Method Article
ניתוח פנוטיפי חצי אוטומטי של תרביות תאים ספרואידים תלת-ממדיים פונקציונליים
In This Article
Summary
אנו מציגים פרוטוקול לגידול ספרואידים בעלי יכולת שחזור גבוהה ואפיונם הפנוטיפי באמצעות לכידת תמונה ופרוטאומיקה.
Abstract
אנו מציגים פרוטוקול המתאר את התכונות והיתרונות של שימוש באינקובטור קלינוסטט עצמאי לגידול, טיפול וניטור תרביות תאים תלת ממדיות. הקלינוסטט מחקה סביבה שבה תאים יכולים להתאסף כספרואידים בעלי יכולת שכפול גבוהה עם כוחות גזירה נמוכים ודיפוזיה של חומרי מזון פעילים. אנו מראים כי הן הפטוציטים הסרטניים והן הפטוציטים שאינם סרטניים (קווי תאים HepG2/C3A ו-THLE-3) דורשים 3 שבועות של גדילה לפני השגת תפקודים דומים לתאי כבד. פרוטוקול זה מדגיש את הנוחות של שימוש באינקובטורים לתאים תלת ממדיים עם מצלמות המנטרות את גדילת התא, שכן ניתן לצלם תמונות כדי לספור ולמדוד ספרואידים בעת הטיפול. אנו מתארים את ההשוואה בין קווי תאים THLE-3 ו- HepG2/C3A, ומראים כיצד ניתן לגדל קווי תאים לא סרטניים כמו גם תאים סרטניים אימורטליים. אנו מדגימים ומדגימים כיצד ניתן לבצע ניסויים בפרוטאומיקה מכמה ספרואידים, שניתן לאסוף מבלי להפריע לאיתות התא, כלומר ללא צורך בטריפסיניזציה. אנו מראים כי ניתוח פרוטאומיקה יכול לשמש לניטור פנוטיפ הכבד הטיפוסי של חילוף החומרים בשרשרת הנשימה וייצור חלבונים המעורבים בניקוי רעלים ממתכות ולתאר מערכת חצי-אוטומטית לספירה ומדידה של שטח הספרואיד. בסך הכל, הפרוטוקול מציג ארגז כלים הכולל אפיון פנוטיפי באמצעות לכידת תמונה וצינור פרוטאומיקה לניסויים במודלים תלת ממדיים של תרביות תאים.
Introduction
תרביות תאים במבחנה הוכחו כהכרחיות ולא יסולא בפז בביסוס ידע בסיסי בביולוגיה. חלק גדול מההבנה המדעית בביולוגיה ובסרטן בפרט הגיע ממערכת התרבית הדו-ממדית, כלומר תאים הגדלים בשכבה חד-שכבתית. למרות שתרבית דו-ממדית הייתה מערכת תרביות התאים הדומיננטית, יש לה חסרונות רבים שעלולים לחנוק התקדמות ביולוגית נוספת. לדוגמה, תרביות דו-ממדיות חסרות אינטראקציות תא-תא החשובות לאיתות ולהתרבות של תאים1. עד כה, מערכות תרבית תלת-ממדיות הוכחו כמודלים טובים יותר של בידול, תגובה לתרופות, פלישת גידולים וביולוגיה 2,3,4,5. מידול תלת ממדי של סרטן ממאיר חיוני במיוחד בשל העלייה באוכלוסייה המזדקנת והתמותה מסרטן. קרצינומה הפטוצלולרית (HCC) היא אחד הגורמים המובילים לתמותה הקשורה לסרטן ברחבי העולם, ולעתים קרובות יש לה פרוגנוזה תהומית6. HCC ידוע כבעל שיעור ריפוי נמוך, תגובה לקויה לתרופות ושיעור גבוה של הופעה חוזרת 6,7,8. פותחו מספר מודלים תלת-ממדיים עבור כבד תקין ו-HCC המחקים את הפיזיולוגיה של רקמת כבד תקינה וממאירה 9,10.
חלק מהמערכות התלת-ממדיות הנוכחיות כוללות שכבות-על נוזליות, ביוריאקטורים, הידרוג'ל, פיגומים ומבנים מודפסים בתלת-ממד. ספרואידים הנוצרים בביוריאקטורים מספקים באופן ספציפי יתרונות ייחודיים מכיוון שהם מחקים את התפלגות הגידול של חשיפה לחומרי מזון, חילופי גזים והתפשטות תאים/ שקט 11. ביוריאקטורים מתאימים במיוחד למודלים של סרטן בשל קלות השימוש בהם, מדרגיות גדולה, דיפוזיה תזונתית ונגישות11. בנוסף, ביוריאקטורים יכולים לאפשר ניסויים בתפוקה גבוהה, יכולת שחזור גבוהה יותר והפחתת טעויות אנוש. הביוריאקטור המשמש במחקר זה ייחודי משום שהוא מדמה מערכת של כבידה מופחתת, אשר ממזערת כוחות גזירה משבשים המופעלים בביוריאקטורים טיפוסיים ומאפשרת שחזור טוב יותר12. כוח הכבידה הרב-כיווני וההפחתה בכוחות הגזירה מאפשרים לתאים להתפתח באופן פיזיולוגי יותר. כראיה, תאי HepG2/C3A הגדלים תחת מתודולוגיה זו מפתחים אברונים כדוריים המייצרים רמות in vivo של ATP, אדנילט קינאז, אוריאה וכולסטרול13,14. בנוסף, הטיפולים התרופתיים במערכת תלת ממדית זו מתקדמים ואוטומטיים יותר בהשוואה לתרביות דו-ממדיות. בתרביות דו-ממדיות, טיפולים תרופתיים חייבים לעתים קרובות להיות במסלול זמן קצר בשל הצורך טריפסיניזציה ושמירה על בריאות התא. עם זאת, במקרה שלנו, אנו יכולים לבצע טיפולים תרופתיים ארוכי טווח של ספרואידים ללא צורך לשבש את המבנה והפיזיולוגיה של התאים. לכן, המעבר מתרבויות דו-ממדיות לתלת-ממדיות הוא הכרחי כדי למדל טוב יותר תופעות ביולוגיות in vivo והתפתחות מדעית נוספת.
מאמר זה מציג מתודולוגיה לגידול ספרואידים בעלי יכולת שחזור גבוהה (איור 1 ואיור 2) ומראה מערכת חצי-אוטומטית לאפיון פנוטיפית של מבנים תלת-ממדיים (איור 3). ברמת התמונה אנו מספקים מידע על ספירה ומדידה של שטח הספרואידים (איור 3). באמצעות שימוש בשיטות ספקטרומטריית מסות, אנו מראים כיצד ניתן להשתמש בפרוטאומיקה כדי להעריך תפקודים ביולוגיים מסוימים (איור 4). על ידי איסוף וניתוח נתונים אלה, אנו מקווים לשפר את ההבנה של הביולוגיה שמאחורי מערכות תרביות תאים תלת-ממדיות.
Protocol
1. חוצצים וריאגנטים
- מצע גדילת תאים לתאי HepG2/C3A: הכינו את מדיום הנשר המעובד של דולבקו (DMEM, 4.5 גרם/ליטר גלוקוז) המכיל 10% נסיוב בקר עוברי (FBS), חומצות אמינו לא חיוניות (1% v/v), L-גלוטמין (1% v/v) ופניצילין/סטרפטומיצין (0.5% v/v). אחסן את מצע הגידול ב- 4 ° C.
- מצע צמיחת תאים לתאי THLE-3: הכינו מדיום גידול תאי אפיתל של הסימפונות [BEGM] המכיל את התוספים שלו (תמצית יותרת המוח מפרות [BPE], הידרוקורטיזון, גורם גדילה אפידרמיס אנושי [hEGF], אפינפרין, טרנספרין, אינסולין, חומצה רטינואית, טריודותירונין, גנטמיצין סולפט-אמפוטריצין [GA]) וכן 10% FBS, 5 ננוגרם/מ"ל hEGF ו-70 נ"ג/מ"ל פוספוטנולמין.
- 500 mM DL-Dithiothreitol (DTT): להכנת 10 מ"ל, יש להשהות מחדש 0.771 גרם DTT ב-10 מ"ל מים באיכות HPLC. יש לאחסן 100 μL aliquots בטמפרטורה של -20°C.
- 200 mM iodoacetamide: כדי להכין 1 מ"ל, resuspend 36 מ"ג של iodoacetamide ב 1 מ"ל של 50 mM אמוניום ביקרבונט. אין לאחסן יודואצטמיד מושעה.
- 5% נתרן דודציל סולפט (SDS): להכנת 50 מ"ל, יש להשעות 2.5 גרם SDS ב-50 מ"ל של 50 מ"ל אמוניום ביקרבונט. יש לאחסן בטמפרטורה של 4°C.
- 12% חומצה זרחתית: להכנת 100 מ"ל, לדלל 14.1 מ"ל של 85% חומצה זרחתית ב 85.9 מ"ל של מים כיתה HPLC. יש לאחסן בבקבוק זכוכית בטמפרטורת החדר (RT).
- חיץ קשירה: כדי להכין 1 L, להוסיף 900 מ"ל של מתנול מרוכז ל 100 מ"ל של 10 mM אמוניום ביקרבונט או TEAB.
- תמיסת 0.1% חומצה טריפלואורואצטית (TFA): יש להוסיף 1 מ"ל TFA מרוכז ל-999 מ"ל מים באיכות HPLC. יש לאחסן בטמפרטורה של 4°C.
- 60% אצטוניטריל/0.1% תמיסת TFA: הוסף 600 מ"ל של אצטוניטריל בדירוג HPLC (60% v/v) ל-399 מ"ל מים באיכות HPLC. לאחר מכן, הוסף 1 מ"ל של TFA מרוכז לפתרון זה, ולאחר מכן לאחסן ב 4 °C.
- Mobile Phase A (MPA) - 0.1% חומצה פורמית: יש להוסיף 1 מ"ל חומצה פורמית מרוכזת ל-999 מ"ל מים באיכות HPLC ולערבב היטב.
- שלב B נייד (MPB) - 80% אצטוניטריל ברמת HPLC + 0.1% חומצה פורמית: הוסף 800 מ"ל של אצטוניטריל ברמת HPLC ל-199 מ"ל מים באיכות HPLC. לאחר מכן, להוסיף 1 מ"ל של חומצה פורמית מרוכזת לתמיסה זו ולערבב.
2. הכנת ספרואידים
הערה: איור 1A מייצג את השלבים הראשונים להכנה ולטיפוח של ספרואידים תלת-ממדיים מקווי תאים.
- הפשירו תאי HepG2/C3A ו-THLE-3 קפואים וגדלו אותם כשכבה חד-שכבתית באמצעות מצע גידול סטנדרטי בתרבית רקמה, בקבוק או צלחת עד שהם מגיעים לכ-80% מפגש.
הערה: כאשר תאים מגיעים למפגש, בדוק את המורפולוגיה התאית הכללית ואת דפוסי הצמיחה באמצעות מיקרוסקופ. לא מומלץ להשתמש בתאים במספרי מעבר גבוהים. - שטפו תאים פעמיים עם תמיסת המלח המאוזנת של האנק (HBSS, השתמשו ב-5 מ"ל עבור בקבוקT75 ס"מ או צלחת של 10 ס"מ).
- הוסיפו 5 מ"ל של 0.05% טריפסין-EDTA מדולל ב-HBSS (דילול 1:2) ודגרו במשך 5 דקות ב-37°C עם 5% CO2. השתמש במיקרוסקופ כדי להעריך את ניתוק התאים והוסף 3 מ"ל של FBS או מצע גידול (המכיל 10% FBS) כדי לנטרל תגובת טריפסין.
- העבר את תרחיף התא לצינור של 15 מ"ל. סובב כלפי מטה במהירות של 270 x גרם ב- RT למשך 5 דקות.
- לשאוף את supernatant ו resuspend תאים ב 5 מ"ל של מדיה צמיחה מלאה.
הערה: אם יש יותר מדי תאים, דלל את תרחיף התאים לפני הספירה. - ספור את מספר התאים ודלל את תרחיף התא במצע צמיחה מלא כדי להשיג 1 x 106 בנפח מרבי של 1.5 מ"ל.
- שיווי משקל של פלטה תחתונה עגולה במיוחד בעלת 24 בארות חיבור המכילה מיקרו-בארות.
- לשטוף את הבארות עם 0.5 מ"ל של מדיה צמיחה.
- צנטריפוגה את הצלחת ב 3,000 x גרם במשך 5 דקות (זה מסיר בועות אוויר מפני השטח של בארות).
- מעבירים את מתלה התא (שהוכן בשלב 1.4) לתוך הצלחת והצנטריפוגה למשך 3 דקות במהירות של 120 x גרם.
הערה: נפח תרחיף התא עשוי להשתנות בהתאם לספירת התאים, אך חשוב להגביל אותו ל-1.5 מ"ל. - לדגור על הצלחת במשך 24 שעות ב 37 ° C עם 5% CO2 כדי להתחיל היווצרות ספרואידים.
3. תרבית ספרואידים לביוריאקטורים (איור 2)
הערה: כדי לשמר את המבנה של ספרואידים, השתמש בקצוות רחבים בעת טיפול בכדורי תלת-ממד.
- אזנו את הביוריאקטור על-ידי מילוי תא הלחות ב-25 מ"ל מים סטריליים ואת תא התא ב-9 מ"ל של מצע גדילה 24 שעות לפני העברת הספרואידים אליו (איור 2A). הקפד להשתמש מזרק 10 מ"ל מחובר למחט ארוכה בעת מילוי לחות ותאי תאים.
- דגור על הביוריאקטור, מסתובב (15 סל"ד) באינקובטור התלת-ממדי (איור 2D,F), במשך 24 שעות ב-37°C עם 5% CO2.
- בעזרת קצוות נשא ברוחב 1 מ"ל, פיפטה עדינה למעלה ולמטה כדי לנתק את הספרואידים מצלחת החיבור הנמוכה במיוחד ולהעביר את הספרואידים לצלחת תרבית רקמה.
- שטפו את צלחת החיבור הנמוכה במיוחד עם 0.5 מ"ל של מצע גידול שחומם מראש כדי ללכוד שאריות ספרואידים והעבירו אותם לאותה צלחת משלב 3.3.
- הערך את הגודל, הקומפקטיות והמעוגלות של הספרואידים באמצעות מיקרוסקופ אור (הגדלה פי 4) ובחר את אלה שנוצרו מספיק.
הערה: תאים נוצרים מספיק כאשר הם קומפקטיים ואינם מתפרקים במהלך הטיפול. חשוב גם שהספרואידים יהיו יחידה אחת ולא יתקבצו יחד. גודל ספרואיד בין 100-200 מיקרומטר מועדף כאשר הוא מועבר לביוריאקטור. - העבירו ספרואידים לביוריאקטור מאוזן מלא ב-5 מ"ל של מצע גידול טרי. לאחר העברת הספרואידים, מלאו לחלוטין את הביוריאקטור במצע גידול טרי, והקפידו להימנע מהוספת בועות לביוריאקטור בעת מילויו במדיה.
- מקמו את הביוריאקטור באינקובטור התלת-ממדי וכווננו את מהירות הסיבוב באמצעות יחידת הבקרה (איור 2C). עבור ספרואידים HepG2/C3A, הגדר את מהירות הסיבוב ל 10-11 סל"ד; עבור ספרואידים THLE-3, הגדר אותו ל 11-12 סל"ד.
הערה: מהירות הסיבוב נקבעת נכון כאשר הספרואידים מפוזרים באופן שווה במרכז הביוריאקטור ולא נוגעים בדפנותיו. איור 2E מראה ביוריאקטור מסתובב. - החלפת מדיה צמיחה כל 2-3 ימים על ידי הסרת 10 מ"ל של מדיה ישנה והחלפתה עם 10 מ"ל של מדיה טרייה; הקפידו לא להסיר ספרואידים בעת החלפת מדיה (איור 2B).
הערה: מומלץ לקיים שגרה של חילופי מדיה (לדוגמה, 48 שעות / 48 שעות / 72 שעות) ולנהל רישום מפורט. - התאם את מהירות הסיבוב בכל פעם שהמדיה משתנה. הגדל את המהירות ככל שהספרואידים גדלים בגודל ובמספר.
- לאחר 15 יום בתרבית, פיצלו את הספרואידים לשני ביוריאקטורים חדשים.
הערה: בהתאם לקו התאים ולקצב הצמיחה, הזמן עשוי להשתנות ויש למטב אותו בנפרד. - לאחר 20 יום, ספרואידים מוכנים לאיסוף.
4. לכידת תמונה וספירה של ספרואידים
הערה: הצינור הפשוט לספירת ספרואידים מוצג באיור 3A. עבור ספירת ספרואידים וקביעת שטח (סעיף 5), קריטי להעריך את הקומפקטיות של המבנים התלת-ממדיים. זה יתרום ניגודיות צבעים משופרת יותר, אשר הכרחי עבור השיטה להיות מדויקת.
- פתח את אפליקציית התלת-ממד המותקנת בטאבלט.
- בחר את הביוריאקטור וצלם תמונה.
הערה: לחלופין, ניתן לצלם תמונה. יש לקחת בחשבון את הפרמטרים הבאים.- מקם רקע שחור מאחורי הביוריאקטור (תמיכה שחורה מסופקת בקופסת הביוריאקטור).
- ודא שאין השתקפות אור על תא התא.
- צלם את התמונה קרוב ככל האפשר לביוריאקטור.
- פתח תמונה אחת בפיג'י (ImageJ).
- הכן את התמונה לניתוח:
- בחרו בכלי אליפסה . צייר עיגול סביב התקע.
- לחץ על מחק בלוח המקשים כדי להפוך את כל מה שנמצא בתוך העיגול לשחור. ציירו עיגול סביב תא התא. ודא שכל הספרואידים נמצאים בתוך המעגל.
- הפעל מאקרו כדי לספור ספרואידים:
- לחץ על תוספים > מאקרו > רשומה. העתק טקסט מאקרו מקובץ משלים 1 למקליט.
- לחץ על צור, וחלון חדש ייפתח. לחץ על הפעל כדי לנתח את התמונה שנפתחה.
- חלון חדש ייפתח עם התוצאות (ספירה, שטח כולל, גודל ממוצע ואזור אחוזים).
- סגור את התמונה וחזור על שלבים 4.4 ו- 4.5 עבור כל תמונה שעבורה יש צורך בספירת הספרואידים. לקבלת עיבוד קל ומהיר יותר, שמור את המאקרו והפעל אותו על-ידי לחיצה על תוספים > מאקרו > הפעל ובחר את המאקרו שנשמר.
5. קביעה פלנימטרית של האזור הספרואידי
הערה: הצינור הפשוט לקביעת האזור הכדורי מוצג באיור 3B.
- צלם תמונות כמתואר בשלב 3.5.
- לפני שמתחילים, הגדר סולם עולמי למדידת שטח הספרואידים.
- פתח בפיג'י תמונה עם סרגל קנה מידה עם ההגדלה הרצויה. בחרו בכלי קו. צייר מעל סרגל קנה המידה (הקו צריך להיות ארוך כמו סרגל קנה המידה).
- Open Analyze > להגדיר קנה מידה. כתוב את המרחק הידוע (אורך הקו מוזן אוטומטית ב - מרחק בפיקסלים. הקפד לסמן את גלובל.
- לחץ על אישור. עכשיו קנה המידה מוגדר לתמונה כי ינותח.
- כדי להתחיל את הקביעה הפלנימטרית, הפעל את המאקרו (קובץ משלים 2).
- לחץ על תהליך > מאקרו > אצווה. בחר את התיקיה המכילה את התמונות לניתוח. תיקיה זו תהיה הקלט.
- בחר את התיקיה שנוצרה בשלב 5.3.1 כפלט. לחץ על תהליך.
- כבדיקת איכות, העריכו אם האזור הכדורי שנמדד תואם לתמונה המקורית.
הערה: המאקרו אינו כולל ספרואידים בקצה שבו ניתן למדוד רק חלק מהספרואידים.
6. אוסף של ספרואידים
הערה: מומלץ מאוד לאסוף ספרואידים באמצעות קצוות רחבים כדי לשמר את המבנה התלת-ממדי שלהם. האיסוף יכול להיעשות באמצעות התקע בחלק הקדמי של הביוריאקטור (איור 2A).
גודל הספרואידים בעת האיסוף עשוי להשתנות בהתאם לקו התא, מספר התאים ההתחלתי ותהליך הפיצול (ימים בתרבית, מספר ספרואידים לכל ביוריאקטור ויחס פיצול).
- כדי לאסוף ספרואידים, הסר 5 מ"ל של מדיה מהביוריאקטור דרך היציאה העליונה באמצעות מזרק המחובר למחט ארוכה. הקפד לתת לכדורים לשקוע למרכז התחתון של הביוריאקטור (ליד היציאה התחתונה).
- פתח את היציאה הקדמית ואסוף ספרואידים באמצעות קצה נשא ברוחב 1 מ"ל. מניחים את הספרואידים בצינורות מיקרוצנטריפוגה. צנטריפוגות את הספרואידים ב 500 x גרם במשך 5 דקות ולהשליך מדיה.
- לשטוף את הספרואידים עם 200 μL של HBSS כדי להסיר את FBS. צנטריפוגה ב 500 x גרם במשך 5 דקות ולהשליך את supernatant.
- יש להקפיא את כדורית הספרואיד בחנקן נוזלי ולאחסן בטמפרטורה של -80°C עד לעיבוד.
7. כדאיות של ספרואידים
הערה: הכדאיות של הספרואיד נקבעה על-ידי מדידת הפעילות של אדנילט קינאז (AK) ששוחרר על-ידי תאים פגומים (איור 4A). בשל שיפוע הדיפוזיה, מדידת AK יעילה כאשר הספרואידים קטנים מ-900 מיקרומטר בקוטר12. אם הספרואידים הופכים גדולים יותר, או אם יש ספק לגבי מדידת הכדאיות, ניתן לבצע בדיקת ATP15.
- אספו את הסופרנאטנט הספרואידי והעבירו 20 μL לצלחת בעלת דופן לבנה 96 בארות (תחתית שטוחה שקופה). הוסף 100 μL של מגיב זיהוי אדנילט קינאז לכל באר הומוגניזציה על ידי pipeting בעדינות למעלה ולמטה.
- הסר בועות על ידי צנטריפוגה בוואקום המהיר למשך 2 דקות ב- RT. דגור על הצלחות במשך 20 דקות ב- RT.
- מקם את הלוח בלומנומטר (קורא לוחות, מצב הארה) והפעל את התוכנית. הגדר את הפרמטרים בהתאם להוראות יצרן הערכה.
הערה: במבחני כדאיות ביולומינסנטיים, ניתן להשתמש בפלט האור הלומנומטר הישיר (בדרך כלל RLUs) כדי לחשב את תגובת התא. מכיוון שאדנילט קינאז ידלוף רק מתאים ששלמות התא שלהם נפגעה, ניתן להשיג שליטה מלאה באדנילט קינאז באמצעות מגיב ליזיס.
8. מיצוי חלבון
הערה: איור 1B מייצג את זרימת העבודה עבור עיבוד ספרואידים ומיצוי חלבונים.
- אספו ספרואידים (סעיף 6) ביום ה-36. להשהות מחדש ספרואידים ב 25 μL של 5% SDS כדי lyse את התאים.
- לאחר הוספת 5% SDS, הומוגניזציה של הגלולה על ידי פיפטציה למעלה ולמטה. במקרים מסוימים כאשר קשה lyse spheroids, להשתמש pestle קטן.
- לדגור על הדגימות עם 20 mM DTT במשך 1 שעה כדי להפחית חלבונים. מערבבים על ידי פיטינג למעלה ולמטה.
- דגרו על דגימות עם יודואצטמיד 40 mM למשך 30 דקות המוגנות מפני חלבונים קלים ואלקילט. מערבבים על ידי פיטינג למעלה ולמטה.
- הוסף 12% תמיסת חומצה זרחתית (10x) לדגימות בריכוז סופי של 1.2%.
- מדללים דגימות בשישה כרכים של חיץ קשירה ומערבבים בעדינות.
- טען את הדגימה ללוח מסנן S-Trap וסובב כלפי מטה במהירות של 500 x גרם למשך 30 שניות.
הערה: אם אמצעי האחסון הכולל של הדגימה בשלב 8.4 חורג מקיבולת נפח העמודה, טען דגימות באצוות וחזור על שלב 8.6 עד שהכל יעבור דרך העמודה. - לשטוף את הדגימות פעמיים עם 150 μL של חיץ קשירה. לאחר כל שטיפה, יש לסובב כלפי מטה ב-500 x גרם למשך 30 שניות ולהשליך את הזרימה.
- לדגור את הדגימות עם 1 מיקרוגרם של טריפסין כיתה ריצוף מדולל 50 mM אמוניום ביקרבונט לילה ב 37 ° C.
הערה: לפחות 20 מיקרוגרם חלבון מומלץ כחומר מוצא לניתוח פרוטאום מקיף. בשביל זה, 1 מיקרוגרם של טריפסין הוא הסכום האידיאלי לעיכול יעיל. הסר בועות בין המאגר למיטת העמודות. - יש לשאוב פפטידים עם 40 μL של 50 mM אמוניום ביקרבונט ולהיכנס לאותו צינור איסוף.
- סובב כלפי מטה ב- 500 x גרם למשך 30 שניות. מוציאים פפטידים עם 40 μL של 0.1% TFA ומאגדים אותם לאותו צינור איסוף.
- סובב כלפי מטה ב- 500 x גרם למשך 30 שניות. יש להצמיד את הפפטידים עם 40 μL של 60% אצטוניטריל ו-0.1% TFA ולאגד אותם באותו צינור איסוף.
- סובב כלפי מטה ב- 500 x גרם למשך 30 שניות. איגום יבש פולט בוואקום המהיר ומאחסן את הדגימות ב -80 מעלות צלזיוס עד לעיבוד.
9. ניקוי דגימה
הערה: לפני שתמשיך לניתוח פרוטאומיקה, יש צורך להסיר את המלח הנוכחי בדגימות. מלחים יכולים להפריע לניתוח ספקטרומטריית מסה של כרומטוגרפיה נוזלית (HPLC-MS) בעלת ביצועים גבוהים כשהם מייננים במהלך התרסיס חשמלי, ומדכאים את האות מפפטידים. ההגדרה לדה-מלחה ששימשה במחקר זה הודגמה בעבר על ידי ג'וזף-צ'אודהורי ועמיתיו16.
- לפני שמתחילים, ודא שיש צלחת איסוף נקייה של 96 בארות כדי לאסוף את הזרימה במהלך השלבים הבאים. ערבבו את שרף C18 (50 מ"ג/מ"ל ב-100% אצטוניטריל) על צלחת ערבוב מגנטית.
- הוסף 70 μL של מתלה שרף C18 לכל באר של צלחת מסנן 96 בארות, לעשות זאת במהירות כדי להבטיח כי שרף C18 לא מצטבר בתחתית פיפטה.
- הפעילו את השואב בעדינות כדי למנוע התזה. השליכו את הזרימה שנאספה על צלחת האיסוף בת 96 הקידוחים.
- לשטוף את השרף עם 100 μL של 0.1% TFA. הפעילו את השואב בעדינות כדי למנוע התזה וערבוב של דגימות והשליכו את הזרימה.
- השהה מחדש כל דגימה ב- 100 μL של 0.1% TFA. בדוק וודא כי ה- pH הוא בין 2-3.
- טען כל דגימה לכל באר של צלחת הסינון. הפעילו את השואב בעדינות כדי למנוע התזה וערבוב של דגימות והשליכו את הזרימה.
- יש לשטוף עם 100 μL של 0.1% TFA. הפעל את השואב בעדינות כדי למנוע התזה וערבוב של דגימות. השליכו את הזרימה.
- החליפו את צלחת האיסוף בת 96 הבארות בצלחת חדשה בת 96 בארות כדי לאסוף את הדגימות שעברו דה-מלח.
- הוסף 60 μL של acetonitrile/0.1% TFA לכל באר. כדי לשלול את הדגימות משרף C18, הפעל את השואב בעדינות כדי למנוע התזה.
- אספו את הזרימה וייבשו בוואקום מהיר ב-RT. המשיכו אל LC-MS/MS או אחסנו את הצלחת בטמפרטורה של -80°C עד לעיבוד הדגימות.
10. ניתוח פרוטאומיקה באמצעות כרומטוגרפיה נוזלית בשילוב ספקטרומטריית מסות
הערה: כדי להפיק את הנתונים עבור כתב יד זה, נעשה שימוש במערכת nLC-MS/MS עם מערך מערכת של שתי עמודות עם השמנה של 300 מיקרומטר ID x 0.5 ס"מ C18 עמודת השמנה וננו-עמודה אנליטית בגודל 75 מיקרומטר x 25 ס"מ C18-AQ (3 מיקרומטר) ארוזה בתוך החברה.
- הכן את השלבים הניידים להפעלה ב- HPLC:
שלב A נייד (MPA): 0.1% חומצה פורמית במים באיכות HPLC
שלב B נייד (MPB): 0.1% חומצה פורמית באצטוניטריל ברמת HPLC - תכנת את שיטת HPLC באופן הבא: 4%-34% MPB במשך 120 דקות, 34%-90% MPB במשך 5 דקות, איזוקרטי 90% MPB במשך 5 דקות; קצב זרימה: 300 nL/min.
- הגדר את שיטת רכישת הטרשת הנפוצה לביצוע רכישה בלתי תלויה בנתונים (DIA) כדי ליצור ספקטרום MS/MS של אותות הפפטיד שזוהו.
- יש להשהות מחדש 1 מיקרוגרם של דגימות ב-10 μL של 0.1% TFA לפני הצבת הדגימות בדוגם האוטומטי nLC.
- הפעל את שיטת nLC-MS כפי שתוכנתה לרכישת פרוטאומיקה קנונית:
- הגדר סריקת MS מלאה ל- 300-1100 m/z במסלול, עם רזולוציה של 120,000 (ב- 200 m/z) ויעד בקרת רווח אוטומטית (AGC) של 125.
- הגדר MS/MS במסלול עם חלון בידוד רציף של 50 m/z עם יעד AGC של 400 ואנרגיית דיסוציאציה התנגשות באנרגיה גבוהה יותר (HCD) של 30.
11. ניתוח נתונים
- יבא את קבצי הנתונים הגולמיים nLC-MS/MS לתוכנת זיהוי שיא התואמת לפלטפורמת MS שבה נעשה שימוש.
- בחר את מסד הנתונים המתאים (אדם, עכבר וכו ').
- הגדר אצטילציה N-terminal כשינוי משתנה וקרבמידומתיל ציסטאין כשינוי קבוע.
- ציין טריפסין כאנזים העיכול עם שני מחשופים חסרים מותרים.
- הגדר סובלנות מסה בהתאם למנתח המסה המשמש לרכישת ספקטרה.
- יצא את הניתוח כגיליון אלקטרוני ועבד את הנתונים הלאה לפי הצורך.
תוצאות
בפרוטוקול הזה אנו מתארים את המאפיינים של אינקובטור תאי תלת-ממד חדשני ללא לחץ, מערכת שתוכננה במיוחד לגידול ספרואידים תלת-ממדיים (איור 2). ביצענו אופטימיזציה של הפרוטוקול לתרבית תלת-ממדית של קווי תאים THLE-3 ו-HepG2/C3A. הפרוטוקול המתואר כאן פשוט לשימוש ומאפשר שחזור וגידול חסכוני של > 100 ספרואידים לכל ביוריאקטור. ברגע שהם מגיעים לביוריאקטור, הספרואידים מטופלים באופן דומה לתאים הנשמרים בתרבית דו-ממדית. תנאי גדילה אופטימליים מושגים על-ידי החלפת המדיה פעמיים עד שלוש בשבוע (איור 2B) והתאמת מהירות הסיבוב בהתאם לגדילה ולגודל של הספרואידים (איור 2C). המערכת הזו, שבה ספרואידים תלת-ממדיים מתורבתים בביוריאקטורים מסתובבים (איור 2E,F), מספקת סביבת גידול אופטימלית למבנים תלת-ממדיים על-ידי חשיפת הספרואידים לכמות שווה ונמוכה מאוד של כוח גזירה.
הראינו בעבר כיצד ניתן להשתמש בספרואידים לניתוח שינוי כרומטין16. כאן אנו מדגימים בפירוט כיצד להשיג ספרואידים בכבד וכיצד ניתן לערוך ניסויים בפרוטאומיקה לצורך ניתוח הפרוטאום המלא (איור 1). בקצרה, הפרוטוקול החל על ידי שימוש בתאים שטוחים THLE-3 או HepG2/C3A עד שהתרבית הגיעה למפגש של 80%. כדי להשתמש בתאי תרבית כספרואידים, כ-2,000 תאים צופו בלוח התקשרות נמוך במיוחד שהכיל מיקרו-בארות כדי לאפשר להם לצבור את עצמם, ולאחר מכן, ספרואידים שנוצרו הועברו לביוריאקטור (איור 1A). למרות שהם פעילים מבחינה תפקודית לאחר 3 שבועות בתרבית, כפי שהודגם בעבר17, אנו מראים תוצאות מספרואידים שנאספו לאחר 36 ימים בתרבית עבור פרוטוקול זה. לאחר האיסוף, ספרואידים סובבו כלפי מטה, וגם גלולה וגם סופרנאטנט אוחסנו לניתוח. כדאיות התא הוערכה מהסופרנאטנט לכימות אדנילט קינאז המשתחרר על ידי תאים פגומים, כפי שתואר קודם לכן17. חלבונים תאיים הופקו מכדורית התא, והפרוטאום המלא נותח על-ידי ספקטרומטריית מסות ברזולוציה גבוהה (איור 1B).
פרוטוקול זה גם מדגים שיטה חצי-אוטומטית לספירת ספרואידים (איור 3A) באמצעות תוכנת עיבוד התמונה הציבורית FIJI (Fiji Is Just ImageJ)18. יש לקחת תמונה באיכות טובה של הספרואיד לצורך הניתוח, ויש לקחת בחשבון כמה פרמטרים כאמור בסעיף 5. לאחר מכן, לאחר הכנת התמונה לניתוח, נעשה שימוש בסקריפט מאקרו (קובץ משלים 1) לספירת הספרואידים. המאקרו פועל על-ידי יצירת תיקייה בשם FIJI Spheroids counting, בתוך התיקיה שבה ממוקמות התמונות הספרואידים. בתיקיה זו, כל המידע מהניתוח נשמר; זה כולל תמונה של הספרואידים שנספרו, עם מספר תעודת זהות על כל ספרואידים. הוא כולל גם קובץ אקסל שנקרא ספירה ספרואידית. קובץ זה מכיל את אזור הפיקסלים ואת מספר הזיהוי של כל ספרואיד שנספר. הנתונים המתאימים לתמונה מנותחת אחת מוצגים בכל כרטיסיה של הקובץ. הכרטיסייה מסומנת לפי שם התמונה המנותחת. מכיוון שגודל הספרואיד יכול להיפגע על ידי גורמים רבים, כולל מספר המבנים בתוך כלי הדם וטיפול תרופתי, חשוב גם לעקוב אחר שטח הפנים שלהם (פלנימטריה). סקריפט המאקרו המוצג כאן (קובץ משלים 2) פועל על-ידי מדידת האזורים השחורים, המתאימים לכדוריות בתמונה (איור 3B). הפלט נאסף בקובץ שנקרא planimetry.xlsx, המכיל את השטח הנמדד, ההיקף והקוטר של כל ספרואיד. יש גם מדידה בשם Feret, המשמשת לחישוב הקוטר. Feret הוא הקוטר הארוך ביותר האפשרי, בעוד minFeret הוא הקצר ביותר. הקוטר הוא הממוצע של שני אלה. בתוך תיקיית הפלט, מלבד קובץ הפלנימטריה.xlsx, יש גם תמונה של הספרואידים שנמדדו.
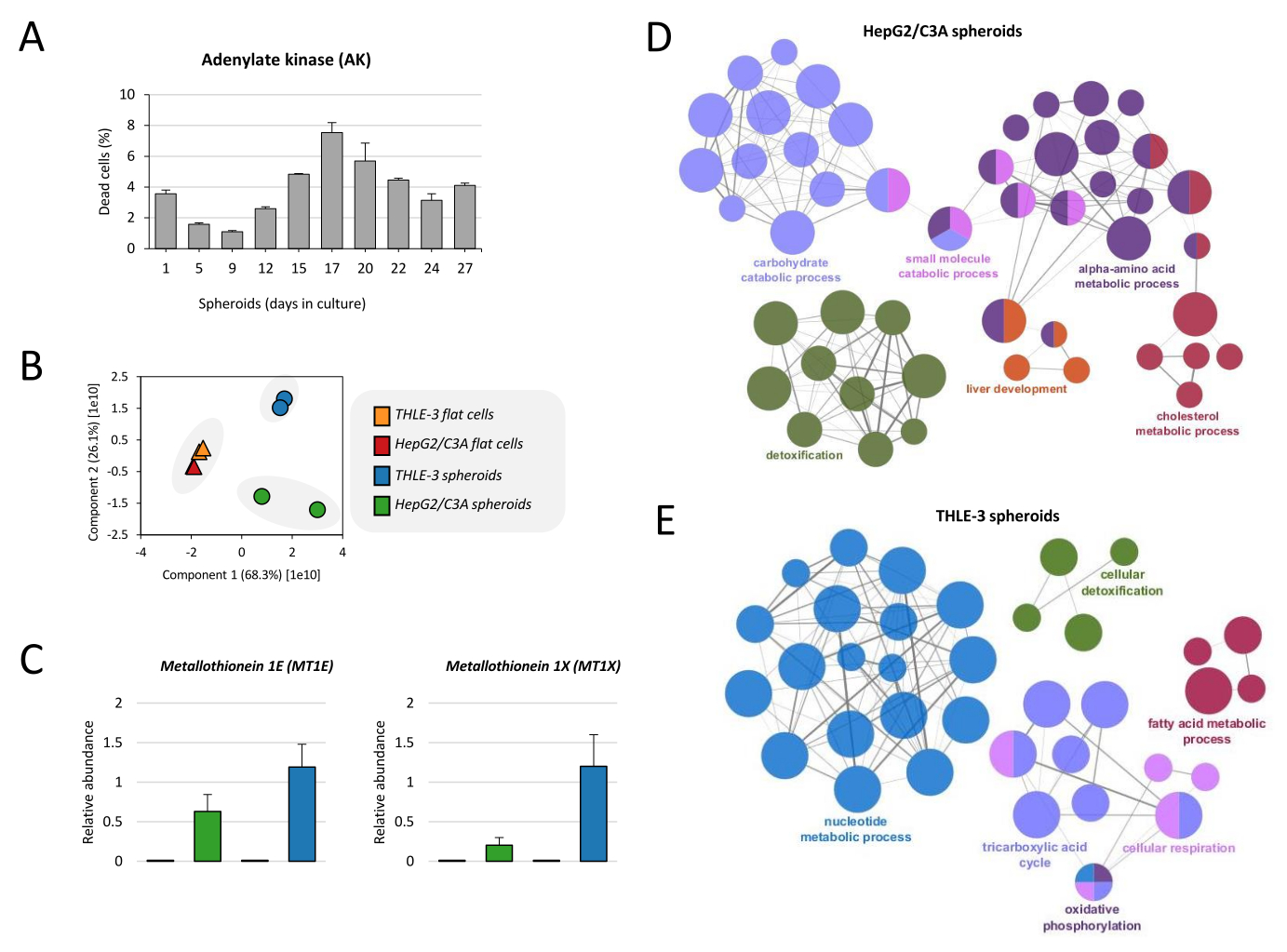
לפני שנמשיך לניתוח הפרוטאום, הוערכה הכדאיות של הספרואידים לאורך זמן התרבית. רמות AK עולות עד היום ה-17 ומגיעות לכ-7% ממקרי המוות של תאים, ואז המוות יורד לרמות מתחת ל-5% (איור 4A), בהתאם לעבודה17 שפורסמה בעבר. פרוטוקול זה מציג גם את ניתוח הפרוטאום המלא לניטור פנוטיפ התא. ראשית, הושוו הפרוטאומים של התאים השטוחים THLE-3 ו-HepG2/C3A והספרואידים. על-ידי ניתוח המרכיב העיקרי הראשון (PC1), ברור שיש הפרדה קפדנית של דגימות ספרואידים מתרביות תאים שטוחים, ונראה שהמתאם של סוג התא (THLE-3 ו-HepG2/C3A) אינו רלוונטי (איור 4B). למרות שספרואידים THLE-3 ו-HepG2/C3A אינם מתקבצים יחד, הם חולקים פרופילים דומים התואמים את תפקודי הכבד. אנו מדגימים בפרוטוקול זה את הדוגמה של מטלותיונינים, שיש להם תפקיד בניקוי רעלים ממתכות המבוצע על ידי הכבד. זיהינו בניתוח הפרוטאומיקה 2 איזופורמים המבוטאים יתר על המידה בספרואידים בהשוואה לתאים שטוחים (MT1E ו-MT1X) (איור 4C). אנו מראים גם את העשרת האונטולוגיה הגנטית (GO) של שני קווי התאים שגדלו כספרואידים. התהליך המטבולי של פחמימות, הכולל את מחזור החומצה הטריקרבוקסילית (מחזור TCA), שרשרת הובלת האלקטרונים (נשימה תאית) ומטבוליזם של פירובט, הוא מונח שכיח ומועשר הן בספרואידים HepG2/C3A והן ב-THLE-3 (איור 4D,E). ניקוי רעלים מתאים, חומצות שומן ומטבוליזם של כולסטרול הם תפקידים נוספים המועשרים בשני הספרואידים. יחד, תפקודים אלה ידועים כחיוניים לתפקוד הכבד.

איור 1: זרימת עבודה עבור תרבית ספרואידים והכנת דגימה . (A) גישה ניסיונית תלת-ממדית לתרבית תאים. תרביות תאים שטוחות במפגש הרצוי עברו טריפסינזציה ונזרעו על צלחת חיבור נמוכה במיוחד 24 בארות המכילה מיקרו-בארות, שבה התאים מתאספים בעצמם לספרואידים. לאחר 24 שעות, ספרואידים הועברו לביוריאקטור וטופחו עד שהם מוכנים לניתוח. (B) לאחר האיסוף, כדוריות היו גלולות ואוחסנו הן כדוריות והן סופרנאטנט תרבית עד לעיבוד. חלבוני היסטונים16ושאינם היסטונים חולצו, עוכלו לפפטידים ונותחו על ידי ספקטרומטריית מסות ברזולוציה גבוהה. קבצים גולמיים שהתקבלו מספקטרומטריית המסות נבדקו מול מסד הנתונים האנושי, והנתונים עובדו עוד יותר. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 2: מערכת תלת-ממדית של תרביות תאים . (A) חלקי ביוריאקטורים. הביוריאקטור מורכב מתא חילופי גזים ולחות המכיל חרוזי מים ותא תא הניתן לפתיחה עם שני תקעים להחלפת מדיה ואיסוף ספרואידים. (B) חילופי מדיה של ביוריאקטור. הביוריאקטור מלא ב -10 מ"ל של מצע צמיחה באמצעות מזרק עם מחט. (C) אפליקציית בקרת המערכת. ניתן לשלוט במהירות הסיבוב, רמת CO2, טמפרטורה, יומן אזעקה ופונקציות אחרות באמצעות יחידת הבקרה. (ד) הצבת הביוריאקטור באינקובטור התלת-ממדי. לכל ביוריאקטור יש מנוע מקושר שיכול לסובב את הביוריאקטור באיטיות. (E) ביוריאקטור בתנועה עם מהירות (סל"ד) הנשלטת על ידי טבליה (C). המהירות (סל"ד) מותאמת בהתאם לגודל הספרואידים. (F) ביוריאקטורים בתוך אינקובטור תלת-ממדי. החממה התלת-ממדית יכולה להכיל עד 6 ביוריאקטורים בשליטה אישית. התמונה באדיבות ג'ייסון טורס צילום. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 3: אפיון פנוטיפי של ספרואידים באמצעות לכידת תמונה . (A) ספירה ספרואידית חצי-אוטומטית. לאחר צילום תמונות של הספרואידים בביוריאקטור, התמונה מוכנה לניתוח בפיג'י. כל ספרואיד נספר, ולכל אחד מהם ניתן מספר תעודת זהות. נעשה שימוש במאקרו, והתוצאות מוצגות ומציגות את המזהה של הספרואיד שנספר, את התווית (שם התמונה שנותחה) ואת האזור (מספר הפיקסלים שנספרו בספרואיד). (B) קביעה פלנימטרית של שטח ספרואידי. באמצעות מאקרו נקבעים השטח, ההיקף והקוטר של ספרואיד מסוים. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 4: ניתוח פרוטאום של ספרואידים בכבד . (A) הכדאיות של ספרואידים חושבה בהתבסס על שחרור אדנילט קינאז (AK) על תרבית סופרנטנט. התוצאות הן אמצעי לנקודות נתונים כפולות ± SD. (B) ניתוח רכיבים עיקרי (PCA) בוצע כדי להשוות את הפרוטאום של תאים שטוחים THLE-3 ו- HepG2/C3A וספרואידים. (C) שפע יחסי של מטלותיונינים, שהם חלבונים המבוטאים על-ידי הכבד האנושי. הנתונים מיוצגים כאמצעים ± SEM. (D) רשת מקובצת פונקציונלית מציגה העשרת GO עבור ספרואידים HepG2/C3A ו-(E) ספרואידים THLE-3, כאשר רק התווית של המונח המשמעותי ביותר לכל קבוצה מוצגת. הרשת נבנתה באמצעות ClueGo19. גודל הצומת מייצג את המונח משמעות העשרה. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}
קובץ משלים 1: סקריפט מאקרו לספירה ספרואידית. אנא לחץ כאן כדי להוריד קובץ זה.
קובץ משלים 2: מאקרו לקביעה פלנימטרית של ספרואידים. אנא לחץ כאן כדי להוריד קובץ זה.
Discussion
הבנת הביולוגיה שמאחורי מבנים תאיים תלת-ממדיים (תלת-ממדיים) חשובה ביותר לידע מקיף יותר על תפקודם. יש עניין גובר בשימוש במודלים תלת-ממדיים לחקר ביולוגיה מורכבת וביצוע סינון רעילות. בעת טיפוח תאים בתלת ממד יש לקחת בחשבון גורמים רבים, כולל הערכה פנוטיפית של מערכת המודל. פנוטיפ מוגדר כקבוצה של מאפיינים נצפים של אורגניזם מסוים, כגון מורפולוגיה, התנהגות, תכונות פיזיולוגיות וביוכימיות20.
בפרוטוקול זה, אנו מדגימים כיצד ניתן לבצע ניסויים בפרוטאומיקה מכמה ספרואידים וניתן להשתמש בהם כדי לעקוב אחר פנוטיפ הכבד הטיפוסי. ספקטרומטריית מסות הפכה לשיטה מיושמת באופן נרחב לאפיון תאים תלת-ממדיים, ומאפשרת לחקור מגוון שאלות ביולוגיות12,16,21,22. לניתוח פרוטאום מקיף, מומלץ להשתמש לפחות ב-20 מיקרוגרם של חומר מוצא חלבוני, שממנו מוזרק 1 מיקרוגרם לספקטרומטר המסות. חשוב לציין כי הוספת פחות דגימה עלולה להוביל לאובדן רגישות, והוספת דגימה נוספת תחמיר בהדרגה את איכות הכרומטוגרפיה ובסופו של דבר תוביל לחסימת העמודה. במחקר זה, הראינו כי הספרואידים HepG2/C3A ו- THLE-3 מועשרים בחלבונים חשובים ממחזור גליקוליזה ו- TCA, שהם מסלולי כבד ספציפיים והם קריטיים לשמירה על רמות הגלוקוז בדם ולייצור אנרגיה23,24. למעשה, ניתוח ספקטרומטריית מסות מספק לא רק מידע ברמת החלבון, אלא גם מאפשר חקירה של שינויים לאחר תרגום חלבון, כפי שהוצג קודם לכן על ידי הקבוצה שלנו16.
היבט נוסף שיש לקחת בחשבון במחקרים פנוטיפיים תלת ממדיים הוא מספר וגודל הספרואידים. מלבד הפיכת הניסויים לניתנים יותר לשחזור, ספירת מספר הספרואידים וקביעת גודלם חיונית כדי לקבוע מתי לפצל את התרבית למספר ביוריאקטורים, שכן מספר המבנים התלת-ממדיים בתוך כלי יכול להשפיע על גודל הספרואידים ועל רמות הפעילות המטבולית. עם זאת, חשוב להדגיש כי מספר וגודל הספרואידים תלויים בקו התא, מספר התאים ההתחלתי, תהליך הפיצול וזמן האיסוף. פרטים על תרבית ספרואידים HepG2/C3A, כגון מספר התאים לכל ספרואידים, תכולת חלבונים וגודל כפונקציה של גיל, סופקו על ידי פיי, קורז'ניובסקה וז'סינסקי25. לניתוח מדויק ומוצלח בשיטה החצי-אוטומטית המתוארת כאן, השלב הקריטי ביותר הוא תמונה טובה של הספרואידים. לשם הפשטות, התמונה יכולה להילקח עם טלפון או טאבלט, אבל הרזולוציה שלה צריך להישמר גבוה ככל האפשר. מכיוון שתמונות ניתנות להשגה במהירות, הן מאפשרות לניסויי סינון בקנה מידה גדול לדמיין תכונות פנוטיפיות ספציפיות או לחקור תגובות לטיפול תרופתי. לכן, בשל המספר ההולך וגדל של בדיקות מבוססות תאים, פותחו במהלך 10 השנים האחרונות מספר תוכנות קוד פתוח לניתוח תמונה26. בפרוטוקול זה, אנו מתארים מערכת חצי אוטומטית המשתמשת בתוכנה FIJI18 כדי לספור ולמדוד את גודל הספרואידים. הצגנו סקריפטים (פקודות תכנות פשוטות) כדי להגדיר רצף של פעולות אלגוריתמיות שניתן להחיל על אוסף תמונות, מה שהופך את הניתוח לתהליך קל ומהיר. עם זאת, בהתאם למאפיין של הספרואידים, יש להשתמש במדידה ידנית. לדוגמה, אם הספרואידים שקופים מדי, כתב FIJI יהיה לא מדויק. אגב, אחד הקריטריונים החשובים ביותר עבור שיטה זו לעבוד הוא קומפקטיות של spheroids. מאפיין זה יתרום לניגודיות צבעים משופרת יותר בין הספרואידים לבין הרקע, דבר הכרחי כדי שהשיטה תהיה מדויקת.
לסיכום, מלבד הצגת מתודולוגיה לגידול ספרואידים בעלי יכולת שחזור גבוהה, תוארה גם מערכת חצי אוטומטית בשילוב אפיון פנוטיפי באמצעות לכידת תמונה ופרוטאומיקה. אנו מצפים שארגז כלים זה לניתוח תאים תלת-ממדיים יהפוך לחזק יותר עם תוכנת ניתוח תמונה אוטומטית מלאה וספקטרומטרים של מסות מהדור הבא.
Disclosures
הלה סדיגי פראנדסן מועסקת כמדענית מחקר בחברת CelVivo ApS, יצרנית מערכת ClinoStar. קרולין מיקלסן היא דוקטורנטית תעשייתית המועסקת ב- CelVivo ApS ומבצעת את עבודת הדוקטורט שלה ב- SDU, אודנסה, דנמרק. לכל שאר המחברים אין אינטרסים כלכליים מתחרים.
Acknowledgements
מעבדת סידולי מודה לקרן לחקר לוקמיה (מענק מחקר חוקר חדש ע"ש הוליס בראונשטיין), AFAR (פרס גרומיקס של רשת סגול), דירפילד (פרס Xseed), Relay Therapeutics, Merck ומשרד המנהל של NIH (1S10OD030286-01).
Materials
| Name | Company | Catalog Number | Comments |
| 1.5 mL microcentrifuge tubes | Bio-Rad | 2239480 | |
| 10 mL syringe | Fisher Scientific | 1481754 | Luer lock tip, graduated to 12 mL |
| 1000 µL wide bore pipet tips | Fisher Scientific | 14222703 | |
| 200 µL wide bore pipet tips | Fisher Scientific | 14222730 | |
| 96-well Orochem filter plate | Orochem | OF1100 | |
| 96-well skirted plate | Axygen | PCR-96-FS-C | |
| 96-well vacuum manifold | Millipore | MAVM0960R | |
| Ammonium bicarbonate | Sigma | A6141-25G | |
| Bronchial Epithelial Cell Growth Medium (BEGM) | Lonza | CC-3170 | |
| Cell culture grade water | Corning | 25-055-CV | |
| ClinoReactor | CelVivo | N/A | Bioreactor for 3D cell culture |
| ClinoStar incubator | CelVivo | N/A | CO2 incubator for 3D cell culture |
| DTT | Sigma | D0632-5G | |
| Dulbecco's Modified Eagle's Medium (DMEM) | Fisher Scientific | MT17205CV | |
| Elplasia 24-well round bottom ultra-low attachment plate containing microwells | Corning | 4441 | |
| Fetal Bovine Serum | Fisher Scientific | MT35010CV | |
| Formic acid | Thermo | 28905 | |
| Hank's Balanced Salt Solution (HBSS) | Fisher Scientific | MT21022CV | |
| hEGF | Corning | 354052 | |
| HERAcell vios 160i | Thermo | 51033557 | CO2 incubator for 2D cell culture |
| HPLC grade acetonitrile | Fisher Scientific | A955-4 | |
| HPLC grade methanol | Fisher Scientific | A452-1 | |
| HPLC grade water | Fisher Scientific | W5-4 | |
| Iodoacetamide | Sigma | I1149-5G | |
| L-glutamine | Fisher Scientific | MT25015CI | |
| Non-essential amino acids | Fisher Scientific | MT25025CI | |
| Oasis HLB Resin 30 µm | Waters | 186007549 | |
| Orbitrap Fusion Lumos Tribrid mass spectrometer | Thermo | IQLAAEGAAPFADBMBHQ | High resolution mass spectrometer |
| PAULA microscope | Leica | ||
| Penicillin-Streptomycin | Fisher Scientific | MT3002CI | |
| PerkinElmer Victor X2 multilabel microplate reader | PerkinElmer | ||
| pH paper | Hydrion | 93 | |
| Phosphoetanolamine | Sigma | P0503 | |
| Phosphoric acid | Fisher Scientific | A260-500 | |
| Pipette gun | Eppendorf | Z666467 (Milipore Sigma) | |
| Refrigerated centrifuge | Thermo | 75-217-420 | |
| Reprosil-Pur resin | MSWIL | R13.AQ.003 | 120 Å pore size, C18-AQ phase, 3 μM bead size |
| SDS | Bio-Rad | 1610301 | |
| Sequencing grade modified trypsin | Promega | V511A | |
| SpeedVac vacuum concentrator (96-well plates) | Thermo | 15308325 | Savant SPD1010 |
| Sterile hood | Thermo | 1375 | |
| Sterile serological pipettes | Fisher Scientific | 1367549 | |
| S-trap | Protifi | C02-micro-80 | |
| Syringe needle (18 G) | Fisher Scientific | 14817100 | 3" length, 0.05" diameter |
| Trifluoroacetic acid (TFA) | Thermo | 28904 | |
| Trypsin-EDTA | Gibco | 25300-054 | |
| Vortex | Sigma | Z258415 | |
| Water bath | Fisher Scientific | FSGPD10 |
References
- Ravi, M., Paramesh, V., Kaviya, S. R., Anuradha, E., Solomon, F. D. 3D cell culture systems: advantages and applications. Journal of Cellular Physiology. 230 (1), 16-26 (2015).
- Nirmalanandhan, V. S., Duren, A., Hendricks, P., Vielhauer, G., Sittampalam, G. S. Activity of anticancer agents in a three-dimensional cell culture model. Assay and Drug Development Technologies. 8 (5), 581-590 (2010).
- Erickson, I. E., Huang, A. H., Chung, C., Li, R. T., Burdick, J. A., Mauck, R. L. Differential maturation and structure-function relationships in mesenchymal stem cell- and chondrocyte-seeded hydrogels. Tissue Engineering Part A. 15 (5), 1041-1052 (2009).
- Vinci, M., et al. Advances in establishment and analysis of three-dimensional tumor spheroid-based functional assays for target validation and drug evaluation. BMC Biology. 10 (29), (2012).
- Liu, J., Abate, W., Xu, J., Corry, D., Kaul, B., Jackson, S. K. Three-dimensional spheroid cultures of A549 and HepG2 cells exhibit different lipopolysaccharide (LPS) receptor expression and LPS-induced cytokine response compared with monolayer cultures. Innate Immunity. 17 (3), 245-255 (2011).
- Khafaga, A. F., Mousa, S. A., Aleya, L., Abdel-Daim, M. M. Three-dimensional (3D) cell culture: a valuable step in advancing treatments for human hepatocellular carcinoma. Cancer Cell International. 22 (1), 243 (2022).
- Llovet, J. M., Burroughs, A., Bruix, J. Hepatocellular carcinoma. The Lancet. 362 (9399), 1907-1917 (2003).
- Sia, D., Llovet, J. M. Liver cancer: Translating '-omics' results into precision medicine for hepatocellular carcinoma. Nature Reviews Gastroenterology & Hepatology. 14 (10), 571-572 (2017).
- Tang, J., et al. A three-dimensional cell biology model of human hepatocellular carcinoma in vitro. Tumour Biology. 32 (3), 469-479 (2011).
- van Zijl, F., Mikulits, W. Hepatospheres: Three dimensional cell cultures resemble physiological conditions of the liver. World Journal of Hepatology. 2 (1), 1-7 (2010).
- Chaicharoenaudomrung, N., Kunhorm, P., Noisa, P. Three-dimensional cell culture systems as an in vitro platform for cancer and stem cell modeling. World Journal of Stem Cells. 11 (12), 1065-1083 (2019).
- Wrzesinski, K., Fey, S. J. Metabolic reprogramming and the recovery of physiological functionality in 3D cultures in micro-bioreactors. Bioengineering(Basel, Switzerland). 5 (1), 22 (2018).
- Breslin, S., O'Driscoll, L. Three-dimensional cell culture: The missing link in drug discovery. Drug Discovery Today. 18 (5-6), 240-249 (2013).
- Kapalczynska, M., et al. 2D and 3D cell cultures - a comparison of different types of cancer cell cultures. Archives of Medical Science: AMS. 14 (4), 910-919 (2018).
- Wrzesinski, K., Frandsen, H. S., Calitz, C., Gouws, C., Korzeniowska, B., Fey, S. J. Clinostat 3D cell culture: Protocols for the preparation and functional analysis of highly reproducible, large, uniform spheroids and organoids. Methods in Molecular Biology. 2273, 17-62 (2021).
- Joseph-Chowdhury, J. N., et al. Global level quantification of histone post-translational modifications in a 3D cell culture model of hepatic tissue. Journal of Visualized Experiments: JoVE. 183, 63606 (2022).
- Wrzesinski, K., Fey, S. J. After trypsinisation, 3D spheroids of C3A hepatocytes need 18 days to re-establish similar levels of key physiological functions to those seen in the liver. Toxicology Research. 2, 123-135 (2013).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Bindea, G., et al. ClueGO: a Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics. 25 (8), 1091-1093 (2009).
- Houle, D., Govindaraju, D. R., Omholt, S. Phenomics: The next challenge. Nature Reviews. Genetics. 11 (12), 855-866 (2010).
- Gonneaud, A., Asselin, C., Boudreau, F., Boisvert, F. M. Phenotypic analysis of organoids by proteomics. Proteomics. 17 (20), (2017).
- Avelino, T. M., et al. Mass spectrometry-based proteomics of 3D cell culture: A useful tool to validate culture of spheroids and organoids. SLAS Discovery. 27 (3), 167-174 (2022).
- Chiang, J., McManus, L. M., MitchellIn, R. N. . Liver physiology: Metabolism and Detoxification. Pathobiology of Human Disease. , 1770-1782 (2014).
- Begriche, K., Massart, J., Robin, M. A., Borgne-Sanchez, A., Fromenty, B. Drug-induced toxicity on mitochondria and lipid metabolism: Mechanistic diversity and deleterious consequences for the liver. Journal of Hepatology. 54 (4), 773-794 (2011).
- Fey, S. J., Korzeniowska, B., Wrzesinski, K. Response to and recovery from treatment in human liver-mimetic clinostat spheroids: a model for assessing repeated-dose drug toxicity. Toxicology Research. 9 (4), 379-389 (2020).
- Smith, K., et al. Phenotypic image analysis software tools for exploring and understanding big image data from cell-based assays. Cell Systems. 6 (6), 636-653 (2018).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved