
Method Article
示差走査蛍光光度を用いたタンパク質 - リガンド相互作用の定量
要約
Differential scanning fluorimetry is a widely used method for screening libraries of small molecules for interactions with proteins. Here, we present a straightforward method to extend these analyses to provide an estimate of the dissociation constant between a small molecule and its protein partner.
要約
広範囲の方法は、現在、タンパク質との解離定数を決定し、小分子の相互作用に利用可能である。しかし、これらのほとんどは、専門機器へのアクセスを必要とし、しばしば効果的に信頼性の高い実験を確立し、データを分析するための専門知識の程度を必要とする。示差走査蛍光光度(DSF)は、ますます生理パートナーを同定するため、またはヒット発見のためのいずれか、小分子相互作用のためのタンパク質の初期スクリーニングのためのロバストな方法として使用されている。この技術は、定量的PCRに適した唯一のPCRマシンを必要とし、従って適切な器具類は、ほとんどの施設で利用可能であるという利点を有する。プロトコルの優れた範囲がすでに利用可能である;および方法の複数の用途のための文学に強い先例があります。過去の研究は、DSFデータから解離定数を計算するが、これらは数学的に求めているのいくつかの手段を提案している。ここでは、DEMonstrate DSF実験データの中程度の量から解離定数を推定する方法。これらのデータは、典型的には、一日以内に収集し、分析することができる。私たちは、単純な結合事象から収集されたデータを適合させるために使用できる方法の異なるモデルを実証し、ここで、協同的結合、または独立した結合部位が存在する。最後に、標準モデルが適用されない場合には、データ分析の一例を提示する。これらの方法は、研究プログラムから商業的に入手可能な対照タンパク質に収集されたデータ、および2つのタンパク質と示されている。全体的に、私たちの方法は、研究者が急速にDSFを用いたタンパク質 - リガンド相互作用へのさらなる洞察を得るするための簡単な方法を提供します。
概要
全てのタンパク質は、他の巨大分子への単純なイオンから他の分子の多様な範囲に、変動する親和性で、結合するであろう。多くの場合、タンパク質は、それらの通常の機能( 例えば、ATPに結合するキナーゼ)の一部として低分子パートナーに結合する。他の相互作用は、機能とは無関係であるが、ツール( 例えば、結晶化の成功を向上させる、または溶液中のタンパク質を維持するのを助けるためにタンパク質を安定化する小分子)として実験的に有用であり得る;活性部位及びタンパク質のアロステリック部位に結合する小分子阻害剤として作用し、したがって酵素の活性を調節することができる一方。
パートナー分子のためのタンパク質の親和性を決定するために使用することができる技術の広い範囲がある。等温滴定熱量計1は、反応に豊富な情報を提供するように広くラベルフリーであり、「ゴールドスタンダード」とみなされる、との機会が限られている実験のrtifacts。しかし、実験設備の計装や自動化の感度の最近の改善にもかかわらず、それは、タンパク質の要件の面ではまだ比較的高価であり、せいぜい低〜中スループットを持ち、中程度からとの相互作用に最適です高親和性(100μMのK dに 10 nM)の2。このような表面プラズモン共鳴または二重層干渉3オファーより高いスループット、および感度を達成しているような他のラベルフリーの方法は、100ダルトンと低い小さな分子を検出する。しかし、これらの方法のためのハイスループット機器は比較的高価であり、関連するプロジェクトの継続的なスループットがあるだろう場所でのみ正当化され、非常に多くの大学の研究室へのアクセス不能である可能性が高いされている。
示差走査蛍光定量(DSF、またはthermofluor)は、第一の薬剤発見のための方法として、2001年4に記載した。このメトでdは、タンパク質は、タンパク質の疎水性領域に結合すると、その蛍光を変化させる蛍光色素(最初はナフタレンスルホン染料を使用した)と共にインキュベートする。タンパク質 - 色素次いで、試料を加熱し、熱が上昇すると蛍光をモニターした。タンパク質のアンフォールディングおよびタンパク質の疎水性部分の曝露、温度( 図1A)の関数としての蛍光の特徴的なパターンを生じさせる。実験は、商業定量的PCR装置で、少量で行うことができ、したがって単一の実験で、多数のサンプルを同時に(装置モデルに応じて、通常、48、96または384サンプル)を試験することができる。実験は通常、試料5のハイスループット解析の可能性を提供し、時間の周りで行うことができる。
方法論へのさらなる改良は、より良いスペクトル特性6,7と色素の採用を見てきました SUPや初期スクリーニング8,9のためのプロトコルを提案した。この方法の応用範囲は、蛋白質10の調製および保存のために、結晶11を支援するために潜在的な結合パートナーを同定することに最適な条件を確立することに特に焦点を当て、拡張されました。弱い結合分子を研究する方法が比較的高いスループット、タンパク質中の比較的低コスト(反応あたり〜2μg)を、そして適用性は、特に学術的文脈12-14に、DSFのフラグメントベースの薬物設計のための貴重なツールを作っています。
タンパク質 - リガンド相互作用を研究するDSFの幅広いアプリケーションにもかかわらず、いくつかの研究では、これらの研究からの解離定数の決定を記載している。しかし、これらは、疎データまたはいくつかの場合(e)で取り付けられなければならない多くのパラメータと、タンパク質のアンフォールディングを記述する詳細な方程式を生成する傾向が7,15-17を stimated。これらの方法は、異常な遷移を表示きつく結合化合物、またはタンパク質のような、困難なケースで特に関連するものである。しかし、多くの研究室では、これらの詳細な分析は、日常的な使用のためにあまりにも扱いにくい。そこで、異なるシナリオのための代替治療法を提案し、これらは、異なるタンパク質 - リガンド相互作用から得られるデータを適合させるために使用することができる方法を示す。提案手法では、オーダーメイドのデータ解析ソフトウェアが提供されているは、StepOne定量PCR機器を使用しています。これは、データ分析を高速化しながら、他の機器からの結果は、以前に公表された方法9を使用して処理することができ、同じ下流の分析を行うことができる。
プロトコル
(一桁以内、すなわち )解離定数の近似値の1。決定
- 表1に詳述液を調製する。
- 利用可能な最高濃度で目的のリガンドの在庫を準備し、この六10倍希釈で。おおよそのK dは 、前のデータから知られている場合には、K dはの上下に少なくとも2濃度を有することを目指しています。
- 定量PCRプレートの8ウェルに分注し、混合物の18μlの。最初のウェルに溶媒の2を添加する。残りの7つの井戸の1つのウェルそれぞれにリガンド希釈系列(ステップ1.2)の各メンバーの2を添加する。
- プレート上のqPCRシールを配置します。プレートの良好なシールを達成するために、プレートの中央に(特定の試薬の表を参照してください)ハンドアプリケータを配置します。片側にシールを下に滑らかで、その後の他の半分に繰り返すプレート。
- 気泡を除去するために2分間500×gでプレートを遠心。
- は、StepOne定量PCR機器にプレートを置きます。 「融解曲線」オプションを選択し、ROXフィルター、および高速ランプ速度を選択します(これは40分かけて99℃までのランプ、その後、2分休止し、25℃で2分間のポーズを、提供します)。熱変性を実行します。
注:実行を実行するためのスクリプトファイルは、httpからオンラインで入手できます。// www.exeter.ac.uk/biosciences/capsular。 - 楽器の実行の終了時に、画面上の「分析」ボタンをクリックしてください。結果ファイルを保存します。
- タンパク質の熱シフトソフトウェアを開きます。
- 新しい研究を作成します。プロパティ]タブで、この名前を付け、[条件]タブにある、細部のリガンド。
- (実験[ファイル]タブに移動し、保存した結果ファイル(XXX.eds)をインポートし、各ウェルの内容を設定するテンプレートフィルES)は、著者から入手可能である。
- [分析]タブに移動し、「分析」ボタンを押してください。
注:これは、結果を分析します。これは、[エクスポート]タブを使用してExcelとのさらなる調査のために結果をエクスポートすることが可能である。結果は、タブ描写形式でエクスポートされます。それは、Excelでエクスポートされたファイルを開いて、すぐにExcel形式で保存するのがベストです。
- 単独の溶媒の存在下で、タンパク質が、図1Aに示したのと同様の結果が得られることを確認してください。次に、「複製」ペインに結果が観察された融解温度を調べる。これは増加したリガンド濃度の融解温度の明らかな増加を示していることを確認してください。
注:理想的には、これは(タンパク質が完全にリガンド結合していると仮定して)明確な最大融解温度を提供し、溶融温度が、リガンドのないタンパク質との中間にあるおおよその K dになり、最大。
解離定数を決定するための2。実験セットアップ
- マスターミックスとして表2に詳述した混合物を準備します。
- 最後の実験で10倍に希釈される15の異なる濃度でリガンドの株式を準備します。理想的には、少なくとも二桁推定K dの上および下の濃度を含み、推定K dに濃度をセンタリング。このいずれかの側に別の4点で、推定K dの大きさのオーダー内の点の7に焦点を当てる。選択肢がある場合には、飽和している値で、より多くのポイントがあります。
注:必要であれば、リガンド株は、リガンドの溶解度が制限されている二重の実験濃度であるような実験条件を変更することが可能である。 - マスターの120μl添加マスターミックスの便利な分配のためのリザーバとして機能するように、U底96ウェルプレートの8ウェルに混合する。 PCRプレートの1列に18μlずつ分注する8チャンネルピペットを使用してください。プレート上6×8のパターンで48満たされた井戸の合計を与えるために、さらに5つの列に対して繰り返します。
- U底96ウェルプレートのウェルを分離するために、20リガンドの株式μlの、または溶媒を追加します。 8チャンネルピペット、吸引、8つの異なるリガンドストック(または溶剤)を2μlを使用した。ステップ2.3でマスターミックスを充填したPCRプレートの1列にこれらを追加します。さらに2つの列に同じ8リガンド/溶媒ストックを繰り返します。吸引する2残りの8つのリガンドまたは溶媒株式μlの、プレートの4列目にこれらを追加します。さらに2つの列のためにこれを繰り返します。これは、すべての16リガンド、溶媒のサンプルについて3回のサンプルを提供します。
- (ステップ1.4参照)をプレート上のqPCRシールを置きます。
- 500×gでプレートを遠心2分。
- 定量PCR機器にプレートを置きます。ステップ1.6に指定されたパラメータを使用して熱変性を実行します。
- 楽器の実行の終了時に、画面上の「分析」ボタンをクリックしてください。結果ファイルを保存します。
- タンパク質の熱シフトソフトウェアを開きます。新しい研究を作成します。プロパティ]タブで、この名前を付け、[条件]タブにある、細部のリガンド。
- 実験ファイル]タブに移動し、保存した結果ファイル(XXX.eds)をインポートし、各ウェルの内容を設定する。
注:テンプレートファイルは、httpからオンラインで入手できます。// www.exeter.ac.uk/biosciences/capsular。 - [分析]タブに移動し、「分析」ボタンを押してください。
- 三連として結果を表示するには、画面の左側にあるメニューから「複製し」タブを選択します。三重はどのようにタイトに基づいてデータの信頼性を評価する。 Shoul三連が悪い再現性を示しているD、密接に生データを調べます。
- 溶融温度を評価するためにボルツマンまたは誘導体法の両方を使用してデータを分析する。 「結果を複製」タブを選択し、 "でプロット:「トグル、「結果プロットを複製」の間のボタン「TM - ボルツマン」と「TM - デリバティブ」。サンプル用の大きな再現性を与える方法を選択します。 [エクスポート]タブを使用してExcelでさらに調査の結果をエクスポートします。
注:複数の遷移を示しているサンプルについては、複数の溶融モードでのデリバティブメソッドを使用するほとんど常に最善の方法です。結果は、タブ描写形式でエクスポートされます。それは、Excelでエクスポートされたファイルを開いて、すぐにExcel形式で保存するのがベストです。 - 結果の再現性を確保するために、独立した日には、リピートを含め少なくとも2回の実験を繰り返します。データ分析(下記のステップ3を参照)はずK dの値が、元の推定値に有意差があることを示し、K dは周囲の値の良好な範囲を確保するために(ステップ2.2参照)に応じてリガンド濃度を変化させる。
熱変性の下での解離定数を決定するために、3。データ解析
- リガンド濃度のExcelで表と溶融温度を作成します。
- グラフパッドプリズムソフトウェアを開き、XYテーブルを作成します。リガンド濃度および温度の結果を溶融させるためのY列のX列を使用して、データを入力します。
注:示されている例が図1Bである。プリロード方程式、およびSPSS統計パッケージを使用するための代替指示にスクリプトは、HTTPでオンラインで入手できます:// www.exeter.ac.uk/biosciences/capsular。 - [分析]タブでは、分析パラメータを変更するオプションを選択秒(Ctrl + Tキー)。 、正しいモデルを入力し、「新規作成」を選択し、「新たな方程式を作成」します。 「単一部位結合リガンド」として表3に詳述され、式を挿入します。
注:これらの手順の例を図1Cに示されている。プリロード式でスクリプトを使用する場合は、関連する方程式は、リストから選択ではなく、入力に向けることができる。この式の導出は付録に記載されている。 - ボックスの「初期値の規則」を選択し、 表3に詳述されるように、初期値のための規則を入力します。
- (リガンド内の指定されたのと同じ単位)とタンパク質の最終濃度を入力する」に等しい定数」として、パラメータPを拘束。
- 分析を実行するために[OK]を選択します。
注:これらの手順の例を図1Dに示されている。グラフ作成ソフトウェアは、データとモデルへの適合を示す図を生成します。 Tの例ヘセ分析は代表的なデータに示されている。
協同組合モデルへのデータのフィッティング4。
協力的なモデルにデータを適合させるために、単純な協力モデル、または2の別個の解離定数が定義されているモデルのいずれかを選択。最初のアプローチは、負の協同の場合において好ましい、または最初の調査などされている。しかし、原理的には、二つの異なる解離定数18をモデル化するために正の協同例に優れています。この場合、モデリングは、リガンドの結合、順次、またはリガンドの結合に依存しないのいずれかを仮定して進めることができる。
- ステップ3.3で、しかし、プロトコル3と同様の初期ステップに従って、「シンプル協力モデル」と記載されている表3の方程式の1、「二つの配位子の逐次結合"を挿入、または18"独立の2つのリガンドの結合」。
- 初期値に関連するルールを選択します表3にこれらの式のそれぞれに関連する。
- データへのモデルの適合を調べます。データが不十分適合する必要があり、別のモデルを考える。
注:それは慎重にプロテインサーマルシフトソフトウェア(ステップ2.9)することで、データへの溶融温度のフィッティングを検討することも重要です。時にはそれが最良の結果を得るために、ここでパラメータを変更する必要がある。 K dのいずれかの側でのデータの限定されたセット、または(特に最高のリガンド濃度での)単一の異常な点のいずれか、大幅に影響を与えることができる:別の考慮事項は、データ点の範囲が理想的であり、いずれの異常な点が存在するかどうかどうかである結果。 - 再現性を確実にするために(ステップ2.12参照)を少なくとも二回の実験を繰り返します。
融解温度のバイナリシフトを表示曲線にデータ近似5。
時折、むしろリガンドへの段階的な応答よりも、タンパク質がされている結合した試料は明らかに結合していない試料から分離されているバイナリの応答を、採用することが観察された。例は代表的な結果( 図4)が設けられている。この場合には、融解温度のフィッティングはK dのための良好なフィット感を提供することはありません。
- タンパク質の熱シフトソフトウェアからの生のデータ出力をエクスポートします。各温度点については、ゼロリガンドに対する平均蛍光、最高リガンド濃度を計算します。これらの隣に各データ点の結果を集計。
注:ここで作成したエラーは、フィットの融解温度の誤差よりも小さい。 - SPSS統計パッケージを開きます。 SPSSのデータウィンドウに各実験のための温度、2平均値のデータセット、およびデータをコピーします。変数]タブで、「低」、「高」が最も高いリガンド濃度の平均データセットとして無リガンドの平均データセットを設定します。
- 使用可能な構文ファイルをダウンロードオンラインでは、http:// www.exeter.ac.uk/biosciences/capsular 。 「ファイル名を指定して実行]→[実行]のすべて」を選択してください。
- 関連するリガンド濃度で、新しいExcelワークブックに結果をバインド割合をコピーします。
- グラフパッドソフトウェアを開き、XYテーブルを作成します。リガンド濃度および温度の結果を溶融させるためのY列のX列を使用して、データを入力します。分析]タブで、「変化分析パラメータ」を選択します。 、正しいモデルを入力し、「新規作成」を選択し、「新たな方程式を作成」します。 「融解温度のバイナリシフトの分析」としてリストされ、表3に示した式を入力します。
- 「初期値の規則」ボックスをオンにして、 表3に詳述された初期値のための規則を入力します。」に等しい定数」として、パラメータPを拘束し、リーガと同じ単位で(タンパク質の最終濃度を入力してくださいnd)がで与えられている。
注:セクション3のプロトコルのためのこれらのボックスを完了する例が図1C、Dに示されている。 - 良好な適合がある場合、結果は、無限のリガンド濃度での期待される結果を外挿することにより改善することができる。各リガンド濃度で結合の割合のモデルから、リガンド濃度の最高値の値を調べます。これが0.99以上であれば、さらなる分析が結果を改善しにくい。
- 比率が0.99未満である場合、追加のステップが、最も高いリガンド濃度試料中の非結合タンパク質の影響を補正する必要がある。ステップ5.2では、セルR2(異なるセルを使用してもよく、R2は、 表3の式で適宜置換さ)にする(ステップ5.7からの)は、最も高いリガンド濃度の点で結合したリガンドの割合を書き込む。最高リガンド濃度結果の平均の後に余分な列を作成します。モミでT細胞は、「無限のリガンド濃度を外挿」として表3に示す式をコピーする。この列の残りのセルにこの数式をコピーします。
注:この計算は、最も高いリガンド濃度で非結合タンパク質の影響を除去します。リガンド遊離タンパク質と最も高いリガンド濃度との差を各温度点での完全結合および非結合タンパク質状態間の予想される差異を提供するために、最も高いリガンド濃度での結合した割合の逆数を乗算する。この差は、完全に、リガンド結合タンパク質の予想される蛍光を与えるために結合していない状態から加算または減算する。 - この新しい列を持つSPSSのデータシートに最大リガンド濃度の列を交換し、データフィッティングを繰り返します。
注: - モデルは、最大リガンドconcentratiで結合割合のさらなる有意な変化を示唆している場合は、5.9を繰り返すことが必要になる場合があり5.7は、手順上(この場合であれば、それはおそらく含ま高いリガンド濃度ポイントと実験を繰り返すことが理想的である)。 - タンパク質は、バイナリシフトして表示協同挙動を示している場合では、方程式は、ステップ4.1からのもので置換されていなければならないステップ5.5で提案されている。 「トップ」と「ボトム」のパラメータは、それぞれ1と0に置き換える必要がある。
- 再現性を確実にするために(ステップ2.12参照)を少なくとも二回の実験を繰り返します。
結果
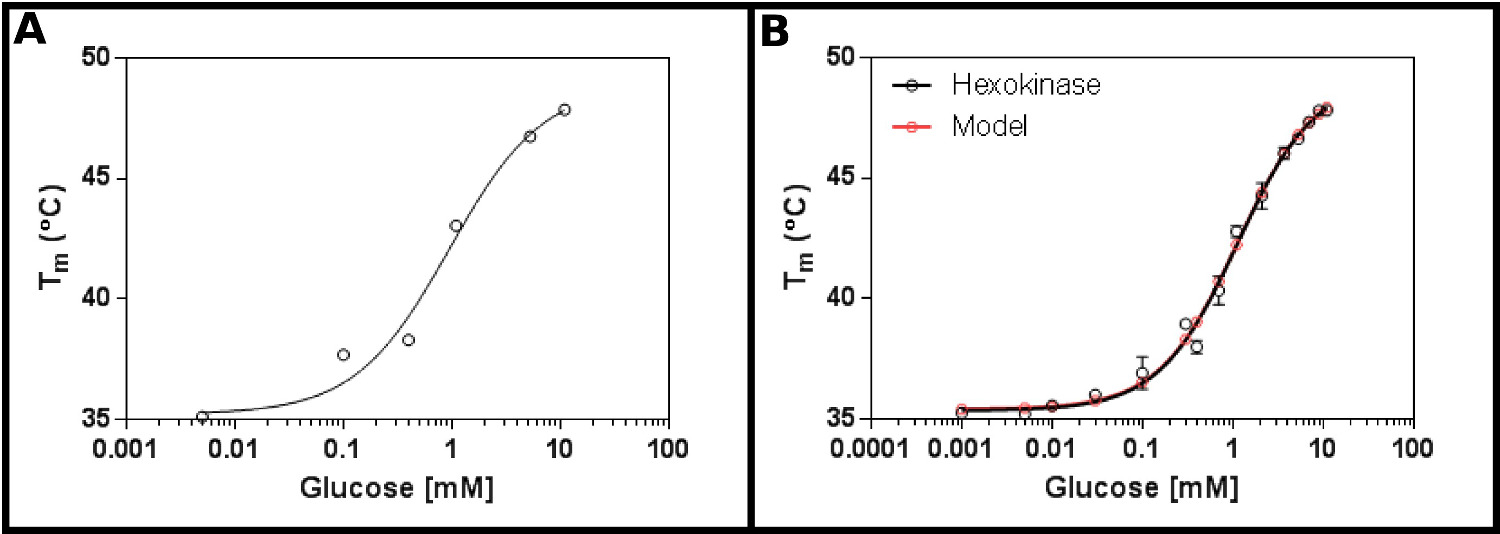
この方法のための優れたテスト基板はヘキソキナーゼです。これは、商業的に容易に入手可能であり、最も研究室で見られる2つの基板を有するという利点を有し、そしてアッセイにおいて明確な、再現性のある結果を提供する。初期濃度画面(プロトコル1)は、ヘキソキナーゼおよびグルコース( 図2A)を使用して、可能性が高いK dは 0.2〜1.7 mMの範囲であることを示唆している。そのため、大画面(プロトコル2)は表4の結果( 図2B)に示されている濃度を用いて、実施されたシングルサイトリガンド結合方程式に良好な適合を示した(プロトコルセクション3.3)[9]、および与えたK 1.2±0.1 mMとD。
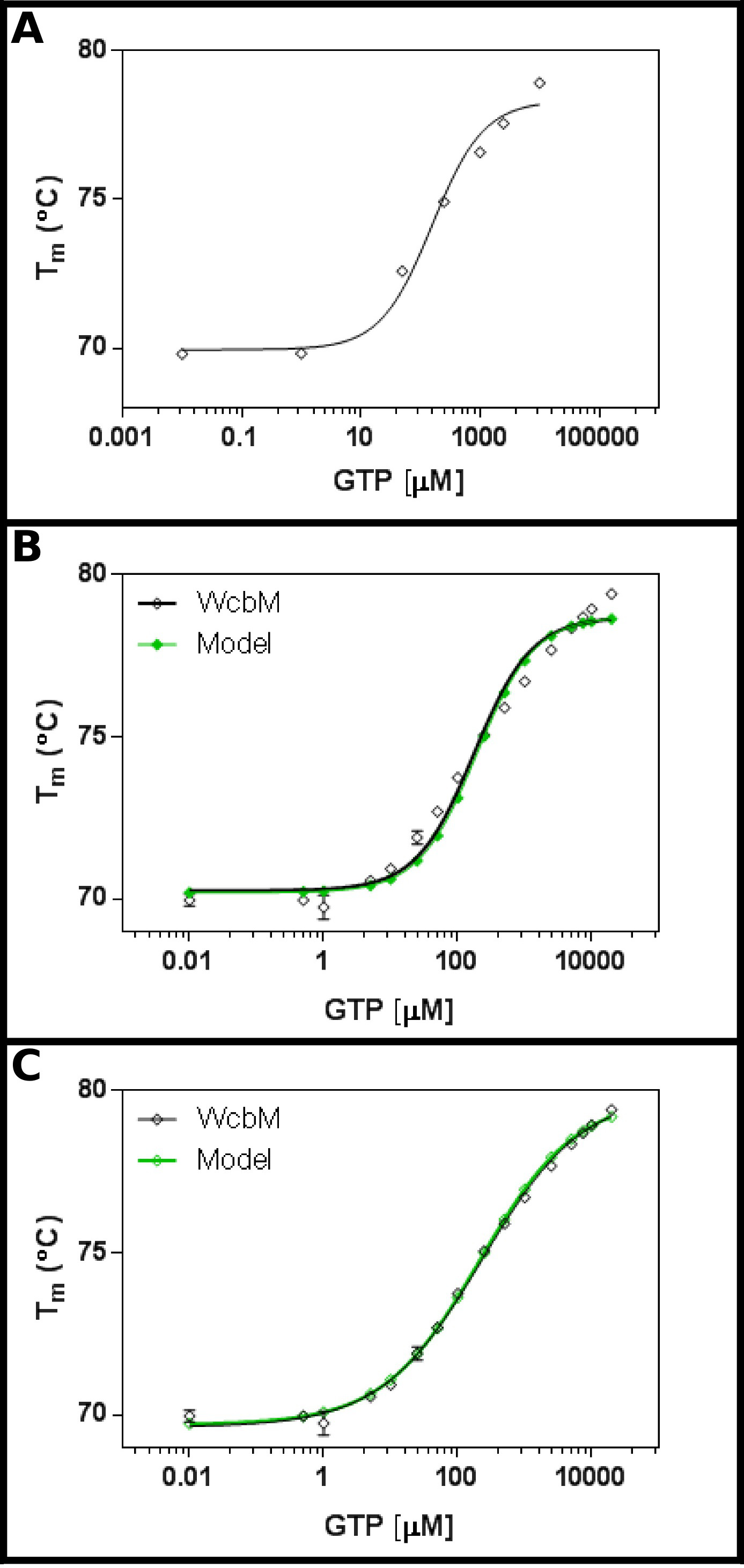
推定のヘプトース-グアニルトランスフェラーゼWcbM 19,20は、GTP( 図3A)との結合に強い熱シフトを示している。初期画面では、Kと示唆dは 100μm程度の範囲であろう。そのため、全画面を表5に示す濃度を使用して、設定された結果のフィッティング3.3を式に程よいフィット感が示された。(0.981のR 2と 、図3B).Howeverは、データとの間に明白な違いがある異なる方程式が必要であることを示唆しているモデル。 WcbM配列を有するタンパク質データバンク21の検索は、構造体が二量体を形成して決定されている最も近いホモログことを示した。データは、したがって、協調シーケンシャル、および2つの配位子(プロトコル4)の独立した結合のための3つの方程式を用いて分析した。順次および独立した結合モデルの両方が0.992のR 2値および0.480と0.461のSy.xを与えたのに対し、協同モデルのフィッティング統計は、R 2 0.998の値と0.215の残差(Sy.x)の標準偏差を与えたそれぞれ。これは、モデルを示唆している協力的なモデルでは、データに最もフィットをした与える:ここでは230±10μMのK½が観察された、0.52±0.02( 図3C)のnの値で。これは、結合に負の協同を示した。 K dの単位はむしろ不十分μM0.52であるように、K½は 、むしろK dよりも、この場合に使用されたことに注意してください。
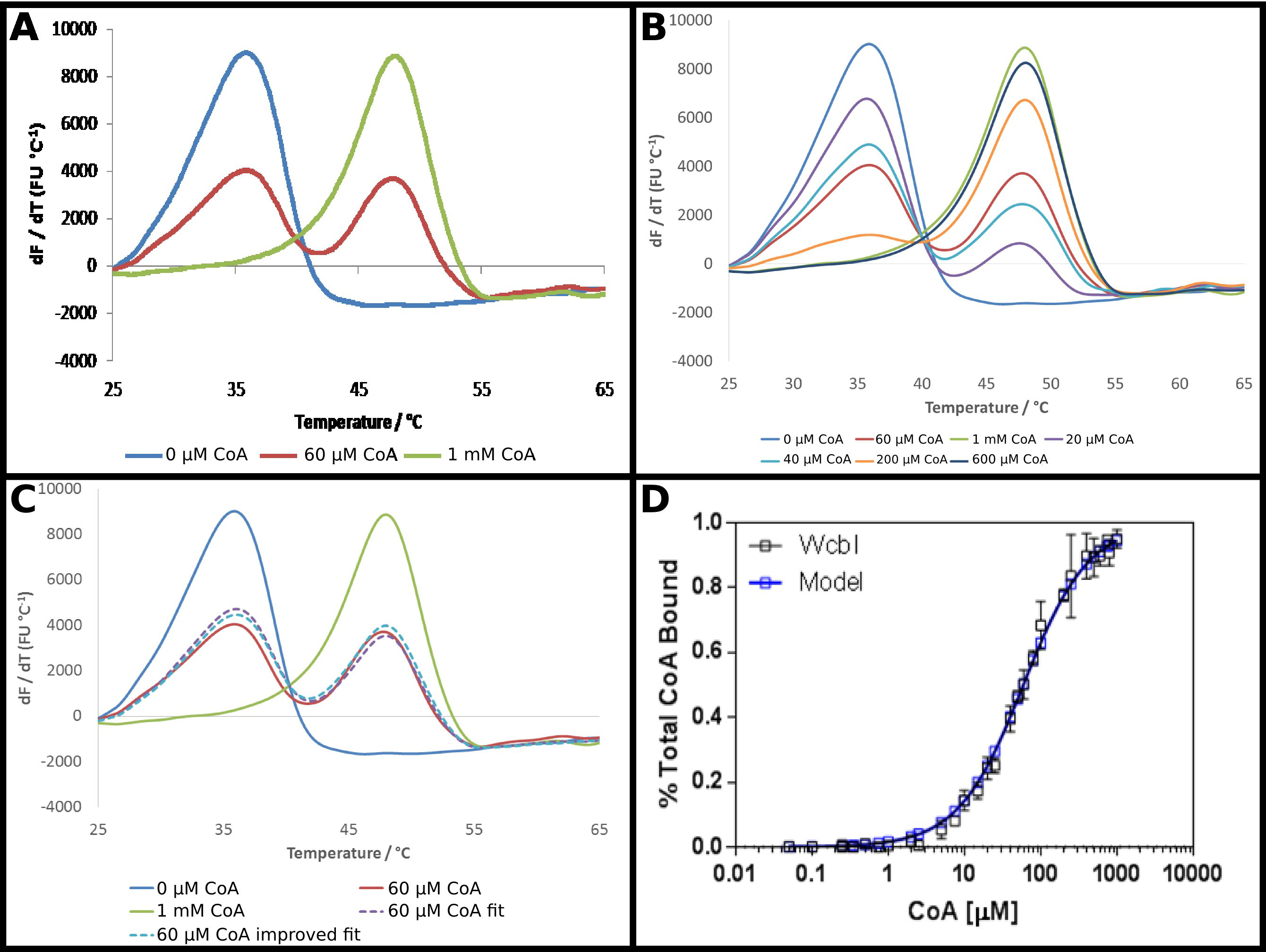
推定上のGDP-6-デオキシ-β-D- マンノ -heptopyranose 2- -acetylase O、WcbI 22は 、示差走査蛍光定量はむしろ珍しい結果を示している。任意のリガンドの非存在下では、それが明確かつ単純な変性( 図4A)を示す。補酵素A(CoAレダクターゼ)は、DSFを使用して、このタンパク質のリガンドとして同定された、プロトコルに記載のように、このパートナーに対するタンパク質の親和性を調べた。 CoAの高濃度、強い秒の存在下で高い温度にhift 15℃の融解温度の変化に伴って観察される。しかしながら、中間の濃度ではなく、中間の溶融温度で溶融相性へのシフトで、WcbIのいずれかのリガンドを含まない温度で溶融するように現れるタンパク質、または完全に結合した融解温度( 図4A)と、二相性の融解を示した。より高い温度( 図4B)で融解割合を増加させる基質濃度の増加に伴って、用量依存的に改変された二つの種の割合。これらのデータの直接分析は、困難であった:ボルツマン方程式に当てはめる派生法は、2つの融解事象が発生していたが、増加するリガンド濃度変化を示す根拠としなかったことを強調しながら、非常に貧弱なフィットを与えた。
これらのデータを分析するための少ない従来のアプローチは、したがって、(プロトコル5)を採用した。蛍光Dリガンドなしの最高リガンド濃度におけるerivative結果は本質的に低い溶融温度のすべてのタンパク質、またはより高い溶融温度の状態を表すとした。残りの微分データが団結( 図4C)に加算された割合が、これら二つの状態のそれぞれの割合の合計として各点で適合させた。得られたデータは、その後、前と同じ式を使用して、見かけのK dを取得するために以前のように適合させた。これが「高」は、リガンドポイントがバインドされた唯一の95%のリガンドである可能性が高いことを強調した。データは、その後、100%結合したタンパク質に対する結果の予測に外挿され、データフィッティングは58±2μMの見かけのK dを与えるために繰り返した。これは、結合モデル( 図4D)の実験結果の優れたフィット感を提供した。
FO:コンテンツ幅= "5インチ" SRC = "/ファイル/ ftp_upload / 51809 / 51809fig1highres.jpg"幅= "500" />
実験のセットアップと分析の1例を図。 (A)(酵母ヘキソキナーゼのデータから取られた)熱変性プロファイルの期待される形状の一例。生データの特徴的な形状(9においてより詳細に説明する)浅い減少に続いて最大蛍光の漸進的な上昇を示している。これは、蛍光の一次導関数における単一のピークを伴う。グラフパッドへのデータ入力(B)の例。リガンド濃度はX軸上に与えられ、Y軸上の融解温度が観察される。グラフパッドにおける方程式の定義の(C)の例。正しく変数の初期値を設定し、タンパク質濃度を固定(D)としては、例えば、解離定数の正確な決定を可能にする。M /ファイル/ ftp_upload / 51809 / 51809fig1highres.jpg "ターゲット=" _ブランク」>この図の拡大版をご覧になるにはこちらをクリックしてください。

示差走査型蛍光定量によって測定されたグルコースとのヘキソキナーゼの図2の相互作用(A)グルコース濃度の広い範囲を検査する最初の実験は、K dが 0.2の範囲である可能性が高いことを示唆している- 。1.7mMの(B)の詳細実験は、グルコースの16の濃度をテストし、1.12±0.05 mmの見かけのK dの決定を可能にする。データは、単一の結合(底部(T1)と、トップはそれぞれ35.4±0.2ºCおよび49.3±0.5ºCにふさわしい(T2)の温度)イベントのモデルに非常によく適合します。あることに注意してくださいこれらのデータは、10mMのMgCl 2の存在下で収集した。これらのイメージは、グラフパッドを用いて調製した。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}

GTPとWcbMの図3の相互作用は、抗協同的結合を明らかにしている。 (A)GTPの広範囲の濃度を試験する初期の実験は、K dは 200の範囲である可能性が高いことを示唆している- 。500μMGTPの16濃度を試験する(B)詳細な実験は、見かけ上の価値を示唆している120±20μM の K d。しかし、対数目盛をx軸に使用される場合、有意なdiscreがあるモデルとデータとの間pancy。の協力モデルと同じデータ(C)の分析では、単純な協力モデルが使用されているデータへの優れたフィット感を示しています。ここで、K 230±20μM の 2分の1(それぞれ69.63±0.06ºCおよび79.9±0.1ºCにフィッティング底部(T1)とトップ(T2)温度の)協同性係数はn = 0.52±0.02で、決定した。 WcbMは二量体であるように見えるように、これは、酵素がそのがGTPへの結合において完全にanticooperativeであることを意味する。これらのイメージは、グラフパッドを用いて調製した。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}

図4 WcbIは二相性の融解パターンを示しているそのリガンド補酵素A(CoA)レダクターゼの存在下で行われる。 (A)WcbIは、リガンド(青)の非存在下で、単純な単相の融解パターンを示している。高いリガンド濃度(1mMの、緑色の線)では、同様のパターンが観察される。しかしながら、中間のリガンド濃度(60μM;赤線)で、リガンドを含まず、リガンド結合状態に対応する二つの別個の融解ピークが観察される(B)のピークの2つのセットの間の遷移は、フルにわたって用量依存的である。濃度の範囲は、。リガンドフリーで高リガンド結果の割合の合計と相性の融解(C)モデリングは、(赤線と比較して、一点パープルライン)のデータへの良好なフィット感を提供します。このフィットは満室(青い点線)から(モデルは示唆している〜95%の占有率)の高いリガンド濃度について観察した結果を外挿することにより改善される。(D)のCoA笙にバインドWcbIの割合について得られたデータを WS 58±2μmのK dで単純な結合モデルへの優れたフィット(これらのデータは、結果の最初のセットに基づいて、二日目のために選択された、わずかに異なるリガンド濃度で、2別の日に収集されたデータを表す)。パネル(A - C)は 、Excelを使用して製造し、パネル(D)は、グラフパッドを使用していた。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
最初の実験のために表1のレシピ。
| 試薬 | ミックスの容量(μL) |
| タンパク質 | 0.11 mg / mlの最終濃度 |
| 0.3 | |
| 0.5MのHEPES pH7.0の | 3.7 |
| の5M NaCl | 5.6 |
| 水 | 180μlに |
これは、タンパク質の「マスターミックス」、検出試薬を説明し、プロトコルセクション1に記載したように、この緩衝液混合物は、一般的なタンパク質に適した、K dの推定値を提供するための初期スカウト実験のためにバッファリング。以前の結果は、他のバッファを使用する必要があります示唆している場合、これらは置換されるべきである。タンパク質ストックは、低濃度( すなわち、0.3未満mg / mlで)である場合、それは追加のバッファの量を低減する必要があるかもしれないタンパク質試料中に既に存在するバッファを補うために添加した。
K dを決定するためのクラス= "jove_content"> 表2レシピ。| 試薬 | ミックスの容量(μL) |
| タンパク質 | 0.11 mg / mlの最終濃度 |
| 5,000倍SYPROオレンジ | 1.78 |
| 0.5MのHEPES pH7.0の | 22.2 |
| の5M NaCl | 33.3 |
| 水 | 180μlに |
これは、タンパク質の「マスターミックス」、検出試薬、およびbについて説明しタンパク質試料の K dの完全な決定のためuffer、プロトコル項2に記載のように、この緩衝液は、一般的なタンパク質に対して適切である。以前の結果は、他のバッファを使用する必要があります示唆している場合、これらは置換されるべきである。タンパク質ストックは、低濃度( すなわち、0.3未満mg / mlで)である場合、それは追加のバッファの量を低減する必要があるかもしれないタンパク質試料中に既に存在するバッファを補うために添加した。
表3方程式とデータ分析のためのパラメータ 。
dの| 実験プロトコルのステップ | 式に必要な | 必要なパラメータ | 変数とパラメータの説明 |
| 3.3 | |||
| 単一部位リガンド結合 | Y =ボトム+((トップ下)*(1 - ((PK D -Xの+ SQRT(((P + X + K dは )^ 2) - (4 * P * X)))/(2 * P )))) | P:タンパク質濃度。 Kdは:解離定数。 P及びKdは、リガンド濃度について使用したのと同じ単位で与えられる。上、下:無限のリガンド濃度をそれぞれ無リガンド濃度での融解温度。 | |
| 3.4 | ボトム= * YMIN | YMIN、Yの最小値(この場合は最低の実験的なタンパク質のTm、) | |
| トップ= * YMAX | YMAX、Yの最大値(最も高い実験タンパク質のTm) | ||
| K dは = * X YMIDで | YMID:YMINとYMAXの平均に相当するYの値。 Xは、対応するX値(ここでは、関連するリガンド濃度)である | ||
| P =(初期値、フィットするために) | |||
| 4.1 | |||
| シンプルな協同モデル | Y =ボトム+((トップ下)*(((X / Kd)を^ N)/(1 +((X / Kd)を^ N)))) | N:H病気係数。これは、タンパク質の協同性、または他の生化学的特性を説明し、必ずしもタンパク質におけるリガンド結合部位の数の推定値ではない。 1のヒル係数は協同性を表しません。 1未満の値が負の協同を示し、1より大きい正の協同値。 | |
| ボトム= * YMIN | |||
| トップ= * YMAX | |||
| K dは = * X YMIDで | |||
| P =(初期値、フィットするように ) | |||
| N =(初期値、フィットするために) | |||
| 二つの配位子の結合を逐次 | Y =ボトム+((トップ下)*((X ^ 2)/(K dは *のK2))/(1 +(X / K d)は +((X ^ 2)/(K dは *のK2))) ) | K2:第二の結合イベントの解離定数。 | |
| ボトム= * YMIN | |||
| トップ= * YMAX | |||
| YMIDでK2 = * X | |||
| P =(初期値、フィットするために) | |||
| 独立2リガンドの結合 | Y =ボトム+((トップ下)*((X ^ 2)/(K dは *のK2))/(1 +(2 * X / K d)は +((X ^ 2)/(K Dの * K2) ))) | ||
| ボトム= * YMIN | |||
| K dは = * X YMIDで | |||
| YMIDでK2 = * X | |||
| P =(初期値、フィットするために) | |||
| 5.5 | |||
| 融解温度のバイナリシフトの分析 | Y = 1 - ((PK のd-X + SQRT(((P + X + K d)の ^ 2) - (4 * P * X)))/(2 * P)) | ||
| ボトム= * YMIN | |||
| トップ= * YMAX | |||
| K dは = * X YMIDで | |||
| P =(初期値、フィットするために) | |||
| 5.8 | |||
| 無限リガンド濃度に外挿する | (C2 - ((1 - $ R $ 2)* B2))/ $ R 2ドル | B2:なしリガンドとの結果を含むセル。 C2:最大リガンドとの結果を含むセル。 $ R $ 2:最大リガンド濃度で結合比率を含むセル。 |
図3、図4および図5は、解析ソフトウェア、及びデータ分析のためのパラメータの開始の正確な定義に詳細な方程式の添加を必要とする手順。各関連ステップの方程式は、パラメータの正しい選択をして、表示されます。変数およびパラメータの意味の説明は、参考のために提供される。
表グルコースとヘキソキナーゼの相互作用のスクリーニングのため4。濃度。
| 標本点 | リガンド(glucose)の濃度(mM)の |
| 1 | 0 |
| 2 | 0.001 |
| 3 | 0.005 |
| 4 | 0.01 |
| 5 | 0.03 |
| 6 | 0.1 |
| 7 | 0.3 |
| 8 | 0.4 |
| 9 | 0.7 |
| 10 | 1.1 |
| 11 | 2.1 |
| 12 | 3.7 |
| 13 | 5.3 |
| 14 | 7 |
| 15 | 9 |
| 16 | 11 |
プロトコールに記載されているように出芽酵母サッカロマイセス·セレビシエからのヘキソキナーゼは、マグネシウムが知られている補因子であるように、10mMのMgCl 2を補充し、マスターミックスに添加した。 K dの初期推定値は、0.5〜2 mMの間であった。実験は、グルコースの示す最終濃度を提供するように設定された。
表のGDPとのWcbMの相互作用のスクリーニングのための5。濃度。
| 標本点 | リガンド(GTP)濃度(μM) |
| 1 | 0 |
| 2 | 0.5 |
| 3 | 1 |
| 4 | 5 |
| 5 | 10 |
| 6 | 25 |
| 7 | 50 |
| 8 | 100 |
| 9 | 250 |
| 10 | |
| 11 | 千 |
| 12 | 2,500 |
| 13 | 5000 |
| 14 | 7,500 |
| 15 | 万 |
| 16 | 2万 |
プロトコールに記載されているよう疽菌からWcbMはマスターミックスに添加した。 K dの初期推定値は約100μmであった。実験は、K dはの上下少なくとも二桁をカバーすることを目指し、GTPの示す最終濃度を提供するように設定された。
ディスカッション
示差走査蛍光定量は、タンパク質を特徴付ける、および潜在的なタンパク質リガンドを同定するための堅牢で汎用性の高い方法として、その力を実証してきました。 (特にあまり資金を研究室で)タンパク質の安定化、薬物の発見を促進し、結晶化10,23-25 において十分に実証の成功は、それ化合物の初期スクリーニングのための魅力的な方法作った。タンパク質に付加する化合物は、見かけ上の溶融温度7,9の明確な用量依存性の増加を示す。しかし、それらの親和性について化合物をランク付けするのを助けるために見かけの結合定数を決定するために、これらの実験からの結果を使用するいくつかの試みがなされてきた。ここでは、体系的リガンドの存在下でのタンパク質の見かけの解離定数を決定するための方法を提示する。
ここに示す結果は、DSFを迅速かつ確実にするための解離定数の推定値を提供できることを実証タンパク質 - リガンドの組み合わせ。観測データは、パラメータの可能性が高い値に関する仮定をする必要なしに、K dの迅速な決定を提供するために、市販のツールを使って操作することができます。この方法は、必要なタンパク質と時間の両方での倹約であることのいくつかの同等の方法に比べて大きな利点があります。ここに記載した実験は、実験当たりタンパク質0.13ミリグラム(三連で繰り返し実験では約0.4 mg)を消費します。これは平均的な40kDaのタンパク質と単一の実験は、同様の量を消費する等温滴定熱量測定(ITC)、と好意的に比較します。このプロトコルのために必要な実験のフルセットは、実験の単一のセットのための準備、など約4時間、消費する。再び、これは、強力ながら、多くの場合、最良のデータを得るために、かなりの最適化が必要ITCまたは表面プラズモン共鳴などの方法よりもかなり迅速である可能性が高い。
本発明者らの結果は慎重に生データを検討する必要性が残っていることを示している、これらのデータのフィットは、融解温度を決定するために、溶融温度データの適合は、解離定数を決定する。最初の課題は、タンパク質の溶解で製造した生データの形である。いくつかのケースでは、形状は、 図1Aで観察されるものに近似しなくてもよい。一般的な問題は、リガンド結合上の低温度変化、高いバックグラウンド蛍光、および温度の異常な複数の遷移が含まれています。低温シフトは、リガンドの数を結合に見られる。この方法では、最も重要なパラメータは、温度シフトに比べて、T mの測定における誤差である。 3回の測定の標準偏差が結合していないと完全に結合したタンパク質との間の融解温度シフトの10%を超えていない場合に、データは、通常、合理的に取り付けることができる。私たちの経験は、どこにそのような気温ということですeratureシフトはわずか2℃での個別のデータポイントが非常に正確である場合、これは、データをフィッティングするために十分であり得る。第二の問題は、非常に曲線形状となっている。リガンド結合は、タンパク質のモードを展開影響を与えるように、これらは多くの場合、無料の蛋白質とリガンド結合形態の間で異なる。これらのケースでは、ユーザは、データが融解温度および解離定数を決定するために使用されるモデルの適切な考慮して使用できるかどうかを考慮しなければならない。もう1つの一般的な問題は、タンパク質の補因子の添加( 例えば、ヘキソキナーゼとの例でのMgCl 2)が最も信頼性の高いデータを得るために必要とされることである。私たちの経験は、最初の読み取りを行う段階で、実験中のすべての可能性が高い要因を慎重に検討が最良の結果を得るために不可欠であることとなっている。さらに、代替理論的治療法は、これらのデータ15,17の特徴を明らかにすることができます。最後に、Cいくつかのタンパク質は珍しいことではありませんontainネイティブ高いバックグラウンド蛍光を示すために疎水性領域を露出した。広範囲に他の場所で6,9概説されているこれらの問題に対する解決策がいくつかあります。
具体的には、ユーザは、( 例えば、 図4)、及びその誘導体を使用する場合には、複数の溶融物をモデル化する必要があるかどうかボルツマンまたは派生モデルを使用するかどうかを考慮しなければならない。熱変性をモデル化する方法は2ボルツマン法がアンフォールディング曲線への定期的なS字状の形状を仮定して、ボルツマン方程式に実験データをフィットするという点で異なる。対照的に、誘導体方法は、各点( 図1Aの下のパネル)での実験データの一次導関数をとり、最も高い一次導関数の点と融解温度を考慮する。 3℃ - 派生方法は、一般的に約2だけ高い溶融温度を返します。ほとんどのタンパク質は、より一貫性のある戻りますその結果、二つの方法のいずれか( すなわち、三連の実験のために溶融温度の標準誤差は低くなる)。これは通常、タンパク質のアンフォールディング曲線の正確な形状に密接に関連し、それは経験的にそれぞれの場合に最適な方法を決定する必要がある。誘導体モデルが使用される場合、それは、複数の融解事象を考慮することも重要である。いくつかのデータは明らかに、複数の遷移の証拠を示し、これらの場合の結果は、これらの複数の融解イベントがモデル化される場合に解釈することが容易である可能性が高い。このプロトコルのコンテキストでは、リガンドの添加、またはその逆 (最も熱的に脆弱なサブドメインを安定化させることにより、 例えば、)単一の遷移に複数の融解転移を有することからシフトするタンパク質を引き起こすことがよくある。そこで、生データを使用するのがベストであろうどのアプローチ検討する前に一緒に検討されていることを提唱したい。
メートルに続いて個別の融解温度のodellingは、更なる問題は、プロトコルセクションに表示モデルにこれらのフィッティングに発生する可能性があります。なお、この分析はしばしば観察されたデータとモデル間の相違を強調したよう慎重に、対数目盛を用いて解離定数方程式に当てはめを調べるために不可欠である(電子.G。 図 3)。得られた結果は、一般的に強固である一方で、解釈のケアはデータから、より良い結果を抽出する機会、そして最も意味を提供しています。
これらのデータが提起した特定の問題は、DSFで、協同性を示し、タンパク質、または複数の結合イベントに配置する必要があります解釈である。私たちは、これまでに、唯一の複数の特定の結合事象( 例えば、WcbM、その最高の相同体26の多量体であり、サイズ排除クロマトグラフィー上の多量体として作用する[データではないタンパク質を有することが期待されるタンパク質においてこの現象を観察している示さ])。むしろ、これはメソッドの広い範囲を使用して、より徹底的に調査されなければならない複雑な結合を示している可能性がありますそれは、DSFの変性で観察された負の協同が酵素で、最終的には負の協同が表示されることを示していることは全く明らかではない。これはそのようなタンパク質のより広範な研究が面白い効果を特定する可能性があること、しかし、私たちに示唆していない。
この方法を用いて解離定数のために与えられた値は、一般に、等温滴定熱量測定及び表面プラズモン共鳴などの他の方法によって提供されるものと同じオーダーである。しかし、観察された絶対値は、これらの方法を使用して観察されるものより頻繁に高い。これは、少なくとも部分的に解離定数は、リガンドとタンパク質の融解温度で観察されるという事実の結果である。このK dは 、生理的温度でのものよりも一般に高い。 dissociationは定数式により反応の温度に関連している:
 [1]
[1]
 [2]
[2]
(Cθは 、標準的な基準濃度であり、ΔはGは反応のギブスの自由エネルギー変化であるR、Rはモル気体定数、ΔHは反応におけるエンタルピー変化であり、ΔSは反応におけるエントロピー変化である。)
この方法の測定可能範囲内の解離定数との反応は、一般に負のΔrは Gを持ち、従って式に温度上昇の影響は、[1]の解離定数を増加させるであろう。どちらのギブスの自由エネルギーを構成ΔH、ΔSの用語は、(式[2])temperaturですE従属27、および解離定数への影響は、これらの温度依存性の大きさと符号に依存し、必ずしも相互作用依存することになる。したがって、この方法によって決定解離定数は、時にはRTで動作する方法によって決定されたものよりも大きいことは予想外ではない。温度依存性はまた、もちろん、生理的温度より低い温度で解離定数を提供する傾向がある多くの他の方法、の警告である。
それはITCとは異なり、ラベルされた方法であることをDSF方法の他の注意点はある。 (SYPROオレンジ)を使用する蛍光標識は、疎水性であるので、場合によってはタンパク質に疎水性リガンドの結合と競合することができる。したがって、いくつかの場合において、得られた解離定数は、人為的に起因するラベルとの競争に昇温される可能性がある。しかし、多様なリガンドの比較、(の主な用途のためにDSF)、差異は、親和性による化合物のランキングに影響を与えるために十分に有意である可能性は低い。
この方法の潜在的な欠点を達成することができる検出限界である。原理的には、正確にタンパク質濃度の50%未満であり、この範囲内の偶数値が疑わしい精度であると思われるK dの値を測定することが可能であってはならない。範囲のこの端部における検出限界は、タンパク質と色素の濃度を低減することによって少し延長することができる一方で、機器の感度は、タンパク質濃度のさらなる低下を防止する。同様に、感度の上端は、リガンドの溶解性によって決定される。 K dの数学的にロバスト推定を得るためには、近似であるとリガンド濃度を必要とするリガンド結合形態中に存在するタンパク質の90%を使用してデータを取得するために最も重要であるLY十倍の K d(協同がないと仮定して)。検出限界は、したがって必然的に、関連するバッファ内のリガンドの溶解性の十分の一になります。これは方法の検出限界は、典型的には、タンパク質とリガンドに応じて、1μMmMの間、および1〜100の範囲であることを意味する。
結論として、示差走査型蛍光定量は、広範囲のタンパク質に適用可能で多目的な技術である。ここに提示される方法を使用して、迅速かつ安価に異なるリガンドに対するタンパク質の親和性を決定することが可能である。これは、特に小さな実験室で、タンパク質精製および安定化の応用、メタゲノムからの酵素の機能または特異性を解明するための大きな可能性を有しており、創薬における。
開示事項
The authors declare that they have nothing to disclose.
謝辞
This work was funded by grant from the BBSRC (grant number BB/H019685/1 and BB/E527663/1) to the University of Exeter.
資料
| Name | Company | Catalog Number | Comments |
| StepOne real time PCR instrument | Life Technologies | 4376357 | DSF can be performed with many other instruments. The StepOne instrument has very convenient software for data analysis. |
| Protein thermal shift software v1.0 | Life Technologies | 4466037 | |
| MicroAmp Fast optical 48-well plates | Life Technologies | 4375816 | |
| Optical sealing tape | Life Technologies | 4375323 | Bio-rad part no. 223-9444 is an alternative supplier |
| U-bottomed 96-well plates | Fisher | 11521943 | |
| SYPRO Orange | Life Technologies | S6650 | For a smaller volume supplier, use Sigma part no. S5692 |
| SPSS statistics version 20 | IBM | Other statistics packages will provide similar functionality | |
| GraphPad Prism 6.02 | GraphPad | Other statistics packages will provide similar functionality | |
| Hand applicator (PA1) | 3M | 75-3454-4264-6 | |
| Hexokinase from Saccharomyces cerevisiae | Sigma-Aldrich | H5000 | |
| Glucose | Fisher scientific | 10141520 |
参考文献
- Freyer, M. W., Lewis, E. A. Isothermal titration calorimetry: experimental design, data analysis, and probing macromolecule/ligand binding and kinetic interactions. Methods Cell Biol. 84, 79-113 (2008).
- Ladbury, J. E. Calorimetry as a tool for understanding biomolecular interactions and an aid to drug design. Biochem Soc Trans. 38, 888-893 (2010).
- Abdiche, Y., Malashock, D., Pinkerton, A., Pons, J. Determining kinetics and affinities of protein interactions using a parallel real-time label-free biosensor, the Octet. Anal Biochem. 377, 209-217 (2008).
- Pantoliano, M. W., et al. High-density miniaturized thermal shift assays as a general strategy for drug discovery. J Biomol Screen. 6, 429-440 (2001).
- Senisterra, G., Chau, I., Vedadi, M. Thermal denaturation assays in chemical biology. Assay Drug Dev Technol. 10, 128-136 (2012).
- Ericsson, U. B., Hallberg, B. M., Detitta, G. T., Dekker, N., Nordlund, P. Thermofluor-based high-throughput stability optimization of proteins for structural studies. Anal Biochem. 357, 289-298 (2006).
- Lo, M. C., et al. Evaluation of fluorescence-based thermal shift assays for hit identification in drug discovery. Anal Biochem. 332, 153-159 (2004).
- Nettleship, J. E., Brown, J., Groves, M. R., Geerlof, A. Methods for protein characterization by mass spectrometry, thermal shift (ThermoFluor) assay, and multiangle or static light scattering. Methods Mol Biol. 426, 299-318 (2008).
- Niesen, F. H., Berglund, H., Vedadi, M. The use of differential scanning fluorimetry to detect ligand interactions that promote protein stability. Nat Protoc. 2, 2212-2221 (2007).
- Geders, T. W., Gustafson, K., Finzel, B. C. Use of differential scanning fluorimetry to optimize the purification and crystallization of PLP-dependent enzymes. Acta Crystallogr Sect F Struct Biol Cryst Commun. 68, 596-600 (2012).
- Vedadi, M., et al. Chemical screening methods to identify ligands that promote protein stability, protein crystallization, and structure determination. Proc Natl Acad Sci USA. 103, 15835-15840 (2006).
- Davis, B. J., Erlanson, D. A. Learning from our mistakes: the 'unknown knowns' in fragment screening. Bioorg Med Chem Lett. 23, 2844-2852 (2013).
- Larsson, A., Jansson, A., Aberg, A., Nordlund, P. Efficiency of hit generation and structural characterization in fragment-based ligand discovery. Curr Opin Chem Biol. 15, 482-488 (2011).
- Scott, D. E., et al. Using a fragment-based approach to target protein-protein interactions. Chembiochem. 14, 332-342 (2013).
- Cimmperman, P., et al. A quantitative model of thermal stabilization and destabilization of proteins by ligands. Biophys. J. 95, 3222-3231 (2008).
- Matulis, D., Kranz, J. K., Salemme, F. R., Todd, M. J. Thermodynamic stability of carbonic anhydrase: measurements of binding affinity and stoichiometry using ThermoFluor. Biochemistry. 44, 5258-5266 (2005).
- Zubriene, A., et al. Measurement of nanomolar dissociation constants by titration calorimetry and thermal shift assay - radicicol binding to Hsp90 and ethoxzolamide binding to CAII. Int J Mol Sci. 10, 2662-2680 (2009).
- Weiss, J. N. The Hill equation revisited: uses and misuses. FASEB J. 11, 835-841 (1997).
- Cuccui, J., et al. Characterization of the Burkholderia pseudomallei K96243 capsular polysaccharide I coding region. Infect Immun. 80, 1209-1221 (2012).
- DeShazer, D., Waag, D. M., Fritz, D. L., Woods, D. E. Identification of a Burkholderia mallei polysaccharide gene cluster by subtractive hybridization and demonstration that the encoded capsule is an essential virulence determinant. Microb Pathog. 30, 253-269 (2001).
- Berman, H., Henrick, K., Nakamura, H. Announcing the worldwide Protein Data Bank. Nat Struct Biol. 10, 980(2003).
- Vivoli, M., Ayres, E., Beaumont, E., Isupov, M., Harmer, N. Structural insights into WcbI, a novel polysaccharide biosynthesis enzyme. IUCr Journal. 1 (1), 28-38 (2014).
- Sorrell, F. J., Greenwood, G. K., Birchall, K., Chen, B. Development of a differential scanning fluorimetry based high throughput screening assay for the discovery of affinity binders against an anthrax protein. J Pharm Biomed Anal. 52, 802-808 (2010).
- Uniewicz, K. A., et al. Differential scanning fluorimetry measurement of protein stability changes upon binding to glycosaminoglycans: a screening test for binding specificity. Anal Chem. 82, 3796-3802 (2010).
- Wan, K. F., et al. Differential scanning fluorimetry as secondary screening platform for small molecule inhibitors of Bcl-XL. Cell Cycle. 8, 3943-3952 (2009).
- Koropatkin, N. M., Holden, H. M. Molecular structure of alpha-D-glucose-1-phosphate cytidylyltransferase from Salmonella typhi. J Biol Chem. 279, 44023-44029 (2004).
- Paleskava, A., Konevega, A. L., Rodnina, M. V. Thermodynamics of the GTP-GDP-operated conformational switch of selenocysteine-specific translation factor SelB. J Biol Chem. 287, 27906-27912 (2012).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved