このコンテンツを視聴するには、JoVE 購読が必要です。 サインイン又は無料トライアルを申し込む。
Method Article
チャンバーおよびプレートベースの呼吸計を使用して細胞および組織の生体エネルギーを評価するための高解像度呼吸法
要約
高分解能呼吸計を用いた酸化的リン酸化の評価は、ミトコンドリアおよび細胞エネルギー代謝の機能解析の不可欠な部分となっている。ここでは、チャンバーおよびマイクロプレートベースの高分解能呼吸計を使用して細胞エネルギー代謝を分析するためのプロトコルを提示し、各デバイスの主な利点について説明します。
要約
高分解能呼吸法(HRR)により、酸化的リン酸化をリアルタイムでモニタリングし、個々の細胞エネルギー状態の分析や、多様な基質非結合剤阻害剤滴定(SUIT)プロトコルを使用した呼吸複合体の評価を行うことができます。ここでは、2つの高解像度呼吸測定装置の使用例を示し、培養細胞、骨格筋線維および心筋線維、ならびに脳および肝臓などの軟部組織の分析に適用可能なプロトコルの基本的なコレクションを提示する。培養細胞および組織のためのプロトコルは、チャンバーベースの呼吸計およびマイクロプレートベースの呼吸数計のための培養細胞のために提供され、両方とも標準的な呼吸プロトコルを包含する。比較の目的で、多発性呼吸器系欠損を引き起こすミトコンドリア翻訳を欠損させたCRISPRが操作したHEK293細胞を両方の装置と共に使用して、呼吸における細胞欠陥を実証する。両方の呼吸計は、研究中の研究課題とモデルに依存するそれぞれの技術的メリットと適合性を備えた細胞呼吸の包括的な測定を可能にします。
概要
ミトコンドリアは、エネルギーの重要な供給を満たし、ヌクレオチド、脂質およびアミノ酸の同化作用、鉄硫黄クラスター生合成などの必須の細胞生体エネルギーおよび代謝プロセスに寄与する区画化された細胞小器官であり、制御された細胞死などのシグナル伝達に関与している1,2,3.酸化的リン酸化によるミトコンドリア生体エネルギー学は、細胞内のほぼすべての細胞プロセスに寄与し、その結果、一次または二次起源のミトコンドリア機能不全は、広範囲の疾患状態と関連している4,5。ミトコンドリア機能障害は、構造やミトコンドリア密度の変化だけでなく、呼吸器系の質や調節も伴います6。この定性的要素は、基質制御、結合特性、翻訳後修飾、クリステダイナミクス、および呼吸スーパーコンプレックス7、8を包含する。したがって、細胞のエネルギー代謝を評価するための実験的および診断的アプローチのためのミトコンドリア生体エネルギー学の正確な分析は、健康および疾患において重要である。
ミトコンドリア酸化リン酸化(OXPHOS)は、アデノシン三リン酸(ATP)を介して細胞エネルギーを生成するための呼吸器系または電子伝達系(ETS)内の一連の反応です9。複合体IおよびIIを通る電子流から複合体IVへのエネルギーを利用する多酵素ステップは、内側ミトコンドリア膜を横切って電気化学的プロトン勾配を生成し、続いて複合体V(F1FOATP合成酵素)を介してアデノシン二リン酸(ADP)からATPへのリン酸化に利用される(図1A)。
まず、トリカルボン酸環(TCA)、解糖系、ピルビン酸酸化の間に2つの電子伝達体が生成される:ニコチンアミドアデニンジヌクレオチド(NADH)およびジヒドロフラビンアデニンジヌクレオチド(FADH2)。NADHは錯体I(NADHデヒドロゲナーゼ)で酸化され、その間に2つの電子がコエンザイムQに伝達され(キノンはキノールに還元され)、プロトンは膜間空間(IMS)にポンプで送られます。第二に、複合体II(コハク酸デヒドロゲナーゼ)はFADH2を酸化し、プロトンをポンピングすることなく電子をコエンザイムQに供給する。第三に、複合体III(シトクロムc酸化還元酵素)では、コエンザイムQからの電子がシトクロムcに移され、プロトンがIMSにポンピングされる。第四に、シトクロムcは電子を複合体IV(シトクロムcオキシダーゼ)に伝達し、最終的な複合体は陽子をポンプし、酸素が電子受容体として機能してプロトンを同化し、最終的に水を形成する。ミトコンドリアが消費するのはこの酸素であり、オキシグラフで測定することができます。最後に、複合体I、複合体III、および複合体IVから生成されたプロトンを使用して複合体Vを回転させ、それによってATP9を生成する。
重要なことに、電子移動は直線的な様式で起こるだけでなく、電子輸送鎖とも表記される。代わりに、電子は複数の呼吸経路を介してコエンザイムQプールに伝達され、収束電子の流れを促進することができる。NADH基質およびコハク酸塩は、例えば、複合体Iおよび複合体IIを介してそれぞれ侵入することができる。脂肪酸酸化からの電子は、電子伝達フラビンタンパク質複合体を介して供与することができる。実際、OXPHOSの包括的な分析には、適切な燃料基材を用いた総合的なアプローチが必要です(図1A)。

図1:ミトコンドリア酸化リン酸化ならびに特異的基質および阻害剤プロトコル。 (A)ミトコンドリアおよび電子伝達系(CI−CIV)およびミトコンドリアF1F0ATP合成酵素(CV)のスキーム。すべての構造体はPDBからのものです。図は、この研究で記載された基質および阻害剤のみを描写している)。(B)mHRR装置における標準プロトコールを用いた無傷のHEK293細胞における酸素フラックスのサンプル微量。(C)cHRR装置における標準プロトコールを用いた無傷のHEK293細胞における酸素フラックスのサンプル微量。(d)それぞれのSUITプロトコールを用いて健康なドナーからの透過処理されたヒト線維芽細胞における微量の酸素流束のサンプル。略語:1 =無傷の細胞の日常的な呼吸;2 = 状態 2;3 = 状態 3(I);4 = cytCを持つ状態3(I)。5 = 状態 3 (I+II);6 = リーク(OM);7 = ETS 容量;8 = S(ROT);9 = ROX;10 = TMPD;ROT=ロテノン、AM=アンチマイシン、ATP=アデノシン三リン酸、Az=アジド、OM=オリゴマイシン、FCCP=カルボニルシアン化物p-トリフルオロメトキシフェニル-ヒドラゾン;Asc = アスコルビン酸塩、TMPD = N,N,N',N'-テトラメチル-p-フェニレンジアミン、スク酸 = コハク酸塩、M = リンゴ酸塩、P = ピルビン酸塩、ADP = アデノシン二リン酸、NAD = ニコチンアミドアデニンジヌクレオチド、IMS = 膜間空間、FAD = フラビンアデニンジヌクレオチド。この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
HRRを用いたミトコンドリアOXPHOS能力の分析は、原発性ミトコンドリア欠損10,11だけでなく、癌や老化12などの生物学の他のすべての領域にも及ぶ診断値の器械的生化学的方法となっている12。HRRは、ミトコンドリアOXPHOS能力の分析による細胞呼吸の決定を可能にし、これは、個体または結合されたミトコンドリア呼吸器複合体欠損を直接反映し、間接的に細胞機能障害および変化したエネルギー代謝に関連する9。方法論的には、呼吸測定は、細胞、組織、または単離されたミトコンドリア11、13、14、15、16の凍結材料のみを部分的に適切なものを用いて行う。凍結組織は、維持された超複合体安定性を有する無傷のETSを有することが示される15。したがって、従来のTCA中間体とは対照的に、それぞれの基質はETSに直接供給される。しかし、ETSとATP合成との間の結合は、凍結損傷(氷結晶形成)によって膜の完全性が損なわれるにつれて失われる。
呼吸実験は、通常、非透過処理または透過処理された細胞または組織のいずれかにおける吸熱について、37°Cの生理学的温度で行われる。前者は細胞質ゾル代謝の文脈を考慮するが、後者は特定の基質(および阻害剤)の添加を通じて個々のOXPHOS複合体およびATPaseのエネルギー的寄与を提供する。基質および阻害剤の配列および変異は、OXPHOS機能に関する様々な科学的問題に対処するための多様なSUITプロトコル17 およびアッセイ18 の開発につながった(12でレビュー)。細胞呼吸の基本プロトコールは、4つの異なる状態を評価する:i) ルーチン呼吸−基質または阻害剤を消費するが内因性基質の添加なしにそれぞれの呼吸媒体中での呼吸。この状態は、一般的なOXPHOSまたは二次誘発呼吸欠陥、例えば代謝産物プロファイルの変化によって引き起こされるものを明らかにすることができる。次に、ATPase阻害剤オリゴマイシンの添加は、プロトンに対するミトコンドリア内膜の透過性を明らかにし、ii) 漏出呼吸として定義する。その後のアンカプラカルボニルシアン化物p-トリフルオロメトキシフェニル-ヒドラゾン(FCCP)などのプロトノフォアの滴定は、iii) 非結合呼吸 として定義される膜開放型プロトン回路モードにおいてETS容量が最大となる状態を決定することを可能にする。重要なことに、非結合状態は、ミトコンドリア膜への過度の機械的損傷を介した実験的介入によっても起こり得る。逆に、非結合状態とは、生理学的に制御された内因的な機構による呼吸脱結合をいう。最後に、複合体III阻害剤アンチマイシンおよび複合体I阻害剤ロテノンの添加によるETSの完全な阻害は、非ミトコンドリア酸素消費プロセスからの 残留酸素消費 (ROX)を決定する(図1A−C)。
ミトコンドリア生体エネルギー学は、5つの異なる呼吸状態19,20からなる。状態1呼吸は、内因的に利用可能なものを除いて、追加の基質またはADPを含まない。ADPの添加後、依然として、基質がないが、状態2呼吸が達成される。基質が添加されると、電子移動およびATP合成が可能になり、状態3呼吸に達する。この状態では、OXPHOS容量は、ADP、無機リン酸、酸素、NADHおよびコハク酸結合基質の飽和濃度で定義することができる。状態4呼吸またはLEAK呼吸は、十分な基質を有しながらADPまたは化学的に阻害されたATP合成酵素を含まない状態として定義することができる。最後に、閉室設定ですべての酸素が枯渇(無酸素)されると、状態5の呼吸が観察される。
Oroboros 2k高分解能呼吸計とタツノオトシゴXF細胞外フラックスアナライザーという、実験モデルと研究課題に応じて適用性が異なる密閉チャンバーシステムにおける酸素の経時的な減少の関数として測定された酸素消費量の分析を通じて、OXPHOSの現在のリアルタイム評価を支配する2つの装置を用いて、細胞エネルギー状態14 を評価するためにいくつかの方法が存在する。両方の装置は、チャンバまたはマイクロプレートウェル内の絶対値として、1秒あたりの酸素(O2)のピコモール(pmol)の減少として酸素消費速度を記録する。質量当たりの比酸素消費量は、細胞数(数百万)、組織重量(mg)、またはタンパク質量当たりの特定の緩衝レシピにおけるそれぞれの酸素消費量を正規化することによって得られる。
O2k(オロボロスインスツルメンツ)は、ポラログラフ酸素センサ(チャンバベースの高解像度呼吸計:cHRRと略される)を備えた閉じた2チャンバシステムです。各実験チャンバは、マグネチックスターラーによって均質に保たれる2mLの液体を保持する。ポラログラフ酸素センサは、アンペロメトリックアプローチを使用して酸素を測定します:金カソード、銀/塩化銀陽極、およびKCI溶液の間に電圧(0.8V)が印加される電気化学セルを作成します。アッセイ培地からの酸素は、25μmのフッ素化エチレンプロピレン膜(O2透過性)を通って拡散し、陰極で還元を受け、過酸化水素を生成する。陽極では、銀が過酸化水素によって酸化され、電流を発生させる。この電流(アンペア)は酸素分圧に直線的に関係している。酸素分圧およびアッセイ媒体の酸素溶解係数は、酸素濃度を計算するために使用される。酸素分圧は実験温度に依存し、ポラログラフ測定は温度に敏感であるため、温度の変動はペルチェ加熱ブロックによる正確な(±0.002°C)調節を必要とする。温度は4°Cおよび47°Cの範囲で制御することができる。
タツノオトシゴXF細胞外フラックス分析装置(Agilent)は、3つの蛍光電極が各ウェルの酸素消費量を経時的に測定する24ウェルまたは96ウェルマイクロプレートフォーマットのプレートベースのシステムです(マイクロプレートベースの高解像度呼吸計:mHRRと略されます)。アッセイカートリッジ内の最大 4 つのポートは、アッセイ中の自動注入に使用できます。アッセイには複数のサイクルが含まれ、それぞれに3つのフェーズ(1)混合、2)待機、および3)測定があります。測定段階では、センサープローブをマイクロプレートに下げて、7〜10μLの容量を含む一時的に閉じたチャンバを作成し、放出された光を測定します。この光は、センサープローブの先端にあるポリマー包埋蛍光色素によって放出され、リン光消光に基づいてO2 を感知します。蛍光シグナルの強度はO2 に比例し、センサおよびアッセイ媒体の温度によって影響される。したがって、正確な酸素推定には、サンプルなしでバックグラウンドウェルを備えた相対的なアプローチが必要です。酸素濃度の回復は、センサーが上下に移動して仮チャンバの上の体積を混合する混合フェーズ中に発生します。各サイクルで 1 つの酸素消費率が計算されます。温度は16°Cおよび42°Cの範囲で制御することができる。
HRRは、原発性およびミトコンドリア関連疾患および一般的な細胞代謝における細胞生体エネルギーを評価するためのゴールドスタンダードです。この研究では、細胞および組織におけるOXPHOS機能を評価するために、HRRの基本的なプロトコールが提供される。

図2:cHRRのための細胞および組織調製のためのワークフロー、ならびにmHRR呼吸法のための細胞調製のためのワークフロー。(B)哺乳動物細胞(ステップ1.2):3 x106細胞に等しいHEK293ペレット(左パネル)。非線維組織(ステップ1.3):2mLテフロン陶器(中央パネル)におけるマウス小脳溶解物の調製。cHRR呼吸法のためのサポニン誘発骨格筋透過(ステップ1.4)右パネル)。(c)mHRR呼吸法のための真核細胞(HEK293)の分析のための標準的なマイクロプレート播種レイアウト(ステップ2.4)およびコンフルエンシーチェック。(D、E)mHRR呼吸法のための注入口装填のスキーム(ステップ2.4)。この図の拡大版を表示するには、ここをクリックしてください。
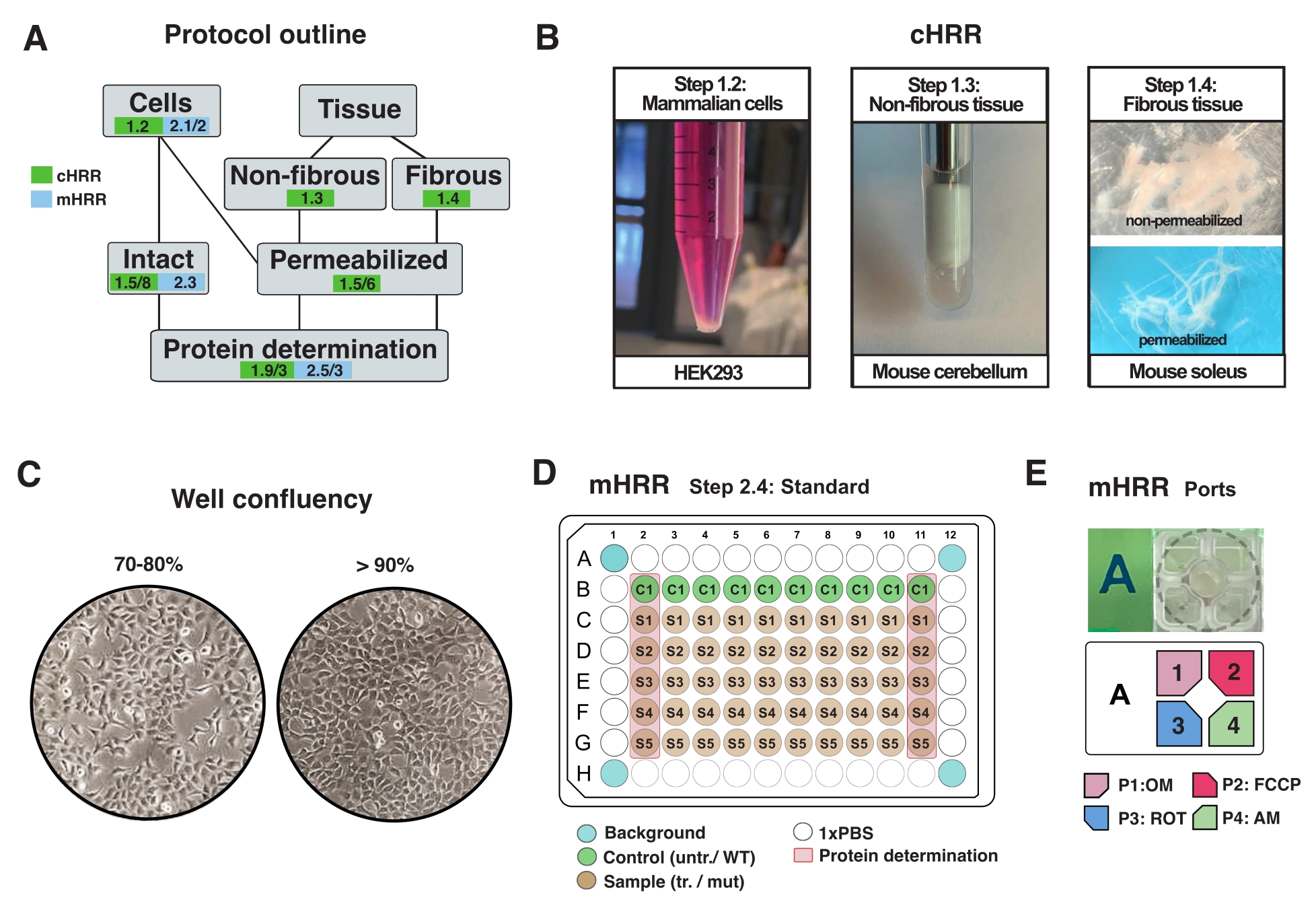{kind=link}
プロトコル
すべての動物実験は、国家動物実験審査委員会および南フィンランド地方国家行政庁に従って行われます。雄のC57BL/6JOlaHsdマウス(生後4~6ヶ月)を本試験に使用した。ヒト細胞株の使用に関する同意は、ヘルシンキ大学の制度倫理委員会から得られた。
1. 高解像度呼吸法:チャンバーベースの呼吸数計(cHRR)
注:プロトコルのこのセクションの実験は、Oroboros O2k-Core:Oxygraph-2k(材料表)を使用して実施されました。
- 酸素センサーの校正
- 2.1 mLのミトコンドリア呼吸培地(MiR05、 表1、溶解度係数:0.92)中で37°Cで37°Cでプレラン呼吸計を>45分間、21に記載したように酸素較正を行った。ベースライン変動が 4 pmol/s 以内±場合は続行します。
注:バックグラウンド信号の大きな変動は、センサー膜の必要なメンテナンス、または以前の実験からチャンバー内に残っているインヒビターの痕跡を示している可能性があります。実験25のバッチの前に、器械的バックグラウンド酸素流束補正が推奨される。 - 酸素校正値を記録して、センサー膜の性能を経時的に監視します。
メモ:これにより、センサー機能、信号対ノイズの安定性、センサーメンブレンのメンテナンスが必要な場合が明らかになります。周囲圧力に応じて、180 ~ 200 μmol の酸素が MiR05 に可溶化されます。 - 呼吸媒体中の任意の試料を添加する前にチャンバー内のすべての液体を除去する。
注:呼吸室の容積を定期的に正確に2mLに評価してください。
- 2.1 mLのミトコンドリア呼吸培地(MiR05、 表1、溶解度係数:0.92)中で37°Cで37°Cでプレラン呼吸計を>45分間、21に記載したように酸素較正を行った。ベースライン変動が 4 pmol/s 以内±場合は続行します。
- 高分解能呼吸測定のための細胞の調製
- 10%熱不活化ウシ胎児血清(FBS)、GlutaMax、非必須アミノ酸、ならびに5%CO2で37°Cのインキュベーター内でOXPHOS欠損代謝を支持するために、高グルコースを添加したダルベッコの改変イーグル培地(DMEM)中の直径10cm2の皿でHEK293細胞を培養した。
注:任意のタイプの真核細胞を培養することができる。ほとんどの細胞タイプでは、10 cm2 ディッシュを培養すると、十分な細胞 (通常 >3 x106 細胞) が得られます。マイコプラズマ感染を定期的にチェックし、細胞代謝や呼吸への影響を回避します。 - 90%コンフルエントを超えずに細胞を増殖させる(図2C)。
注:>90%のコンフルエンシーを有する細胞は、呼吸に対して増殖依存性の阻害効果を示すことがある(同期していないか、または有糸分裂後である場合)。 - 細胞を1x PBSで洗浄し、1mLの温かい0.25%トリプシンで剥離し、温かいDMEM(5mL/10cm2プレート)を加えてトリプシンを失活させ、血球計数器で細胞をカウントする。
- 2.5 x 106 個の細胞溶液を 300 x g で 5 分間穏やかに遠心分離し、上清を完全に除去し、2.5 mL の温かい MiR05 (1 x106 cells/mL) に再懸濁します (図 2A)。
- 浮遊細胞の場合、2.5 x106 細胞に等しい溶液を数えて除去し、ペレット化し、ステップ1.2.4で述べたように続行する。
- 透過処理の最適化(ステップ1.6)、透過処理された細胞または組織(ステップ1.5)、または無傷の細胞(ステップ1.7)のためのSUITプロトコルを実行します。
注: 一貫した結果を得るには、細胞濃度を一定に保つことをお勧めします (例: 1 x106 cells/mL)。呼吸数は呼吸計24における細胞密度とは無関係であるが、細胞数が一定に保たれていれば、基質および阻害剤は実験を通して同等の濃度にある。
- 10%熱不活化ウシ胎児血清(FBS)、GlutaMax、非必須アミノ酸、ならびに5%CO2で37°Cのインキュベーター内でOXPHOS欠損代謝を支持するために、高グルコースを添加したダルベッコの改変イーグル培地(DMEM)中の直径10cm2の皿でHEK293細胞を培養した。
- 高分解能呼吸測定のための非繊維組織(例えば、脳、肝臓)の調製
- 均質な組織片、重量30〜40mgを切除するか、臓器全体(この場合はマウス小脳)を使用する。
注: 組織をすぐに使用しない場合は、2 mL の氷冷 MiR05 に入れて保管し、ほとんどの組織で最大 2 時間保存できるようにします。個々の組織の保存時間は、時系列で評価する必要があります。 - Whatmanろ紙で乾いた組織を拭き取ります(注意:柔らかい組織物質が付着する傾向があります)。
- 30〜40mgの組織片を氷冷した2mLのポリテトラフルオロエチレンポッターElvehjemホモジナイザーに入れる。
- 適量のMiR05を加えて20mg/mLを得、組織対バッファー比を維持します。適切な機械的透過処理のために不十分または過剰な流体を避けるために、総量>1.5mLおよび<2mLを保管してください。
- 乳棒を挿入し、過度の組織損傷を引き起こす真空の発生を避けながら、乳棒を慎重に引っ込めることによって組織をゆっくりと溶解する。
- 溶解するまで(大きな破片のない濁った液体として見える)合計7回のストローク(上下のストロークとして1xと定義)を実行します(図2B)。
注:適切な溶解のための脳卒中の数は、シトクロムC応答を介してミトコンドリア外膜の完全性を評価することによって、各組織について試験する必要がある(ステップ1.5.11)。溶解しにくい結合組織または血管部分が残ることがあります。 - 溶解した組織を15mL遠沈管にデカントする。
- 溶解工程で使用した等量の MiR05 (例: 1.5 mL) で陶芸家の内部を洗浄し、3 ~ 4 mL の MiR05 が入った 15 mL チューブに 10 mg/mL の組織溶解液で加えます。
- チャンバーあたり2mLのプレーンMiR05を加え、37°Cに加温する。
- チャンバーあたり各溶解液の500 μL(5 mgに等しい)をゆっくりとピペッティングする前に、チューブを等分布に旋回させて、寒さから37°Cまでのストレスを最小限に抑えます。
- チャンバーの内容物>37°Cに温まるまで3分間待ってからチャンバーを閉じます。ストッパーの上部にある余分な液体を除去する(閉鎖後のチャンバーあたりの量:4mg)。
- 標準透過処理用の SUIT プロトコルを実行します (ステップ 1.5)。
- 均質な組織片、重量30〜40mgを切除するか、臓器全体(この場合はマウス小脳)を使用する。
- 高分解能呼吸測定のための繊維組織(骨格筋、心筋)の調製
- 硬組織を抽出し、解剖顕微鏡下で2mLの氷冷BIOPS(表2)中の鋭い鉗子を用いて筋肉から結合組織および脂肪を除去する。
- 繊維束(〜4mg)を長手方向軸に沿って鋭い鉗子で分離する。繊維を十分に引き抜き、網目状の構造を得る(図2B)。
注:適切な機械的繊維分離および透過処理は、赤色色素ミオグロビンの喪失および透光性の増加によって示される。 - 繊維束を洗浄し、サポニン(BIOPSで50μg/mL、新鮮に調製)で4°Cで20分間透過処理する(繊維は半透明になり、完全な透過処理を示す、 図2B)。
- 4°Cで1回の洗浄につき5分間、MiR05で繊維を2回洗浄します。
- ろ紙でブロットドライし、2.1mLのMiR05で満たされたチャンバーに加える前に秤量する。
- 完全に閉じずにストッパーを導入し、20mLシリンジを使用して2mLの純粋なO2 でチャンバを酸素化し、回転運動でストッパーをねじってチャンバを閉じます。酸素拡散の制限を避けるために、実験中はO2 濃度を300〜500μMに保ちます。
- 細胞または組織における日常的な呼吸を評価するためのプロトコル
- ステップ1.5.2-1.5.3で述べたようにチャンバーにサンプルを加える。
- 2.3 mL の温かい MiR05 細胞懸濁液を追加します (標準入力: ステップ 1.2 では 1 x106 cells/mL、ステップ 1.3 では 2 mg/mL)
- 骨格筋および心筋(ステップ1.4):ステップ1.4.4-1.4.6を考慮して、予め温めた2.3mLの暖かいMiR05に約4mgのサポニン透過繊維を加える
- チャンバーを37°Cで実行し、攪拌速度700rpmとした。>3分間待ってから、媒体が脱気し、回転運動でストッパーをねじってチャンバーを閉じます。ペルチェブロック安定化は、設定温度に達することを示す。
- (オプション)スターラーの速度を 300 rpm に変更して、残りの気泡がストッパーのキャピラリーから逃げるようにします。
- 余分な液体をストッパーの上に吸引します。任意のサンプルタイプで安定した酸素流束信号が達成されるまで10分間待って、ルーチン/状態1呼吸を記録します( 図1B)。
- 透過処理された細胞および組織における呼吸測定については、ステップ1.6に進みます。手順 1.8 の無傷のセルの場合。
- 透過処理された細胞または組織におけるOXPHOS分析のためのプロトコル
- 溶解(透過処理)組織サンプルを使用するか、終濃度5 μg/mLで1 μLのジギトニン(ジメチルスルホキシド(DMSO)中の8.1 mMジギトニンストック)を加えて細胞を透過処理し、細胞を透過処理します。フラックスは低下し、>5分で安定するはずです。
注意: ジギトニンは、気道に急性毒性があります, 皮膚と接触して, または飲み込んだとき.
注:すべての化学物質の注入は、精密ガラスシリンジで行われます。シリンジは、交差汚染を避けるために指示された化学物質にのみ使用し、使用後は水とEtOHで徹底的に洗ってください。ブロックされたシリンジは、暖かいddH2 Oまたはクリーニングワイヤーで超音波処理を使用して、化学的な目詰まりを取り除く必要があります。チャンバーに空気が導入されないように、それぞれの原液の余剰分を常にシリンジに引き込んでください。各注射後に空気が導入されていないかチャンバーの内部を検査します。フラックスがプラトーするまで各ステップを記録します。 - 5 μLの0.4 Mリンゴ酸塩(M)を終濃度1 mM、5 μLの2.0 Mピルビン酸(P;新たに調製)、終濃度5 mM、4 μLの2.5 Mグルタミン酸(G)を終濃度5 mMで急速に連続して加える。
- 以前のフラックスを頭打ちした後、5 μL(筋肉組織の場合は10 μL)の0.5 Mアデノシン二リン酸(ADP、-80°Cで保存されたアリコート)を最終濃度1.25 mMで加える。
注:筋肉などの組織は、飽和に達するために異なる濃度を必要とするかもしれません。 - 5 μL の 4 mM シトクロム C (cytC) を最終濃度 10 μM で加える。
注: 細胞が透過処理の品質を評価するためのオプションです。 - 16 μL の 1.25 M コハク酸塩 (S) を終濃度 10 mM で加える。(オプション)3 μL の 0.5 M ADP を最終濃度 2 mM で加え、ADP 濃度の飽和を制御します。
- 細胞および非繊維組織の場合、2 μL の 1 mg/mL オリゴマイシン (OM) を最終濃度 1 μg/mL で加えます。
警告: 使用されるすべての ETS 阻害剤は毒性が強いです。
注:オリゴマイシンは、ETS能力を抑制することができ、筋肉組織では省略されているため、最適な濃度のために滴定が必要な場合があります。筋肉組織がアッセイされ、O2 が300μM未満の場合、ここで再酸素化される。 - 2 mM ストックから FCCP を滴定し、呼吸の増加がなくなるまで 0.6 μL を加え、その後の 0.2 μL ステップを加えます (理論上:非結合)。
- 終濃度0.5 μMの場合、1 μLの1 mMロテノン(ROT)を加え、終濃度1 μg/mLのアンチマイシン(AM)ストックを2 μL加える。
- チャンバを再酸素化して、プランジャーをねじれ運動でゆっくりと持ち上げることによって、すべてのチャンバで同様の酸素レベル(〜150μM)を達成します。
- 5 μLの0.8 Mアスコルビン酸塩を終濃度2 mMで直ちに加え、続いて5 μLの0.2 MN,N,N',N'-テトラメチル-p-フェニレンジアミン(TMPD)を終濃度0.5 mMで加え、複合体IV活性を評価します(オプション)。
- TMPDでピークO2フラックスに達したらすぐに終濃度10mMの4Mアジ化物を5μL 加える。>5分間実行を継続し、複合体IV塩基レベル計算のためにTMPDの自動酸化をアッセイする。
- セルを再カウントして、実行前にセル数を確認し、手順 1.9 に進みます。
注:デジトニン透過処理(細胞のみ)は、最大流束に達し、ミトコンドリア膜の完全性に影響を与えないように、試験実験で滴定する必要があります(ステップ1.7を参照)。シトクロムcの添加後に呼吸数が>10%増加する透過処理サンプル(特に筋肉組織)は、ミトコンドリア外膜損傷のためにさらなる分析から除外されるべきである。EtOH溶解化学物質の添加後、フラックスに短時間浸漬することが予想される。
- 溶解(透過処理)組織サンプルを使用するか、終濃度5 μg/mLで1 μLのジギトニン(ジメチルスルホキシド(DMSO)中の8.1 mMジギトニンストック)を加えて細胞を透過処理し、細胞を透過処理します。フラックスは低下し、>5分で安定するはずです。
- 細胞に最適な透過処理条件を決定するためのプロトコル
- 手順 1.2 および 1.5.2 の説明に従ってセルを追加します。
- 10 μL の 10 mg/mL ジギトニンストックを摂取し、10 μL の DMSO を加えて 5 mg/mL に希釈します。
- 1 μLのロテノン(1 mMストック)を加える。10 μL のコハク酸塩 (2 mM ストック) および 5 μL の ADP (0.5 M ストック) を加える。
- 1 μLのジギトニン(1ステップあたり2.5 mg)を繰り返し滴定し、呼吸がそれ以上増加しず、最大になるまで滴定する。
注:呼吸の減少は、ジギトニンの過剰な濃度を示す。
- 無傷の細胞におけるOXPHOS分析のためのプロトコル
- ルーチン呼吸後(ステップ1.6.1〜1.6.6)、最終濃度10nMの0.01mMオリゴマイシンを2μL加える。
- 2 mM ストックから FCCP を滴定し、呼吸と呼吸が最大限結合解除されるまで、0.6 μL を後続の 0.2 μL ステップで加えます (理論:非結合)
- 終濃度0.5 μMの場合、1 μLの1 mMロテノンを加え、最終濃度1 μg/mLのアンチマイシンストックを2 μL加える。
- チャンバーを同じ酸素レベル(約150μM)に再酸素化するには、プランジャーをねじれ運動でゆっくりと持ち上げます。
- 5 μLの0.8 Mアスコルビン酸塩を加え、最終濃度2 mMにする。直ちに5 μLの0.2 M TMPDを終濃度0.5 mMで加え、複合体IV活性を評価した。
注:TMPDは自動酸化を起こしやすいため、より大きな実験セットの前に新しいバッチを準備してください。活性は、-20°Cで保存すると時間の経過とともに低下する可能性があります。 - TMPDでピークO2フラックスに達したらすぐに終濃度10mMの4Mアジ化物を5μL 加える。>5分間実行を継続し、複合体IV塩基レベル計算のためのTMPDの自動酸化をアッセイする。
- セルを再カウントして、実行前にセル数を確認し、手順 1.9 に進みます。
- 実行後のサンプル コレクション
- 各チャンバーから正確に 1 mL の MiR05 懸濁液を (スターラーをオンにして) 1.5 mL チューブに集めます。
- 透過処理された細胞の場合は1000 x gで、組織溶解物の場合は20,000 x gで遠心分離機。上清を除去し、ペレットを-80°Cで凍結してさらに処理する(セクション3)。
- SUITプロトコルの分析
- 基質または阻害剤を添加した後、各プラトーの酸素フラックス(pmol/s、入力に正規化)を分析します(図1C および 図3A)。値をスプレッドシートにエクスポートします。
- 各実験実行のすべての値から残留酸素消費量(ROX、 図1C および 図3C)値を差し引く。TMPDからアジ化物残留呼吸を差し引いて、複雑なIV呼吸を得る。
- 細胞(図3A、B)または組織入力(図5A、B)について正規化した絶対値をプロットします。フラックス制御比を計算するか(ステップ1.11)、タンパク質入力に正規化します(図3C)。
- 磁束制御比計算
- フラックス制御比(FCR)9,26を用いて呼吸機能とカップリング制御の指標を取得する。
注:これにより、ミトコンドリアの量とは無関係に、固有のミトコンドリアの品質を評価することができます。さらに、フラックス制御比(FCR)は、試薬の品質管理を可能にする同じ細胞株内で同等である(各FCRは、図1B-Dおよび図3Cに示された番号付き基準値を介して得られる)。 - 式1を使用して、OXPHOSとLEAKの結合に対する呼吸制御比を計算します。
式 1: FCRADP = 5/6 = 状態 3 / 状態 4 - 式2を使用してNADH依存呼吸を評価するためにFCRを計算する
式 2: FCR 状態 3 (I) = 3/5 = 状態 3 (I) /状態 3 ( I+II) - 式3を使用してコハク酸依存性呼吸を評価するためにFCRを計算する。
式 3: FCR状態 3 (II) = 8/7 = S腐敗 / ETS容量 - 式4を使用して、結合と非結合を評価するFCRを計算します。
式4:FCR結合/非結合 = 5/7 =状態3(I + II)/ ETS容量 - ミトコンドリア外膜の完全性をテストするには、式5を使用します。
式5:%ミトコンドリア外膜損傷= 3/4 = 状態3(I)/状態3(I)とcyt c
- フラックス制御比(FCR)9,26を用いて呼吸機能とカップリング制御の指標を取得する。
2. 高解像度呼吸法:マイクロプレートベースの呼吸数計(mHRR)
注:プロトコルのこのセクションの実験は、タツノオトシゴXFe96細胞外フラックスアナライザー(材料表)を使用して実施されました。
- 細胞培養
- 任意のタイプの細胞を培養する。付着剤(例えば、コラーゲン、ラミニン)は、細胞付着を促進するために使用され得る。ここで、HEK293細胞は、先と同様に培養される(工程1.3)。
- 実験の前日に細胞を剥離し、指定されたmHRR 96ウェルマイクロプレートに移して、実験当日に理想的なコンフルエンシー(80%-100%)を得た(図2C)。
注: mHRR の場合、マイクロプレートの細胞密度は非常に重要です。増殖に影響を及ぼす細胞株または治療の個々の増殖特性は、実験当日に同等のコンフルエンシーを確保するために考慮される必要がある。
- 高分解能呼吸測定のための細胞の調製
- 播種前に細胞を採取し、十分に再懸濁する
注: 複製のために同じ希釈液から細胞を播種することをお勧めします。 - 個々の細胞株の増殖速度または処理中の増殖特性に従って細胞を播種する。
注: 標準の 96 ウェルマイクロプレートで最適化し、細胞密度を 96 ウェルアッセイ固有のマイクロプレートに外挿します。このセットアップでは、7 x104 HEK293 WT細胞を、96ウェルのウェルあたりに播種した。96ウェルプレートの最初と最後のカラムは、タンパク質測定に使用されます(図2C)。4つのコーナーウェルには細胞が含まれてはならず、実験的なバックグラウンド補正に使用されます。理想的には、エッジ効果を最小限に抑えるために、エッジに近いウェルは空である(例えば、細胞は温度効果によって引き起こされる変化した増殖速度を示す)(図2C、D)。
- 播種前に細胞を採取し、十分に再懸濁する
- センサープレートの作製、阻害剤の装填
- アッセイの日に、38.8mLの培地に0.4mLの1Mグルコース、0.4mLの200mMグルタミン、および0.4mLの100mMNa-ピルビン酸塩を補充した。
注:mHRR呼吸には、pH 7.4の特殊な非緩衝DMEM培地が必要です。一般に、1つの96ウェルマイクロプレートで1回の実験を行うには40mLで十分です。 - 呼吸アッセイ培地を37°Cに加温し、細胞培養培地を1ウェルあたり80 μLで2回洗浄して呼吸アッセイ培地に交換した。
- アッセイの前に60分間CO2 を含まない37°Cのインキュベーターに細胞を入れたプレートをセットする。
注:CO2 は呼吸結果に影響を与える可能性があり、培地中の血清はアッセイ中に気泡を生成する可能性があるため、このステップはプレートを脱気するために不可欠です。 - OM、FCCP、ROT、およびAMの阻害剤アリコートを37°Cに予温し、センサープレートをインキュベーターから取り出します。
- OM、FCCP、ROT、および AM を 3 mL のアッセイ培地で希釈し、それぞれ最終ウェル濃度が 1.5 μM、1.125 μM、および 1 μM になるようにします。 図2Eに示すように、別々のポートに入力します。
メモ: センサーカートリッジを充填するには、マルチチャンネルピペットを使用することをお勧めします。加圧空気は化合物の注入に使用されるため、ポートが化合物で満たされるたびに、すべてのポートに同量の液体容量を充填する必要があります。ROTとAMは1つのポートで組み合わせることができます。阻害剤は、EtOHまたはDMSOに溶解させることができる。 - 注入ポートを検査し、各ポートのローディング・ボリュームが均等であることを確認します。
メモ: すべてのポートの底部には、注入用の穴があります。センサープレートを動かすときは注意が必要です。気泡は針を用いて除去することができる。
- アッセイの日に、38.8mLの培地に0.4mLの1Mグルコース、0.4mLの200mMグルタミン、および0.4mLの100mMNa-ピルビン酸塩を補充した。
- 無傷の細胞における酸素評価のためのプロトコル
- アッセイの前日に、ステップ2.4.2〜2.4.7を行う。
- 20 mLのキャリブラント溶液を50 mLの円錐管にアリコートする。
- 細胞外フラックスアッセイキットを開き、内容物を取り出します。
- センサーカートリッジを反転させてユーティリティプレートの横に置きます。200 μLのキャリブラント溶液をユーティリティプレートの各ウェルにピペットで入れます。
- センサーカートリッジをユーティリティプレートに取り付け、すべてのセンサーが水没していることに注意してください。
- プレートをCO2を含まない37°Cのインキュベーターに一晩または最低12 時間置く。インキュベーター内の湿度がキャリバートの蒸発を防ぐのに十分であることを確認します。
- マイクロプレートベースのシステムとコンピュータの電源を入れて、翌日使用できる状態にします(アッセイを実施する前に、37°Cに平衡化するために最低3時間かかります)。
メモ: 信号の安定性を保つために、呼吸状態ごとに測定サイクルを 3 回ではなく 6 回に増やしてください。各サイクルは、3分間の混合と3分間の測定で構成されています。 - XFアッセイの日に、ステップ2.4.9〜2.4.20を行う。
- 細胞培養プレートのコンフルエンシー、細胞の形態、およびバックグラウンドウェルが空であることを確認します。
- ステップ2.4.11-2.4.12で述べたように、調製した呼吸培地で細胞を洗浄する。
- 各ウェルから20 μLを除くすべての培養培地を除去します。培養液が一晩の蒸発により80 μLであった場合、55 μL(約5 μL)を除去した。
- 細胞を90 μLのアッセイ培地で2回洗浄する。最後に、100μLのアッセイ培地を添加する。最終容量は120μLでなければならない。
注: このステップでは、各実験条件に同じ洗浄手順が適用されていることを確認するために、マルチチャンネルピペットをお勧めします(プレートの設定によって異なります)。吸引するときは、プレートを45°の角度に傾け、ピペットチップをウェルの角に置き、液体の吸引と注入を行います。特定の細胞が細胞培養プレートの底から容易に剥離する可能性があるため、洗浄中に注意することが不可欠です。 - アッセイの前に60分間CO2 を含まない37°Cのインキュベーターにプレートをセットする。
- 水和センサーカートリッジプレートをCO2フリーインキュベーターから取り出します。
- 古いキャリブラント溶液を捨て、37°Cに予温した新しいキャリブラント溶液と交換してください。
- インヒビターとアッセイ培地 (インヒビターあたり 3 mL、合計 12 mL のアッセイ培地) を調製し、ポートへのインヒビターのロードにピペットリザーバを使用します。
- ソフトウェアを開き、事前に設計されたテンプレートまたは新しいテンプレートを実行します。プレートマップに入力し、滴定と測定サイクルを調整してから、 開始 を押して光学センサーのキャリブレーションを開始します。
- ロードされたカートリッジから蓋を取り外し、自動的にマシンからスライドするスロットに置き、プレートの右下隅のマーキングがスロットの右下隅の三角形と揃っていることを確認します。
- [ 続行] をクリックして自動キャリブレーションを実行し、約20分間持続します。
- 校正後、キャリバートを含むユーティリティプレートを取り外します。
- 細胞が入ったマイクロプレートから蓋を取り外し、機械の指示に従ってプレートをスロットに置きます。[ 続行] をクリックして実行を開始します。
- 実行後のサンプル コレクション
- プレートを機械から取り出し、細胞を乱すことなく残りのアッセイ培地を慎重に取り出し、プレート全体を-80°Cで凍結してさらに処理します(セクション3)。
ビシンコニン酸アッセイ(BCAアッセイ)を用いたタンパク質の定量
- タンパク質抽出に使用した緩衝液で希釈したウシ血清アルブミン(BSA)を調製し、BCAと適合する:2mg/mL、1.5mg/mL、1mg/mL、0.5mg/mL、0.25mg/mLおよび0mg/mLの標準曲線を複製する。
- mHRRの場合は1ウェルあたり20 μL、またはcHRRの場合は1.5 mLチューブ内に含まれるペレットあたり100 μLの適切な溶解バッファー(RIPAなど)に再懸濁してタンパク質を抽出します。
- タンパク質溶解物を含むmHRRプレートまたは1.5mLチューブを氷上で30分間インキュベートする。
- タンパク質溶解物を入れた1.5 mLチューブを4°C、20,000 x g で20分間遠心分離し、得られた上清を新しいきれいな1.5 mLチューブに移した。
- マイクロタイタープレートの複製および標準でサンプルあたり10 μLを使用します。200 μLのBCA作動試薬を加え、>15分間インキュベートする。
- 波長562nmの標準分光光度計でプレートを読み取り、BSA標準曲線を使用してタンパク質濃度を計算します。
- 呼吸結果をタンパク質濃度に正規化する。
注:タンパク質量への正規化により、細胞播種密度または湿重量入力を裏付けることができる。抽出されたタンパク質は、例えばETSのサブユニットに対するその後のイムノブロッティングに適しているが、天然サンプルを完全には表していない(例えば、リン酸化部位の喪失)。
結果
ここでは、真核細胞、非線維組織(例えば、小脳)、および線維組織(例えば、骨格筋)におけるミトコンドリア生体エネルギーを決定するためのプロトコルを提供する。真核細胞について、複数の(CRISPRKO1)および重度/完全なOXPHOS欠損症(CRISPR KO2)をもたらすミトコンドリア翻訳に関連する2つの異なるタンパク質のCRISPR操作によるノックアウトを有するHEK293を、cHRR(図3A-C)またはmHRR(<...
ディスカッション
伝統的に、ミトコンドリア生体エネルギー学は、クラーク型酸素電極を用いて研究されてきた。しかし、解像度とスループットの欠如は、技術の進歩を正当化しました。今日まで、O2k(cHRRと呼ばれる)およびタツノオトシゴXF96フラックスアナライザー(mHRRと呼ばれる)は、細胞バイオエネルギー学の分野で広く採用されている。ここでは、cHRRまたはmHRRのいずれかを使用してミトコンドリア呼吸?...
開示事項
開示する利益相反の禁止。
謝辞
この研究は、フィンランドアカデミー(C.B.J)、Magnus Ehrnroot Foundation(C.B.J)、および統合生命科学大学院(R.A.)の博士フェローシップからの資金提供によって支援されました。
資料
| Name | Company | Catalog Number | Comments |
| 2 mL Potter-Elvehjem Glass/PTFE Tissue Grinder/Homogenizer | Omni International | 07-358029 | |
| 95% O2, 5% CO2 medical gas mixture | Potter for tissue grinding | ||
| ADP | Sigma | A 4386 | |
| Antimycin A | Sigma | A 8674 | Chemical |
| Ascorbate | Merck | PHR1279-1G | Chemical, dissolve in ethanol |
| BSA (fatty accid free) | Sigma | A 6003 | Chemical |
| CaCO3 | Sigma | C 4830 | Chemical |
| Cytochrome c | Sigma | C 7752 | Chemical |
| Digitonin | Sigma | D 5628 | Chemical |
| Dithiothreitol | Sigma | D 0632 | Chemical, dissolve in DMSO |
| D-Sucrose | Roth | 4621.1 | Chemical |
| Dulbecco’s modified Eagle’s medium (High glucose) | Fisher Scientific | 41965-039 | Chemical |
| Dulbecco’s modified Eagle’s medium (No Glucose) | Fisher Scientific | A14430-01 | |
| EGTA | Sigma | E 4378 | |
| Etomoxir | Sigma | E1905 | Chemical |
| Falcon 15 ml Conical Centrifuge Tubes | Fisher Scientific | AM12500 | Chemical |
| Falcon 50 ml Conical Centrifuge Tubes | Fisher Scientific | AM12501 | |
| FCCP | Sigma | C 2920 | |
| Glucose | Sigma | G7021 | Chemical, dissolve in ethanol |
| Glutamate | Sigma | G 1626 | Chemical |
| GlutaMax (100x) (200 nM L-alanyl-L-glutamine dipeptide) | Fisher Scientific | 35050061 | Chemical |
| HEK293 cells | ATTC | CRL-1573 | |
| Hemocytometer | Fisher Scientific | 0267151B | Instrument for cell counting |
| Hepes | Sigma | H 7523 | Chemical |
| Imidazole | Fluka | 56750 | Chemical |
| KCl | Merck | 1.04936 | Chemical |
| L-carnitine | Sigma | C0283 | Chemical |
| Malate | Sigma | M 1000 | Chemical |
| MES hydrate | Sigma | M8250 | Chemical |
| MgCl2 | Sigma | M 9272 | Chemical |
| Na2ATP | Sigma | A 2383 | Chemical |
| Na2Phosphocreatine | Sigma | P 7936 | Chemical |
| Na-pyruvate (100 mM) (100x) | Fisher Scientific | 11360070 | |
| NEAA (Non-essential amino acids) 100x | Fisher Scientific | 11140035 | |
| Normal FBS (10x) | Fisher Scientific | 10500064 | |
| O2k-Core: Oxygraph-2k | Oroboros Instruments | 10000-02 | High-resolution respirometry instrument |
| O2k-Titration Set | Oroboros Instruments | 20820-03 | Hamilton syringes for chemical injections |
| Oligomycin | Sigma | O 4876 | Chemical, dissolve in ethanol |
| Palmitoylcarnitine | Sigma | P 4509 | Chemical |
| Penicillin-Streptomycin | Fisher Scientific | 15140122 | |
| Pierce BCA Protein Assay Kit | Fisher Scientific | 23227 | |
| Pyruvate | Sigma | P 2256 | Chemical |
| RIPA-Buffer | Fisher Scientific | 89900 | Chemical |
| Rotenone | Sigma | R 8875 | Chemical, dissolve in ethanol |
| Saponin | Sigma | S7900 | Chemical |
Seahorse XF DMEM assay medium pack, pH 7.4 | Agilent, Santa Clara, CA | 103680-100 | |
| Seahorse XFe96 Extracellular Flux Analyzer | Agilent, Santa Clara, CA | High-throughput respirometry instrument | |
| Seahorse XFe96 FluxPak | Agilent, Santa Clara, CA | Includes assay plates, cartridges, loading guides for transferring compounds to the assay cartridge, and calibrant solution. | |
| Small scissors | Fisher Scientific | 08-951-20 | |
| Sodium azide | Sigma | S2002 | Chemical |
| Succinate | Sigma | S 2378 | Chemical |
| Taurine | Sigma | T 8691 | Chemical |
| TMPD | Sigma | T 3134 | Chemical |
| Trypan Blue solution | Merck | 72-57-1 | Chemical |
| Trypsin 0.25% EDTA | Fisher Scientific | 25200056 | |
| Two thin-edged forceps | Fisher Scientific | 12-000-122 | |
| Uridine stock (500x) | Sigma | U3750 | Chemical |
参考文献
- McBride, H. M., Neuspiel, M., Wasiak, S. Mitochondria: More than just a powerhouse. Current Biology. 16 (14), 551-560 (2006).
- Mehta, M. M., Weinberg, S. E., Chandel, N. S. Mitochondrial control of immunity. Beyond ATP. Nature Reviews Immunology. 17 (10), 608-620 (2017).
- Spinelli, J. B., Haigis, M. C. The multifaceted contributions of mitochondria to cellular metabolism. Nature Cell Biology. 20 (7), 745-754 (2018).
- Gnaiger, E. Capacity of oxidative phosphorylation in human skeletal muscle. New perspectives of mitochondrial physiology. International Journal of Biochemistry and Cell Biology. 41 (10), 1837-1845 (2009).
- Gorman, G. S., et al. Mitochondrial diseases. Nature Reviews Disease Primers. 2, 1-23 (2016).
- Boushel, R., Gnaiger, E., Schjerling, P., Skovbro, M., Kraunsøe, R., Dela, F. Patients with type 2 diabetes have normal mitochondrial function in skeletal muscle. Diabetologia. 50 (4), 790-796 (2007).
- Cogliati, S., et al. Mitochondrial cristae shape determines respiratory chain supercomplexes assembly and respiratory efficiency. Cell. 155 (1), 160-171 (2013).
- Kühlbrandt, W. Structure and function of mitochondrial membrane protein complexes. BMC Biology. 13, 1-11 (2015).
- Gnaiger, E. Mitochondrial pathways and Respiratory control. An introduction to OXPHOS analysis. Bioenergetics communications. 5th ed. , (2020).
- Jackson, C. B., et al. Mutations in SDHD lead to autosomal recessive encephalomyopathy and isolated mitochondrial complex II deficiency. Journal of Medical Genetics. 51 (3), 170-175 (2014).
- Pesta, D., Gnaiger, E. High-resolution respirometry: OXPHOS protocols for human cells and permeabilized fibers from small biopsies of human muscle. Methods in Molecular Biology. 810, 25-58 (2012).
- Horan, M. P., Pichaud, N., Ballard, J. W. O. Review: Quantifying mitochondrial dysfunction in complex diseases of aging. Journals of Gerontology - Series A Biological Sciences and Medical Sciences. 67 (10), 1022-1035 (2012).
- Doerrier, C., Garcia-Souza, L. F., Krumschnabel, G., Wohlfarter, Y., Mészáros, A. T., Gnaiger, E. High-resolution fluorespirometry and oxphos protocols for human cells, permeabilized fibers from small biopsies of muscle, and isolated mitochondria. Methods in Molecular Biology. 1782, 31-70 (2018).
- Zhang, J., et al. Measuring energy metabolism in cultured cells, including human pluripotent stem cells and differentiated cells. Nature Protocols. 7 (6), 1068-1085 (2012).
- García-Roche, M., Casal, A., Carriquiry, M., Radi, R., Quijano, C., Cassina, A. Respiratory analysis of coupled mitochondria in cryopreserved liver biopsies. Redox Biology. 17, 207-212 (2018).
- Acin-Perez, R., et al. A novel approach to measure mitochondrial respiration in frozen biological samples. The EMBO Journal. 39 (13), 1-18 (2020).
- Cell metabolism assay kits. Seahorse assay kits and media Available from: https://www.agilent.com/en/product/cell-analysis/real-time-cell-metabolic-analysis/xf-assay-lits-reagents-cell-assay-media (2021)
- Chance, B., Williams, G. R. A method for the localization of sites for oxidative phosphorylation. Nature. 176 (4475), 250-254 (1955).
- Gnaiger, E., et al. Mitochondrial respiratory states and rates. MitoFit Preprint Arch. , (2019).
- Gnaiger, E. O2k-procedures: SOP O2k quality control 1: Polarographic oxygen sensors and accuracy of calibration Section Page. Oroboros. 03 (18), 1-21 (2020).
- Robinson, B. H., Petrova-Benedict, R., Buncic, J. R., Wallace, D. C. Nonviability of cells with oxidative defects in galactose medium: A screening test for affected patient fibroblasts. Biochemical Medicine and Metabolic Biology. 48 (2), 122-126 (1992).
- King, M. P., Attardi, G. Human cells lacking mtDNA: Repopulation with exogenous mitochondria by complementation. Science. 246 (4929), 500-503 (1989).
- Makrecka-Kuka, M., Krumschnabel, G., Gnaiger, E. High-resolution respirometry for simultaneous measurement of oxygen and hydrogen peroxide fluxes in permeabilized cells, tissue homogenate and isolated mitochondria. Biomolecules. 5 (3), 1319-1338 (2015).
- Fasching, M., Gnaiger, E. O2k quality control 2: Instrumental oxygen background correction and accuracy of oxygen flux. Mitochondrial Physiology Network. 14 (06), 1-14 (2016).
- Gnaiger, E., Lassnig, B., Kuznetsov, A., Rieger, G., Margreiter, R. Excess capacity of cytochrome c oxidase. Journal of Experimental Biology. 1139, 1129-1139 (1998).
- Gnaiger, E., et al. Mitochondria in the Cold. Life in the Cold. , 431-442 (2000).
- Fontana-Ayoub, M., Fasching, E., Gnaiger, Selected media and chemicals for respirometry with mitochondrial preparations. Mitochondrial Physiology Network. 02 (17), 1-9 (2014).
- Gerencser, A. A., et al. Quantitative microplate-based respirometry with correction for oxygen diffusion. Analytical Chemistry. 81 (16), 6868-6878 (2009).
- Krumschnabel, G., Eigentler, A., Fasching, M., Gnaiger, E. Use of safranin for the assessment of mitochondrial membrane potential by high-resolution respirometry and fluorometry. Methods in Enzymology. 542, 163-181 (2014).
- Nászai, A., Terhes, E., Kaszaki, J., Boros, M., Juhász, L. Ca(2+)N it be measured? Detection of extramitochondrial calcium movement with high-resolution fluorespirometry. Scientific Reports. 9 (1), 1-13 (2019).
- Pajak, B., et al. 2-Deoxy-d-Glucose and its analogs: From diagnostic to therapeutic agents. International Journal of Molecular Sciences. 21 (1), 234 (2019).
- Mercier-Letondal, P., Marton, C., Godet, Y., Galaine, J. Validation of a method evaluating T cell metabolic potential in compliance with ICH Q2 (R1). Journal of Translational Medicine. 19 (1), 1-15 (2021).
- Sauerbeck, A., et al. Analysis of regional brain mitochondrial bioenergetics and susceptibility to mitochondrial inhibition utilizing a microplate based system. Journal of Neuroscience Methods. 198 (1), 36-43 (2011).
- Jackman, M. R., Willis, W. T. Characteristics of mitochondria isolated from type I and type IIb skeletal muscle. American Journal of Physiology - Cell Physiology. 270 (2), 673-678 (1996).
- Ponsot, E., et al. Mitochondrial tissue specificity of substrates utilization in rat cardiac and skeletal muscles. Journal of Cellular Physiology. 203 (3), 479-486 (2005).
- Schönfeld, P., Reiser, G. Why does brain metabolism not favor burning of fatty acids to provide energy-Reflections on disadvantages of the use of free fatty acids as fuel for brain. Journal of Cerebral Blood Flow and Metabolism. 33 (10), 1493-1499 (2013).
- Calderon-Dominguez, M., Mir, J. F., Fucho, R., Weber, M., Serra, D., Herrero, L. Fatty acid metabolism and the basis of brown adipose tissue function. Adipocyte. 5 (2), 98-118 (2016).
- Divakaruni, A. S., Rogers, G. W., Murphy, A. N. Measuring mitochondrial function in permeabilized cells using the seahorse XF analyzer or a clark-type oxygen electrode. Current Protocols in Toxicology. 2014, 1-16 (2014).
- Iuso, A., Repp, B., Biagosch, C., Terrile, C., Prokisch, H. Assessing mitochondrial bioenergetics in isolated mitochondria from various mouse tissues using Seahorse XF96 analyzer. Methods in Molecular Biology. 1567, 217-230 (2017).
- Rogers, G. W., et al. High throughput microplate respiratory measurements using minimal quantities of isolated mitochondria. PLoS ONE. 6 (7), 21746 (2011).
- Jordá, A., Zaragozá, R., Portolés, M., Báguena-Cervellera, R., Renau-Piqueras, J. Long-term high-protein diet induces biochemical and ultrastructural changes in rat liver mitochondria. Archives of Biochemistry and Biophysics. 265 (2), 241-248 (1988).
- Jackson, C. B., Gallati, S., Schaller, A. QPCR-based mitochondrial DNA quantification: Influence of template DNA fragmentation on accuracy. Biochemical and Biophysical Research Communications. 423 (3), 441-447 (2012).
- Hirsch, H. M. Tissue autoxidation inhibitors: II. The presence of inhibitor in intact cells; Assay of liver and hepatoma effect on radio-oxidations. Cancer Research. 16 (11), 1076-1082 (1956).
- Picard, M., et al. Mitochondrial structure and function are disrupted by standard Isolation methods. PLoS ONE. 6 (3), 18317 (2011).
- Tanumihardja, E., Slaats, R. H., Van Der Meer, A. D., Passier, R., Olthuis, W., Van Den Berg, A. Measuring both pH and O2 with a single On-Chip sensor in cultures of human pluripotent stem cell-derived cardiomyocytes to track induced changes in cellular metabolism. ACS Sensors. 6 (1), 267-274 (2021).
- Harms, F., Stolker, R. J., Mik, E. Cutaneous respirometry as novel technique to monitor mitochondrial function: A feasibility study in healthy volunteers. PLoS ONE. 11 (7), 159544 (2016).
- Levitsky, Y., et al. Micro-respirometry of whole cells and isolated mitochondria. RSC Advances. 9 (57), 33257-33267 (2019).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved