איזוקרטית.
- ודא כי בקבוקון הכניסה של מערכת CE נמצא באותו גובה כמו קצה המרסס של מנתח המסה.
- השתמש בתחנת העבודה MassHunter (גרסה 10.1) או בתוכנת MS דומה כדי לשלוט במערכת כולה, וכן לצורך איסוף וניתוח נתונים.
2. הכנת מאגר, נימי דם ומערכת CE-MS
- הכן חיץ ריצה CE: התאם את ה- pH של אמוניום אצטט של 40 mM ל- 9.0 עם אמוניום הידרוקסיד. מומלץ להשתמש בבקבוק נפחי של 250 מ"ל. סנן את המאגר בגודל 250 מ"ל באמצעות מסנני ממברנה בגודל נקבוביות של 0.2 מיקרומטר. הקפידו להשתמש רק במים טהורים במיוחד שעברו דה-יוניזציה ובריאגנטים ברמה של טרשת נפוצה.
הערה: ניתן לשמור חיץ זה בטמפרטורת החדר למשך 2-3 שבועות או למשך מספר חודשים במקרר. - הכינו את נוזל הנדן: ערבבו 100 מ"ל מים טהורים במיוחד ו-100 מ"ל איזופרופנול בדרגה LC-MS בבקבוק של 250 מ"ל. החלף את נוזל הנדן לפחות פעם בשבוע. הוסף התייחסות מסה לנוזל הנדן בעת שימוש בספקטרומטר מסה ברזולוציה גבוהה.
- התקן נוזל נדן: נקה במהירות של 5 מ"ל/דקה למשך 5 דקות והגדר את קצב הזרימה על 1 מ"ל/דקה (10 μL/min לתוך מרסס CE-MS). לחץ המשאבה יהיה בערך 180 בר. ודא שצינורות המיחזור מתחברים בחזרה לבקבוק נוזל הנדן כדי לעשות שימוש חוזר בממס.
- הכן נימי: רכוש נימי CE-MS (50 μm i.d. ו- 365 μM o.d. באורך של 125 ס"מ) עם חלון זיהוי UV. המשתמש יכול גם להשיג הרבה יותר זול נימי סיליקה התמזגות בר ממפיצים מיוחדים. חותכים נימי באורך של 100 ס"מ. חותכים כראוי את שני הקצוות הנימיים עם חותך עמוד נימי עם להב יהלום מסתובב ומסירים 2-3 ס"מ של ציפוי פולימיד משני הקצוות עם מצית. נקו את המשטח הנימי עם איזופרופנול.
- התקן נימי: התאם את הנימים לקלטת CE-MS. לחץ על כפתור שינוי קלטת והתקן את הקלטת במכשיר CE. קצה הכניסה של הנימים נמוך בכ-2 מ"מ מהאלקטרודה. ודא שקצה הכניסה נמוך משטח הדגימה במהלך תהליך ההזרקה.
- הפעלת נימי: לפני השימוש הראשון, יש לשטוף את הנימים עם 1 M NaOH, לאחר מכן מים למשך 10 דקות, וחיץ ריצה CE למשך 15 דקות.
- הכנס את הקצה הנימי למרסס CE-MS: הכנס בעדינות את הנימי למרסס CE-MS וודא שהקצה הנימי בולט כ-0.1 מ"מ מקצה המרסס. הקפידו לבצע כוונון מדויק של קצה השקע הנימי באמצעות זכוכית מגדלת ובורג הכוונון במרסס. החזירו את המרסס למקור היונים והימנעו מלגעת בבורג הכוונון. ה- MS נמצא במצב המתנה בעת ביצוע פעולה זו.
- בדוק את ספריי ESI: בדוק את היציבות של מרסס ESI במצב סריקה מלאה. התנודתיות של סך האלקטרופרוגרמות של היונים חייבת להיות בטווח של 5%.
- בצע הרצת בדיקה עם תקני InsP: השתמש בתערובת של 2 μM InsP3-InsP 8 תקנים (מותאם על ידי כמות 31P NMR15,27) עבור ריצות בדיקה עם זריקה ב 50 mbar עבור 10 שניות (10 nL). הגדר את הפרמטרים המפורטים של ESI ו- MS כפי שמוצג בטבלה 1. זרם CE הוא בערך 26 μA. רוחב השיא הוא סביב 0.4-0.5 דקות. ודא שיחס האות לרעש מגיע ל-400 לפחות.
3. מיצוי פוספטים מסיסים של אינוזיטול מתאי יונקים
הערה: תאי HCT116NIH היו מתנה חביבה מסטיבן שירס28. תאי HCT116UCL היו ממעבדה26 של סאיארדי.
- זריעת תאים
- תרבית תאי HCT116NIH או HCT116UCL ב-T75 צלוחיות בטמפרטורה של 37 מעלות צלזיוס באטמוספירה של CO2 עם לחות גבוהה ב-5% (המכונה גם תנאים סטנדרטיים) במדיום הנשר המעובד (DMEM) של דולבקו, בתוספת 10% סרום בקר עוברי (FBS).
- יש לשטוף את תרביות המלאי HCT116NIH ו-HCT116UCL עם תמיסות מלח עם מאגרי פוספט (PBS) (5 מ"ל) ולדגור על התאים עם חומצה טטרצטית טריפסין-אתילנדיאמין (EDTA) (3 מ"ל, 0.25%) בתנאים סטנדרטיים עד לניתוקם מוחלט. להרוות את פעילות הטריפסין על ידי הוספת מדיום (7 מ"ל), לאסוף את התאים לתוך צינור צנטריפוגה, צנטריפוגה (200 x גרם, 3 דקות).
- הסר את הסופרנטנט ושלח את התאים במדיום (10 מ"ל). לספור את התאים ולקבוע את הכדאיות באמצעות הרחקה כחולה טריפאן.
- זרעו את התאים (6 מיליון תאי HCT116 בכל בדיקה) לצלחת של 150 מ"מ והתאימו 20 מ"ל של תרבית תאים בסך הכל. מערבבים מראש את המדיום ואת התאים בצינור צנטריפוגה לפני הזריעה כדי להשיג חלוקה שווה של התאים בצלחת. הכן מנה מקבילה כאשר נדרש נורמליזציה לפי מספר התא.
- תרבית את התאים בתנאים סטנדרטיים במשך 72 שעות. התאים יגיעו לכ-80%-90% מפגש.
- אפנון רמות הפוספט של אינוזיטול עם NaF וקצירת תאים
- טיפול NaF: יש להוסיף NaF (10 מ"מ) שעה אחת לפני הקטיף למדיום. מערבבים את המדיום על ידי מערבולת הצלחת/פיפטינג ודוגרים על התאים למשך 60 דקות בתנאים סטנדרטיים.
- לאחר טיפול NaF, הסר את המדיום מהתאים והנח את התאים על קרח.
- לשטוף את התאים פעמיים עם PBS (5 מ"ל, 4 מעלות צלזיוס) ולהסיר את PBS לחלוטין מן הכלים.
- הוסף חומצה פרכלורית (PA) (1 מ"ל, 1 מ', 4 °C ). הקפידו לכסות את כל פני השטח ב-PA (התאים יהפכו לבנים ככל שהחלבונים יזרזו). דגירה של התאים במשך 10 דקות על שולחן הטיה בטמפרטורה של 4 מעלות צלזיוס.
- לאסוף PA לתוך צינור צנטריפוגה ולהסיר את הפסולת המזהמת על ידי צנטריפוגה (17,000 x גרם, 5 דקות, 4 מעלות צלזיוס). הוסף את הסופרנאטנט לחרוזי TiO2 המוכנים למשיכת InsPs.
- לשטוף את המנה לאחר מיצוי פעמיים עם PBS (5 מ"ל, r.t.) עבור deacidification; להסיר PBS לחלוטין מן המנה.
- לבודד את החלבונים שעל הצלחת באמצעות תוספת של חיץ תזה תאית (1.5 מ"ל, r.t.; 0.1% נתרן דודציל סולפט [SDS] ב-0.1 M NaOH). דגרו את המנה במשך 15 דקות על שולחן הטיה ב- r.t. מעבירים את התא ליזאט לתוך צינור צנטריפוגה וצנטריפוגה (17,000 x גרם, 5 דקות, 4 מעלות צלזיוס). יש לאחסן את הסופר-נטנט בטמפרטורה של -80°C עד לקביעת ריכוז החלבון באמצעות בדיקת חלבון DC באמצעות אלבומין בסרום בקר כתקן כיול (בדרך כלל, מנה אחת של 150 מ"מ מכילה כ-10 מ"ג חלבונים).
- קביעת מספרי תאים ממנות מקבילות: קצירת התאים של המנה המקבילה באמצעות טריפסין כמתואר בשלב 3.1.2 (השתמש בטריפסין-EDTA של 5 מ"ל עבור צלחת 150 מ"מ) והסר את המדיום. החזירו את כדור התא ב-PBS (5 מ"ל), ערבבו כראוי וספרו את התאים. בצע שלב זה ממש לפני הקציר באמצעות מרווה ישירה כדי לקבל ספירות תאים מייצגות. בנוסף, מדוד את נפח התאים בשיטה מתאימה (למשל, באמצעות מכונה רב-ממדית).
- העשרת TiO2 של פוספטים אינוזיטול
הערה: כדי למנוע פירוק חומצי של תרכובות זרחניות, בצע את כל שלבי ההעשרה עד לאלוטציה על קרח, וקצר את כל הריאגנטים ל -4 מעלות צלזיוס. שמור את הזמן למיצוי למינימום (1.5-2 שעות). בצע את כל שלבי החילוץ עם 1 M PA.- הכנת חרוזים: לשטוף TiO2 חרוזים (5 מ"ג לדגימה) עם ddH2O (1 מ"ל) וצנטריפוגה (3,500 x גרם, 1 דקה, 4 °C). הסר ddH2O ולשטוף את החרוזים עם PA (1 מ"ל). הסר PA על ידי צנטריפוגה (3,500 x גרם, דקה אחת, 4 °C). יש להשעות את החרוזים ב-PA (50 μL לדגימה).
- הוסף את הסופרנטנט המכיל תרכובות זרחניות (השווה סעיף 3.2, שלב 3.2.5) לתרחיף החרוזים, למערבולת, ולאחר מכן סובב את הדגימה למשך 20 דקות ב-4 מעלות צלזיוס.
- צנטריפוגה של הדגימה (3,500 x גרם, 1 דקה, 4 מעלות צלזיוס) ולהשליך את supernatant. לשטוף את החרוזים עם PA (500 μL) וצנטריפוגה (3,500 x גרם, 1 דקה, 4 °C). יש להשליך את הסופר-נטנט ולחזור על שלב הכביסה.
- הוסף NH4OH (200 μL, 3%) לחרוזים ולהחייאה. סובב את הדגימה במשך 5 דקות ב- r.t.
- צנטריפוגה את הדגימה (3,500 x גרם, 1 דקה) ולהעביר את supernatant לתוך צינור צנטריפוגה חדש.
- חזור על שלבים 3.3.4 ו- 3.3.5 ושלב את האלואנטים. השליכו את החרוזים.
- צנטריפוגה את eluents המשולבים (17,000 x גרם, 1 דקה, 4 °C) כדי להסיר כל שאריות מסיסות.
- יבש לחלוטין את הסופרנטנט תחת אידוי ואקום (70 דקות, 60 מעלות צלזיוס, V-AQ). יש להוסיף ddH2O (50 μL) לתמציות המיובשות המכילות InsPs. מערבולת לערבב את הדגימה עד להמסה מלאה. אחסן את הדגימה בטמפרטורה של -20 °C עד לניתוח CE-ESI-MS.
4. ביצוע ריצות CE-ESI-MS
- הכינו תערובת של תקנים פנימיים המכילים 40 μM [13 C 6]1,5-(PP)2-InsP 4, 80 μM [13 C 6]5-PP-InsP 5, 80 μM [13 C 6]1-PP-InsP 5, 400 μM [13 C 6]InsP 6 ו-400 μM [13C 6]Ins(1,3,4,5,6)P 5. לקבוע את הריכוזים של תמיסות SIL IS על ידי NMR כמותי של 31P ו- 1H, בעזרת תקן ייחוס מוסמך, כלומר, חומצה זרחתית.
הערה: כל התקנים הפנימיים של SIL (IS) הנ"ל עם טוהרים גבוהים מ-96% סונתזו וסופקו על ידי קבוצת פידלר15,27. בדומה לנוקלאוטידים, ה-IS האלה יכולים לשאת מולקולות רבות של מים גבישיים ויוני נגד מגוונים. במקום לשקול את החומר ולחשב את הריכוז, מומלץ לקבוע ריכוז של 13C6 פתרונות סטנדרטיים על ידי NMR כמותי 31P. - מערבבים 10 μL של הדגימה עם 0.5 μL של תערובת תקנים פנימיים בבקבוקון מדגם CE. 2 μM [13 C 6]1,5-(PP)2-InsP 4, 4 μM [13 C 6]5-PP-InsP 5, 4 μM [13 C 6]1-PP-InsP 5, 20 μM [13 C 6]InsP 6, ו-20 μM [13C 6]Ins(1,3,4,5,6)P 5 הם הריכוזים הסופיים בתוך הדגימות.
- בעת שימוש במערכת החידוש, לחץ על כפתור שינוי בקבוק, הכנס את 250 מ"ל מוכן של CE פועל חיץ לתוך בקבוק אלקטרוליטים, ולחץ על צינורות נקיים. שמור את מחט חידוש בבקבוקון מים.
- הגדר פרמטרים של ESI ו- MS כפי שמוצג בטבלה 1. מטב את פרמטרי המקור באמצעות ממטב מקור עם תערובת של תקני פוליפוספט אינוזיטול. השג את הגדרות ניטור התגובות המרובות (MRM) באמצעות Masshunter Optimizer עם כל התקנים. התאם את הגדרות ESI ו- MS/MS עבור מכשור שונה.
- בצע ריצה עבור תמציות InsP ובדוק את התוצאה (איור 2). קבעו רצף כשיש דוגמאות נוספות.
- תן ל-MS להיות במצב המתנה לאחר המדידות. אין לכבות את משאבת ה-LC. זרימת נוזל הנדן מגנה על מחט המרסס. החלף את מרסס CE-ESI-MS במרסס LC-ESI-MS כאשר אין אספקת נוזל נדן.
5. ניתוח נתונים
- פתח את תוכנת ניתוח כמותי (עבור QQQ), צור אצווה עבור כל הדגימות.
- צור שיטה חדשה מנתוני MRM שנרכשו. הגדר את ה- INSPS [13C6] כסטנדרטים פנימיים - (ISTD). בדוק את הגדרת תרכובת MRM, הגדרת זמן שמירה, הגדרת ISTD, הגדרת ריכוז והגדרת מזהה. העבר את האימות וצא כדי להחיל את השיטה על האצווה הנוכחית. שמור את השיטה.
- בדוק אם כל שיא באצווה משולב כראוי; אחרת, שלב את השיא באופן ידני.
- ייצא את התוצאות לגיליון אלקטרוני. בצע את הכמות של אינוזיטול (פירו)פוספטים על ידי השוואת תגובת השיא האנליטית עם תגובת השיא המתאימה של SIL IS עם ריכוזים ידועים. עקומות כיול תיאורטיות וניסיוניות מוצגות בטבלה 2 עבור 5-PP-InsP 5, InsP 6 ו- Ins(1,3,4,5,6)P 5 עם הטווח הליניארי.
הערה: התנאי המקדים להפעלת עקומת כיול תיאורטית הוא שימוש בתבנית איזוטופית איכותית של אנליטים ממוקדים ובריכוז אמין. הערך את הטווח הליניארי. עקומת כיול חיונית לכמות מוחלטת כאשר הרכישה המלאה של התקנים האיזוטופיים הנ"ל אינה מעשית. - עם הריכוז הנמדד בתמיסת תמצית InsP ונפחו, חשב את הכמויות המוחלטות. יתר על כן, לנרמל את הסכום על ידי ספירת תאים או תוכן חלבון. חשב את הריכוז התאי בהתבסס על ספירת תאים ונפח תא ממוצע של HCT116 (1.68 fL).
תוצאות
התוצאות המוצגות כאן נועדו להמחיש את הפוטנציאל של ניתוח CE-ESI-MS. הנתונים המדווחים מתארים ריצת CE-ESI-MS ללא רבב מבחינה טכנית. ראשית, מוצגת תערובת של תקני אינוזיטול פירופוספט (איור 1) ותמצית תאי יונקים (איור 2). שנית, ניתנת השוואה של שני קווי תאים מסוג HCT116 (איור 3) ותאי HCT116 שטופלו ב-NaF (איור 4).
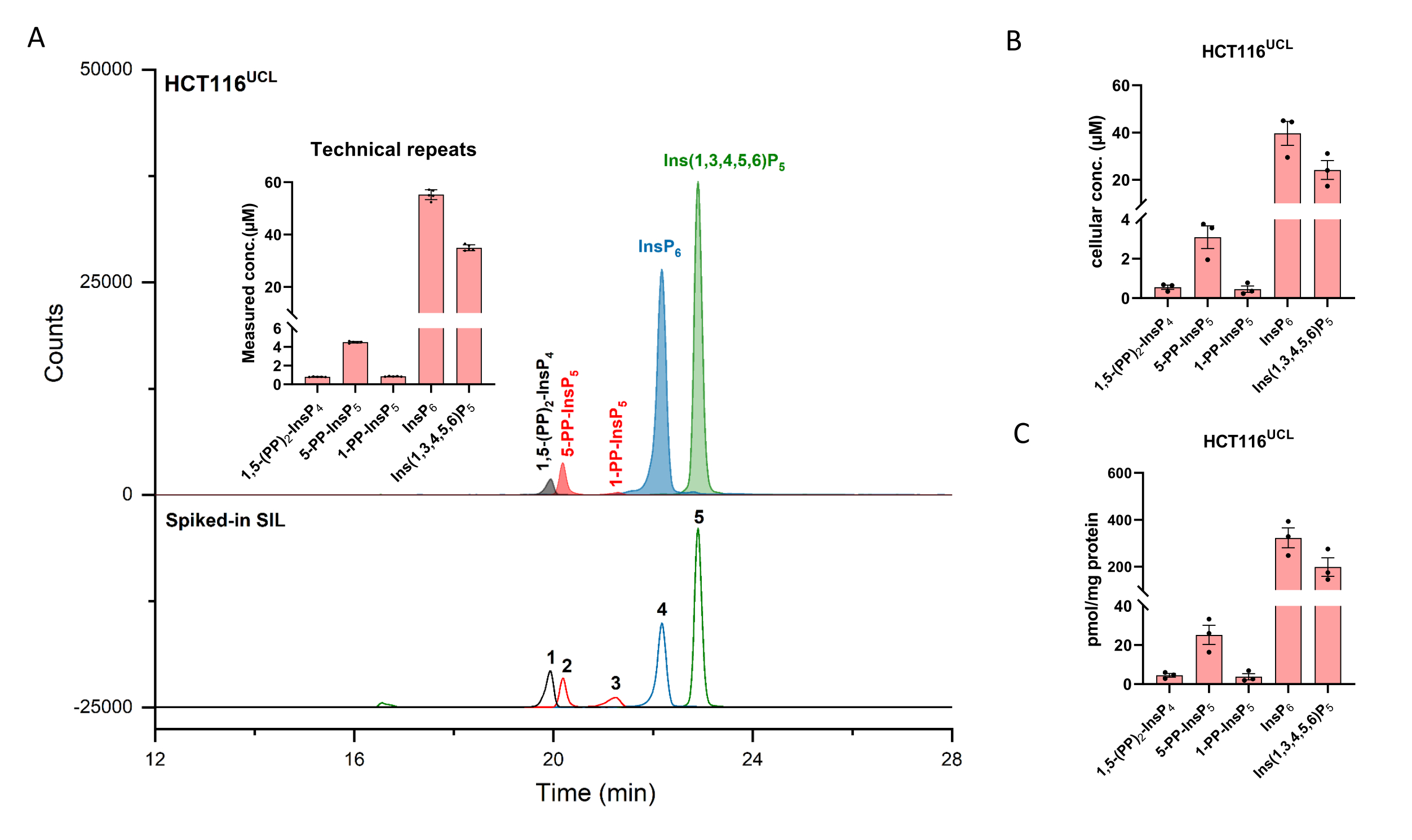
אלקטרופרוגרמות יונים מופקות (EIEs) של תקני אינוזיטול (פירו)פוספט בריכוז של 2 μM מוצגות באיור 1. מטבוליזם של אינוזיטול pyrophosphates ביונקים עם מבנים פשוטים שלהם מוכנס. ארבעת הפירופוספטים של אינוזיטול ביונקים, 1,5-(PP)2-InsP 4, 5-PP-InsP 5, 1-PP-InsP 5 ו-5-PP-Ins(1,3,4,6)P 4 נבדלים היטב באמצעות השיטה המתוארת. הרצת CE-ESI-MS של HCT116UCL מתוארת באיור 2. בעזרת תקנים פנימיים יציבים המסומנים באיזוטופים (SIL), ניתן להשיג בקלות כמות מוחלטת על ידי השוואת תגובת האות עם ה-SIL המחורץ של הריכוז הידוע. מוצגים ה- EIEs המשולבים של הפוספט של אינוזיטול מ- InsP5 ל- (PP)2-InsP 4 ו- EIEs לא משולבים של התבניות האיזוטופיות שלהם. RSDs של כל האנליטים משש חזרות טכניות הם בטווח של 4%. עם הריכוז הנמדד ואת נפח התמציות, כמות האנליטים ניתן לחשב. עם ספירת התאים ונפח התא, או תכולת החלבון, ריכוז תאי מוחלט (μM) או כמות מנורמלת על ידי תכולת חלבון (pmol/mg חלבון) הם בדרך כלל התוצאות הסופיות של ניתוח כזה.
על פי מחקר קודם, שתי קבוצות של תאי HCT116 מתפצלים יש וריאציה של רמות InsP 8, תאי HCT116UCL מכילים רמות גבוהות פי 6 של InsP8 מאשר תאי HCT116NIH 29. בשיטת CE-MS, ניתן לכמת בקלות 1,5-(PP)2-InsP 4 ב-HCT116 NIH (איור 3), ותאי HCT116 UCL מכילים רמות גבוהות פי 7 של InsP8 מאשר ב-HCT116 NIH. בנוסף, הצטברות משמעותית של 1,5-(PP)2-InsP 4 בתאי HCT116UCL מקבילה לעלייה משמעותית של 1-PP-InsP5, המוצגת כעת באופן כמותי באיור 3.
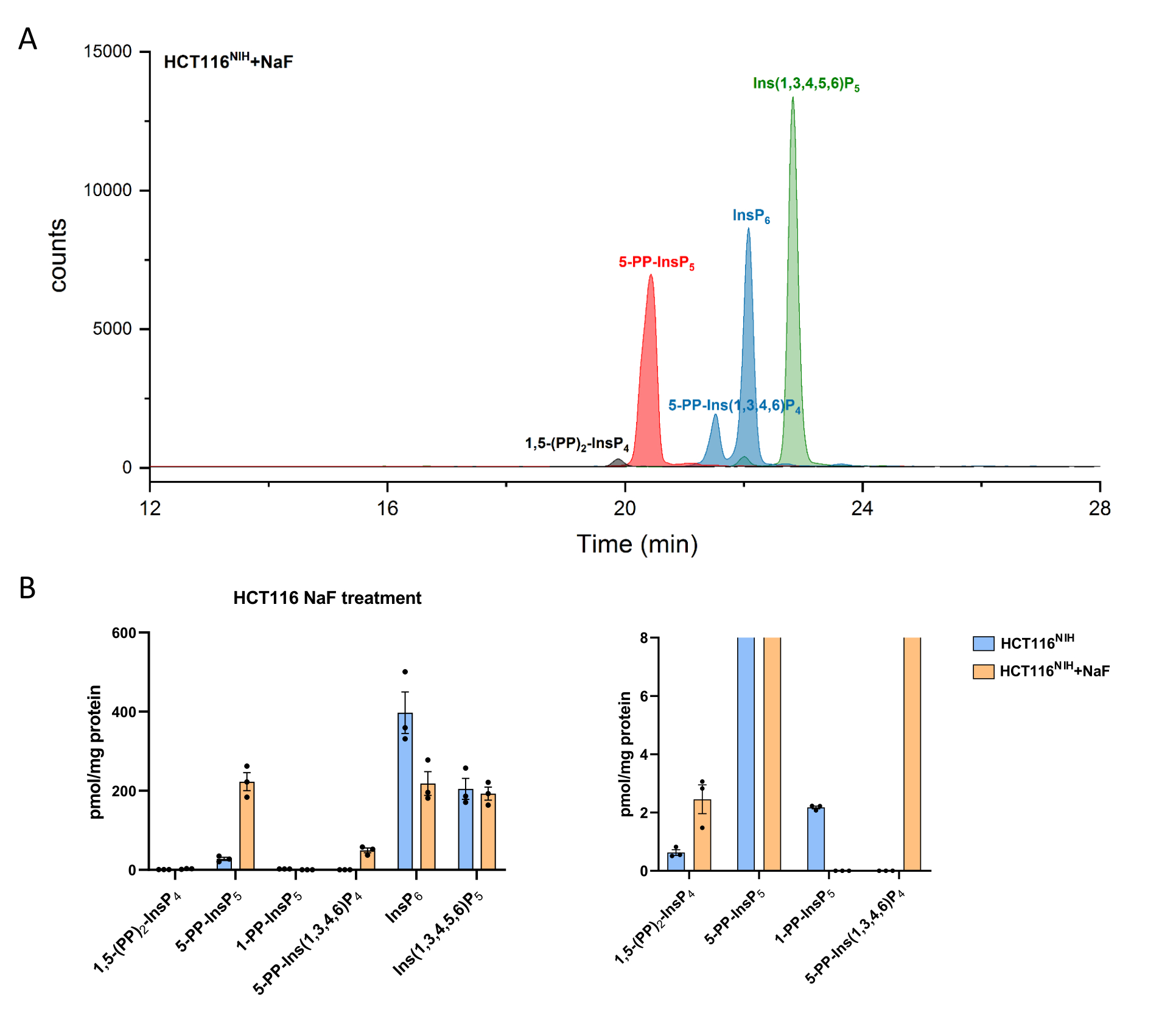
רמות PP-InsPs עולות על ידי עיכוב הדה-פוספורילציה שלהם באמצעות נתרן פלואוריד. ניתוח CE-ESI-MS של תאי HCT116NIH שטופלו ב-NaF הדגים את הגובה של -5-PP-InsP 5 יחד עם ירידה ב-InsP 6 והופעה של 5-PP-Ins(1,3,4,6)P 4 (איור 4). חוץ מזה, הגובה של רמות InsP8 מורגש, בעוד 1-PP-InsP5 יורד במידה מסוימת. 1-PP-InsP5 אינו נעדר לחלוטין ב-HCT116NIH שטופל ב-NAF, אך בעיקר תחת מגבלת הגילוי או הכמות.

איור 1: אלקטרופרוגרמות יונים טיפוסיות (EIEs) של תקני אינוזיטול (פירו)פוספט בניתוח CE-ESI-MS באמצעות הפרוטוקול המתואר. הריכוז של כל אנאליט הוא 2 μM. נפח הדגימה המוזרק הוא בערך 10 nL עם זריקה ב 50 mbar במשך 10 שניות. מוסיף להראות את חילוף החומרים של אינוזיטול pyrophosphates ביונקים. IPPK: אינוזיטול פנטקיספוספט 2-קינאז, IP6K: אינוזיטול הקסקיספוספט קינאז, PPIP5K: דיפוספינוזיטול פנטקיספוספט קינאז, DIPP1: דיפוספינוזיטולפוליפוספט פוספוהידרולאז 1. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 2: פרופיל InsP מייצג של תאי HCT116 UCL. (A) EIEs של האינוסיטול הראשי (פירו)פוספטים ב-HCT116 NIH ו-SIL ISs 2 μM [13 C 6]1,5-(PP)2-InsP 4 (1), 4 μM [13 C 6]5-PP-InsP 5 (2), 4 μM [13 C 6]1-PP-InsP5 (3), 20 μM [13C 6]InsP 6 (4), ו-20 μM [13C 6]Ins(1,3,4,5,6)P 5 (5). התוספות מראות שש חזרות טכניות של ניתוח InsP על ידי CE-ESI-MS, הנתונים מוצגים כאמצעי ±SD. (B) ריכוז תאי של PP-InsPs ו- InsPs בקווי תאים אנושיים HCT116UCL ו- (C) PP-InsPs ו- InsPs כמות מנורמלת על ידי תכולת חלבון. נתונים הם אמצעים ± SEM משלושה ניסויים עצמאיים. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 3: שונות ברמות InsP8 בין שני תאי HCT116 מתפצלים. (א) EIEs של אינוזיטול פירופוספט ב- HCT116 UCL ו- HCT116NIH. InsP8 ב-HCT116UCL נפוץ יותר באופן ניכר מאשר ב-HCT116NIH. (B) היחס בין אינוזיטול פירופוספט ל-InsP6 (%) בשני תאי HCT116. תאי HCT116UCL מכילים רמות גבוהות פי 7 של InsP8 בהשוואה ל-HCT116NIH, בעוד שרמות 5-PP-InsP5 שוות. נתונים הם אמצעים ± SEM משלושה ניסויים עצמאיים. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 4: רמות אינוזיטול (פירו)פוספט בתאי HCT116NIH, עם טיפול ב-NAF. (A) EIEs של אינוזיטול (פירו)פוספט ב- HCT116NIH עם טיפול בנתרן פלואוריד (NaF, 10 mM). רמות של אינוזיטול פירופוספט כולל 1,5-(PP)2-InsP 4, 5-PP-InsP 5 ו-5-PP-Ins(1,3,4,6)P4 עולות באמצעות חסימת הדה-פוספורילציה שלהם באמצעות NaF. (B) רמות אינוזיטול (פירו)פוספט (הכמויות מנורמלות לפי תכולת חלבונים) בתאי HCT116NIH שלא טופלו וטופלו ב-NaF. נתונים הם אמצעים ± SEM משלושה ניסויים עצמאיים. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}
טבלה 1: הגדרות פרמטר CE-ESI-MS. פרמטרים של מקור ופרמטרים של iFunnel ממוטבים על-ידי Source ו-iFunnel Optimizer. הגדרות פרמטר MSM עבור אינוזיטול (פירו)פוספטים ממוטבות על ידי MassHunter Optimizer. אנא לחץ כאן כדי להוריד טבלה זו.
טבלה 2: משוואת רגרסיה תיאורטית וניסויית. הריכוז של [13 C 6]5-PP-InsP 5 [13 C 6]InsP 6, ו-[13C 6]Ins (1, 3, 4, 5, 6)P5 הוא 4 μM, 20 μM ו-20 μM, בהתאמה. עבור משוואת הרגרסיה, ריכוז 5-PP-InsP 5 הוא 0.04 μM, 0.1 μM, 0.2 μM, 0.4 μM, 1 μM, 2 μM, 4 μM, 8 μM, 16 μM, 24 μM. InsP 6 ו- Ins (1, 3, 4, 5,6)P 5 הריכוז הוא 0.2 μM, 0.5 μM, 1 μM, 2 μM,5 μM, 10 μM, 20 μM, 40 μM, 80 μM, 120 μM. x הוא ריכוז, y הוא (שטח InsP)12C/(שטח InsP)13C. אנא לחץ כאן כדי להוריד טבלה זו.
Discussion
מוצגת כאן שיטה מעשית ורגישה לכמות של אינוזיטול pyrophosphates טעון מאוד בתאי יונקים. שילוב גישת ניתוח זו עם מיצוי InsP המתקדם הנוכחי עם חומצה פרכלורית ואחריו העשרה עם TiO2, ניתוח CE-ESI-MS יש יתרונות חסרי תקדים. לגבי התפוקה, הרגישות, היציבות, הכמות האבסולוטית, זיהוי האיזומרים והתלות במטריצה, שיטה זו בולטת בהשוואה לגישות אחרות. פרוטוקול זה ישים לתאי יונקים, אך אכן אסטרטגיה זו מצליחה בדגימות רבות ושונות (למשל, שמרים, צמחים, טפילים, רקמות עכבר וכו ').
פרוטוקול המיצוי המיושם משחזר באופן מלא PP-InsPs ו- InsP6 מתמציות תאי יונקים16,19. הוא גם יחלץ מטבוליטים אניוניים רבים אחרים, במיוחד מינים המכילים פוספטים, כגון פוספטים סוכר ונוקלאוטידים. הערכה של התאוששות ופירוק עבור analytes של המשתמש עם פרוטוקול זה יהיה צורך.
באופן כללי, מערכת CE-ESI-MS פועלת בצורה חלקה ויכולה להכיל כ-200 דגימות מדי שבוע באמצעות פרוטוקול זה. שלא כמו HPLC, עם זאת, CE נחשב כשיטה עבור מומחים ואנשים מיוחדים במשך זמן רב, אשר הגביל את השוק שלה והגביל את היישום שלה. לפיכך, מכשיר CE-ESI-MS נעדר בדרך כלל בפקולטות אנליטיות. אנשים שרוצים לבצע ניתוח CE-ESI-MS כנראה חסרים ניסיון CE וישקיעו יותר זמן בפתרון בעיות. כאן, השלבים הקריטיים מודגשים. בראש ובראשונה איכות החתך הנימי. הרגישות והיציבות של ספריי ESI מסתמכות בעיקר על חתך נימי ממדרגה ראשונה. שנית, קצה היציאה הנימית צריך להיות בדיוק 0.1 מ"מ מקצה המרסס. מחט המרסס ונימי CE צריכים להיות בכיוון הצירי. איכות תרסיס ESI היא קריטית לכמות; יש לבצע ריצות טכניות כדי להעריך את יכולת החזרה.
עם הפרוטוקול המתואר, גבול הכמות (LOQ) עבור PP-InsPs הוא 40 ננומטר עם זריקה ב 50 mbar עבור 10 שניות (10 nL). ישנן מספר גישות כדי להגדיל עוד יותר את הרגישות לשיטה. ראשית, זריקה ב 100 mbar עבור 20 s (40 nL) עדיין יביא לצורת שיא טובה ורזולוציה מספקת עבור regioisomers 5-PP-InsP 5 ו 1-PP-InsP5. שנית, תמציות InsP ניתן להמיס בכמות קטנה יותר של מים. שלישית, ניתן להאריך את זמן השהייה בעת שימוש בפחות מעברי MRM לכמות. בנוסף, מקור יונים CE-MS המשתמש בזרימת נוזל נדן נמוכה במיוחד יגדיל משמעותית את הרגישות.
מאגר CE פועל עם pH 9 מספק את הרזולוציה הטובה ביותר בין InsP6-InsP 8. כאשר מגדילים את ה- pH ל -9.7, הרזולוציה בקרב InsP3-InsP 6 תשתפר באופן משמעותי. בשל הרזולוציה המצוינת, אורך נימי קצר יותר של 72 ס"מ מומלץ להגדלת התפוקה עוד יותר. חוץ מזה, טמפרטורת קלטת CE גבוהה יותר ב 40 מעלות צלזיוס מקטינה את צמיגות המאגר האלקטרופורטי מימי ומאיצה את תנועתם תחת EOF. על פי דרישות מחקר שונות, שינויים בשיטה זו יכולים להקל עוד יותר על ניתוח InsPs ו- PP-InsPs. לכן, לפרוטוקולי CE-ESI-MS המתוארים יש פוטנציאל לפתוח אפיקי מחקר חדשים למשפחה רבת פנים זו של מולקולות איתות.
Disclosures
המחברים מצהירים שאין אינטרסים מתחרים.
Acknowledgements
פרויקט זה קיבל מימון ממועצת המחקר האירופית (ERC) במסגרת תוכנית המחקר והחדשנות Horizon 2020 של האיחוד האירופי (הסכם מענקים מס' 864246, ל-HJJ). DQ מודים על התמיכה הכספית של תוכנית בריג'יט-שליבן-לנגה. AS נתמך על ידי מענק תוכנית MRC MR / T028904/1.
Materials
| Name | Company | Catalog Number | Comments |
| Materials | |||
| 1.5 mL microcentrifuge tubes | Greiner Bio-One | 616201 | - |
| 15 cm tissue culture dishes | Thermo Fisher | 168381 | - |
| 2.0 mL microcentrifuge tubes | Greiner Bio-One | 623201 | - |
| 50 mL centrifuge tubes | Greiner Bio-One | 227261 | - |
| 96-well plates | Thermo Fisher | 260836 | for the DC protein assay |
| CE fused silica capillary | CS Chromatographie | 105180 | 50 µM i.d. 360 µM o.d. |
| Pipette tips | Starlab | I1054-0001, S1111-6701, S1113-1700, S1111-3700 | 10 mL, 1000 µL, 200 µL, 10 µL pipette tips |
| Serological pipets | TPP | 94550, 94525, 94010, 94005 | 50 mL, 25, mL, 10 mL, 5 mL serological pipettes |
| T75 flasks | TPP | 90076 | - |
| Chemicals and Reagents | |||
| NaOH | AppliChem | A6829,0500 | sodium hydroxide pellets for molecular biology, for preparation of cell lysis buffer |
| 0.25% trypsin-EDTA | Gibco | 25200056 | - |
| Ammonium acetate | Thermo Fisher | 1677373 | HPLC grade |
| BSA | Thermo Fisher | 23209 | albumin standard (2.0 mg/mL) for standard curve preparation |
| DC protein assay | Biorad | 5000116 | DC protein assay reagents package |
| DMEM | Gibco | 41966029 | high glucose, pyruvate |
| FBS | Gibco | 10270106, 10500064 (heat inactivated) | 10270106 for HCT116UCL, 10500064 for HCT116NIH |
| Isopropanol | Carl Roth | AE73.2 | 99.95% LC-MS grade |
| NH4OH, 10% | Carl Roth | 6756.1 | for preparation of 3% NH4OH |
| PBS | Gibco | 10010015 | - |
| Perchloric acid, 70% | Carl Roth | 9216.1 | for preparation of 1 M perchloric acid |
| SDS | SERVA | 20760.02 | for preparation of cell lysis buffer |
| Sodium fluoride | Sigma Aldrich | S7920 | - |
| TiO2 beads | GL Sciences | 5020-75000 | 5 µm particle size |
| Trypan blue solution | Gibco | 15250061 | trypan blue stain (0.4%) |
| Ultrapure (Type 1) water | Milli-Q | ZRQSVP3WW | model: Direct-Q 3 UV Water Purification System |
| Equipment | |||
| Analytical balance | Mettler Toledo | 30105893 | model: XPE26; for weighing of beads (5-6 mg per sample) |
| Automated cell counter | Logos Biosystems | L40002 | model: LUNA-II Automated Cell Counter |
| Benchtop centrifuge | Hettich | 1401 | model: UNIVERSAL 320 |
| Benchtop centrifuge with cooling | VWR | 521-1647P | model: Microstar 17R |
| CE system | Agilent | G7100A | - |
| CE/MS Adapter Kit | Agilent | G1603A | - |
| CE/MS Sprayer Kit | Agilent | G1607A | - |
| Cell counting slides | Logos Biosystems | L12001 | LUNA Cell Counting Slides |
| Centrifugal evaporator | Eppendorf | 5305000304 | model: Concentrator plus complete system |
| ESI source | Agilent | AJS ESI | - |
| Super Support Film | Nisshin EM Co. Ltd, Tokyo | 647 | |
| Humidified incubator | Binder | 9040-0088 | model: CB E6.1, for cultivation of mammalian cells |
| Ice box | - | - | should provide enough space for samples, dishes, etc. |
| Isocratic LC system | Agilent | G7110B 1260 Iso Pump | model: Infinity II Quaternary system |
| MSD | Agilent | G6495C | triple quadrupole |
| Multiplate reader | Tecan | 30086375 | model: SPARK 10 M |
| Pipette filler | Thermo Fisher | 10072332 | for serological pipettes |
| Pipettes | Brand | 705884, 705880, 705878, 705872, 705870 | various pipettes |
| Rotator | Labnet | H5500 | model: Mini LabRoller Rotator |
| Shortix capillary column cutter | SGT | S0020 | - |
| Test tube shaker (vortex mixer) | Carl Roth | HXH6.1 | model: Rotilabo-Mini Vortex |
| Tilt table | Labnet | S0600 | model: EDURO MiniMix Nutating Mixer |
| Water bath | Thermo Fisher | FSGPD05 | model: Isotemp GPD 05 |
| Software | |||
| MassHunter Workstation | Agilent | Version 10.1 | - |
| MassHunter Workstation LC/MS Data Acquisition | Agilent | Version 10.1 | - |
| MassHunter Workstation Optimizer | Agilent | Version 10.1 | - |
| MassHunter Workstation Qualitative Analysis | Agilent | Version 10.0 | - |
| QQQ Quantitaion Analysis | Agilent | Version 10.1 | - |
References
- Stephens, L., et al. The detection, purification, structural characterization, and metabolism of diphosphoinositol pentakisphosphate(s) and bisdiphosphoinositol tetrakisphosphate(s). The Journal of Biological Chemistry. 268 (6), 4009-4015 (1993).
- Menniti, F. S., Miller, R. N., Putney, J. W. Jr, Shears, S. B. Turnover of inositol polyphosphate pyrophosphates in pancreatoma cells. The Journal of Biological Chemistry. 268 (6), 3850-3856 (1993).
- Shears, S. B. Inositol pyrophosphates: Why so many phosphates. Advances in Biological Regulation. 57, 203-216 (2015).
- Irvine, R. F., Schell, M. J. Back in the water: the return of the inositol phosphates. Nature Reviews Molecular Cell Biology. 2 (5), 327-338 (2001).
- Szijgyarto, Z., Garedew, A., Azevedo, C., Saiardi, A. Influence of inositol pyrophosphates on cellular energy dynamics. Science. 334 (6057), 802-805 (2011).
- Wild, R., et al. Control of eukaryotic phosphate homeostasis by inositol polyphosphate sensor domains. Science. 352 (6288), 986-990 (2016).
- Chakraborty, A., et al. Inositol pyrophosphates inhibit Akt signaling, thereby regulating insulin sensitivity and weight gain. Cell. 143 (6), 897-910 (2010).
- Bittner, T., et al. Photolysis of caged inositol pyrophosphate InsP8 directly modulates intracellular Ca2+ oscillations and controls C2AB domain localization. Journal of the American Chemical Society. 142 (24), (2020).
- Hauke, S., et al. Photolysis of cell-permeant caged inositol pyrophosphates controls oscillations of cytosolic calcium in a β-cell line. Chemical Science. 10 (9), 2687-2692 (2019).
- Saiardi, A., Sciambi, C., McCaffery, J. M., Wendland, B., Snyder, S. H. Inositol pyrophosphates regulate endocytic trafficking. Proceedings of the National Academy of Sciences of the United States of America. 99 (22), 14206-14211 (2002).
- Koldobskiy, M. A., et al. p53-mediated apoptosis requires inositol hexakisphosphate kinase-2. Proceedings of the National Academy of Sciences of the United States of America. 107 (49), 20947(2010).
- Rao, F., et al. Inositol hexakisphosphate kinase-1 mediates assembly/disassembly of the CRL4-signalosome complex to regulate DNA repair and cell death. Proceedings of the National Academy of Sciences. 111 (45), 16005(2014).
- Williams, S. P., Gillaspy, G. E., Perera, I. Y. Biosynthesis and possible functions of inositol pyrophosphates in plants. Frontiers in Plant Science. 6, 67(2015).
- Wilson, M. S. C., Livermore, T. M., Saiardi, A. Inositol pyrophosphates: between signalling and metabolism. The Biochemical Journal. 452 (3), 369-379 (2013).
- Harmel, R. K., et al. Harnessing 13C-labeled myo-inositol to interrogate inositol phosphate messengers by NMR. Chemical Science. 10 (20), 5267-5274 (2019).
- Qiu, D., et al. Analysis of inositol phosphate metabolism by capillary electrophoresis electrospray ionization mass spectrometry. Nature Communications. 11 (1), 6035(2020).
- Azevedo, C., Saiardi, A. Extraction and analysis of soluble inositol polyphosphates from yeast. Nature Protocols. 1 (5), 2416-2422 (2006).
- Wilson, M. S. C., Saiardi, A. Importance of radioactive labelling to elucidate inositol polyphosphate signalling. Phosphate Labeling and Sensing in Chemical Biology. , 67-87 (2017).
- Wilson, M. S. C., Bulley, S. J., Pisani, F., Irvine, R. F., Saiardi, A. A novel method for the purification of inositol phosphates from biological samples reveals that no phytate is present in human plasma or urine. Open Biology. 5 (3), 150014(2015).
- Losito, O., Szijgyarto, Z., Resnick, A. C., Saiardi, A. Inositol pyrophosphates and their unique metabolic complexity: analysis by gel electrophoresis. PloS One. 4 (5), 5580(2009).
- Dong, J., et al. Inositol pyrophosphate InsP8 acts as an intracellular phosphate signal in arabidopsis. Molecular Plant. 12 (11), 1463-1473 (2019).
- Riemer, E., et al. ITPK1 is an InsP6/ADP phosphotransferase that controls systemic phosphate homeostasis in Arabidopsis. bioRxiv. , (2020).
- Whitfield, H., et al. An ATP-responsive metabolic cassette comprised of inositol tris/tetrakisphosphate kinase 1 (ITPK1) and inositol pentakisphosphate 2-kinase (IPK1) buffers diphosphosphoinositol phosphate levels. The Biochemical Journal. 477 (14), 2621-2638 (2020).
- Ito, M., et al. Hydrophilic interaction liquid chromatography-tandem mass spectrometry for the quantitative analysis of mammalian-derived inositol poly/pyrophosphates. Journal of chromatography. A. 1573, 87-97 (2018).
- Mantilla, B. S., Amaral, L. D. D., Jessen, H. J., Docampo, R. the inositol pyrophosphate biosynthetic pathway of Trypanosoma cruzi. ACS Chemical Biology. 16 (2), 283-292 (2021).
- Wilson, M. S. C., Saiardi, A. Inositol phosphates purification using titanium dioxide beads. Bio-Protocol. 8 (15), 2959(2018).
- Puschmann, R., Harmel, R. K., Fiedler, D. Scalable chemoenzymatic synthesis of inositol pyrophosphates. Biochemistry. 58 (38), 3927-3932 (2019).
- Gu, C., et al. KO of 5-InsP7 kinase activity transforms the HCT116 colon cancer cell line into a hypermetabolic, growth-inhibited phenotype. Proceedings of the National Academy of Sciences of the United States of America. 114 (45), 11968-11973 (2017).
- Gu, C., Wilson, M. S. C., Jessen, H. J., Saiardi, A., Shears, S. B. Inositol Pyrophosphate Profiling of Two HCT116 Cell Lines Uncovers Variation in InsP8 Levels. PloS One. 11 (10), 0165286(2016).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved