
Method Article
Osservazione in tempo reale della DNA Strand Exchange reazione mediata da Rad51
In questo articolo
Riepilogo
Fluorescenza risonanza energia basati su trasferimento osservazione in tempo reale sistemi del DNA strand reazione di scambio mediato da Rad51 sono stati sviluppati. Utilizzando i protocolli presentati qui, siamo in grado di rilevare la formazione di intermedi di reazione e loro conversione in prodotti, mentre si analizza anche la cinetica enzimatica della reazione.
Abstract
La reazione di scambio strand DNA mediata da Rad51 è un passo fondamentale di ricombinazione omologa. In questa reazione, Rad51 costituisce un filamento nucleoproteina sul DNA a singola elica (ssDNA) e cattura di DNA a doppia elica (dsDNA) non specifico per interrogare i esso per una sequenza omologa. Dopo aver incontrato omologia, Rad51 catalizza DNA strand exchange per mediare abbinamento del ssDNA con filamento complementare del dsDNA. Questa reazione è altamente regolamentata da numerose accessary proteine in vivo. Sebbene analisi biochimiche convenzionali sono state impiegate con successo per esaminare il ruolo di tale proteina accessoria in vitro, l'analisi cinetica di formazione intermedia e la sua progressione in un prodotto finale ha dimostrato impegnativo a causa della natura instabile e transitorio degli intermedi di reazione. Per osservare queste fasi di reazione direttamente in soluzione, trasferimento di energia di risonanza di fluorescenza (FRET)-base di osservazione in tempo reale, sistemi di questa reazione sono stati stabiliti. Analisi cinetica di osservazioni in tempo reale mostra che la reazione di scambio strand DNA mediata da Rad51 obbedisce a un modello di reazione di tre fasi che coinvolgono la formazione di un DNA di tre capi intermedio, maturazione di questo intermedio e il rilascio di ssDNA da l'intermedio maturo. Il complesso di Swi5-Sfr1, una proteina accessary conservato negli eucarioti, migliora fortemente la seconda e la terza procedura di questa reazione. Le analisi basato su FRET presentate qui ci permettono di scoprire i meccanismi molecolari attraverso cui ricombinazione accessary proteine stimolano l'attività di cambio del filo di DNA di Rad51. L'obiettivo primario del presente protocollo è per migliorare il repertorio delle tecniche disponibili per i ricercatori nel campo della ricombinazione omologa, in particolare quelli che lavorano con le proteine da specie diverse da Schizosaccharomyces pombe, affinché la conservazione evolutiva dei risultati presentati nel presente documento può essere determinato.
Introduzione
Ricombinazione omologa (HR) facilita il rimescolamento dell'informazione genetica tra due diverse molecole di DNA. HR è essenziale per due fenomeni biologici fondamentali: la generazione della diversità genetica durante la gametogenesi1 e la riparazione di DNA double-strand breaks (DSBs)2 durante la mitosi. DSBs sono la forma più grave di danno del DNA e costituiscono una rottura nel cromosoma. Una riparazione errata di DSBs può causare ampie riorganizzazioni cromosomiche e l'instabilità genomica, che sono entrambi marchi di garanzia di cancro3.
La reazione di scambio del filo del DNA è la fase centrale di HR. La proteina Rad51, che è un membro della famiglia RecA-tipo altamente conservata di ricombinasi, è la proteina chiave che catalizza questa reazione in eucarioti4,5. In questa reazione, Rad51 si lega al DNA di singola elica (ssDNA) generato dall'elaborazione nucleolitico della fine DSB e forme una nucleoproteina elicoidale complessa definito il filamento presinaptico. Questo filamento cattura intatto DNA a doppia elica (dsDNA) non specifico per la ricerca di una sequenza omologa. Quando il filamento si trova una sequenza omologa, si forma un'intermedio contenente tre filamenti DNA di reazione e il filamento di Rad51 media strand scambio all'interno di questa struttura6,7,8. Per realizzare questa reazione in modo efficiente, Rad51 richiede diversi tipi di proteine accessorie come BRCA1 e BRCA2, prodotti di seno cancro suscettibilità geni9,10.
Intesa come accessori fattori regolano Rad51 è un passaggio integrale nello scoprire le cause di instabilità genomica durante il tumorigenesis. Anche se gran parte della ricerca è interessato con gli effetti di questi fattori sulla formazione dei filamenti presinaptici e stabilità11,12,13,14,15,16, la contributo di questi fattori di formazione di tre capi intermedi e la relativa elaborazione nel prodotto finale non è ancora chiaro. Osservando questi passaggi di reazione tramite esperimenti biochimici convenzionali è molto difficile perché l'intermedio di tre capi è instabile e soggetta al collasso da manipolazioni sperimentali comuni quali deproteinizzazione dei campioni o elettroforesi.
Per ovviare a questo problema, abbiamo adattato due sistemi di osservazione in tempo reale precedentemente sviluppata della reazione DNA strand exchange utilizzando il trasferimento di energia di risonanza di fluorescenza (FRET): il DNA strand abbinamento e DNA strand cilindrata saggi17, 18 (Figura 1). Nel filo del DNA abbinamento dosaggio, Rad51 forme un filamento presinaptico con fluorescina amidite (FAM) - etichettato ssDNA e quindi omologa carboxy-x-rodamina (ROX) - etichettato dsDNA è aggiunto per avviare la reazione di scambio del filo. Quando il filamento cattura il dsDNA ROX-etichettati e costituisce l'intermedio di tre capi, i due fluorofori mai avvicinare e l'emissione di fluorescenza di FAM è placata da ROX (Figura 1A). Nell'analisi di spostamento del filo del DNA, un filamento presinaptico formato il ssDNA senza etichetta viene incubato con FAM e ROX dsDNA double-etichettati. Quando cambio strand è completato e il FAM etichettato ssDNA viene rilasciato da tre-strand intermedio, l'emissione di FAM aumenta perché FAM non è più in prossimità di ROX (Figura 1B). Queste analisi ci permettono di osservare la formazione di tre capi intermedi e loro trasformazione in prodotti finali in tempo reale senza alcun disturbo per la reazione.
Utilizzando questo sistema di osservazione in tempo reale, abbiamo trovato che la reazione di scambio strand DNA mediata da Rad51 procede in tre fasi tra cui la formazione della prima reazione intermedia (C1), transitioning di primo intermedio un intermedio secondo (C2) e il rilascio di ssDNA da C219. Abbiamo anche trovato quel lievito di fissione (S. pombe) Swi5-Sfr1, che è un evolutivamente conservati Rad51 proteina accessoria complesso13,16,20,21,22, stimola la transizione di C1-C2 e il rilascio di ssDNA da C2 in un modo che dipende di idrolisi dell'ATP da Rad5119.
Se questi risultati sono evolutivamente conservati rimane sconosciuto. Questo protocollo viene fornito con la speranza che i ricercatori nel campo delle risorse umane, soprattutto coloro che lavorano con le proteine da organismi diversi da S. pombe, possono applicare queste tecniche per determinare la misura in cui il meccanismo molecolare di Rad51-driven filo cambio è conservata. Inoltre, queste tecniche si sono dimostrate altamente efficaci nel determinare il ruolo di S. pombe Swi5-Sfr1. Così, è una previsione razionale che queste tecniche sarà preziose nello scoprire i ruoli precisi di altri fattori accessori di HR.
Protocollo
1. preparazione di substrati di DNA e proteine
- Purificare proteine S. pombe Rad51 e Swi5-Sfr1 ad omogeneità (giudicati macchiando di Coomassie), come precedentemente segnalato13,21.
- Preparare i substrati di DNA del oligonucleotide elencati nella tabella 118.
Nota: I oligonucleotides sono stati acquistati (Vedi Tabella materiali) e sintetizzati in grado HPLC. Per il filamento di DNA abbinamento reazione, oligonucleotidi 16FA(-), 16A (-) _40bp e 16AR (+) _40bp sono necessari. Per l'analisi del DNA di cilindrata, oligonucleotidi 16A(-), 16FA (-) _40bp e 16AR (+) _40bp sono obbligatori (Figura 1 e tabella 1). Tutte le concentrazioni di DNA in questo protocollo si riferiscono per frammentare le concentrazioni in contrasto con le concentrazioni del nucleotide. - Per formare il donatore dsDNA, mescolare quantità equimolari di fili complementari in un tubo con pareti sottili di PCR con ricottura tampone (10 mM Tris-HCl pH 7.5, NaCl 100 mM, 10 mM MgCl2), garantendo un volume totale maggiore di 20 µ l. eseguire questa miscelazione su un ripiano metallico prechilled ghiaccio (tra 2 ° C e 4 ° C).
Nota: La combinazione di oligonucleotidi per il dosaggio di accoppiamento sono 16A (-) _40bp e 16AR (+) _40bp. Per il dosaggio di cilindrata, tempri oligonucleotidi 16FA (-) _40bp e 16AR (+) _40bp. - Riscaldare la miscela ricottura a 90 ° C per 5 minuti e raffreddare oltre 3 h a 30 ° C, utilizzando una macchina PCR. Conservare il DNA ricotto a-20 ° C.
2. DNA Strand Pairing e saggi di cilindrata
- Eseguire il filo del DNA, analisi di accoppiamento.
- Preparare 1,6 mL di tampone di reazione A (30 mM HEPES-KOH a pH 7.5, 1 mM dithiothreitol [DTT], 15 mM MgCl2, 0,25 mM ATP, albumina di siero bovino 0,1 mg/mL [BSA] e 0,0075% polyoxyethylenesorbitan monolaurato) contenente 36 nM 16FA(-) in una micro-centrifuga 2,0 mL tubo di plastica (polipropilene) e pre-Incubare a 37 ° C per 5 min.
- Per formare filamenti Rad51-ssDNA, aggiungere la proteina Rad51 ad una concentrazione finale di 1,5 µM nel buffer di reazione pre-incubate e incubare a 37 ° C per 5 min.
- Aggiungi Swi5-Sfr1 proteina alla miscela per una concentrazione finale di 0,15 µM e incubare a 37 ° C per ulteriori 5 minuti.
- Prendere 1,5 mL della miscela e trasferirlo in una cuvetta di quarzo 1.0 x 1.0 cm contenente un agitatore magnetico e impostare la cuvetta in una spettrofluorimetro. Configurare il controller di temperatura peltier dello spettrofotometro a 37 ° C e impostare l'agitatore magnetico a 450 giri/min per assicurare la rapida miscelazione del campione iniettato.
- Iniziare a misurare l'emissione di fluorescenza di FAM a 525 nm (larghezza di banda: 20 nm) al momento di eccitazione a 493 nm (larghezza di banda: 1 nm). Raccogliere dati ogni secondo.
- Dopo aver avviato la misurazione per 100 s, iniettare ROX-labeled donatore dsDNA ad una concentrazione finale di 36 nM nella miscela con una siringa e misura il cambiamento in emissione a intervalli di 1 s per ulteriori 30 minuti.
- Eseguire analisi del DNA strand displacement.
- Preparare 1,6 mL di tampone di reazione A contenente 36 nM 16A(-) in plastica micro-centrifuga 2,0 mL tubo e pre-Incubare a 37 ° C per 5 min.
- Formano filamenti Rad51-ssDNA in presenza di Swi5-Sfr1 a 37 ° C, come descritto nei passaggi 2.1.2. e 2.1.3.
- Prendere 1,5 mL della miscela e trasferirlo nella cuvetta di quarzo contenente un agitatore magnetico e impostare la cuvetta nello spettrofotometro, come descritto al punto 2.1.4.
- Iniziare a misurare l'emissione di fluorescenza e dopo 100 s, iniettare FAM - e ROX-labeled donatore dsDNA come descritto al punto 2.1.6. Misurare la variazione nell'emissione di fluorescenza a intervalli di 1 s per ulteriori 30 minuti.
3. analisi dei dati sperimentali dall'accoppiamento e le analisi di spostamento
- Stimare la massima efficienza FRET.
- Preparare 16FA(-) ricotto con 16AR (+) _40bp e 16FA _40bp (-) ricotto con 16AR (+) _40bp utilizzando la stessa procedura descritta ai punti 1.3 e 1.4.
- Preparare 130 μL di tampone di reazione A contenente 36 nM di entrambi 16FA(-), 16FA(-) ricotto con 16AR (+) _40bp, 16FA (-) _40bp ricotto con 16AR (+) _40bp o 16FA _40bp (-) in una cuvetta di quarzo di 0,2 x 1.0 cm.
- Impostare la cuvetta nello spettrofluorimetro e incubare a 37 ° C per 5 min.
- Misurare spettri di fluorescenza da 500 a 600 nm eccitazione a 493 nm.
- Per testare l'effetto di Rad51 su emissioni di FAM e tempra di FAM da ROX, aggiungere Rad51 ad una concentrazione finale di 1,5 μM alla miscela e incubare a 37 ° C per 5 min.
- Misurare spettri di fluorescenza da 500 a 600 nm eccitazione a 493 nm.
- Calcolare la massima efficienza FRET (Emassimo) utilizzando l'equazione descritta di seguito:
Emassima = (intensità di fluorescenza a 525 nm di dsDNA etichettato con FAM e ROX) / (intensità di fluorescenza a 525 nm di FAM etichettato ssDNA)
- Analizzare dati sperimentali del dosaggio di cilindrata.
- Per convertire il cambiamento in fluorescenza osservata nell'analisi della cilindrata per la variazione della quantità di prodotto finale, normalizzare raw dati sperimentali ottenuti con questo test usando l'equazione descritta di seguito, dove Fcrudo è l'intensità di fluorescenza da dati grezzi e Fnormalizzato è la variazione di fluorescenza calcolato dalla seguente equazione.
Fnormalizzato = ([Fcrudo al tempo x]-[Fcrudo al tempo 0]) / (([Fcrudo al tempo 0] / Emassimo)-[Fcrudo al tempo 0])
Fcrudo al tempo 0 è la fluorescenza media controllata per i primi 5 s dopo il tempo morto (cioè, il tempo necessario per la miscelazione dopo un injectant è stato introdotto nella cuvetta). - Per escludere gli effetti di photobleaching e spostamento spontaneo, sottrarre Fnormalizzato senza proteine da Fnormalizzato del campione per ottenere FD, che è la variazione della quantità di prodotto finale in questo test.
FD = [Fnormalizzato del campione] - [Fnormalizzato senza proteine]
- Per convertire il cambiamento in fluorescenza osservata nell'analisi della cilindrata per la variazione della quantità di prodotto finale, normalizzare raw dati sperimentali ottenuti con questo test usando l'equazione descritta di seguito, dove Fcrudo è l'intensità di fluorescenza da dati grezzi e Fnormalizzato è la variazione di fluorescenza calcolato dalla seguente equazione.
- Analizzare dati sperimentali del dosaggio di accoppiamento.
- Normalizzare raw dati sperimentali ottenuti con il test di accoppiamento usando l'equazione descritta di seguito, dove Fcrudo è l'intensità di fluorescenza dai dati grezzi e Fnormalizzato è la variazione di fluorescenza calcolato dalla seguente equazione.
Fnormalizzato = (Fcrudo al tempo x) / (Fcrudo al tempo 0)
Fcrudo al tempo 0 è la fluorescenza media controllata per gli ultimi 20 s prima di iniziare la reazione iniettando il substrato di dsDNA. - Per convertire il cambiamento in fluorescenza in cambiamento nella quantità di substrato ed escludere gli effetti di photobleaching e l'associazione spontanea, normalizzare Fnormalizzato del campione usando l'equazione descritta di seguito, dove FP è il cambiamento nella quantità di substrato in questo test.
FP = 1 - (([Fnormalizzato senza proteine] - [Fnormalizzato del campione]) / [1-Emassimo]) - Per esaminare la cinetica di reazione di scambio di DNA strand, eseguire l'analisi di regressione non lineare di ultima-square della reazione abbinamento utilizzando il programma di analisi23 (Vedi Tabella materiali).
- Preparare un file in formato. txt contenente i dati di corso di tempo di FP.
- Avviare il programma e incollare lo script nel File di codice supplementare in una finestra del programma:
- Avviare l'analisi di regressione non lineare di ultima-square. I risultati di questa analisi verranno visualizzati nella stessa finestra.
- Normalizzare raw dati sperimentali ottenuti con il test di accoppiamento usando l'equazione descritta di seguito, dove Fcrudo è l'intensità di fluorescenza dai dati grezzi e Fnormalizzato è la variazione di fluorescenza calcolato dalla seguente equazione.
Risultati
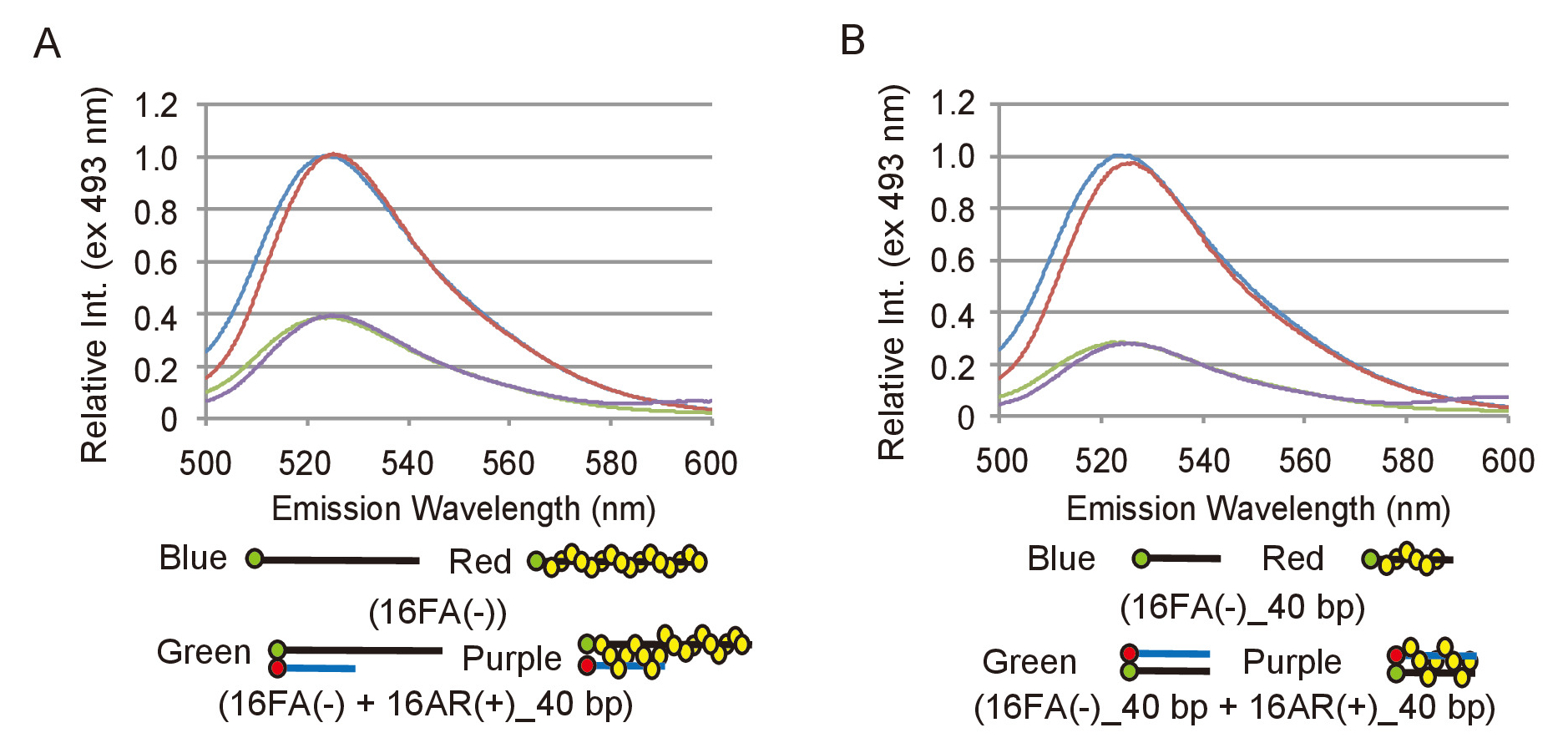
Al fine di analizzare efficacemente i dati sperimentali da saggi di abbinamento e lo spostamento, è necessario definire come un cambiamento nella emissione di fluorescenza di FAM corrisponde a una conversione di substrati di DNA nei prodotti. Per raggiungere questo obiettivo, la gamma relativa dell'intensità della fluorescenza deve essere determinata. Per il dosaggio di accoppiamento, l'emissione di fluorescenza di 16FA(-), che corrisponde al substrato di ssDNA, viene confrontato con l'emissione di 16FA(-) ricotto con 16AR (+) _40bp, che corrisponde ai prodotti finali di questa reazione (Figura 2A). Ciò equivale alla massima efficienza FRET e quindi la massima riduzione dell'intensità di fluorescenza che sarebbe essere previsto se tutti ssDNA substrato è stato convertito in prodotto dsDNA. Per il dosaggio di cilindrata, l'emissione di 16FA (-) _40bp ricotto con 16AR (+) _40bp, che corrisponde al substrato, viene confrontato con l'emissione di 16FA (-) _40bp, che corrisponde al prodotto finale (Figura 2B). In questo caso, il valore massimo dell'intensità di fluorescenza di FAM trasmette uno scenario in cui tutto il substrato di dsDNA è convertito in prodotto di ssDNA. S. pombe Rad51 non ha colpito l'emissione di fluorescenza di FAM o tempra efficienza di FAM da ROX in entrambe le analisi (Figura 2). La massima efficienza FRET deve essere rimisurata con ogni nuova preparazione di oligonucleotidi come dipende l'efficienza d'etichettatura di oligonucleotidi.
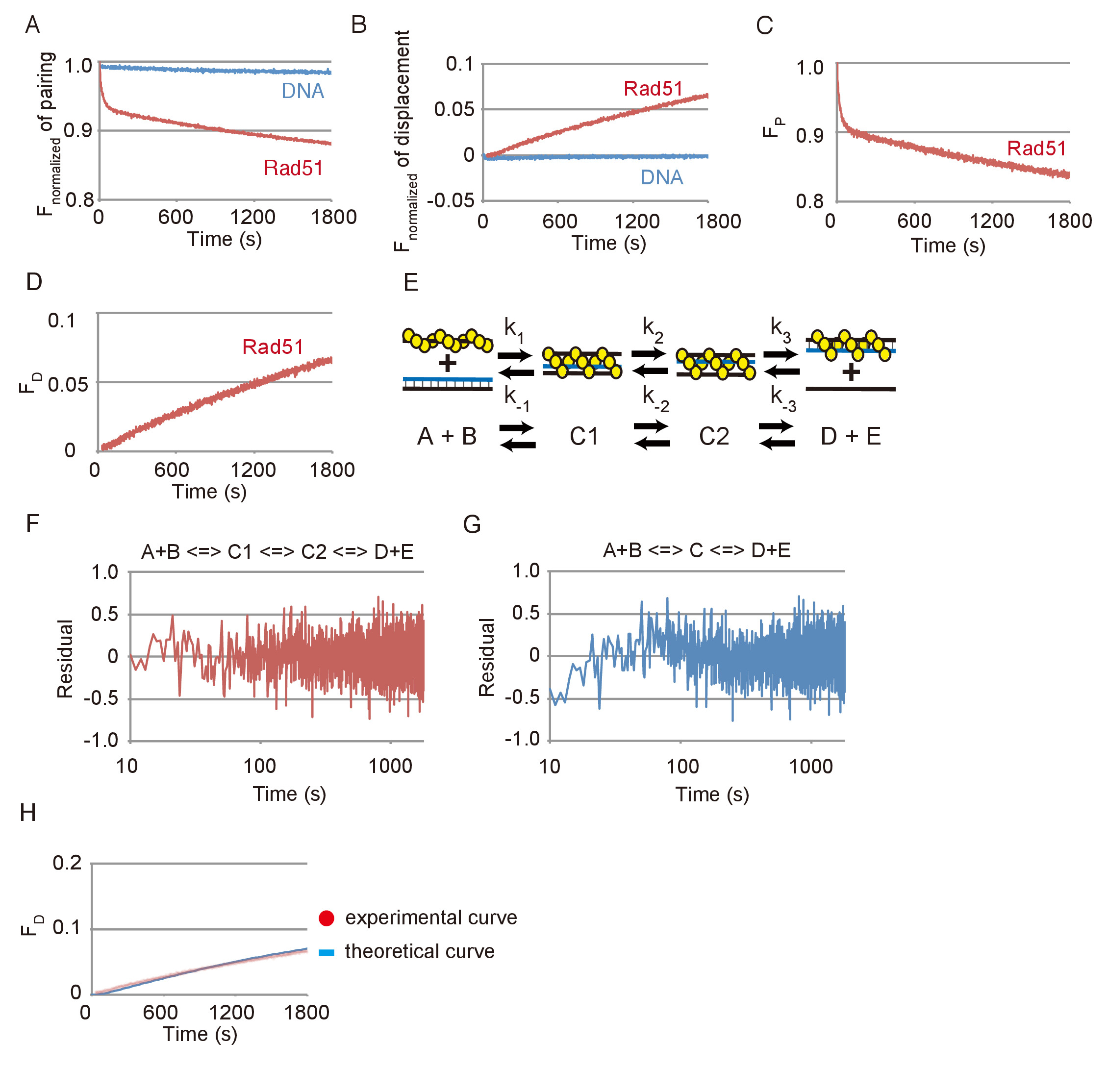
Dati rappresentativi delle reazioni di accoppiamento e spostamento strand DNA sono mostrati nella Figura 3. Gli effetti delle reazioni spontanee tra substrato DNAs e photobleaching erano piccoli in entrambe le analisi, come rivelato dai cambiamenti trascurabili visti l'emissione di FAM senza rispetto ai sostanziali cambiamenti veduti con Rad51 Rad51 (Figura 3A e Figura 3B). Sulla base dei dati mostrati in Figura 3A o 3B di figura, la variazione di fluorescenza è stata convertita nel cambiamento nella quantità di substrato (FP) o prodotto finale (FD), rispettivamente, utilizzando le equazioni descritte nei passaggi 3.2.2 o 3.3.2 ( Figura 3 e Figura 3D).
La reazione di accoppiamento è stata simulata utilizzando un modello di reazione di tre fasi sequenziali, consistendo della formazione del primo tre capi intermedio (C1), transitioning di primo intermedio il secondo intermedio (C1-C2) e il rilascio di ssDNA dal secondo intermedio per formare i due prodotti (D + E) (Figura 3E). Per verificare se la simulazione utilizzando un modello di reazione di tre fasi sequenziali fit i dati sperimentali, residui tra dati sperimentali del filo del DNA abbinamento dosaggio e una curva teorica ottenuti dalla simulazione sono stati calcolati (Figura 3F). In più, residui tra la reazione di accoppiamento e una curva teorica generata utilizzando un modello di reazione in due fasi sequenziali erano inoltre calcolato (Figura 3). I residui per la reazione di accoppiamento e il modello in due passaggi mostrano una deviazione sistematica nella fase iniziale, mentre i residui per la reazione di accoppiamento e il modello a tre fasi non mostrano tale scarto. Questo indica che il modello a tre fasi è una misura migliore rispetto al modello in due fasi per simulare la reazione di accoppiamento.
Per verificare se il modello a tre fasi è coerenza con la reazione di spostamento che rileva il passo fine dello scambio di DNA strand, abbiamo generato una curva teorica della reazione di spostamento utilizzando i parametri cinetici ottenuti dalla simulazione dell'accoppiamento reazione mostrata in Figura 3 e confrontato con i dati sperimentali della reazione spostamento illustrato nella Figura 3D (Figura 3 H). La curva teorica adatti i dati sperimentali del dosaggio cilindrata. Da questi risultati, concludiamo che la simulazione utilizzando il modello a tre fasi è in grado di valutare ragionevolmente la reazione di scambio strand DNA mediata da Rad51.
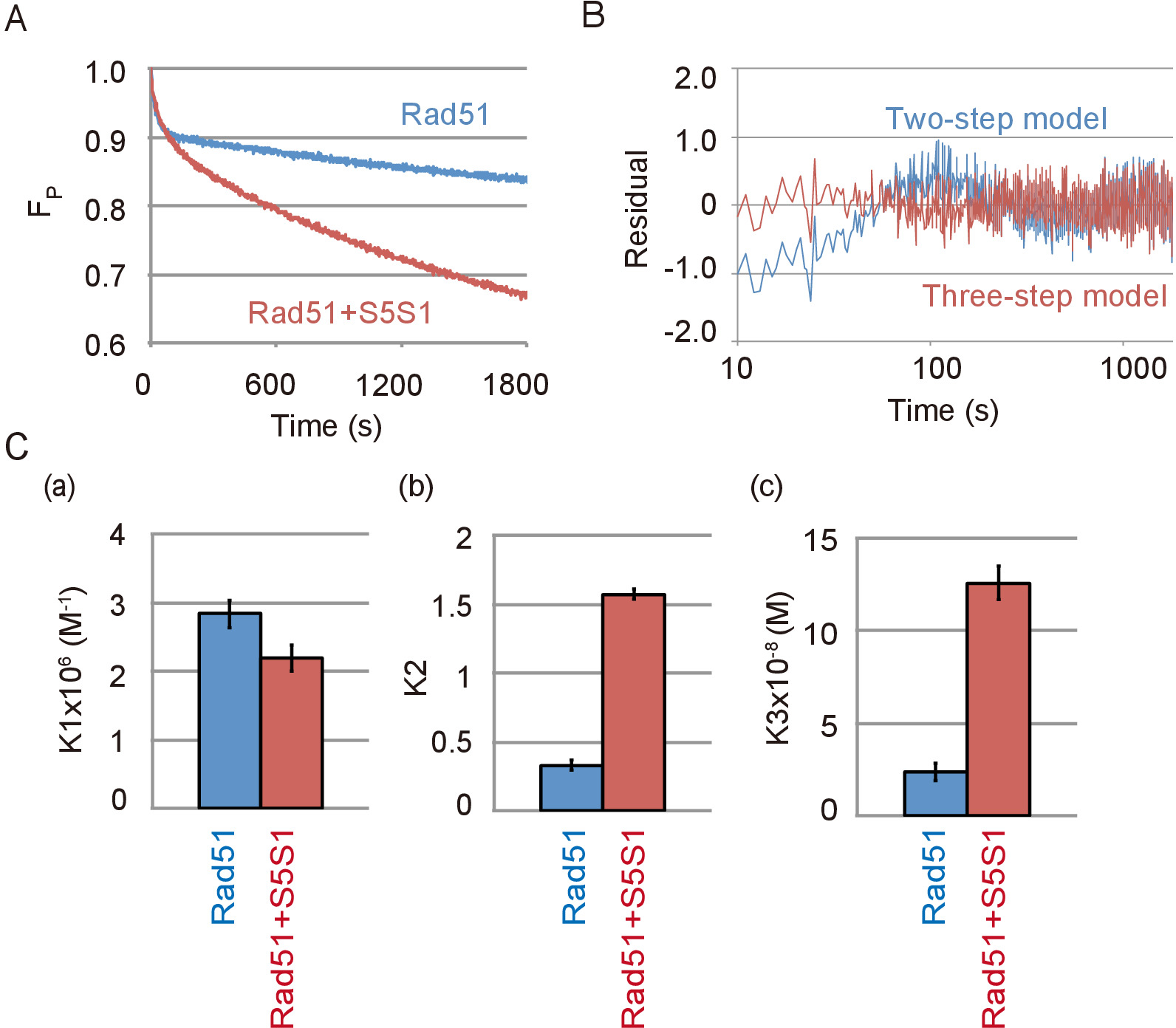
Dati rappresentativi del filo del DNA contenente Rad51 e il complesso Swi5-Sfr1, una proteina accessary di Rad51, reazione di accoppiamento sono mostrati in Figura 4A. Il complesso di Swi5-Sfr1 fortemente stimolato l'attività abbinamento di Rad51. Come si è visto in assenza di Swi5-Sfr1, la reazione di accoppiamento meglio si adatta al modello di tre-passo rispetto al modello in due fasi in presenza di Swi5-Sfr1 (Figura 4B). Attraverso la simulazione della reazione utilizzando il modello a tre fasi, costanti di equilibrio di reazione di ogni passaggio di reazione con o senza Swi5-Sfr1 sono state calcolate. Le costanti di equilibrio di reazione indicato che il complesso Swi5-Sfr1 non stimolare il primo passo di reazione (Figura 4, Pannello di un), in cui un intermedio di tre capi è formato, ma stimola fortemente la transizione C1-C2 ( Figura 4, Pannello b) e il rilascio di ssDNA dall'intermedio di C2 (Figura 4, Pannello c).

Figura 1: disegno sperimentale del DNA strand saggi abbinamento e cilindrata. Schemi di DNA strand accoppiamento (A) e saggi di spostamento (B). Cerchi gialli rappresentano Rad51 monomeri. Green circles contenente "F" e della fluorescina amidite (FAM) e carbossi-X-rodamina (ROX), rappresentano cerchi rossi contenenti "R" rispettivamente. Filamenti di DNA di nero sono identici in sequenza e complementare al blu di filamenti di DNA. Sottili linee nere con le punte delle frecce scegliere il nome di ogni oligonucleotide, come illustrato nella tabella 1. Questa figura è stata adattata da Ito et al. 19 e modificate. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: misure della FRET massima efficienza dei saggi pairing e cilindrata. (A) confronto di spettri di fluorescenza tra il substrato di ssDNA, 16FA(-) e il prodotto di dsDNA, 16FA(-) accoppiato con 16AR (+) _40bp, del dosaggio abbinamento. Linee blu e rosso rappresentano gli spettri di fluorescenza del substrato senza e con Rad51, rispettivamente. Linee verdi e viola mostrano gli spettri di fluorescenza del prodotto finale senza e con Rad51, rispettivamente. (B) confronto di spettri di fluorescenza tra il substrato di dsDNA, 16FA (-) _40bp accoppiato con 16AR (+) _40bp e prodotto di ssDNA, 16FA _40bp (-), del dosaggio cilindrata. Linee blu e rosso rappresentano gli spettri di fluorescenza del prodotto finale senza e con Rad51, rispettivamente. Linee verdi e viola mostrano gli spettri di fluorescenza del substrato senza e con Rad51, rispettivamente. Questa figura è adattata da Ito et al. 19 e modificate. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: reazioni di accoppiamento e spostamento strand DNA mediata da Rad51. (A) corso di tempo della fluorescenza normalizzata della reazione abbinamento con o senza Rad51. (B) corso di tempo della fluorescenza normalizzata della reazione di spostamento con o senza Rad51. (C) Corso di tempo del cambiamento nella quantità di substrato nella reazione di accoppiamento con Rad51. (D) corso di tempo del cambiamento nella quantità di substrato nella reazione di spostamento con Rad51. (E), A schema del modello di reazione di tre fasi sequenziali. A e B corrispondono al dsDNA presinaptico di filamento e donatore. C1 corrisponde al primo intermedio tre capi (immaturo). C2 corrisponde alla seconda (maturo) tre capi intermedi. D ed E corrispondono a un eteroduplex e ssDNA rilasciato da C2. (F e G) residui tra dati sperimentali del filo del DNA abbinamento dosaggio e una curva teorica ottenuta mediante simulazione utilizzando il modello in due fasi (G) o tre fasi (F). (H) punti rossi indicano i dati sperimentali dalla reazione di spostamento con Rad51 mostrato nel pannello blu D. linea indica la curva teorica dei prodotti finali. La curva teorica è stata generata da simulazione utilizzando le costanti di tasso di reazione ottenute con il test di abbinamento mostrato nel pannello C. Questa figura è adattata da Ito et al. 19 e modificate. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Swi5-Sfr1 stimola il secondo e terzi passaggi del DNA strand exchange reazione. (A) corso di tempo del cambiamento nella quantità di substrato nella reazione di accoppiamento con o senza Swi5-Sfr1 (S5S1). (B) residui tra dati sperimentali dell'accoppiamento strand DNA assay con Swi5-Sfr1 e una curva teorica ottenuta mediante simulazione utilizzando il modello di tre-passo (linea rossa) o in due fasi (linea blu). (C) l'abbinamento reazione mostrata in Figura 4A è stato simulato dal modello a tre fasi usando il programma di analisi23 (Vedi Tabella materiali). Le costanti di equilibrio di reazione di ogni passo di reazione, K1 (a), (b) K2 e K3 (c), sono stati ottenuti dalla simulazione. Questa figura è adattata da Ito et al. 19 e modificate. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}
| Oligonucleotidi per filo del DNA, analisi di accoppiamento | ||||
| 16FA(-) | 5'-[FAM] - AAATGAACATAAAGTAAATAAGTATAAGGATAATACA AAA AAATAAGTAAATGAATAAACATAGAAAATAAAGTAAAGGATAT -3 ' | |||
| 16a (-) _40bp | 5'-AAATGAACATAAAGTAAATAAGTATAAGGATAATACAAAA-3' | |||
| 16AR (+) _40bp | 5 '- TTTTGTATTATCCTTATACTTATTTACTTTATGTTCATTT-[ROX] -3' | |||
| Oligonucleotidi per dosaggio di DNA strand displacement | ||||
| 16A(-) | 5'-AAATGAACATAAAGTAAATAAGTATAAGGATAATACAAAATA AAA AGTAAATGAATAAACATAGAAAATAAAGTAAAGGATAT -3 ' | |||
| 16FA (-) _40bp | 5'-[FAM] - AAATGAACATAAAGTAAATAAGTATAAGGATAATACAAAA-3' | |||
| 16AR (+) _40bp | 5 '- TTTTGTATTATCCTTATACTTATTTACTTTATGTTCATTT-[ROX] -3' | |||
Tabella 1: un elenco di oligonucleotidi utilizzati nelle analisi DNA strand pairing e cilindrata. Ove applicabile, le posizioni di fluorofori (fluorescina amidite, FAM; carbossi-x-rodamina, ROX) sono indicate quadrato parentesi.
Discussione
Qui, abbiamo descritto un protocollo dettagliato che utilizza FRET per misurare Rad51-driven DNA strand scambio in tempo reale. Importante, queste misure consentono per la determinazione della cinetica di reazione. Mentre le descrizioni di cui sopra sono sufficienti per riprodurre i nostri risultati pubblicati, ci sono diversi punti critici che verranno descritti in questa sezione. Inoltre, i vantaggi e svantaggi di metodologie basate su FRET per studiare cambio strand DNA saranno discusse, con l'applicazione di tali tecniche per studiare altri aspetti del metabolismo del DNA.
Come con tutte le ricostituzioni biochimici, è fondamentale assicurare che tutti i substrati di reazione sono di elevata purezza. È negligente presumere l'assenza di contaminanti attività basata esclusivamente sulla purezza apparente di una preparazione di proteina giudicata dalla macchiatura Coomassie. In particolare, la presenza di tracce di nucleasi o elicasi drasticamente possa influenzare i risultati dei saggi di accoppiamento e spostamento. Pertanto, consigliamo vivamente test per tali attività ogni volta che un nuovo lotto di proteina è purificato. Inoltre, è opportuno verificare la purezza dei substrati DNA sintetizzati mediante elettroforesi in gel di poliacrilamide nativi. Nonostante molte aziende garantendo la purezza dei oligonucleotides, abbiamo spesso trovato attraverso i nostri propri test che la purezza del DNA sintetizzato può variare tra i batch.
È importante considerare i seguenti due punti quando conducendo esperimenti con cuvette di quarzo. In primo luogo, alcune proteine sono inclini a associare cuvette di quarzo non specifico. Per contrastare ciò, monolaurato di BSA e polyoxyethylenesorbitan sono inclusi nei buffer di reazione. In secondo luogo, la temperatura ha un effetto drastico sull'intensità di fluorescenza e di velocità di reazione. Per minimizzare questo effetto, la cuvette in quarzo deve essere pre-incubate a 37 ° C prima dell'uso.
Anche se analisi biochimiche convenzionali sono stati estremamente utile nello studio dello scambio sul DNA strand, presentano numerosi svantaggi. In un tipico esperimento di corso di tempo, una reazione viene incubata a una certa temperatura e aliquote sono ritirate timepoints desiderata e deproteinizzate mediante trattamento con proteasi per terminare la reazione e detergente. Al termine del tempo-corso, campioni sono poi sottoposti ad elettroforesi per separare substrati DNA dai prodotti. Il principale vantaggio del metodo descritto qui è che permette l'osservazione in tempo reale della reazione senza alcun disturbo. Qualsiasi timepoint durante la reazione può essere controllato senza interruzioni per la reazione si e non c'è nessuna necessità di deproteinize campioni o sottoporli a forze potenzialmente dirompente di elettroforesi. Questo è particolarmente rilevante durante il monitoraggio di strutture di DNA labile.
Nonostante questi punti di forza nel corso di analisi convenzionali, il metodo qui descritto presentano alcuni svantaggi. Mentre l'uso di substrati del DNA del oligonucleotide per cambio filo semplifica l'interpretazione dei risultati, è importante ricordare che tali substrati non assomigliano i substrati DNA coinvolti in HR nella cella. Alcuni saggi convenzionali utilizzano substrati DNA plasmide di dimensioni medie, che sono più propensi a riflettere il numero di coppie di basi che sono scambiati in vivo. Inoltre, l'utilizzo di substrati di dsDNA circolare topologicamente vincolato in un sottoinsieme dei dosaggi convenzionali è in grado di ricreare almeno in parte le tensioni nel DNA fisiologico.
L'applicazione del metodo descritto qui ha iniziato a svelare i meccanismi molecolari di exchange strand DNA Rad51-driven. Tuttavia, ci sono molte domande interessanti che restano da risolvere. Non vi è chiara evidenza che HR durante la meiosi richiede sia Rad51 e Dmc1, la ricombinasi di RecA-tipo meiosi specifici in eucarioti24. Tuttavia, la mancanza di biochimiche principali differenze tra questi due ricombinasi ha sconcertato i ricercatori nel settore per anni. Inoltre, i ruoli di numerosi gruppi distinti di ricombinazione accessorio fattori è stato un tema focale della ricerca nel campo delle risorse umane. Oltre a chiarire le differenze biochimiche tra Rad51 e Dmc1, miriamo a studiare e confrontare gli effetti di ricombinazione diversi fattori accessorio sulla cinetica del cambio di filo di DNA nell'immediato futuro. Infine, è importante sottolineare che la metodologia basata su FRET qui descritta non è limitata allo studio di cambio filo del DNA. Con modifiche relativamente minori, prevediamo molti tipi di applicazioni per questa tecnica in indagando funzionalmente diverse proteine coinvolte nel metabolismo del DNA25,26,27,28. Speriamo che gli sviluppi descritti qui fornirà ulteriori opzioni ai ricercatori appartenenti a molte diverse discipline.
Divulgazioni
Gli autori non hanno nulla a rivelare.
Riconoscimenti
Questo lavoro è stato finanziato dalla localizzativi per la ricerca scientifica (A) (18H 03985) e su settori innovativi (15H 05974) a HI, per giovani scienziati (B) (17K 15061) a Buenos Aires e per scientifico ricerca (B) (18h 02371) a HT della Japan Society per la promozione della scienza ( JSP).
Materiali
| Name | Company | Catalog Number | Comments |
| 0.2 x 1.0 cm quartz cuvette | Hellma Analytics | 105-250-15-40 | |
| 1.0 x 1.0 cm quartz cuvette | Hellma Analytics | 101-10-40 | |
| adenosine triphosphate (ATP) | Sigma | A2383 | |
| DynaFit | BioKin, Ldt. | DynaFit is a program to analyze kinetics of biochemical reactions. | |
| Fluorescent labeled and non-labeled oligonucleotides | Eurofins Genomics | The sequences of oligos are listed in Table. 1. | |
| Magnetic stirrer | Aisis (Japan) | CM1609 | |
| PCR machine | TAKARA (Japan) | TP600 | TAKARA PCR Thermal Cycler Dice |
| Spectrofluorometer | JASCO | FP8300 | Contains a peltier temperature controller and magnetic stirrer system |
| Syringe | HAMILTON | 1702RN 25ul SYR (22s/2"/2) |
Riferimenti
- Camerini-Otero, R. D., Hsieh, P. Homologous recombination proteins in prokaryotes and eukaryotes. Annual Review of Genetics. 29 (1), 509-552 (1995).
- Cromie, G. A., Connelly, J. C., Leach, D. R. Recombination at double-strand breaks and DNA ends: conserved mechanisms from phage to humans. Molecular Cell. 8 (6), 1163-1174 (2001).
- Pierce, A. J., et al. Double-strand breaks and tumorigenesis. Trends in cell biology. 11 (11), S52-S59 (2001).
- Haber, J. E. Genome Stability. , Garland Science. (2013).
- Kowalczykowski, S. C. An Overview of the Molecular Mechanisms of Recombinational DNA Repair. Cold Spring Harbor perspectives in biology. 7 (11), a016410(2015).
- Bianco, P. R., Tracy, R. B., Kowalczykowski, S. C. DNA strand exchange proteins: a biochemical and physical comparison. Frontiers in bioscience: a journal and virtual library. 3, D570-D603 (1998).
- Renkawitz, J., Lademann, C. A., Jentsch, S. Mechanisms and principles of homology search during recombination. Nature Reviews Molecular Cell Biology. 15 (6), 369-383 (2014).
- Greene, E. C. DNA Sequence Alignment during Homologous Recombination. The Journal of biological chemistry. 291 (22), 11572-11580 (2016).
- Sung, P., Krejci, L., Van Komen, S., Sehorn, M. G. Rad51 recombinase and recombination mediators. Journal of Biological Chemistry. 278 (44), 42729-42732 (2003).
- Prakash, R., Zhang, Y., Feng, W., Jasin, M. Homologous recombination and human health: the roles of BRCA1, BRCA2, and associated proteins. Cold Spring Harbor perspectives in biology. 7 (4), a016600(2015).
- Sung, P. Function of yeast Rad52 protein as a mediator between replication protein A and the Rad51 recombinase. Journal of Biological Chemistry. 272 (45), 28194-28197 (1997).
- Sung, P. Yeast Rad55 and Rad57 proteins form a heterodimer that functions with replication protein A to promote DNA strand exchange by Rad51 recombinase. Genes & Development. 11 (9), 1111-1121 (1997).
- Kurokawa, Y., Murayama, Y., Haruta-Takahashi, N., Urabe, I., Iwasaki, H. Reconstitution of DNA strand exchange mediated by Rhp51 recombinase and two mediators. PLoS biology. 6 (4), e88(2008).
- Jensen, R. B., Carreira, A., Kowalczykowski, S. C. Purified human BRCA2 stimulates RAD51-mediated recombination. Nature. 467 (7316), 678-683 (2010).
- Liu, J., et al. Rad51 paralogues Rad55-Rad57 balance the antirecombinase Srs2 in Rad51 filament formation. Nature. 479 (7372), 245-248 (2011).
- Lu, C. -H., et al. Swi5-Sfr1 stimulates Rad51 recombinase filament assembly by modulating Rad51 dissociation. Proceedings of the National Academy of Sciences of the United States of America. , (2018).
- Bazemore, L. R., Takahashi, M., Radding, C. M. Kinetic analysis of pairing and strand exchange catalyzed by RecA. Detection by fluorescence energy transfer. Journal of Biological Chemistry. 272 (23), 14672-14682 (1997).
- Gupta, R. C., Bazemore, L. R., Golub, E. I., Radding, C. M. Activities of human recombination protein Rad51. Proceedings of the National Academy of Sciences. 94 (2), 463-468 (1997).
- Ito, K., Murayama, Y., Takahashi, M., Iwasaki, H. Two three-strand intermediates are processed during Rad51-driven DNA strand exchange. Nature Structural & Molecular Biology. 25 (1), 29-36 (2018).
- Akamatsu, Y., Dziadkowiec, D., Ikeguchi, M., Shinagawa, H., Iwasaki, H. Two different Swi5-containing protein complexes are involved in mating-type switching and recombination repair in fission yeast. Proceedings of the National Academy of Sciences. 100 (26), 15770-15775 (2003).
- Haruta, N., et al. The Swi5-Sfr1 complex stimulates Rhp51/Rad51- and Dmc1-mediated DNA strand exchange in vitro. Nature Structural & Molecular Biology. 13 (9), 823-830 (2006).
- Argunhan, B., Murayama, Y., Iwasaki, H. The differentiated and conserved roles of Swi5-Sfr1 in homologous recombination. FEBS Letters. 591 (14), 2035-2047 (2017).
- Kuzmic, P. Program DYNAFIT for the analysis of enzyme kinetic data: application to HIV proteinase. Analytical biochemistry. 237 (2), 260-273 (1996).
- Brown, M. S., Bishop, D. K. DNA strand exchange and RecA homologs in meiosis. Cold Spring Harbor perspectives in biology. 7 (1), a016659(2014).
- Rudert, W. A., et al. Double-labeled fluorescent probes for 5' nuclease assays: purification and performance evaluation. BioTechniques. 22 (6), 1140-1145 (1997).
- Xiao, J., Singleton, S. F. Elucidating a key intermediate in homologous DNA strand exchange: structural characterization of the RecA-triple-stranded DNA complex using fluorescence resonance energy transfer. Journal of Molecular Biology. 320 (3), 529-558 (2002).
- Grimme, J. M., et al. Human Rad52 binds and wraps single-stranded DNA and mediates annealing via two hRad52-ssDNA complexes. Nucleic Acids Research. 38 (9), 2917-2930 (2010).
- Algasaier, S. I., et al. DNA and Protein Requirements for Substrate Conformational Changes Necessary for Human Flap Endonuclease-1-catalyzed Reaction. The Journal of biological chemistry. 291 (15), 8258-8268 (2016).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati