
Method Article
Diferansiyel Taramalı florimetri Kullanarak Protein-ligand Etkileşimleri belirlenmesi
Bu Makalede
Özet
Differential scanning fluorimetry is a widely used method for screening libraries of small molecules for interactions with proteins. Here, we present a straightforward method to extend these analyses to provide an estimate of the dissociation constant between a small molecule and its protein partner.
Özet
Yöntem çok çeşitli henüz protein arasındaki ayrışma sabiti belirlenmesi ve küçük molekülleri etkileşim için kullanılabilir. Ancak bunların çoğu uzman ekipmana ihtiyaç ve genellikle etkili güvenilir deneyler kurmak ve verileri analiz etmek için bir uzmanlık derecesi gerektirir. Diferansiyel tarama fluorimetry (DSF) artan ya da fizyolojik ortakları belirlemek için ya da vurma çıkarılması için küçük moleküller için etkileşen proteinlerin başlangıç taranması için güçlü bir yöntem olarak kullanılmaktadır. Bu teknik, nicel PCR için uygun olan ve en enstrümantasyon kurumlarda kullanılabilir böylece uygun bir PCR makinesi sadece gerektirmesi avantajına sahiptir; protokollerin mükemmel bir aralık zaten mevcuttur; ve yöntemin birden fazla kullanım için literatürde güçlü örneği yoktur. Geçmiş çalışma DSF verilerinden ayrışma sabitleri hesaplarken çeşitli araçları önerdi, ancak bu matematiksel talep ediyorlar. Burada, demlenmeyeonstrate DSF deney verileri orta düzeyde bir miktarının sapma sabitlerini hesaplamak için bir yönteme ilişkindir. Bu veriler, tipik olarak toplanmış ve bir gün içerisinde analiz edilebilir. Biz basit bir bağlama olaylardan toplanan verileri sığdırmak için nasıl kullanılabileceğini farklı model göstermek ve kooperatif bağlanma veya bağlama bağımsız siteler mevcut nerede. Son olarak, biz standart modeller geçerli değildir bir durumda veri analizi bir örnek sunuyoruz. Bu yöntemler, ticari olarak temin edilebilen kontrol proteinleriyle ilgili toplanan verilerin, ve araştırma programı iki protein ile gösterilmiştir. Genel olarak, bizim yöntem araştırmacılar hızla DSF kullanarak protein-ligand etkileşimleri içine daha fazla fikir edinmek için basit bir yol sağlar.
Giriş
Bütün proteinler, diğer çok büyük moleküllerin basit iyon diğer moleküllerin çeşitli bir yelpazede değişen afinitelere sahip, bağlayacaktır. Birçok durumda, proteinler (örneğin, kinaz ATP bağlanma) normal fonksiyon parçası olarak küçük moleküllü ortaklara bağlanmaktadırlar. Diğer etkileşimleri fonksiyonu ile ilişkili olabilir, ancak araçları (kristalleşme başarısını yükseltir, veya çözelti içinde proteinlerin muhafazasına yardımcı olacak şekilde proteinleri stabilize örneğin küçük molekül) deneysel olarak yararlıdırlar; önleyicileri olarak hareket eden ve böylece enzimin aktivitesini modüle edebilen aktif alanlarının ve proteinlerin allosterik sitelere bağlanan küçük moleküller iken.
Ortak protein molekülleri için afinitesini belirlemek için kullanılabilecek teknikler, geniş bir yelpazede bulunmaktadır. Izotermal titrasyon kalorimetre 1 bu tepkiler üzerine zengin bilgiler sunmaktadır yaygın olarak etiket ücretsiz, bir "altın standart" olarak görülüyor ve fırsatları sınırlıdırDeney rtifacts. Ancak, enstrümantasyon ve deneysel set-up otomasyon duyarlılığı son gelişmelere rağmen, hala protein gereksinimi açısından nispeten pahalı en iyi düşük-orta verim olmasıdır, ve orta ile etkileşimleri en uygundur yüksek afinite (Kd 100 uM ila 10 nM) 2'dir. Böyle kayn veya iki tabakalı enterforemetre 3 teklif daha throughputs ve hassasiyet elde etmiş gibi diğer etiket ücretsiz yöntemler 100 Da gibi düşük küçük moleküllerin tespit. Ancak, bu yöntemler için yüksek verimlilik araçları nispeten pahalı, ilgili projelerin sürekli bir verimlilik olacak, sadece nerede haklı, ve pek çok akademik laboratuvarlara erişilemez olması muhtemel olan.
Diferansiyel tarama fluorimetry (DSF veya thermofluor) birinci ilaç keşfi için bir yöntem olarak 2001 4'te tarif edilmiştir. Bu metho olarakd, proteinleri bir floresan boya ile kuluçkalanır (başlangıçta naftalen-sülfonik boyalar kullanılmıştır), proteinlerin hidrofobik bölgelerine bağlanması üzerine flüoresanını değiştirir edilmektedir. Protein boya Numune daha sonra ısıtılır ve ısı arttıkça floresan izlenir. Proteinin, açılma ve proteinin hidrofobik parçaları maruz kalma sıcaklığı (Şekil 1A) bir fonksiyonu olarak floresansta bir karakteristik örneğini ortaya çıkarmaktadır. Deney, herhangi bir ticari kantitatif PCR cihazda küçük hacimlerde gerçekleştirilebilir ve bu nedenle tek bir deneyde, bir numune aynı anda çok sayıda (enstrüman modeline bağlı olarak, genellikle 48, 96 ya da 384 örnekleri,) test edilebilir. Deneyler genellikle örnekleri 5 yüksek kapasiteli analiz etme olanağı sağlayan bir saat civarında gerçekleştirilebilir.
Metodolojiye daha fazla iyileştirmeler, daha iyi spektral özellikleri 6,7 ile boyaların benimsenmesini gördük , genel veri analizi için araçlar ve ilk tarama 8,9 protokolleri önerdi. Yöntemin uygulama yelpazesi hazırlanması ve proteinlerin 10 depolanması için en iyi koşulları oluşturulması ve kristalleşme 11 yardım için potansiyel bağlanma ortakları belirlenmesi üzerine özel bir odaklanma ile, genişletilmiştir. Yöntemin, nispeten yüksek miktarda protein, görece düşük maliyetli bir (~ 2 reaksiyon başına ug) ve zayıf bağlanma molekülleri okuyan uygulanabilirliği Akademik bağlamda, özellikle 12-14, DSF parçası bazında ilaç tasarımı için değerli bir araç yaptı.
Protein-ligand etkileşimleri okuyan DSF geniş uygulama olmasına rağmen, az sayıda çalışma bu çalışmalardan ayrılma sabitlerinin belirlenmesini tarif etmişlerdir. Ancak, bu seyrek veri veya bazı durumlarda e takılmalıdır birçok parametre ile, proteinin açılımı anlatan detaylı denklem üretmek eğilimi var7,15-17 stimated. Bu yöntemler, örneğin olağandışı geçişleri gösteren sıkı bağlayıcı bileşikler, ya da proteinler olarak, zor durumlarda özellikle önemlidir. Ancak, birçok laboratuvarlar için, bu ayrıntılı analizler rutin kullanım için çok zahmetlidir. Bu nedenle, farklı senaryolar için alternatif tedaviler öneriyoruz, ve bu farklı protein-ligand etkileşimleri kaynaklanan veri sığdırmak için nasıl kullanılabileceğini göstermektedir. Bizim yöntem ısmarlama veri analiz yazılımı mevcut olduğu için StepOne QPCR enstrümanı kullanır; bu, veri analizi hızlandırır iken, diğer cihazlarla elde edilen sonuçlar, daha önce yayınlanmış yöntemler kullanılarak 9 işlenebilir ve aynı alt analizi gerçekleştirilebilir.
Protokol
(Bir mertebe dahilinde yani) ayrışma sabiti için bir Yaklaşık Değer 1. Belirlenmesi
- Tablo 1 'de ayrıntılı olarak açıklanan bir karışım hazırlayın.
- Mümkün olan en yüksek konsantrasyonda söz konusu ligandın stokları hazırlanması ve bu daha sonra en altı ile on kez seyreltilmiş hali. Yaklaşık Kd önceki veri bilindiği durumda, en az iki konsantrasyonları yukarıda ve Kd altında olması hedeflenmektedir.
- Bir kısım QPCR plaka sekiz kuyulara karışımı 18 ul. Birinci oyuğuna çözücünün 2 ul ekleyin. Kalan yedi kuyuda olarak her biri ligand seyreltme serisi (adım 1.2) her bir üyesi, 2 ul ekle.
- Tablada QPCR sızdırmazlık yerleştirin. Levhanın iyi bir sızdırmazlık elde etmek için, plakanın ortasında (özel reaktiflerin tabloya bakınız) bir el aplikatörü yerleştirin. Bir tarafa mühür aşağı pürüzsüz, ve daha sonra diğer yarısı üzerinde tekrarplakası.
- Hava kabarcıklarını çıkarmak için iki dakika boyunca 500 xg'de plaka santrifüj.
- Bir StepOne QPCR enstrüman plakası yerleştirin. (Bu 40 dakika boyunca 99 ° C bir rampa ardından 25 ° C'de 2 dakika duraklama, ve daha sonra 2 dakika duraklama sağlar) "Erime eğrisi" seçeneği, ROX filtreleri seçin, ve hızlı rampa hızı seçin. Termal denatürasyon çalıştırın.
NOT: bir çalışma gerçekleştirmek için komut dosyaları http online olarak mevcuttur: // www.exeter.ac.uk/biosciences/capsular. - Alet koşmak sonunda, ekranda "Analiz" butonuna tıklayınız. Sonuç dosyayı kaydedin.
- Protein Termal Shift yazılımını açın.
- Yeni bir çalışma oluşturun; özellikler sekmesinde, bu bir ad verin ve Koşullar sekmesinde, detay ligandlar.
- Deney Dosyaları sekmesinde taşı, ve kaydedilen sonuçlar dosyası (XXX.eds) ithal ve her iyi (şablon fil içeriğini ayarlamakes) yazarlarda mevcuttur.
- Analiz sekmeye geçmek ve "analiz" düğmesine basın.
NOT: Bu sonuçları analiz eder. Bu Excel İhracat sekmesini kullanarak daha fazla araştırma için sonuçları ihracat mümkündür. Sonuçlar bir sekme tasvir biçiminde ihraç edilmektedir. Excel'de ihraç dosyasını açın ve hemen Excel formatında kaydetmek için en iyisidir.
- Sadece çözücü mevcudiyetinde protein Şekil 1A 'de gösterilene benzer bir sonuç verir olduğunu kontrol edin. Sonra, "suret" bölmesinde sonuçlarında gözlenen erime sıcaklıkları inceleyin. Bu artan ligand konsantrasyonu ile erime sıcaklığı belirgin bir artış gösterdiğinden emin olun.
Not: erime sıcaklığı, ligand içermeyen protein arasındaki yarım olduğu İdeal olarak, bu (protein tam ligand bağlı olduğunu varsayarak) açık bir maksimum erime sıcaklığı ve yaklaşık Kd sağlayacaktırmaksimum.
Ayrışma sabitinin belirlenmesi için 2. Deney hazırlığı
- Bir ana karışımı olarak Tablo 2'de ayrıntıları verilen bir karışım hazırlayın.
- Son deneyde on kat seyreltilerek on beş farklı konsantrasyonlarda stok ligand hazırlayın. İdeal olarak, konsantrasyonlarının yaklaşık Kd altında ve üstünde büyüklük en az iki sıra içerir ve tahmin edilen Kd konsantrasyonlarını merkezi. Bu iki tarafında başka dört puan ile, tahmini K d büyüklükte bir düzen içinde noktaların yedi odaklanın; bir seçim varsa, doyurarak değerlere daha fazla puanı bulunur.
NOT: Gerekli, bu ligand stokları ligand çözünürlüğünün sınırlayıcı çift deneysel konsantrasyonda olduğu deneysel koşullar böyle değiştirmek mümkün değilse. - Master'ın 120 ul ekleyinana karışımı elverişli bir şekilde sevk için bir rezervuar olarak hareket etmek, U-tabanlı 96 oyuklu bir plaka içerisinde sekiz çukurlara karıştırın. PCR plakasının bir kolonuna 18 ul dağıtılması için 8 kanallı pipet kullanın. Plaka üzerinde 8 x 6 deseninde 48 dolu gözenekli bir toplam elde etmek üzere bir başka beş sütun için tekrarlayın.
- U-tabanlı 96 oyuklu bir plaka içerisinde, ayrı ayrı kuyucuklarına, 20 ligand stokların ul, veya solventin ekleyin. 8 kanallı pipet, aspire sekiz farklı bağlayıcı hisse senetleri (veya çözücü) 2 ul kullanma. Adım 2.3 master karışımı ile dolu PCR plaka bir sütun bu ekleyin. Iki başka sütunlar için aynı sekiz ligand / çözücü stokları ile tekrarlayın. Aspire 2 Geriye kalan sekiz ligandı ya da çözücü stoklarının ul, ve plakanın bir dördüncü sütunda bu ekleyin. Iki başka sütunlar için bu işlemi tekrarlayın. Bu, tüm 16 ligand ve çözücü numuneler için üç nüsha örnekler vereceğim.
- Plaka üzerine yerleştirin QPCR conta (adım 1.4).
- 500 xg'de plaka santrifüjeİki dakika.
- QPCR enstrüman plakası yerleştirin. Adım 1.6 belirtilen parametreleri kullanarak bir termal denatürasyon çalıştırın.
- Alet koşmak sonunda, ekranda "Analiz" butonuna tıklayınız. Sonuç dosyayı kaydedin.
- Protein Termal Shift yazılımını açın. Yeni bir çalışma oluşturun; özellikler sekmesinde, bu bir ad verin ve Koşullar sekmesinde, detay ligandlar.
- Deney Dosyaları sekmesinde taşı, ve kaydedilen sonuçlar dosyası (XXX.eds) ithalat, ve de her içeriğini ayarlayın.
NOT: şablon dosyalarını http online olarak mevcuttur: // www.exeter.ac.uk/biosciences/capsular. - Analiz sekmeye geçmek ve "analiz" düğmesine basın.
- Triplicates olarak sonuçlarını göstermek için ekranın sol tarafındaki menüden "çoğaltır" sekmesini seçin. Kopyaların nasıl sıkı dayalı verilerin güvenilirliğini değerlendirmek. ShoulTriplikat yoksul tekrarlanabilir olduğunu göstermek d yakından ham verileri incelemek.
- Erime sıcaklığını değerlendirmek için Boltzmann'ı veya Türev yöntemleri kullanılarak verileri analiz edin. "Sonuçları Replicatea" sekmesini seçin ve "tarafından Plot:" geçiş, "sonuç arsa Replicatea" in arasındaki düğme "Tm - Boltzmann" ve "Tm - Türev". Numune için büyük üretkenlik sağlar yöntemini seçin. Excel Export sekmesini kullanarak daha fazla araştırma için sonuçları ihracat.
NOT: Birden fazla geçişler göstermek numuneler için, birden fazla eriyik modunda Türev yöntemi kullanmak hemen hemen her zaman en iyisidir. Sonuçlar bir sekme tasvir biçiminde ihraç edilmektedir. Excel'de ihraç dosyasını açın ve hemen Excel formatında kaydetmek için en iyisidir. - Sonuçların yeniden üretilebilirlik sağlamak için, ayrı bir günde bir tekrarı dahil olmak üzere, en az iki kez deney tekrarlayın. Veri analizi (aşağıya bkz: adım 3) ShouldKd değeri, ilk tahmin etmek için önemli ölçüde farklı olduğunu göstermektedir, yaklaşık Kd değerleri iyi bir dizi sağlamak için (adım 2.2) uygun olarak, ligand konsantrasyonlarının değiştirilmesi.
3. Veri Analizi renatürasyonun altında ayrışma sabiti belirleme
- Ligand konsantrasyonlarının Excel'de bir tablo ve erime sıcaklığını oluşturun.
- GraphPad Prism yazılımı açın ve bir XY tablo oluşturun. Ligand konsantrasyonu ve sıcaklık sonucu erime için Y sütun için X kolonu kullanılarak, veri girin.
NOT: Gösterilen bir örnek Şekil 1B olduğunu. Önceden yüklenmiş denklemler ve SPSS istatistik paketi kullanılarak alternatif talimatları ile bir komut dosyası, http online olarak mevcuttur: // www.exeter.ac.uk/biosciences/capsular. - Analiz sekmesinde, analiz parametresini değiştirmek için seçeneği seçins (Ctrl + T). , Doğru modeli giriniz "Yeni" seçin ve "yeni bir denklem oluşturma" için. "Tek sitesi ligand bağlanma" olarak Tablo 3'te detaylı denklemi yerleştirin.
NOT: Bu adımların bir örneği Şekil 1C gösterilmiştir. Denklemleri önceden yüklenmiş ile komut kullanırken, ilgili denklem listede yerine girdi seçilebilir yönlendirilebilir. Bu denkleminin ekte verilmiştir. - Kutusunda "Başlangıç Değerlere için Kurallar" seçin ve Tablo 3'te ayrıntılı olarak başlangıç değerleri için kurallar girin.
- (Ligand verilmektedir aynı birimlerinde) ve protein nihai konsantrasyona girmek "ile sürekli eşit" olarak, parametre P sınırlandırmaktadır.
- Analizi gerçekleştirmek için Tamam seçeneğini seçin.
NOT: Bu adımların bir örneği Şekil 1D gösterilmiştir. Grafik yazılımı veri ve modele uyum gösteren bir figür oluşturur. T örnekleriöz konusu analizler temsilci verileri gösterilmiştir.
Kooperatif Modelleri 4. Uydurma Veri
Basit bir kooperatif modeli, ya da iki ayrı ayrılma sabitler tanımlanmıştır model seçmesini, bir kooperatif modeli veri sığdırmak için. Birinci yaklaşım, negatif işbirliğinden durumunda tercih edildiği veya ilk araştırma olarak edilir. Ancak, prensipte iki farklı ayrışma sabiteleri 18 modellemek için pozitif bir birliktelik durumlarda daha iyidir. Bu durumda, ardışık modelleme ligand bağlanma ya da ligandların bağlanma ya da bağımsız olarak varsayarak devam edebilir.
- , "Basit kooperatif modeli" olarak listelenen Tablo 3 denklemlerin birini yerleştirin, adım 3.3 de, Ancak protokolde 3. olarak "iki ligandların Sıralı bağlama" aynı ilk adımları uygulayın, veya 18 "Bağımsız iki ligandların bağlanması".
- Başlangıç değerleri için ilgili kuralları seçinTablo 3'te bu denklemlerin her biri ile ilişkili.
- Verilere modelin uyum inceleyin. Veri kötü uygun olmalıdır, başka bir model düşünün.
NOT: ayrıca dikkatle Protein Termal Shift yazılımıyla (adım 2.9) ile veri erime sıcaklığına takılmasını incelemek önemlidir: bazen iyi sonuçları almak için buraya parametrelerini değiştirmek için gereklidir. K d iki tarafında sınırlı bir veri seti, ya da (özellikle yüksek ligand konsantrasyonlarda) tek bir anormal noktası, ya önemli ölçüde etkileyebilir: Başka bir göz veri noktalarının aralığı ideal ve herhangi bir anormal noktaları olup olmadığını olmadığıdır Sonuçlar. - Yeniden üretilebilirlik sağlamak için en az iki kez (adım 2.12) deney tekrarlayın.
Erime Sıcaklığı İkili kaydırır gösteriliyor Curves 5. Uydurma Veri
Bazen, yerine ligand bir derecelendirilmiş cevaptan daha, proteinler olmuşturÖrnek açık bir şekilde bağlanmış, bağlanmamış numune ayrılır bir ikili yanıtı kabul görülmektedir. Bir örnek, temsili sonuçlar (Şekil 4) sağlanır. Bu durumda, erime sıcaklıklarının montaj Kd için iyi bir uyum sağlamaz.
- Protein Termal Shift yazılım ham veri çıkışı ihracat. Her sıcaklık noktası için, sıfır ligand için ortalama floresan ve en yüksek ligand konsantrasyonlarının hesaplanması. Bu yanında, her bir veri noktasından sonuçları çizelgeye.
NOT: Burada oluşturulan hata monte erime sıcaklıklarında hata daha azdır. - SPSS istatistik paket açın. SPSS bir veri pencerede her bir deney için sıcaklıkları, iki ortalama veri setlerini ve verileri kopyalayın. Değişken sekmesinde, "düşük" olarak ligandsız için ortalama veri kümesi ve "yüksek" olarak en yüksek ligand konsantrasyonu için ortalama veri kümesi ayarlayın.
- Mevcut sözdizimi dosyasını indirinonline olarak http: // www.exeter.ac.uk/biosciences/capsular . "Çalıştır → Çalıştır tüm" seçiniz.
- İlgili ligand konsantrasyonlarında, yeni bir Excel çalışma kitabına orantılı bağlı sonuçları kopyalayın.
- Graphpad yazılımını açın ve bir XY tablo oluşturun. Ligand konsantrasyonu ve sıcaklık sonucu erime için Y sütun için X kolonu kullanılarak, veri girin. Analiz sekmesinde, "değişim analizi parametreleri" seçeneğini seçin. , Doğru modeli giriniz "Yeni" seçin ve "yeni bir denklem oluşturma" için. "Erime sıcaklığı ikili vardiya Analizi" olarak listelenen Tablo 3'te verilen denklemi girin.
- "İlk Değerler Kuralları" kutusunu seçin ve Tablo 3'te detaylı başlangıç değerleri için kurallar girmek. "Için Sabit eşit" olarak, parametre P sınırlama ve liga gibi aynı birimlerde (protein nihai konsantrasyonu girinnd) verilmiştir.
NOT: 3. bölümde protokol için bu kutucukları doldurulduktan örnekleri Şekil 1C, D gösterilmiştir. - Iyi bir uyum varsa, sonuçlar sonsuz ligand konsantrasyonunda beklenen sonuca ekstrapolasyonuyla geliştirilebilir. Her bir ligand konsantrasyonu bağlanmış olan oranının model, ligand konsantrasyonu ve en yüksek değeri değeri inceleyin. Bu 0.99 veya daha büyükse, daha fazla analiz sonuçları iyileştirmek mümkün değildir.
- 0,99 oranı daha az ise, ilave bir aşama, en yüksek ligand konsantrasyonu numunede bağlanmamış proteinin etkilerini gidermek için gereklidir. Aşama 5.2 'de, (başka bir hücre kullanılabilir, ve R2'nin Tablo 3'te denkleme uygun şekilde değiştirilmesi) hücre R2 içinde (adım 5.7), en yüksek ligand konsantrasyonu noktasında bağlanmış ligand oranını mal. Yüksek ligand konsantrasyonu sonuçlarının sonra ortalama fazladan bir sütun oluşturun. FirsT hücresi "lıgand konsantrasyonunu sonsuz uyarlanabilme" olarak Tablo 3'te listelenen denklemi kopyalama. Bu sütundaki kalan hücrelere bu formülü kopyalayın.
NOT: Bu hesaplama, en yüksek ligand konsantrasyonuna bağlı olmayan proteinin etkisini ortadan kaldırır. Ligand serbest protein ve en yüksek ligand konsantrasyonu arasındaki fark, her bir sıcaklık noktasında tamamen bağlanmış ve bağlanmamış proteinin durumlar arasında beklenen farkı sağlamak için yüksek ligand konsantrasyonu bağlanmış olan oranının tersi ile çarpılır. Bu fark, ilave ya da tam ligand bağlı proteini için beklenen floresan vermek üzere bağlanmamış durumda çıkartılmaktadır. - Bu yeni sütun ile SPSS veri sayfasında maksimum ligand konsantrasyonu için sütun değiştirin ve veri uydurma tekrarlayın.
NOT: - model maksimum konsantrasyon derecesinden bağ bağlanmış olan orantılı bir başka önemli bir değişikliği ortaya çıkardığı takdirde 5.9 tekrar edilmesi gerekebilir 5.7 adımları(Bu durumda eğer, muhtemelen dahil daha yüksek bir ligand konsantrasyonu noktası deneyi tekrar ideal olacaktır) üzerine. - Protein, bir ikili kaydırma ve görüntüler kooperatif davranışı gösterir durumda, denklem aşama 4,1 olanlar ile ikame edilmelidir aşama 5.5 önerdi. "Top" ve "Bottom" parametreleri sırasıyla 1 ve 0 ile değiştirilmesi gerekir.
- Yeniden üretilebilirlik sağlamak için en az iki kez (adım 2.12) deney tekrarlayın.
Sonuçlar
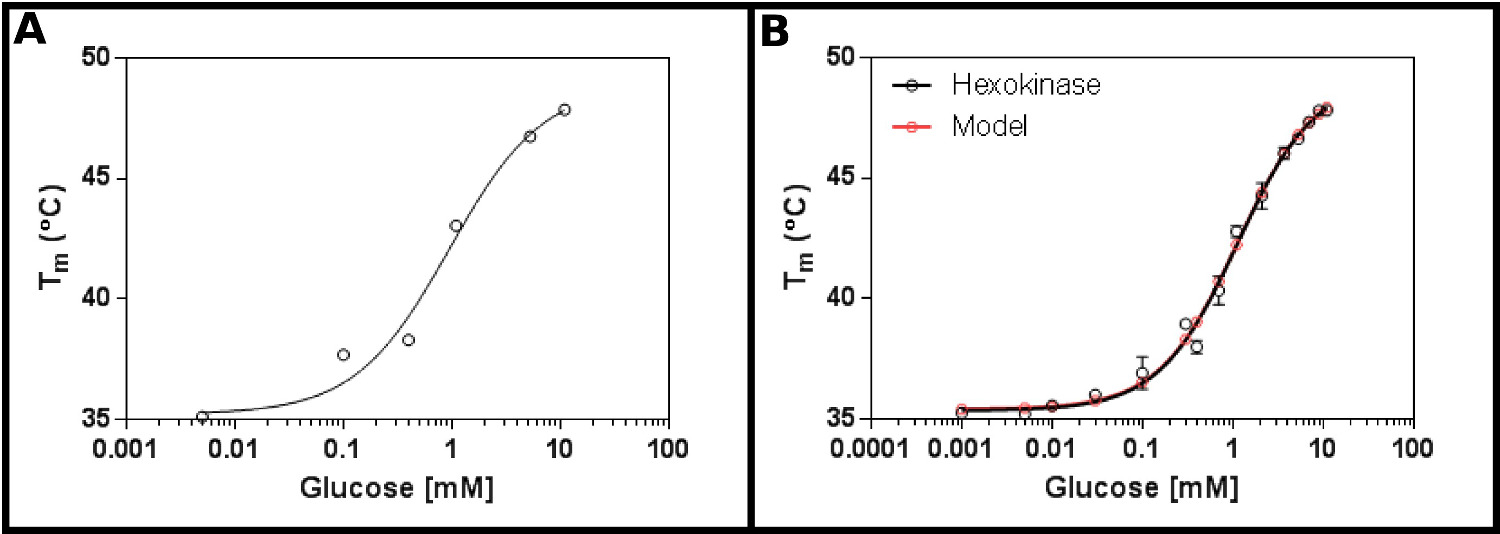
Bu yöntemin için mükemmel bir test substrat Heksokinaz olduğunu. Bu hali hazırda ticari olarak mevcut olan ve en çok laboratuvarda bulunan ve açık deneyde, yeniden üretilebilir sonuçlar elde edilmiştir, iki maddenin sahip olma avantajlarına sahiptir. Bir başlangıç konsantrasyonu ekranı (Protokol 1), heksokinaz ve glükoz (Şekil 2A) ile, büyük olasılıkla Kd 0.2 ila 1.7 mM aralığında olacağını göstermektedir. Bu nedenle, daha büyük bir ekran (Protokol 2), Tablo 4. Sonuçlar (Şekil 2B) 'de gösterilen konsantrasyonu kullanılarak, gerçekleştirildi tek yerli ligand bağlanma denklemine iyi bir uyum gösterir (Protokol bölüm 3.3) [9] ve verdi K 1.2 ± 0.1 mM d.
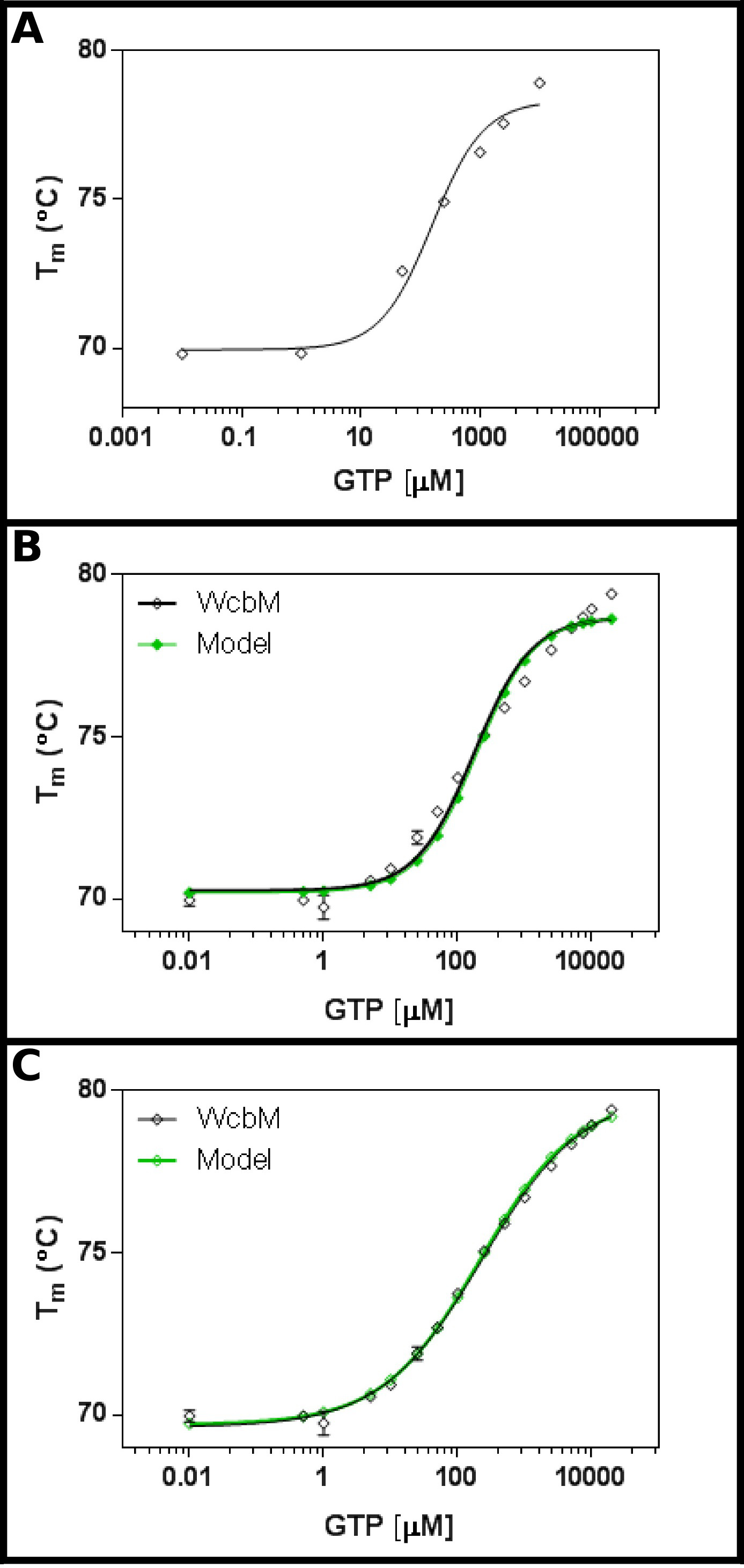
Varsayılan heptose-guanil transferaz WcbM 19,20 GTP (Şekil 3A) bağlanabilen güçlü bir ısı kayması göstermektedir. Bir başlangıç ekranı önerdi Kd yaklaşık 100 uM aralığında olacaktır. . Nedenle, tam ekran 3.3 denklem sonuçların takılması Tablo 5'te gösterilen konsantrasyonlarda kullanarak, ayarlanmış bir makul uyum (0,981 arasında R 2, Şekil 3B) gösterdi .Ancak, veri ve arasında belirgin bir fark yoktur modeli farklı bir denklem gerekli olduğunu düşündürmektedir. WcbM dizisi ile protein veri bankası 21 ara yapılar bir şekilde dimerler belirlenmiş olan en yakın homologlarını gösterdi. Verileri, bu sebepten, kooperatif sıralı ve iki ligand (Protokol 4) bağlanması için bağımsız üç denklemler kullanılarak analiz edildi. Sıralı ve bağımsız bağlama modelleri hem 0,992 bir R2 değeri ve 0.480 ve 0.461 bir Sy.x verdi ise bir kooperatif modeli için uydurma istatistikler, bir R 2 0,998 değerini ve 0.215 artıkları (Sy.x) standart sapma verdi sırasıyla. Bu modeli önerirkooperatif modelinin veri için en uygun olan vermiştir: Burada, 230 ± 10 uM altındaki bir KD ½ gözlenmiştir, 0.52 ± 0.02 (Şekil 3C) arasında bir n değerine sahip. Bu bağlanma için negatif bir birliktelik göstermektedir. Kd için bahsedilen birimler, oldukça tatmin edici uM 0,52 olacak şekilde K ½ yerine Kd daha bu durumda kullanılan dikkat etmek gerekir.
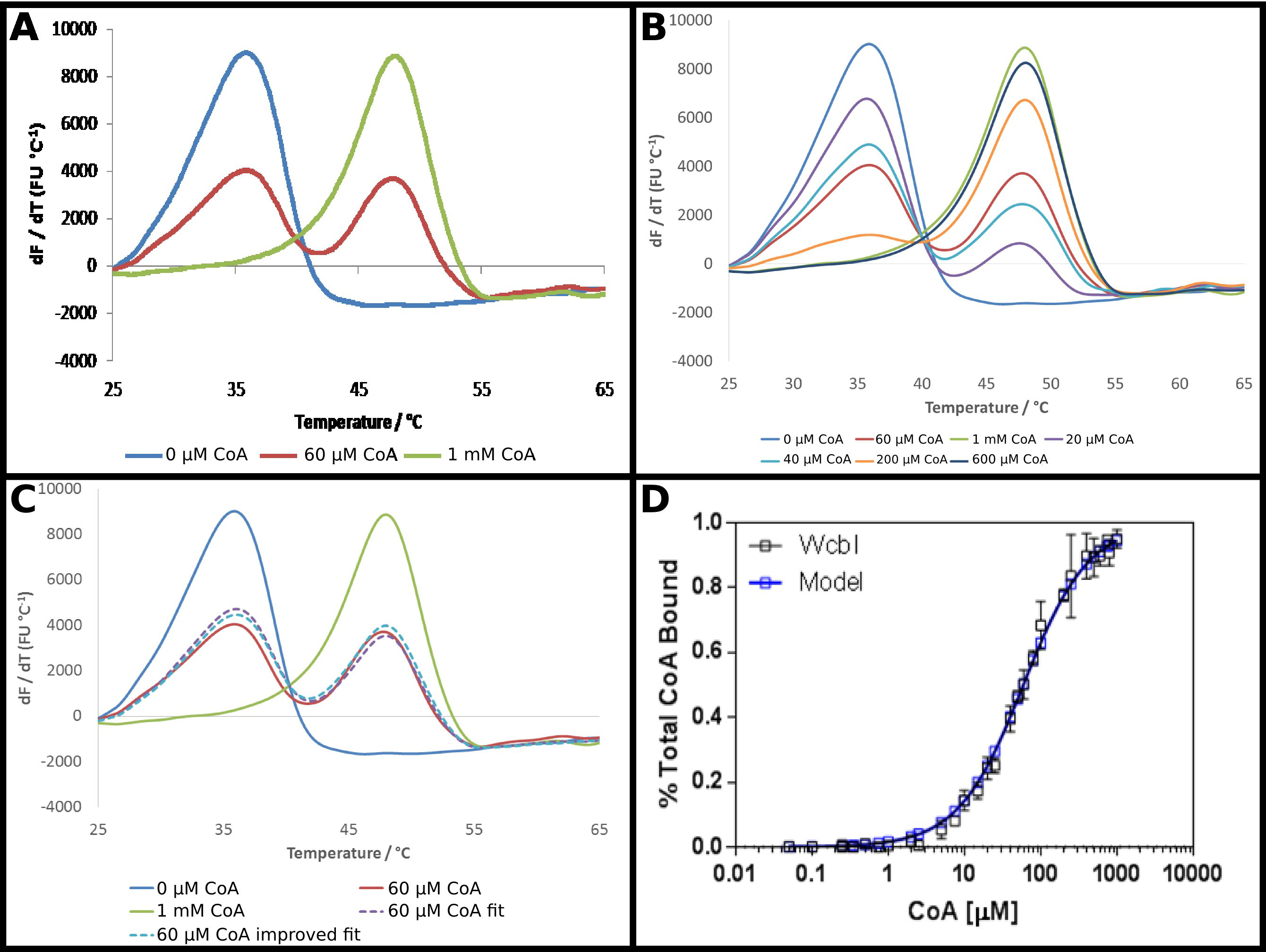
Varsayılan GDP-6-deoksi-β-D-manno -heptopyranose 2 -acetylase, O, WcbI 22, diferansiyel tarama florimetri oldukça sıradışı bir sonucunu göstermektedir. Herhangi bir ligand olmadığında, açık ve basit bir denatürasyon (Şekil 4A) göstermektedir. Protokolde tarif edildiği gibi koenzim A (CoA) DSF kullanarak bu proteinin bir ligand ve bu ortak proteinin afinite olarak tespit edilmiştir araştırılmıştır. CoA'nın yüksek konsantrasyonlarda güçlü bir in mevcudiyetindedaha yüksek bir sıcaklığa hift 15 ° C erime sıcaklığında bir değişiklikle gözlenmektedir. Bununla birlikte, orta konsantrasyonlarda, daha çok bir ara erime sıcaklığında bir tek fazlı erimesine kayması daha WcbI protein ligand içermeyen bir sıcaklıkta, ya tam olarak bağlı erime sıcaklığı (Şekil 4A), ya da eriyik azalmasıyla, iki fazlı bir erime göstermiştir . Daha yüksek bir sıcaklıkta (Şekil 4B) de eritilmiş oranı artan substrat konsantrasyonları artan, doza bağlı bir şekilde değişmiş iki türün oranları. Bu verilerin doğrudan analizi zor oldu: türev yöntemleri iki erime olayların meydana geldiği, ancak artan ligand konsantrasyonu ile bir değişiklik gösteren yardımcı olmadığını vurgulanan iken Boltzmann denklemine uydurma, çok kötü uyuyor verdi.
Bu verileri analiz etmek için bir daha az geleneksel bir yaklaşım, bu nedenle (Protokolü 5) kabul edildi. Floresan dbağ olmadan ve yüksek bağ konsantrasyonda erivative sonuç, esas olarak düşük bir erime sıcaklığına tüm protein ya da daha yüksek erime sıcaklığı durumunu temsil olarak alınmıştır. Kalan türevi veriler birlik (Şekil 4C) toplanarak oranda, bu iki devletin her bir oranının toplamı olarak her noktada takıldı. Elde edilen veriler, daha sonra önce olduğu gibi aynı denklemler kullanılarak, bir görünür Kd elde etmek için daha önce olduğu gibi yerleştirilmiştir. Bu "yüksek" bağ noktası yalnızca 95% ligand bağlı olması muhtemel olduğunu vurguladı. Veriler daha sonra,% 100 bağlanan protein için sonuç tahmininde bir ekstrapole edilmiş ve veriler, 58 ± 2 uM kadar bir görünür Kd elde etmek için tekrarlanır donanımıdır. Bu bağlanma modeli (Şekil 4D) için deneysel sonuçların mükemmel bir uyum sağlanır.
fo: İçerik-width = "5in" src = "/ files / ftp_upload / 51809 / 51809fig1highres.jpg" width = "500" />
Deneme set-up ve analizi 1. Örnekler Şekil. (A), bir termal denatürasyon profili, beklenen şekli Örneği (maya, heksokinaz için veri alınmıştır). Ham veri karakteristik şekli sığ bir düşüş takip maksimum floresan ilerleyen bir artış, (9 içinde daha detaylı olarak tartışılacaktır) göstermektedir. Bu floresan birinci türevinin tek bir tepe eşlik eder. GraphPad veri girişi (B) Örnek. Ligand konsantrasyonu x ekseni üzerinde verilir ve Y-ekseni üzerinde erime sıcaklıkları görülmektedir. GraphPad denklem tanımı (C) Örnek. (D) Örnekler doğru ve protein konsantrasyonu tespit değişkenlerin başlangıç değerlerinin ayarlanması, ayrışma sabitinin doğru tespitine imkan vermek.m / files / ftp_upload / 51809 / 51809fig1highres.jpg "target =" _blank "> Bu rakamın büyük bir versiyonunu görmek için lütfen buraya tıklayınız.

. Diferansiyel tarama florimetri ile ölçülen glikoz ile heksokinazın Şekil 2. etkileşimi (A), glukoz konsantrasyonları, geniş bir dizi test İlk deneyde, Kd 0.2 aralığında olması muhtemel olduğunu -. 1.7 mM (B) 'ayrıntılı deney, glukoz 16 konsantrasyonlarını test, 1.12 ± 0.05 mM olarak görünür K d belirlenmesini sağlar. Veri (35.4 ± 0.2 oC ve 49.3 ± 0.5 ºC sırasıyla uydurma alt (T1) ve üst (T2) sıcaklıkları ile) tek bir bağlanma olay için model son derece iyi uyuyor. DikkatBu veriler, 10 mM MgCl2 mevcudiyetinde toplanmıştır. Bu görüntüler GraphPad kullanılarak hazırlanmıştır. , bu rakamın büyük bir versiyonunu görmek için lütfen buraya tıklayınız.
{kind=link}

GTP ile WcbM Şekil 3. Etkileşim bir anti-kooperatif bağlanma ortaya koymaktadır. (A), GTP konsantrasyonları, geniş bir dizi test İlk deneyde, Kd 200 aralığında olması muhtemel olduğunu -. 500 uM GTP 16 konsantrasyonlarda test (B), ayrıntılı bir deneydir, belirgin bir değer göstermektedir 120 ± 20 mcM K d. Ancak, logaritmik ölçek x-ekseni için kullanıldığı zaman, önemli bir discre oradamodeli ve veri arasında Pancy. bir kooperatif modeli ile aynı verilerin (C) Analizi basit kooperatif modeli kullanılmıştır verilere mükemmel bir uyum gösterir. Burada, K 230 ± 20 mcM ½ (sırasıyla 69.63 ± 0.06 º C ve 79.9 ± 0.1 ºC uydurma alt (T1) ve üst (T2) sıcaklıkları ile) kooperativiteye katsayısı n = 0.52 ± 0.02 ile tespit edilmiştir. WcbM dimerik gibi göründüğü için, bu enzim, kendi GTP bağlanma mükemmel anticooperative olduğunu ima eder. Bu görüntüler GraphPad kullanılarak hazırlanmıştır. , bu rakamın büyük bir versiyonunu görmek için lütfen buraya tıklayınız.
{kind=link}

Şekil 4. WcbI iki fazlı bir erime modelini göstermektedirligandı koenzim A (CoA) mevcudiyetinde. (A) 'WcbI ligant (mavi) yokluğunda, basit bir tek fazlı bir erime modelini göstermektedir. Yüksek ligand konsantrasyonlarda (1 mM; yeşil çizgi), benzer bir model görülmektedir. Bununla birlikte, ara bağ konsantrasyonlarda mevcuttur. (60 uM; kırmızı çizgi), iki farklı erime piki ligand içermeyen ve ligand-bağlı reseptör durumuna karşılık gelen görülmektedir (B), zirvelerin iki grup arasındaki geçiş doza bağımlı tam karşısında konsantrasyonlarının aralığı. ligand serbest ligand ve yüksek bir oranda bir toplamı olarak iki fazlı ergime (C) Modelleme (kırmızı çizgi ile karşılaştırıldığında, kesikli çizgi mor), bir veri iyi bir uyum sağlar. Bu uyum tam doluluk (kesikli mavi çizgi) ile (model göstermektedir ~% 95 doluluk) yüksek ligand konsantrasyonu için gözlemlenen sonucu ekstrapolasyonuyla geliştirilir. (D) CoA'dan Sho bağlı WcbI oranı için elde edilen veriler WS 58 ± 2 uM bir Kd ile basit bağlanma modeline, mükemmel bir uyum için (bu veriler sonuçların birinci kümesine bağlı olarak ikinci gün için seçilen biraz farklı ligand konsantrasyonları, iki ayrı günde toplanan verileri temsil eder). Paneller. - Graphpad kullanan (A C) Excel kullanılarak hazırlanmış ve paneli (D) verildi , bu rakamın büyük bir versiyonunu görmek için lütfen buraya tıklayınız.
{kind=link}
İlk deneyler için Tablo 1. Tarif.
| Reaktif | Karışımı hacmi (ul) |
| Protein | 0.11 mg / ml nihai konsantrasyona |
| 0.3 | |
| , 0.5 M HEPES pH 7.0 | 3.7 |
| 5 M NaCI | 5.6 |
| Su | 180 ul |
Bu protokol, bölüm 1. Bu tampon karışımı genel proteinler için uygun olduğu tarif edildiği gibi, Kd bir tahminde bulunmak için bir ilk keşif deney için protein, saptama reaktif ve tampon A "ana karışımı" açıklar. Önceki sonuçlar diğer tamponlar kullanılmalıdır önermek durumlarda, bu ikame edilmelidir. Stok protein düşük bir yoğunlukta (yani, en az 0.3 mg / ml) ise, ilave tampon miktarını azaltmak için gerekli olabilir, protein numune tamponu içinde halihazırda mevcut olan telafi etmek için ilave edildi.
K d belirlenmesi için Tablo 2. Tarif.| Reaktif | Karışımı hacmi (ul) |
| Protein | 0.11 mg / ml nihai konsantrasyona |
| 5,000X SYPRO Turuncu | 1.78 |
| , 0.5 M HEPES pH 7.0 | 22.2 |
| 5 M NaCI | 33.3 |
| Su | 180 ul |
Bu protein, saptama reaktif maddesi ve b "ana karışımı" nı tarifuffer bir protein numunesi için Kd tam tayini için, protokol bölüm 2'de tarif edildiği gibi, genel tampon karışımı proteinler için uygundur. Önceki sonuçlar diğer tamponlar kullanılmalıdır önermek durumlarda, bu ikame edilmelidir. Stok protein düşük bir yoğunlukta (yani, en az 0.3 mg / ml) ise, ilave tampon miktarını azaltmak için gerekli olabilir, protein numune tamponu içinde halihazırda mevcut olan telafi etmek için ilave edildi.
Tablo 3. Denklem ve veri analizi için parametreleri.
d| Deney protokolünde Aşama | Denklem gerekli | Parametreler gerekli | Değişkenlerin ve parametrelerin açıklaması |
| 3.3 | |||
| Bağlanma Tek sitesi ligand | Y = Alt + ((Alt-Üst) * (1 - ((PK d -X + sqrt (((P + X + K d) ^ 2) - (4 * P * X))) / (2 * P )))) | P: protein konsantrasyonu. Kd: Sabit ayrışma. P ve KD ligand konsantrasyonları için kullanılan aynı birimler cinsinden verilmiştir. Üst, Alt: sonsuz ligand konsantrasyonu ve sırasıyla hiçbir bağ konsantrasyonda erime sıcaklıkları. | |
| 3.4 | Alt = * Ymin | Ymin: (Bu durumda düşük deneysel protein Tm) Y minimum değeri | |
| En = * Ymax | Ymax: Y maksimum değeri (en yüksek deneysel protein Tm) | ||
| Kd = * X, en YMID | YMID: yMin ve Ymax ortalamasına tekabül Y değer. X (burada, ilgili ligand konsantrasyonu) dahilinde ilgili X değeri | ||
| P = (Başlangıç değer, uygun olması için) | |||
| 4.1 | |||
| Basit kooperatif modeli | Y = Alt + ((Alt-Üst) * (((X / Kd) ^ n) / (1 + ((X / Kd) ^ n)))) | K Hhasta katsayısı. Bu protein bir birliktelik ya da diğer biyokimyasal özellikleri açıklanmaktadır, ve kesin olarak protein ligand bağlama sitelerinin sayısına ilişkin bir tahmin değildir. Birine göre bir Hill katsayısı herhangi bir birliktelik temsil eder; Birden düşük değerler negatif bir birliktelik gösterir ve birden büyük pozitif bir birliktelik değerleri. | |
| Alt = * Ymin | |||
| En = * Ymax | |||
| Kd = * X, en YMID | |||
| P = (Başlangıç değer, uygun olmak ) | |||
| n = (Başlangıç değer, uygun olması için) | |||
| İki ligandların bağlanma Sıralı | Y = Alt + ((Alt-Üst) * ((X ^ 2) / (K d * K2)) / (1 + (X / K d) + ((X ^ 2) / (K d * K2))) ) | K2: ikinci bağlanma olayı için sürekli ayrışma. | |
| Alt = * Ymin | |||
| En = * Ymax | |||
| YMID K2 = * X, | |||
| P = (Başlangıç değer, uygun olması için) | |||
| Bağımsız iki ligand bağlanmasının | Y = Alt + ((Alt-Üst) * ((X ^ 2) / (K d * K2)) / (1 + (2 * X / K d) + ((X ^ 2) / (K d * K2) ))) | ||
| Alt = * Ymin | |||
| Kd = * X, en YMID | |||
| YMID K2 = * X, | |||
| P = (Başlangıç değer, uygun olması için) | |||
| 5.5 | |||
| Erime sıcaklığı ikili kaymalar analizi | Y = 1 - ((PK d = X + sqrt (((P + X + Kd) ^ 2) - (4 * p * X,))) / (2 x P) göre) | ||
| Alt = * Ymin | |||
| En = * Ymax | |||
| Kd = * X, en YMID | |||
| P = (Başlangıç değer, uygun olması için) | |||
| 5.8 | |||
| Sonsuz ligand konsantrasyonu Dışdeğerbiçim | (C2 - ((1 $ R $ 2) * B2)) / $ R $ 2 | B2: hiçbir ligand sonucunu içeren bir hücre. C2: Maksimum ligand ile sonucunu içeren hücre. $ R $ 2: maksimum ligand konsantrasyonunda bağlı oranını içeren hücre. |
4, 3 ve 5 numaralı adımları analiz yazılımı içine ayrıntılı denklemlerin eklenmesi ve veri analizi için başlangıç parametrelerin kesin tanımını gerektirir. Her bir ilgili adım için denklemler parametrelerin doğru bir seçim ile gösterilir. Değişkenlerin ve parametrelerin anlamı bir açıklama başvuru için sağlanmıştır.
Tablo glikoz ile heksokinazın etkileşim taranması için 4. Konsantrasyonları.
| Örneklem nokta | Ligand (da glikoze) konsantrasyonu (mM) |
| 1 | 0 |
| 2 | 0.001 |
| 3 | 0.005 |
| 4 | 0.01 |
| 5 | 0.03 |
| 6 | 0.1 |
| 7 | 0.3 |
| 8 | 0.4 |
| 9 | 0.7 |
| 10 | 1.1 |
| 11 | 2.1 |
| 12 | 3.7 |
| 13 | 5.3 |
| 14 | 7 |
| 15 | 9 |
| 16 | 11 |
Protokolde tarif edildiği gibi bilinen bir magnezyum kofaktör olarak çiçek mayası Saccharomyces cerevisiae ikinci heksokinaz, 10 mM MgCl2 ile takviye edilmiş, ana karışıma ilave edildi. Kd başlangıç tahmini 0.5 ile 2 mm arasında olmuştur. Deneyler glikoz belirtilen nihai konsantrasyonlar elde etmek için oluşturulmuştur.
Tablo GSYİH ile WcbM etkileşiminin taranması için 5. Konsantrasyonları.
| Örneklem nokta | Ligand (GTP) konsantrasyonu (uM) |
| 1 | 0 |
| 2 | 0.5 |
| 3 | 1 |
| 4 | 5 |
| 5 | 10 |
| 6 | 25 |
| 7 | 50 |
| 8 | 100 |
| 9 | 250 |
| 10 | |
| 11 | 1.000 |
| 12 | 2.500 |
| 13 | 5.000 |
| 14 | 7.500 |
| 15 | 10.000 |
| 16 | 20.000 |
Protokolde tarif edildiği gibi, Burkholderia pseudomallei ikinci WcbM ana karışıma ilave edildi. K d ilk tahmini yaklaşık 100 uM'ydi. Deneyler yukarıda ve aşağıda Kd büyüklük en az iki sıra kapatılmasına yönelik, GTP belirtilmedikçe nihai konsantrasyonları sağlamak için kurulmuştur.
Tartışmalar
Diferansiyel tarama fluorimetry proteinleri karakterize ve potansiyel protein ligandları teşhis etmek için güçlü ve çok yönlü bir yöntem olarak gücünü göstermiştir. (Özellikle daha az iyi finanse laboratuvarlarda) protein stabilizasyonu, ilaç keşif ve hızlandırılması kristalleşme 10,23-25 de iyi belgelenmiş başarıları bu bileşiklerin ilk taraması için cazip bir yöntem yaptık. Proteinlerine ilave bileşikler, belirgin bir erime sıcaklığı, 7,9 açıkça doza bağlı bir artış göstermektedir. Ancak, afinite için bileşikleri sıralaması yardımcı olmak için görünür bağlanma sabitleri belirlemek için bu deneylerin sonuçlarını kullanmak için birkaç girişimleri olmuştur. Burada, sistematik bir ligandın varlığında, proteinler için belirgin bir ayrışma sabiti belirlenmesi için bir yöntem sunulmaktadır.
Burada sunulan sonuçlar DSF hızlı ve güçlü bir şekilde için ayrışma sabitinin tahminleri sağlayabilir göstermektedirBir protein-lijand kombinasyonu. Gözlemlenen veriler parametrelerin olası değerine ilişkin varsayımlar yapmak için gerek kalmadan, K d hızlı bir belirlenmesini sağlamak üzere piyasada mevcut araçları ile manipüle edilebilir. Yöntem, istenilen protein hem de zaman içinde hasis olma, bazı benzer yöntemlere kıyasla önemli bir avantajı vardır. Burada tarif edilen deney, deney başına (üç kopya halinde tekrarlanan deneylerde yaklaşık 0.4 mg) bir protein 0.13 mg tüketecektir. Bu ortalama bir 40 kDa proteini ile tek bir deney benzer miktarda tüketmek izotermal titrasyon kalorimetri (ITC), olumlu ile karşılaştırır. Bu protokol için gerekli deneylerin tam set deney tek bir set için, hazırlık olmak üzere yaklaşık 4 saat etkilerdi. Yine, bu tür güçlü iken, genellikle en iyi verilerin elde etmek için önemli bir optimizasyon gerektirebilir ITC veya yüzey plazmon rezonansı gibi yöntemlere göre önemli ölçüde daha hızlı olması muhtemeldir.
Bizim sonuçlar dikkatli ham verileri incelemek için bir gereklilik olduğunu ortaya koymaktadır, bu verilerin uygun erime sıcaklığını belirlemek için, ve erime sıcaklığı verilerinin uygun ayrışma sabiti belirlemek için. İlk sorun, protein erimesi üretilen ham veri şeklidir. Bazı durumlarda, şekil Şekil 1A gözlenene yaklaşık olmayabilir. Ortak sorunlar ligand düşük sıcaklık değişimleri, yüksek arka floresan ve sıcaklık sıradışı çoklu geçişleri içerir. Düşük sıcaklık geçişi ligandlann bir dizi bağlayıcı görülür. Bu yöntem için, en önemli parametre sıcaklık geçişi ile karşılaştırıldığında, Tm ölçüm hatasıdır. , Üçlü Ölçümlerin standart sapması bağlanmamış ve tam bağlı protein arasındaki erime sıcaklık geçişinin% 10'unu yok olduğunda veri genellikle gayet iyi takılabilir. Deneyimlerimiz bu nerede böyle tempıcaklık kaymalar, tek tek veri noktalan son derece hassas ise sadece 2 ° C, bu ayar, verilerin yerleştirilmesi için yeterli olabilir bulunmaktadır. İkinci bir konu alışılmadık eğrileri şekillenir. Ligan bağlama proteininin açılımı modları etkiler gibi bu, genellikle, serbest protein ve ligand bağlı şekilde farklılık gösterir. Bu durumda, kullanıcı, veri erime sıcaklığı ve ayrışma sabiti belirlemek için kullanılacak olan modellerin uygun dikkate alınarak kullanılabilen olmadığını değerlendirmelidir. Başka bir genel sorun, proteine bir kofaktör eklenmesi (örneğin, heksokinaz örneğimizde MgCI2), en güvenilir veri elde etmek için gerekli olmasıdır. Bizim deneyim ilk okutmalarında iyi sonuçları elde etmek esastır aşamasında deneyde tüm olası faktörlerin dikkatli dikkate olmuştur. Ayrıca alternatif teorik tedaviler bu verilerin 15,17 özelliklerini ortaya çıkarabilir. Son olarak, bu C edilen bazı proteinler için nadir değildirontain doğal olarak yüksek eşiğe göstermek için hidrofobik bölgeler maruz bırakılmıştır. Başka yerde yoğun 6,9 gözden geçirilmiş, bu problemlere çözümler vardır.
Özel olarak, kullanıcı (örneğin, Şekil 4) ve türevlerinin kullanılması durumunda, birden fazla erir modellenmelidir olup Boltzmann veya türevi modelleri kullanmak için olmadığını değerlendirmelidir. Açılımı termal modelleme iki yöntem Boltzmann yöntemi tutmasına eğrisine düzenli bir sigmoid şekli varsayarak, Boltzmann denklemi deneysel verilere uygun farklı. Bunun aksine, yöntem, her türev noktası (Şekil 1A 'da alt panel) deneysel veriler birinci türevi alır ve en birinci türevinin noktası olarak erime sıcaklığı dikkate alır. 3 ° C - türevi Yöntem genel olarak, yaklaşık 2 ile daha yüksek bir erime sıcaklığına döndürür. Çoğu protein, daha tutarlı döndürüriki yöntemden biri için sonucu (yani, üçlü deney için erime sıcaklığının standart hatası daha düşüktür). Bu genellikle iyice eğrisi açılma proteinin tam bir şekli ile ilgili olup, bu deneysel her durumda en iyi yöntemi belirlemek için gereklidir. Türevi modeli kullanıldığında, birden fazla erime olayları dikkate almak da önemlidir. Bazı veriler açıkça çoklu geçişler için kanıt göstermek ve bu durumlarda sonuçlar, bu birden fazla erime olayları modellenmiştir olmadığını yorumlamak daha kolay olması muhtemeldir. Bu protokolün bağlamında, ligand eklenmesi bir protein tersi (en termal kırılgan subdomain stabilize ederek, örneğin) bir tek geçiş birden fazla erime geçişleri sahip kaydırmaya veya neden olabilir genellikle böyledir. Bu nedenle ham veri kullanmak için iyi olacak hangi yaklaşım dikkate alınarak önce birlikte incelendiğinde olduğunu savunmaktadır.
M takibenBireysel erime sıcaklıklarının odelling, başka sorunları protokol bölümünde sunulan modeller bu uydurma ortaya çıkabilir. Bu analiz, çoğunlukla gözlenen veri ve modelin arasındaki tutarsızlıkları vurgular gibi dikkatle, bir logaritmik ölçek kullanarak ayrışma sabiti denkleme uyum incelemek için zorunludur (e .g., Şekil 3). Elde edilen sonuçlar genel olarak sağlam iken, yorumlanması bakım verilerden daha iyi sonuçlar elde etmek için fırsat ve en anlama sahiptir.
Bu verilerin gündeme belirli bir konu DSF de, birliktelik gösterebilir proteinler, ya da birden fazla bağlayıcı olaylara konulmalıdır yorumudur. Bu, şimdiye kadar sadece çok özel bağlama etkinlikleri (örneğin, WcbM, boyut dışlama kromatografisi ile, bir multimer olarak işlev gören en homologu 26 bir multimer olan protein, [bir veri sahip olmaları beklenir proteinler bu olguyu gözlemledikgösterilmiştir]). Bu günün tüm açık DSF denatürasyondan gözlenen olumsuz kooperativiteye enzim sonuçta olumsuz bir birliktelik göstereceğini belirtir ki değil: ziyade, bu yöntemleri daha geniş bir yelpazede kullanarak daha fazla iyice incelenmesi gerekir karmaşık bağlayıcı bir göstergesi olabilir. Bu tür proteinlerin daha kapsamlı çalışmaların ilginç etkilerini tespit etmek olasıdır ki, bize düşündürmektedir.
Bu yöntemi kullanarak ayrışma sabiti için verilen değerler, genel olarak, izotermal titrasyon kalorimetrisi ve yüzey plazmon rezonansı gibi yöntemler tarafından sağlananlar gibi aynı mertebededir. Ancak, gözlenen mutlak değerleri, bu yöntemler kullanılarak gözlemlenen daha sıklıkla yüksektir. Bu, en azından kısmen ayrışma sabiti ligandı ile proteinin erime sıcaklığında görülmektedir gerçeğinin bir sonucudur. Bu Kd fizyolojik sıcaklıklarda daha yüksektir. Dissociation, sabit denklemler ile reaksiyonun sıcaklığı ile ilgili olup:
 [1]
[1]
 [2]
[2]
(C θ standart referans konsantrasyonunu, Δ G reaksiyonun Gibbs serbest enerjisi değişim r, R, molar gaz sabitidir, Δ H reaksiyonda entalpi değişimi ve Δ S reaksiyonunda değişim entropi .)
Bu yöntemin ölçülebilir aralığında ayrışma sabitleri ile reaksiyonları, genellikle olumsuz bir Δ Rg sahip olacak ve bu nedenle denkleme sıcaklığındaki bir artışın etkisi [1] ayrışma sabiti arttırılması olacaktır. Her iki Gibbs serbest enerjisini oluşturan Δ H ve Δ S terimler (denklemi [2]) sıcaklık vardıre göre 27 ve ayrışma sabitinin üzerindeki etki, bu sıcaklık bağımlılıkları büyüklüğü ve işareti bağlıdır ve zorunlu olarak etkileşim bağlı olacaktır. Sonuç olarak, bu yöntemle belirlenen ayrılma sabitleri oda sıcaklığında çalışmasına yöntemlerle belirlenen zaman daha yüksek olduğu beklenmedik bir durum değildir. Sıcaklık bağımlılık da, tabii ki, fizyolojik sıcaklıkta daha düşük sıcaklıklarda ayrışma sabiti temin etme eğilimindedir birçok yöntem, bir sorun teşkil etmektedir.
Bu ITC farklı olarak, etiketlenmiş bir yöntem olduğu DSF yöntemin diğer bir sorun teşkil etmektedir. (SYPRO Turuncu) kullanılan flüoresan etiket hidrofobiktir ve bu nedenle bazı durumlarda proteinlerin hidrofobik ligandlan ile rekabet edebilir. Sonuç olarak, bazı durumlarda, elde edilen ayrışma sabiti yapay bağlı etiket ile rekabete yükseltilecek muhtemeldir. Bununla birlikte, çeşitli ligandların karşılaştırma (birincil kullanım içinDSF), afinite ile fark bileşiklerin sıralamasını etkiler için yeterince büyük olması olası değildir.
Bu yöntemin olası bir dezavantajı, elde edilebilir tespit sınırıdır. Prensip olarak, doğru protein konsantrasyonunun% 50'den daha düşük olduğu, ve bu aralıkta daha değerleri şüpheli doğruluk olması muhtemeldir için Kd değerini ölçmek mümkün olmamalıdır. Aralığın bu sonunda algılama sınırı protein ve boya konsantrasyonlarının azaltılması ile biraz uzatılabilir iken, cihazın, söz konusu protein konsantrasyonundaki azalma daha da engeller. Benzer şekilde, duyarlılık üst ucu ligandın çözünürlükleri tarafından belirlenir. Kd için matematiksel olarak sağlam bir tahmininin elde edilmesi için, yaklaşık olarak bir ligand konsantrasyonu gerektirmektedir ligand-bağlı biçimde de mevcut olan protein,% 90 veri elde edilmesi için çok önemlidirly on kat K d (hayır birliktelik varsayarak). Saptama sınırı bu sebeple, ilgili tampon içinde ligandın çözünürlüğüne onda biri olacaktır. Bu yöntem algılama sınırları tipik olarak protein ve ligand bağlı mM 1 uM ila yaklaşık 1 ila 100 arasında olacağı anlamına gelir.
Sonuç olarak, diferansiyel tarama fluorimetry proteinlerin geniş bir aralığına uygulanabilirdir çok yönlü bir tekniktir. Burada yer alan yöntem kullanılarak, hızla ve ucuz bir şekilde farklı ligandlar için bir proteinin afinitesini belirlemek mümkündür. Bu durum özellikle küçük laboratuvarlarda, protein saflaştırma ve istikrar uygulama, metagenomes enzimlerin işlevini veya spesifisitesini ortaya çıkması için büyük bir potansiyele sahiptir ve ilaç keşfi.
Açıklamalar
The authors declare that they have nothing to disclose.
Teşekkürler
This work was funded by grant from the BBSRC (grant number BB/H019685/1 and BB/E527663/1) to the University of Exeter.
Malzemeler
| Name | Company | Catalog Number | Comments |
| StepOne real time PCR instrument | Life Technologies | 4376357 | DSF can be performed with many other instruments. The StepOne instrument has very convenient software for data analysis. |
| Protein thermal shift software v1.0 | Life Technologies | 4466037 | |
| MicroAmp Fast optical 48-well plates | Life Technologies | 4375816 | |
| Optical sealing tape | Life Technologies | 4375323 | Bio-rad part no. 223-9444 is an alternative supplier |
| U-bottomed 96-well plates | Fisher | 11521943 | |
| SYPRO Orange | Life Technologies | S6650 | For a smaller volume supplier, use Sigma part no. S5692 |
| SPSS statistics version 20 | IBM | Other statistics packages will provide similar functionality | |
| GraphPad Prism 6.02 | GraphPad | Other statistics packages will provide similar functionality | |
| Hand applicator (PA1) | 3M | 75-3454-4264-6 | |
| Hexokinase from Saccharomyces cerevisiae | Sigma-Aldrich | H5000 | |
| Glucose | Fisher scientific | 10141520 |
Referanslar
- Freyer, M. W., Lewis, E. A. Isothermal titration calorimetry: experimental design, data analysis, and probing macromolecule/ligand binding and kinetic interactions. Methods Cell Biol. 84, 79-113 (2008).
- Ladbury, J. E. Calorimetry as a tool for understanding biomolecular interactions and an aid to drug design. Biochem Soc Trans. 38, 888-893 (2010).
- Abdiche, Y., Malashock, D., Pinkerton, A., Pons, J. Determining kinetics and affinities of protein interactions using a parallel real-time label-free biosensor, the Octet. Anal Biochem. 377, 209-217 (2008).
- Pantoliano, M. W., et al. High-density miniaturized thermal shift assays as a general strategy for drug discovery. J Biomol Screen. 6, 429-440 (2001).
- Senisterra, G., Chau, I., Vedadi, M. Thermal denaturation assays in chemical biology. Assay Drug Dev Technol. 10, 128-136 (2012).
- Ericsson, U. B., Hallberg, B. M., Detitta, G. T., Dekker, N., Nordlund, P. Thermofluor-based high-throughput stability optimization of proteins for structural studies. Anal Biochem. 357, 289-298 (2006).
- Lo, M. C., et al. Evaluation of fluorescence-based thermal shift assays for hit identification in drug discovery. Anal Biochem. 332, 153-159 (2004).
- Nettleship, J. E., Brown, J., Groves, M. R., Geerlof, A. Methods for protein characterization by mass spectrometry, thermal shift (ThermoFluor) assay, and multiangle or static light scattering. Methods Mol Biol. 426, 299-318 (2008).
- Niesen, F. H., Berglund, H., Vedadi, M. The use of differential scanning fluorimetry to detect ligand interactions that promote protein stability. Nat Protoc. 2, 2212-2221 (2007).
- Geders, T. W., Gustafson, K., Finzel, B. C. Use of differential scanning fluorimetry to optimize the purification and crystallization of PLP-dependent enzymes. Acta Crystallogr Sect F Struct Biol Cryst Commun. 68, 596-600 (2012).
- Vedadi, M., et al. Chemical screening methods to identify ligands that promote protein stability, protein crystallization, and structure determination. Proc Natl Acad Sci USA. 103, 15835-15840 (2006).
- Davis, B. J., Erlanson, D. A. Learning from our mistakes: the 'unknown knowns' in fragment screening. Bioorg Med Chem Lett. 23, 2844-2852 (2013).
- Larsson, A., Jansson, A., Aberg, A., Nordlund, P. Efficiency of hit generation and structural characterization in fragment-based ligand discovery. Curr Opin Chem Biol. 15, 482-488 (2011).
- Scott, D. E., et al. Using a fragment-based approach to target protein-protein interactions. Chembiochem. 14, 332-342 (2013).
- Cimmperman, P., et al. A quantitative model of thermal stabilization and destabilization of proteins by ligands. Biophys. J. 95, 3222-3231 (2008).
- Matulis, D., Kranz, J. K., Salemme, F. R., Todd, M. J. Thermodynamic stability of carbonic anhydrase: measurements of binding affinity and stoichiometry using ThermoFluor. Biochemistry. 44, 5258-5266 (2005).
- Zubriene, A., et al. Measurement of nanomolar dissociation constants by titration calorimetry and thermal shift assay - radicicol binding to Hsp90 and ethoxzolamide binding to CAII. Int J Mol Sci. 10, 2662-2680 (2009).
- Weiss, J. N. The Hill equation revisited: uses and misuses. FASEB J. 11, 835-841 (1997).
- Cuccui, J., et al. Characterization of the Burkholderia pseudomallei K96243 capsular polysaccharide I coding region. Infect Immun. 80, 1209-1221 (2012).
- DeShazer, D., Waag, D. M., Fritz, D. L., Woods, D. E. Identification of a Burkholderia mallei polysaccharide gene cluster by subtractive hybridization and demonstration that the encoded capsule is an essential virulence determinant. Microb Pathog. 30, 253-269 (2001).
- Berman, H., Henrick, K., Nakamura, H. Announcing the worldwide Protein Data Bank. Nat Struct Biol. 10, 980(2003).
- Vivoli, M., Ayres, E., Beaumont, E., Isupov, M., Harmer, N. Structural insights into WcbI, a novel polysaccharide biosynthesis enzyme. IUCr Journal. 1 (1), 28-38 (2014).
- Sorrell, F. J., Greenwood, G. K., Birchall, K., Chen, B. Development of a differential scanning fluorimetry based high throughput screening assay for the discovery of affinity binders against an anthrax protein. J Pharm Biomed Anal. 52, 802-808 (2010).
- Uniewicz, K. A., et al. Differential scanning fluorimetry measurement of protein stability changes upon binding to glycosaminoglycans: a screening test for binding specificity. Anal Chem. 82, 3796-3802 (2010).
- Wan, K. F., et al. Differential scanning fluorimetry as secondary screening platform for small molecule inhibitors of Bcl-XL. Cell Cycle. 8, 3943-3952 (2009).
- Koropatkin, N. M., Holden, H. M. Molecular structure of alpha-D-glucose-1-phosphate cytidylyltransferase from Salmonella typhi. J Biol Chem. 279, 44023-44029 (2004).
- Paleskava, A., Konevega, A. L., Rodnina, M. V. Thermodynamics of the GTP-GDP-operated conformational switch of selenocysteine-specific translation factor SelB. J Biol Chem. 287, 27906-27912 (2012).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır