È necessario avere un abbonamento a JoVE per visualizzare questo. Accedi o inizia la tua prova gratuita.
Method Article
Respirometria ad alta risoluzione per valutare la bioenergetica nelle cellule e nei tessuti utilizzando respirometri a camera e a piastre
In questo articolo
Riepilogo
La valutazione della fosforilazione ossidativa mediante respirometri ad alta risoluzione è diventata parte integrante dell'analisi funzionale dei mitocondri e del metabolismo energetico cellulare. Qui, presentiamo protocolli per l'analisi del metabolismo energetico cellulare utilizzando respirometri ad alta risoluzione basati su camera e micropiastre e discutiamo i principali vantaggi di ciascun dispositivo.
Abstract
La respirometria ad alta risoluzione (HRR) consente di monitorare la fosforilazione ossidativa in tempo reale per l'analisi dei singoli stati energetici cellulari e la valutazione dei complessi respiratori utilizzando protocolli diversificati di titolazione substrato-disaccoppiatore-inibitore (SUIT). Qui viene dimostrato l'uso di due dispositivi di respirometria ad alta risoluzione e viene presentata una raccolta di base di protocolli applicabili per l'analisi di cellule in coltura, fibre muscolari scheletriche e cardiache e tessuti molli come il cervello e il fegato. I protocolli per cellule e tessuti in coltura sono forniti per un respirometro a camera e cellule coltivate per un respirometro basato su micropiastre, entrambi comprendenti protocolli di respirazione standard. A scopo comparativo, le cellule HEK293 ingegnerizzate con CRISPR carenti di traduzione mitocondriale che causano una carenza multipla del sistema respiratorio vengono utilizzate con entrambi i dispositivi per dimostrare difetti cellulari nella respirazione. Entrambi i respirometri consentono una misurazione completa della respirazione cellulare con i rispettivi meriti tecnici e l'idoneità a seconda della domanda di ricerca e del modello in studio.
Introduzione
I mitocondri soddisfano la fornitura chiave di energia e sono un organello compartimentato che contribuisce a processi bioenergetici e metabolici cellulari essenziali come l'anabolismo di nucleotidi, lipidi e amminoacidi, biogenesi del cluster ferro-zolfo e sono implicati nella segnalazione come la morte cellulare controllata 1,2,3 . La bioenergetica mitocondriale attraverso la fosforilazione ossidativa contribuisce a quasi tutti i processi cellulari all'interno della cellula e, di conseguenza, le disfunzioni mitocondriali di origine primaria o secondaria sono associate a un ampio spettro di condizioni di malattia 4,5. La disfunzione mitocondriale comporta non solo alterazioni della struttura o della densità mitocondriale, ma anche della qualità e della regolazione dell'apparato respiratorio6. Questo elemento qualitativo comprende il controllo del substrato, le caratteristiche di accoppiamento, le modifiche post-traduzionali, la dinamica delle cristae e i supercomplessi respiratori 7,8. Pertanto, un'analisi accurata della bioenergetica mitocondriale per approcci sperimentali e diagnostici per valutare il metabolismo energetico della cellula è importante per la salute e la malattia.
La fosforilazione ossidativa mitocondriale (OXPHOS) è una sequenza di reazioni all'interno del sistema respiratorio o del sistema di trasferimento di elettroni (ETS) per la generazione di energia cellulare attraverso l'adenosina trifosfato (ATP)9. La fase multienzimatica per sfruttare l'energia dal flusso di elettroni attraverso i complessi I e II al complesso IV genera un gradiente protonico elettrochimico attraverso la membrana mitocondriale interna, successivamente utilizzato per la fosforilazione dell'adenosina difosfato (ADP) all'ATP tramite il complesso V (F1FO ATP sintasi) (Figura 1A).
In primo luogo, i vettori di due elettroni vengono generati durante il ciclo tricarbossilico (TCA), la glicolisi e l'ossidazione del piruvato: nicotinamide adenina dinucleotide (NADH) e diidroflavina adenina dinucleotide (FADH2). Il NADH viene ossidato nel complesso I (NADH deidrogenasi), durante il quale due elettroni vengono trasferiti al coenzima Q (il chinone viene ridotto a chinolo), mentre i protoni vengono pompati nello spazio intermembrana (IMS). In secondo luogo, il complesso II (Succinato deidrogenasi) ossida FADH2 e alimenta gli elettroni al coenzima Q senza pompare protoni. In terzo luogo, nel complesso III (citocromo c ossidoreduttasi), gli elettroni del coenzima Q vengono trasferiti al citocromo c mentre i protoni vengono pompati nell'IMS. In quarto luogo, il citocromo c trasferisce gli elettroni al complesso IV (citocromo c ossidasi), il complesso finale per pompare protoni e dove l'ossigeno funziona come un accettore di elettroni per assimilare i protoni, formando infine acqua. È questo ossigeno che i mitocondri consumano che può essere misurato da un ossigrafo. Infine, i protoni generati dal complesso I, dal complesso III e dal complesso IV vengono utilizzati per ruotare il complesso V, generando così ATP9.
È importante sottolineare che il trasferimento di elettroni avviene non solo in modo lineare, altrimenti indicato come la catena di trasporto degli elettroni. Invece, gli elettroni possono essere trasferiti al pool di coenzima Q attraverso più vie respiratorie e facilitare il flusso di elettroni convergenti. I substrati nadH e il succinato, ad esempio, possono entrare rispettivamente attraverso il complesso I e il complesso II. Gli elettroni dell'ossidazione degli acidi grassi possono essere donati tramite il complesso flavoproteico che trasferisce elettroni. In effetti, un'analisi completa di OXPHOS richiede un approccio olistico con substrati di carburante appropriati (Figura 1A).

Figura 1: Fosforilazione ossidativa mitocondriale e protocolli specifici di substrato e inibitori. (A) Mitocondrio e schema del sistema di trasferimento di elettroni (CI-CIV) e F1F0 ATP sintasi mitocondriale (CV). Tutte le strutture provengono da PDB. Le figure raffigurano solo substrati e inibitori descritti in questo studio). (B) Traccia campione di flusso di ossigeno in celle HEK293 intatte utilizzando il protocollo standard in un dispositivo mHRR. (C) Traccia campione di flusso di ossigeno in celle HEK293 intatte utilizzando il protocollo standard in un dispositivo cHRR. (D) Traccia campione del flusso di ossigeno nei fibroblasti umani permeabilizzati da un donatore sano con il rispettivo protocollo SUIT. Abbreviazioni: 1 = Respirazione di routine delle cellule intatte; 2 = Stato 2; 3 = Stato 3(I); 4 = Stato 3(I) con cytC; 5 = Stato 3 (I+II); 6 = Perdita(OM); 7 = capacità ETS; 8 = S(ROT); 9 = ROX; 10 = TMPD; 11 = Az. ROT = Rotenone, AM = Antimicina, ATP = Adenosina trifosfato, Az = Azide, OM = Oligomicina, FCCP = cianuro di carbonile p-trifluoro-metossifenil-idrazzone; Asc = Ascorbato, TMPD = N,N,N′,N′-tetrametil-p-fenilendiammina, Succ = Succinato, M = Malato, P = Piruvato, ADP = Adenosina difosfato, NAD = Nicotinamide adenina dinucleotide, IMS = Spazio intermembrana, FAD = Flavin adenina dinucleotide. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
L'analisi della capacità di OXPHOS mitocondriale utilizzando HRR è diventata un metodo biochimico strumentale di valore diagnostico non solo per i difetti mitocondriali primari10,11 ma estendendosi a tutti gli altri regni della biologia come il cancro e l'invecchiamento12. HRR consente la determinazione della respirazione cellulare mediante l'analisi della capacità mitocondriale oxphos, che riflette direttamente il deficit del complesso respiratorio mitocondriale individuale o combinato, e indirettamente è associato a disfunzione cellulare e metabolismo energetico alterato9. Metodologicamente, le misurazioni della respirazione vengono eseguite utilizzando cellule, tessuti o mitocondri isolati 11,13,14, con materiale congelato solo parzialmente adatto 15,16. Il tessuto congelato ha dimostrato di avere un ETS intatto con stabilità supercomplessa mantenuta15. Pertanto, a differenza dei tradizionali intermedi TCA, i rispettivi substrati vengono immessi direttamente nell'ETS. Tuttavia, l'accoppiamento tra la sintesi di ETS e ATP viene perso poiché l'integrità della membrana è compromessa da danni da congelamento (formazione di cristalli di ghiaccio).
Gli esperimenti di respirazione si svolgono normalmente a una temperatura fisiologica di 37 °C per gli endotermi in cellule o tessuti non permeabilizzati o permeabilizzati. Mentre il primo considera il contesto metabolico citosolico, il secondo fornisce il contributo energetico dei singoli complessi OXPHOS e dell'ATPasi attraverso l'aggiunta di specifici substrati (e inibitori). La sequenza e la variazione di substrati e inibitori hanno portato allo sviluppo di una vasta gamma di protocolli SUIT17 e saggi18 per affrontare varie questioni scientifiche sulla funzione di OXPHOS (esaminate al punto12). Il protocollo di base della respirazione cellulare valuta quattro diversi stati: i) respirazione di routine - la respirazione in un rispettivo mezzo respiratorio senza alcuna aggiunta di substrati o inibitori che consumano substrati ma endogeni. Questo stato può rivelare OXPHOS generale o difetti respiratori indotti secondariamente causati, ad esempio, da profili metabolici alterati. Successivamente, l'aggiunta dell'oligomicina inibitore dell'ATPasi rivela la permeabilità della membrana mitocondriale interna ai protoni, definita come ii) perdita respiratoria. La successiva titolazione di un protonoforo come il cianuro di carbonile uncoupler p-trifluoro-metossifenil-idrazzone (FCCP) consente di determinare lo stato in cui la capacità ETS è massima in un circuito protonico aperto-transmembrana, definito come iii) respirazione disaccoppiata. È importante sottolineare che uno stato disaccoppiato può verificarsi anche da interventi sperimentali attraverso un eccessivo danno meccanico alle membrane mitocondriali. Al contrario, lo stato non accoppiato si riferisce al disaccoppiamento respiratorio da parte di un meccanismo intrinseco fisiologicamente controllato. Infine, la completa inibizione dell'ETS mediante aggiunta dell'antimicina inibitore del complesso III e dell'inibitore del complesso I rotenone determina il consumo residuo di ossigeno (ROX) da processi non mitocondriali che consumano ossigeno (Figura 1A-C).
La bioenergetica mitocondriale è costituita da cinque distinti stati respiratori19,20. La respirazione dello stato 1 è senza substrati aggiuntivi o ADP, ad eccezione di ciò che è disponibile endogenamente. Dopo l'aggiunta di ADP, ma ancora, nessun substrato, si ottiene la respirazione dello stato 2. Quando vengono aggiunti substrati, consentendo il trasferimento di elettroni e la sintesi di ATP, viene raggiunta la respirazione dello stato 3. In questo stato, la capacità di OXPHOS può essere definita a concentrazioni saturanti di ADP, fosfato inorganico, ossigeno, NADH- e substrati legati al succinato. La respirazione allo stato 4 o la respirazione LEAK può essere definita come uno stato senza ADP o ATP sintasi chimicamente inibite pur avendo substrati sufficienti. Infine, quando tutto l'ossigeno è esaurito (anossico) in un ambiente a camera chiusa, si osserva la respirazione allo stato 5.
Esistono diversi metodi per valutare gli stati energetici cellulari14 con due dispositivi che dominano l'attuale valutazione in tempo reale di OXPHOS attraverso l'analisi del consumo di ossigeno, misurato come funzione della diminuzione dell'ossigeno nel tempo in un sistema a camera chiusa con diversa applicabilità a seconda del modello sperimentale e della domanda di ricerca: il respirometro ad alta risoluzione Oroboros 2k e l'analizzatore di flusso extracellulare Seahorse XF. Entrambi i dispositivi registrano i tassi di consumo di ossigeno come una diminuzione dei picomoli (pmol) di ossigeno (O2) al secondo come valore assoluto all'interno della camera o del pozzo della micropiastra. Il consumo specifico di ossigeno per massa si ottiene normalizzando il rispettivo consumo di ossigeno in una specifica ricetta tampone per numero di cellule (milioni), peso dei tessuti (mg) o quantità di proteine.
L'O2k (Oroboros Instruments) è un sistema chiuso a due camere dotato di un sensore polarografico di ossigeno (abbreviato in respirometro ad alta risoluzione basato su camera: cHRR). Ogni camera sperimentale contiene 2 ml di liquido che viene mantenuto omogeneo da agitatori magnetici. Il sensore polarografico di ossigeno utilizza un approccio amperometrico per misurare l'ossigeno: contiene un catodo d'oro, un anodo di cloruro d'argento / argento e tra una soluzione KCI che crea una cella elettrochimica su cui viene applicata una tensione (0,8 V). L'ossigeno proveniente dal mezzo di analisi si diffonde attraverso una membrana fluorurata di etilene propilene da 25 μm (permeabile A2) e subisce una riduzione al catodo, producendo perossido di idrogeno. All'anodo, l'argento viene ossidato dal perossido di idrogeno, generando una corrente elettrica. Questa corrente elettrica (ampere) è linearmente correlata alla pressione parziale dell'ossigeno. La pressione parziale dell'ossigeno e il fattore di solubilità dell'ossigeno del mezzo di saggio vengono utilizzati per calcolare la concentrazione di ossigeno. Poiché la pressione parziale dell'ossigeno dipende dalla temperatura sperimentale e le misurazioni polarografiche sono sensibili alla temperatura, le fluttuazioni di temperatura richiedono una regolazione precisa (±0,002 °C) da parte di un blocco riscaldante Peltier. La temperatura può essere controllata in un intervallo di 4 °C e 47 °C.
L'analizzatore di flusso extracellulare Seahorse XF (Agilent) è un sistema basato su piastre con formato micropiastra a 24 o 96 pozzetti in cui tre elettrodi di fluorescenza misurano il consumo di ossigeno nel tempo in ciascun pozzo (abbreviato in respirometro ad alta risoluzione basato su micropiastre: mHRR). Un massimo di quattro porte nella cartuccia del test sono disponibili per l'iniezione automatica durante il test. Un test contiene più cicli, ciascuno con tre fasi: 1) miscelazione, 2) attesa e 3) misurazione. Durante la fase di misurazione, le sonde del sensore vengono abbassate nella micropiastra creando una camera temporaneamente chiusa contenente un volume di 7-10 μL per misurare la luce emessa. Questa luce viene emessa da fluorofori incorporati in polimeri sulla punta delle sonde del sensore, che rilevano O2 in base alla tempra a fosforescenza. L'intensità del segnale di fluorescenza è proporzionale a O2 e influenzata dalla temperatura del sensore e del mezzo di saggio. Pertanto, una stima accurata dell'ossigeno richiede un approccio relativo con un pozzo di fondo senza alcun campione. Il ripristino della concentrazione di ossigeno avviene durante la fase di miscelazione quando il sensore si muove su e giù per mescolare il volume sopra la camera temporanea. Ogni ciclo calcola un tasso di consumo di ossigeno. La temperatura può essere controllata in un intervallo di 16 °C e 42 °C.
HRR è il gold standard per valutare la bioenergetica cellulare nelle malattie primarie e associate ai mitocondri e nel metabolismo cellulare generale. In questo studio, vengono forniti protocolli di base per HRR per valutare la funzione di OXPHOS nelle cellule e nei tessuti.

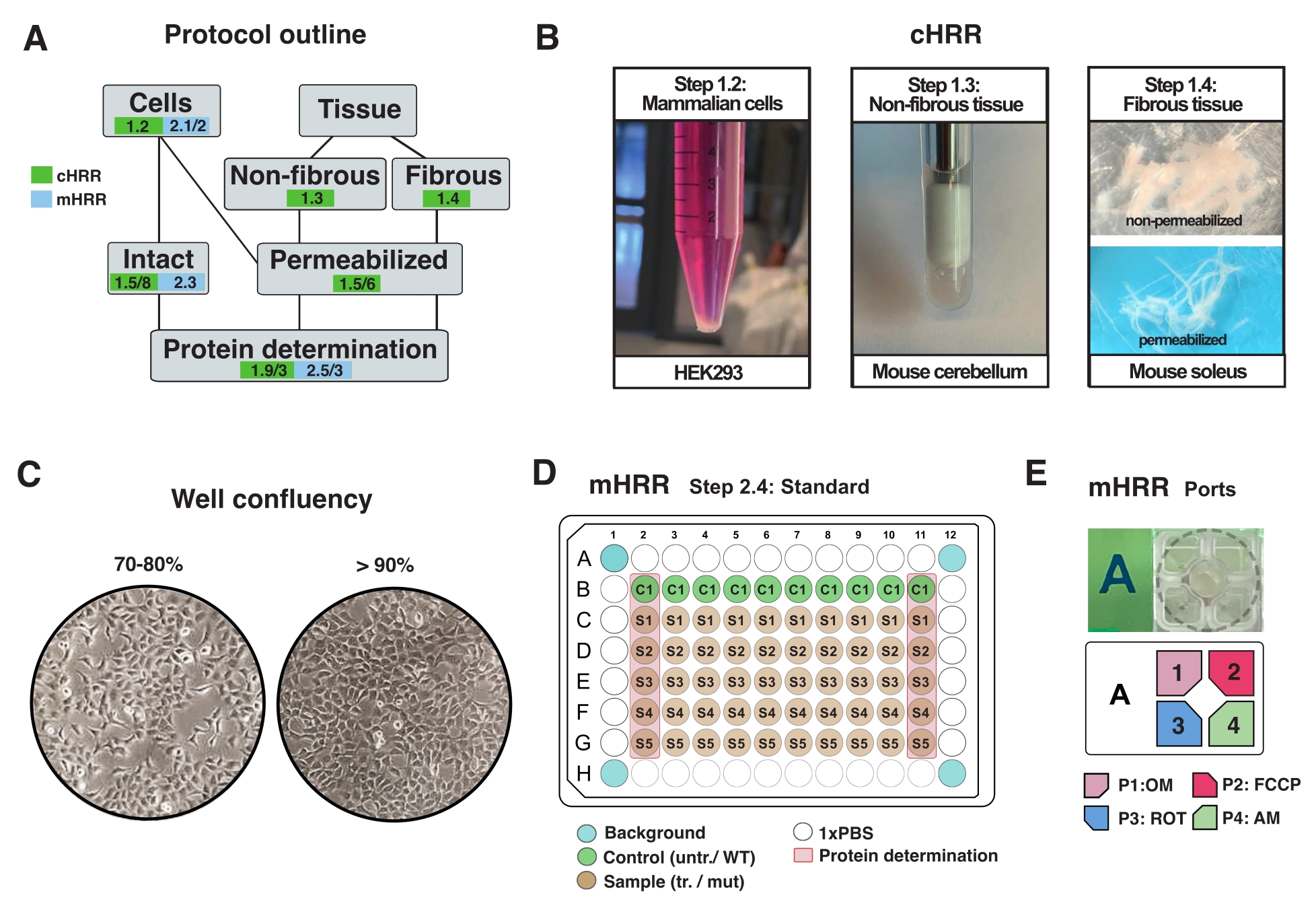
Figura 2: Flusso di lavoro per i preparati cellulari e tissutali per la cHRR e preparazione cellulare per la respirometria mHRR. (A) Schema dei protocolli forniti. (B) Cellule di mammifero (fase 1.2): hek293 pellet pari a 3 x 106 celle (pannello di sinistra). Tessuto non fibroso (fase 1.3): Preparazione del lisato di cervelletto murino in vasaio di teflon da 2 ml (pannello centrale). Permeabilizzazione del muscolo scheletrico indotta da saponina (pannello destro passo 1.4) per la respirometria cHRR. (C) Layout standard di semina delle micropiastre (fase 2.4) e controllo della confluenza per l'analisi delle cellule eucariotiche (HEK293) per la respirometria mHRR. (D, E) Schema di carico della porta di iniezione per la respirometria mHRR (fase 2.4). Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Protocollo
Tutta la sperimentazione animale viene eseguita in conformità con il National Animal Experiment Review Board e l'Agenzia amministrativa regionale statale per la Finlandia meridionale. In questo studio sono stati utilizzati topi maschi C57BL/6JOlaHsd (4-6 mesi) Il consenso per l'uso di linee cellulari umane è stato ottenuto dal comitato etico istituzionale dell'Università di Helsinki.
1. Respirometria ad alta risoluzione: respirometro a camera (cHRR)
NOTA: Gli esperimenti in questa sezione del protocollo sono stati eseguiti utilizzando l'Oroboros O2k-Core: Oxygraph-2k (Tabella dei materiali)
- Calibrazione dei sensori di ossigeno
- Respirometri pre-run a 37 °C in 2,1 mL di mezzo respiratorio mitocondriale (MiR05, Tabella 1, fattore di solubilità: 0,92) per >45 min ed eseguire la calibrazione dell'ossigeno come descritto21. Procedere se la variazione basale è compresa tra ± 4 pmol/s.
NOTA: Grandi fluttuazioni nel segnale di fondo potrebbero indicare la necessità di mantenere la membrana del sensore o tracce di inibitori rimasti nella camera da precedenti sperimentazioni. Si raccomanda una correzione strumentale del flusso di ossigeno di fondo prima di una serie di esperimenti25. - Registrare i valori di calibrazione dell'ossigeno per monitorare le prestazioni della membrana del sensore nel tempo.
NOTA: questo rivela la funzione del sensore, la stabilità dal segnale al rumore e quando è necessaria la manutenzione della membrana del sensore. A seconda della pressione ambiente, tra 180-200 μmol di ossigeno viene solubilizzato in MiR05. - Rimuovere tutto il liquido nella camera prima dell'aggiunta di qualsiasi campione nel mezzo respiratorio.
NOTA: Valutare regolarmente il volume delle camere respiratorie in modo che sia esattamente di 2 ml.
- Respirometri pre-run a 37 °C in 2,1 mL di mezzo respiratorio mitocondriale (MiR05, Tabella 1, fattore di solubilità: 0,92) per >45 min ed eseguire la calibrazione dell'ossigeno come descritto21. Procedere se la variazione basale è compresa tra ± 4 pmol/s.
- Preparazione di celle per respirometria ad alta risoluzione
- Coltiva cellule HEK293 in piatti da 10 cmdi diametro 2 nel mezzo dell'aquila modificata (DMEM) di Dulbecco con glucosio alto integrato con il 10% di siero bovino fetale inattivato dal calore (FBS), GlutaMax, aminoacidi non essenziali e Na-piruvato22 e uridina23 per supportare il metabolismo difettoso di OXPHOS in un incubatore a 37 °C al 5% di CO2.
NOTA: Qualsiasi tipo di cellula eucariotica può essere coltivata. Per la maggior parte dei tipi di cellule, la coltivazione di un piatto di 10 cm2 porta a cellule sufficienti (di solito >3 x 106 cellule). Controllare regolarmente l'infezione da micoplasma per evitare effetti sul metabolismo cellulare e sulla respirazione. - Far crescere le cellule senza superare il 90% di confluenza (Figura 2C).
NOTA: Le cellule con confluenza >90% possono mostrare effetti inibitori dipendenti dalla crescita sulla respirazione (se non sincronizzati o post-mitotici). - Lavare le cellule con 1x PBS, staccare con 1 mL di tripsina calda allo 0,25%, disattivare la tripsina aggiungendo DMEM caldo (5 ml / 10 cm2 piastra) e contare le cellule con un emocitometro.
- Centrifugare delicatamente la soluzione cellulare pari a 2,5 x 106 celle a 300 x g per 5 minuti, rimuovere completamente il surnatante e risospese in 2,5 mL di MiR05 caldo (1 x 106 celle/mL) (Figura 2A).
- Per le celle in sospensione, contare e rimuovere la soluzione pari a 2,5 x 106 celle, pellet e continuare come indicato al punto 1.2.4.
- Eseguire il protocollo SUIT per l'ottimizzazione della permeabilizzazione (passaggio 1.6), la cellula o il tessuto permeabilizzato (passaggio 1.5) o le cellule intatte (passaggio 1.7)
NOTA: Per risultati coerenti, si raccomanda di mantenere costante la concentrazione cellulare (ad esempio, 1 x 106 cellule/ml). Sebbene la respirazione sia indipendente dalla densità cellulare nel respirometro24, i substrati e gli inibitori sono in concentrazione comparabile durante gli esperimenti se il numero di cellule viene mantenuto costante.
- Coltiva cellule HEK293 in piatti da 10 cmdi diametro 2 nel mezzo dell'aquila modificata (DMEM) di Dulbecco con glucosio alto integrato con il 10% di siero bovino fetale inattivato dal calore (FBS), GlutaMax, aminoacidi non essenziali e Na-piruvato22 e uridina23 per supportare il metabolismo difettoso di OXPHOS in un incubatore a 37 °C al 5% di CO2.
- Preparazione di tessuto non fibroso (ad esempio, cervello, fegato) per la respirometria ad alta risoluzione
- Asportare un pezzo di tessuto omogeneo, 30-40 mg di peso, o utilizzare l'intero organo (cervelletto di topo in questo caso).
NOTA: se il tessuto non viene utilizzato immediatamente, conservare 2 mL di MiR05 ghiacciato consentendo la conservazione fino a 2 ore per la maggior parte dei tessuti. I singoli tempi di conservazione dei tessuti devono essere valutati in serie temporali. - Asciugare il tessuto con una carta da filtro Whatman (attenzione: la materia dei tessuti molli tende ad attaccarsi).
- Posizionare il pezzo di tessuto da 30-40 mg in un omogeneizzatore Elvehjem da 2 mL raffreddato a ghiaccio.
- Aggiungere una quantità appropriata di MiR05 per ottenere 20 mg/mL per mantenere il rapporto tessuto-tampone. Mantenere la quantità totale >1,5 ml e <2 ml per evitare fluidi insufficienti o eccessivi per un'adeguata permeabilizzazione meccanica.
- Inserire il pestello, lisare lentamente il tessuto ritraendo attentamente il pestello evitando la generazione di un vuoto causando un danno tissutale eccessivo.
- Eseguire 7 colpi in totale (1x definito come un tratto verso l'alto e verso il basso) fino a lisato (evidente come un liquido torbido senza detriti maggiori) (Figura 2B).
NOTA: Il numero di ictus per una lisi appropriata deve essere testato per ciascun tessuto valutando l'integrità della membrana mitocondriale esterna tramite la risposta al citocromo C (fase 1.5.11). Il tessuto connettivo difficile da lisare o le parti dei vasi potrebbero rimanere. - Decantare il tessuto lisato in un tubo centrifugo da 15 ml.
- Lavare l'interno del vasaio con una quantità uguale di MiR05 utilizzato nella fase di lisatura (ad esempio, 1,5 mL) e aggiungere al tubo da 15 mL ora contenente 3-4 mL di MiR05 a 10 mg / mL di lisato tissutale.
- Aggiungere 2 mL di MiR05 semplice per camera per riscaldare a 37 °C.
- Ruotare lentamente il tubo per un'equa distribuzione prima di pipettare 500 μL (pari a 5 mg) di ciascun lisato per camera per ridurre al minimo lo stress da freddo a 37 °C.
- Attendere >3 minuti affinché il contenuto della camera si riscaldi a 37 °C prima di chiudere la camera. Rimuovere il liquido in eccesso sulla parte superiore del tappo (quantità per camera dopo la chiusura: 4 mg).
- Eseguire il protocollo SUIT per permeabilizzato standard (passaggio 1.5).
- Asportare un pezzo di tessuto omogeneo, 30-40 mg di peso, o utilizzare l'intero organo (cervelletto di topo in questo caso).
- Preparazione del tessuto fibroso (muscolo scheletrico, muscolo cardiaco) per la respirometria ad alta risoluzione
- Estrarre il tessuto duro, rimuovere il tessuto connettivo e il grasso dai muscoli usando una pinza affilata in 2 ml di BIOPS ghiacciato (Tabella 2) al microscopio di dissezione.
- Separare i fasci di fibre (~ 4 mg) lungo l'asse longitudinale con pinze affilate. Stuzzicare le fibre sufficientemente per ottenere una struttura a rete (Figura 2B).
NOTA: La corretta separazione e permeabilizzazione meccanica delle fibre è indicata dalla perdita della mioglobina del pigmento rosso e dall'aumento della traslucenza. - Lavare e permeabilizzare il fascio di fibre in saponina (50 μg/mL in BIOPS, preparato fresco) per 20 min a 4 °C (le fibre diventano traslucide, indicando la completa permeabilizzazione, Figura 2B).
- Lavare le fibre due volte in MiR05 per 5 minuti per lavaggio a 4 °C.
- Asciugare con carta da filtro e pesare prima di aggiungere alla camera riempita con 2,1 mL MiR05.
- Introdurre i tappi senza chiudersi completamente, quindi ossigenare le camere con 2 mL di O2 puro utilizzando una siringa da 20 mL e chiudere le camere ruotando i tappi con un movimento rotatorio. Mantenere la concentrazione di O2 tra 300-500 μM durante l'esperimento per evitare la limitazione della diffusione dell'ossigeno.
- Protocollo per la valutazione della respirazione di routine nelle cellule o nei tessuti
- Aggiungere il campione alla camera come indicato nei passaggi 1.5.2-1.5.3.
- Aggiungere 2,3 mL di sospensione cellulare MiR05 calda (ingresso standard: 1 x 106 celle/mL come nel passaggio 1.2 o 2 mg di tessuto/mL come nel passaggio 1.3)
- Muscolo scheletrico e cardiaco (fase 1.4): Aggiungere ~ 4 mg di fibre permeabilizzate alla saponina a 2,3 ml di MiR05 caldo preriscaldato considerando i passaggi 1.4.4-1.4.6
- Camere di funzionamento a 37 °C e una velocità di agitazione di 700 giri/min. Attendere >3 minuti per consentire ai fluidi di degassare e chiudere le camere ruotando il tappo con un movimento rotante. La stabilizzazione del blocco di Peltier indica il raggiungimento della temperatura impostata.
- (FACOLTATIVO) Modificare la velocità dell'agitatore a 300 giri / min per consentire alle bolle rimanenti di fuoriuscire attraverso il capillare del tappo.
- Aspirare il liquido in eccesso sulla parte superiore del tappo. Attendere 10 minuti fino a quando non si ottiene un segnale di flusso di ossigeno stabile con qualsiasi tipo di campione per registrare la respirazione di routine / stato 1, Figura 1B).
- Per le misurazioni della respirazione in cellule e tessuti permeabilizzati, continuare con il passaggio 1.6. Per cellule intatte con passo 1.8.
- Protocollo per l'analisi OXPHOS in cellule o tessuti permeabilizzati
- Utilizzare un campione di tessuto lisato (permeabilizzato) o permeabilizzare le cellule aggiungendo 1 μL di digitonina (8,1 mM di stock di digitonina in dimetilsolfossido (DMSO)) per una concentrazione finale di 5 μg/mL per permeabilizzare le cellule. Il flusso diminuirà e dovrebbe stabilizzarsi a >5 min.
ATTENZIONE: La digitonina è acutamente tossica per le vie respiratorie, a contatto con la pelle o quando viene ingerita.
NOTA: l'iniezione di tutte le sostanze chimiche viene eseguita con siringhe di vetro di precisione. Utilizzare siringhe solo per sostanze chimiche indicate per evitare contaminazioni incrociate e lavare accuratamente in acqua ed EtOH dopo l'uso. Le siringhe bloccate possono richiedere l'ultrasonicazione in ddHcaldo 2O o un filo di pulizia per rimuovere eventuali zoccoli chimici. Ritrarre sempre un surplus della rispettiva soluzione madre nella siringa per evitare di introdurre aria nelle camere. Ispezionare l'interno delle camere per l'introduzione di aria dopo ogni iniezione. Registra ogni passo fino a quando non si stabilizzano il flusso. - Aggiungere in rapida successione: 5 μL di 0,4 M di malato (M) per una concentrazione finale di 1 mM, 5 μL di 2,0 M piruvato (P; preparato fresco), per una concentrazione finale di 5 mM, 4 μL di 2,5 M di glutammato (G) per una concentrazione finale di 5 mM.
- Dopo che il flusso precedente si è stabilizzato, aggiungere 5 μL (10 μL per il tessuto muscolare) di 0,5 M di adenosina difosfato (ADP, aliquote immagazzinate a -80 °C) per una concentrazione finale di 1,25 mM.
NOTA: tessuto come il muscolo potrebbe aver bisogno di una concentrazione diversa per raggiungere la saturazione. - Aggiungere 5 μL di 4 mM di citocromo C (cytC) per una concentrazione finale di 10 μM.
NOTA: Facoltativo per le cellule per valutare la qualità della permeabilizzazione. - Aggiungere 16 μL di 1,25 M di succinato (S) per una concentrazione finale di 10 mM. (FACOLTATIVO) Aggiungere 3 μL di 0,5 M ADP per una concentrazione finale di 2 mM per controllare la saturazione della concentrazione di ADP.
- Per le cellule e i tessuti non fibrosi, aggiungere 2 μL di 1 mg/mL di oligomicina (OM) per una concentrazione finale di 1 μg/mL.
ATTENZIONE: Tutti gli inibitori ETS utilizzati sono altamente tossici.
NOTA: l'oligomicina può richiedere la titolazione per una concentrazione ottimale in quanto può reprimere la capacità ETS e viene omessa per il tessuto muscolare. Riossigenare qui quando il tessuto muscolare viene dosato e se O2 è inferiore a 300 μM. - Titolare FCCP da uno stock di 2 mM, aggiungere 0,6 μL con successivi passi di 0,2 μL fino a quando nessun aumento della respirazione e della respirazione è massimamente disaccoppiato (teorico: non accoppiato).
- Aggiungere 1 μL di 1 mM di rotenone (ROT) per una concentrazione finale di 0,5 μM. Aggiungere 2 μL di 1 mg/mL di antimicina (AM) per una concentrazione finale di 1 μg/mL.
- Riossigenare le camere per ottenere un livello di ossigeno simile (~150 μM) in tutte le camere sollevando lentamente lo stantuffo in movimento di torsione.
- Aggiungere 5 μL di 0,8 M di ascorbato per una concentrazione finale di 2 mM immediatamente seguita da 5 μL di 0,2 M N,N,N′,N′-tetrametil-p-fenilendiammina (TMPD) per una concentrazione finale di 0,5 mM per valutare l'attività IV complessa (opzionale).
- Aggiungere 5 μL di 4 M azide per una concentrazione finale di 10 mM immediatamente quando viene raggiunto il picco di flusso O2 con TMPD. Continuare la corsa per >5 minuti per analizzare l'auto-ossidazione di TMPD per il calcolo del livello di base IV complesso.
- Riconteggiare le celle per confermare il conteggio delle celle prima dell'esecuzione e continuare con il passaggio 1.9.
NOTA: La digitonizzazione (solo per le cellule) deve essere titolata negli esperimenti di prova per raggiungere il flusso massimo e non influenzare l'integrità della membrana mitocondriale (vedere passo 1.7). I campioni permeabilizzati (in particolare il tessuto muscolare) con >10% di aumento della frequenza respiratoria dopo l'aggiunta di citocromo c devono essere esclusi da ulteriori analisi a causa di danni alla membrana mitocondriale esterna. Si prevede un breve calo del flusso dopo l'aggiunta di sostanze chimiche disciolte dall'EtOH.
- Utilizzare un campione di tessuto lisato (permeabilizzato) o permeabilizzare le cellule aggiungendo 1 μL di digitonina (8,1 mM di stock di digitonina in dimetilsolfossido (DMSO)) per una concentrazione finale di 5 μg/mL per permeabilizzare le cellule. Il flusso diminuirà e dovrebbe stabilizzarsi a >5 min.
- Protocollo per determinare le condizioni ottimali di permeabilizzazione per le cellule
- Aggiungere celle come descritto nei passaggi 1.2 e 1.5.2.
- Prendere 10 μL di 10 mg/mL di digitonina e aggiungere 10 μL di DMSO per diluire a 5 mg/mL.
- Aggiungere 1 μL di rotenone (1 mM di stock). Aggiungere 10 μL di succinato (2 mM di stock) e 5 μL di ADP (0,5 M di stock).
- Titolare 1 μL di digitonina (2,5 mg per passo) ripetutamente fino a quando la respirazione non aumenta ulteriormente ed è massima.
NOTA: Una diminuzione della respirazione indica un'eccessiva concentrazione di digitonina.
- Protocollo per l'analisi oxphos in cellule intatte
- Dopo la respirazione di routine (fase 1.6.1-1.6.6), aggiungere 2 μL di 0,01 mM di oligomicina per una concentrazione finale di 10 nM.
- Titolare FCCP da 2 mM stock, aggiungere 0,6 μL con successivi passi di 0,2 μL fino a quando nessun ulteriore aumento della respirazione e della respirazione è massimamente disaccoppiato (teorico: non accoppiato)
- Aggiungere 1 μL di 1 mM di rotenone per una concentrazione finale di 0,5 μM. Aggiungere 2 μL di 1 mg/mL di antimicina per una concentrazione finale di 1 μg/mL.
- Riossigenare la camera allo stesso livello di ossigeno (~150 μM) sollevando lentamente lo stantuffo in movimento di torsione.
- Aggiungere 5 μL di 0,8 M di ascorbato per una concentrazione finale di 2 mM. Aggiungere immediatamente 5 μL di 0,2 M TMPD per una concentrazione finale di 0,5 mM per valutare l'attività IV complessa.
NOTA: Preparare un nuovo lotto prima di qualsiasi serie più grande di esperimenti poiché il TMPD è soggetto all'auto-ossidazione. L'attività potrebbe diminuire nel tempo se conservata a -20 °C. - Aggiungere 5 μL di 4 M azide per una concentrazione finale di 10 mM immediatamente quando viene raggiunto il picco di flusso O2 con TMPD. Continuare a correre per >5 minuti per analizzare l'auto-ossidazione di TMPD per il calcolo del livello di base IV complesso.
- Riconteggi le celle per confermare il conteggio delle celle pre-eseguito e continuare con il passaggio 1.9.
- Raccolta di esempi post-esecuzione
- Raccogliere esattamente 1 mL di sospensione MiR05 da ciascuna camera (con agitatori accesi) in un tubo da 1,5 mL da 1,5 mL.
- Centrifuga a 1000 x g per cellule permeabilizzate o a 20.000 x g per lisato tissutale. Rimuovere il surnatante e congelare il pellet a -80 °C per un'ulteriore lavorazione (sezione 3).
- Analisi dei protocolli SUIT
- Analizzare il flusso di ossigeno (pmol/s, normalizzato in ingresso) ad ogni plateau dopo aver aggiunto un substrato o un inibitore (Figura 1C e Figura 3A). Esportare i valori in un foglio di calcolo.
- Sottrarre il valore del consumo residuo di ossigeno (ROX, Figura 1C e Figura 3C) da tutti i valori di ogni esecuzione sperimentale. Sottrarre la respirazione residua di azide dal TMPD per ottenere la respirazione IV complessa.
- Tracciare i valori assoluti normalizzati per l'input cellulare (Figura 3A, B) o tissutale (Figura 5A,B). Calcolare i rapporti di controllo del flusso (fase 1.11) o normalizzarli in input proteico (Figura 3C).
- Calcolo del rapporto di controllo del flusso
- Acquisire un indice della funzione respiratoria e del controllo dell'accoppiamento utilizzando i rapporti di controllo del flusso (FCR)9,26.
NOTA: Questo permette di valutare la qualità mitocondriale intrinseca, indipendentemente dalla quantità mitocondriale. Inoltre, i rapporti di controllo del flusso (FCR) sono comparabili all'interno delle stesse linee cellulari consentendo il controllo della qualità dei reagenti (i rispettivi FCR sono ottenuti attraverso i valori di riferimento numerati indicati nella Figura 1B-D e nella Figura 3C). - Calcolare il rapporto di controllo respiratorio per l'accoppiamento di OXPHOS a LEAK utilizzando l'equazione 1.
Equazione 1: FCRADP = 5/6 = Stato 3 / Stato 4 - Calcola la FCR per valutare la respirazione NADH-dipendente usando l'equazione 2
Equazione 2: Stato FCR3 (I) = 3/5 = Stato 3 (I) / Stato 3 (I+II) - Calcola la FCR per valutare la respirazione succinato-dipendente usando l'equazione 3.
Equazione 3: stato FCR3 (II) = 8/7 =S rot /capacità ETS - Calcola l'FCR per valutare accoppiato a disaccoppiato usando l'equazione 4.
Equazione 4: FCRaccoppiato/disaccoppiato = 5/7 = Stato 3 (I+II) /Capacità ETS - Per testare l'integrità della membrana esterna mitocondriale, utilizzare l'equazione 5.
Equazione 5: % danno mitocondriale della membrana esterna = 3/4 = Stato 3 (I) / Stato 3 (I) con cyt c
- Acquisire un indice della funzione respiratoria e del controllo dell'accoppiamento utilizzando i rapporti di controllo del flusso (FCR)9,26.
2. Respirometria ad alta risoluzione: respirometro a base di micropiastre (mHRR)
NOTA: Gli esperimenti in questa sezione del protocollo sono stati eseguiti utilizzando l'analizzatore di flusso extracellulare Seahorse XFe96 (Tabella dei materiali)
- Coltura cellulare
- Coltivare qualsiasi tipo di cellula. Gli aderenti (ad esempio, collagene, laminina) potrebbero essere utilizzati per facilitare l'attaccamento cellulare. Qui, le cellule HEK293 vengono coltivate come prima (passaggio 1.3).
- Il giorno prima dell'esperimento, staccare le cellule e trasferirle in una micropiastra mHRR a 96 pozzetti designata per ottenere la confluenza ideale il giorno dell'esperimento (80%-100%) (Figura 2C).
NOTA: per mHRR, le densità delle celle micropiastre sono fondamentali. Le proprietà di crescita individuali delle linee cellulari o dei trattamenti che influenzano la crescita devono essere prese in considerazione per garantire una confluenza comparabile il giorno dell'esperimento.
- Preparazione di celle per respirometria ad alta risoluzione
- Raccogliere e risospese le cellule sufficientemente prima della semina
NOTA: si consiglia di seminare cellule dalla stessa diluizione per le repliche. - Seminare le cellule in base ai tassi di crescita delle singole linee cellulari o alle proprietà di crescita in trattamento.
NOTA: ottimizzare su una micropiastra standard a 96 pozzetti ed estrapolare la densità della cella in una micropiastra specifica del saggio a 96 pozzetti. In questa configurazione, 7 x 104 celle WT HEK293 sono state seminate per pozzetto di un pozzo 96. La prima e l'ultima colonna della piastra a 96 pozzetti sono utilizzate per la determinazione delle proteine (Figura 2C). I quattro pozzetti angolari non devono contenere cellule e vengono utilizzati per la correzione sperimentale dello sfondo. Idealmente, i pozzetti vicini ai bordi sono vuoti per ridurre al minimo l'effetto bordo (ad esempio, le celle mostrano tassi di crescita alterati causati dagli effetti della temperatura) (Figura 2C, D).
- Raccogliere e risospese le cellule sufficientemente prima della semina
- Preparazione di piastre sensore, caricamento di inibitori
- Il giorno del test, integrare 38,8 mL di mezzo con 0,4 mL di 1 M di glucosio, 0,4 mL di 200 mM di glutammina e 0,4 mL di 100 mM Na-piruvato.
NOTA: la respirazione mHRR richiede un mezzo DMEM specializzato non tamponato a pH 7,4. In generale, 40 ml dovrebbero essere sufficienti per un esperimento con una micropiastra a 96 pozzetti. - Riscaldare il mezzo di saggio respiratorio a 37 °C e scambiare il terreno di coltura cellulare con il mezzo di saggio respiratorio lavando due volte con 80 μL per pozzetto.
- Posizionare la piastra con le celle in un incubatore a 37 °C senza CO2 per 60 minuti prima del test.
NOTA: Questo passaggio è essenziale per degassare la piastra poiché la CO2 può influenzare i risultati della respirazione e il siero nel mezzo può produrre bolle durante il test. - Aliquote di inibitori pre-riscaldamento per OM, FCCP, ROT e AM a 37 °C e tolgono la piastra del sensore dall'incubatore.
- Diluire OM, FCCP, ROT e AM in 3 mL di mezzo di saggio a una concentrazione finale del pozzo di 1,5 μM, 1,125 μM e 1 μM, rispettivamente. Compilare porte separate come indicato nella Figura 2E.
NOTA: si consiglia una pipetta multicanale per riempire la cartuccia del sensore. Poiché l'aria pressurizzata viene utilizzata per iniettare composti, tutte le porte devono essere riempite con una quantità uguale di volume liquido ogni volta che una porta viene riempita con un composto. ROT e AM possono essere combinati in un'unica porta. Gli inibitori possono essere sciolti in EtOH o DMSO. - Ispezionare le porte di iniezione e verificare un volume di caricamento uniforme per ciascuna porta.
NOTA: tutte le porte contengono un foro nella parte inferiore per l'iniezione. Prestare attenzione quando si sposta la piastra del sensore. Le bolle d'aria possono essere rimosse usando un ago.
- Il giorno del test, integrare 38,8 mL di mezzo con 0,4 mL di 1 M di glucosio, 0,4 mL di 200 mM di glutammina e 0,4 mL di 100 mM Na-piruvato.
- Protocollo per la valutazione dell'ossigeno in cellule intatte
- Il giorno prima del test, eseguire i passaggi 2.4.2-2.4.7.
- Aliquota 20 mL della soluzione calibrante in un tubo conico da 50 mL.
- Aprire il kit di analisi del flusso extracellulare e rimuovere il contenuto.
- Posizionare la cartuccia del sensore invertita accanto alla piastra di utilità. Pipettare 200 μL di soluzione calibrante in ciascun pozzetto della piastra di utilità.
- Collegare la cartuccia del sensore alla piastra di utilità facendo attenzione che tutti i sensori siano sommersi.
- Posizionare la piastra in un incubatore a 37 °C senza CO2 durante la notte o un minimo di 12 ore. Verificare che l'umidità all'interno dell'incubatore sia sufficiente per evitare l'evaporazione del calibrante.
- Accendere il sistema basato su micropiastre e il computer per essere pronti all'uso il giorno successivo (la macchina richiede un minimo di 3 ore per equilibrarsi a 37 °C prima di eseguire un test).
NOTA: per la stabilità del segnale, aumentare i punti di misurazione a 6 anziché 3 cicli di misurazione per stato respiratorio. Ogni ciclo consiste in 3 min di miscelazione e 3 min di misurazione. - Il giorno del test XF, eseguire i passaggi 2.4.9-2.4.20.
- Verificare la confluenza della piastra di coltura cellulare, la morfologia delle cellule e che i pozzetti di fondo siano vuoti.
- Lavare le cellule con il mezzo respiratorio preparato come indicato nei passaggi 2.4.11-2.4.12.
- Rimuovere tutti tranne 20 μL del terreno di coltura da ciascun pozzetto. Rimuovere 55 μL se il terreno di coltura era di 80 μL a causa dell'evaporazione durante la notte (circa 5 μL).
- Lavare le cellule due volte con 90 μL di mezzo di saggio. Infine, aggiungere 100 μL di mezzo di saggio. Il volume finale dovrebbe essere di 120 μL.
NOTA: per questa fase si consiglia una pipetta multicanale per garantire che la stessa procedura di lavaggio sia stata applicata a ciascuna condizione sperimentale (dipende dalla configurazione della piastra). Durante l'aspirazione, inclinare la piastra con un angolo di 45 ° e posizionare le punte della pipetta nell'angolo dei pozzetti per l'aspirazione e l'iniezione di liquidi. È imperativo fare attenzione durante il lavaggio poiché alcune cellule possono facilmente staccarsi dal fondo della piastra di coltura cellulare. - Posizionare la piastra in un incubatore a 37 °C senza CO2 per 60 minuti prima del test.
- Recuperare la piastra della cartuccia del sensore idratata dall'incubatore privo di CO2.
- Scartare la vecchia soluzione di calibrante e sostituirla con una soluzione di calibrante fresco, preriscaldata a 37 °C.
- Preparare gli inibitori e il mezzo di saggio (3 mL per inibitore per un totale di 12 mL di mezzo di saggio) e utilizzare un serbatoio di pipetta per il caricamento dell'inibitore nelle porte.
- Aprire il software ed eseguire un modello predefinito o nuovo. Riempire la mappa della piastra, regolare le titolazioni e i cicli di misurazione, quindi premere Start per avviare la calibrazione dei sensori ottici.
- Rimuovere il coperchio dalla cartuccia caricata e posizionarlo nello slot che scorre automaticamente fuori dalla macchina, verificando che i segni nell'angolo in basso a destra della piastra si allineino con il triangolo nell'angolo in basso a destra dello slot.
- Fare clic su Continua per eseguire la calibrazione automatica, della durata di circa 20 minuti.
- Dopo la calibrazione, rimuovere la piastra di utilità contenente il calibrante.
- Rimuovere il coperchio dalla micropiastra contenente le celle e posizionare la piastra nella fessura quando richiesto dalla macchina. Fare clic su Continua per avviare la corsa.
- Raccolta di esempi post-esecuzione
- Estrarre la piastra dalla macchina, rimuovere con attenzione il mezzo di saggio rimanente senza disturbare le celle e congelare l'intera piastra a -80 °C per un'ulteriore lavorazione (sezione 3).
3. Determinazione delle proteine mediante il saggio dell'acido bicinchoninico (BCA assay)
- Preparare l'albumina sierica bovina diluita (BSA) in un tampone utilizzato per l'estrazione delle proteine e compatibile con BCA: 2 mg/mL, 1,5 mg/mL, 1 mg/mL, 0,5 mg/mL, 0,25 mg/mL e 0 mg/mL per la curva standard nei duplicati.
- Estrarre le proteine riesempendo in un appropriato tampone di lisi (ad esempio, RIPA) con 20 μL per pozzetto per mHRR o 100 μL per pellet contenuto all'interno di un tubo da 1,5 mL per cHRR.
- Incubare la piastra mHRR o il tubo da 1,5 ml contenente lisati proteici per 30 minuti sul ghiaccio.
- Centrifugare il tubo da 1,5 mL contenente il lisato proteico a 4 °C a 20.000 x g per 20 minuti e trasferire il surnatante risultante in un nuovo tubo pulito da 1,5 ml.
- Utilizzare 10 μL per campione in duplicati e standard in una piastra di microtitolazione. Aggiungere 200 μL di reagente funzionante BCA e incubare per >15 min.
- Leggere la piastra in uno spettrofotometro standard a una lunghezza d'onda di 562 nm e calcolare le concentrazioni proteiche utilizzando una curva standard BSA.
- Normalizzare i risultati della respirazione alla concentrazione proteica.
NOTA: la normalizzazione alla quantità proteica consente di corroborare le densità di semina cellulare o l'input di peso umido. Le proteine estratte sono adatte per il successivo immunoblotting contro subunità dell'ETS, ad esempio, ma non rappresentano pienamente il campione nativo (ad esempio, perdita di siti di fosforilazione).
Risultati
Qui, forniamo protocolli per determinare la bioenergetica mitocondriale nelle cellule eucariotiche, nel tessuto non fibroso (ad esempio, cervelletto) e nel tessuto fibroso (ad esempio, muscolo scheletrico). Per le cellule eucariotiche, HEK293 con knockout ingegnerizzato CRISPR di due diverse proteine associate alla traduzione mitocondriale con conseguente deficit multiplo (CRISPRKO1) e grave/completo di OXPHOS (CRISPRKO2) è stato misurato con cHRR (Figura 3A-C<...
Discussione
Tradizionalmente, la bioenergetica mitocondriale è stata studiata con elettrodi di ossigeno di tipo Clark. Una mancanza di risoluzione e throughput, tuttavia, giustificata per il progresso tecnologico. Ad oggi, l'O2k (indicato come cHRR) e il Seahorse XF96 Flux Analyzer (denominato mHRR) sono stati ampiamente adottati nel campo della bioenergetica cellulare. Qui, presentiamo una raccolta comprensibile di protocolli per l'analisi del metabolismo energetico cellulare attraverso la valutazione della respirazione mitocondri...
Divulgazioni
Nessun conflitto di interessi da divulgare.
Riconoscimenti
Questo lavoro è stato sostenuto da finanziamenti dell'Accademia di Finlandia (C.B.J),della Magnus Ehrnroot Foundation (C.B.J) e da una borsa di dottorato della Integrated Life Sciences Graduate School (R.A.).
Materiali
| Name | Company | Catalog Number | Comments |
| 2 mL Potter-Elvehjem Glass/PTFE Tissue Grinder/Homogenizer | Omni International | 07-358029 | |
| 95% O2, 5% CO2 medical gas mixture | Potter for tissue grinding | ||
| ADP | Sigma | A 4386 | |
| Antimycin A | Sigma | A 8674 | Chemical |
| Ascorbate | Merck | PHR1279-1G | Chemical, dissolve in ethanol |
| BSA (fatty accid free) | Sigma | A 6003 | Chemical |
| CaCO3 | Sigma | C 4830 | Chemical |
| Cytochrome c | Sigma | C 7752 | Chemical |
| Digitonin | Sigma | D 5628 | Chemical |
| Dithiothreitol | Sigma | D 0632 | Chemical, dissolve in DMSO |
| D-Sucrose | Roth | 4621.1 | Chemical |
| Dulbecco’s modified Eagle’s medium (High glucose) | Fisher Scientific | 41965-039 | Chemical |
| Dulbecco’s modified Eagle’s medium (No Glucose) | Fisher Scientific | A14430-01 | |
| EGTA | Sigma | E 4378 | |
| Etomoxir | Sigma | E1905 | Chemical |
| Falcon 15 ml Conical Centrifuge Tubes | Fisher Scientific | AM12500 | Chemical |
| Falcon 50 ml Conical Centrifuge Tubes | Fisher Scientific | AM12501 | |
| FCCP | Sigma | C 2920 | |
| Glucose | Sigma | G7021 | Chemical, dissolve in ethanol |
| Glutamate | Sigma | G 1626 | Chemical |
| GlutaMax (100x) (200 nM L-alanyl-L-glutamine dipeptide) | Fisher Scientific | 35050061 | Chemical |
| HEK293 cells | ATTC | CRL-1573 | |
| Hemocytometer | Fisher Scientific | 0267151B | Instrument for cell counting |
| Hepes | Sigma | H 7523 | Chemical |
| Imidazole | Fluka | 56750 | Chemical |
| KCl | Merck | 1.04936 | Chemical |
| L-carnitine | Sigma | C0283 | Chemical |
| Malate | Sigma | M 1000 | Chemical |
| MES hydrate | Sigma | M8250 | Chemical |
| MgCl2 | Sigma | M 9272 | Chemical |
| Na2ATP | Sigma | A 2383 | Chemical |
| Na2Phosphocreatine | Sigma | P 7936 | Chemical |
| Na-pyruvate (100 mM) (100x) | Fisher Scientific | 11360070 | |
| NEAA (Non-essential amino acids) 100x | Fisher Scientific | 11140035 | |
| Normal FBS (10x) | Fisher Scientific | 10500064 | |
| O2k-Core: Oxygraph-2k | Oroboros Instruments | 10000-02 | High-resolution respirometry instrument |
| O2k-Titration Set | Oroboros Instruments | 20820-03 | Hamilton syringes for chemical injections |
| Oligomycin | Sigma | O 4876 | Chemical, dissolve in ethanol |
| Palmitoylcarnitine | Sigma | P 4509 | Chemical |
| Penicillin-Streptomycin | Fisher Scientific | 15140122 | |
| Pierce BCA Protein Assay Kit | Fisher Scientific | 23227 | |
| Pyruvate | Sigma | P 2256 | Chemical |
| RIPA-Buffer | Fisher Scientific | 89900 | Chemical |
| Rotenone | Sigma | R 8875 | Chemical, dissolve in ethanol |
| Saponin | Sigma | S7900 | Chemical |
Seahorse XF DMEM assay medium pack, pH 7.4 | Agilent, Santa Clara, CA | 103680-100 | |
| Seahorse XFe96 Extracellular Flux Analyzer | Agilent, Santa Clara, CA | High-throughput respirometry instrument | |
| Seahorse XFe96 FluxPak | Agilent, Santa Clara, CA | Includes assay plates, cartridges, loading guides for transferring compounds to the assay cartridge, and calibrant solution. | |
| Small scissors | Fisher Scientific | 08-951-20 | |
| Sodium azide | Sigma | S2002 | Chemical |
| Succinate | Sigma | S 2378 | Chemical |
| Taurine | Sigma | T 8691 | Chemical |
| TMPD | Sigma | T 3134 | Chemical |
| Trypan Blue solution | Merck | 72-57-1 | Chemical |
| Trypsin 0.25% EDTA | Fisher Scientific | 25200056 | |
| Two thin-edged forceps | Fisher Scientific | 12-000-122 | |
| Uridine stock (500x) | Sigma | U3750 | Chemical |
Riferimenti
- McBride, H. M., Neuspiel, M., Wasiak, S. Mitochondria: More than just a powerhouse. Current Biology. 16 (14), 551-560 (2006).
- Mehta, M. M., Weinberg, S. E., Chandel, N. S. Mitochondrial control of immunity. Beyond ATP. Nature Reviews Immunology. 17 (10), 608-620 (2017).
- Spinelli, J. B., Haigis, M. C. The multifaceted contributions of mitochondria to cellular metabolism. Nature Cell Biology. 20 (7), 745-754 (2018).
- Gnaiger, E. Capacity of oxidative phosphorylation in human skeletal muscle. New perspectives of mitochondrial physiology. International Journal of Biochemistry and Cell Biology. 41 (10), 1837-1845 (2009).
- Gorman, G. S., et al. Mitochondrial diseases. Nature Reviews Disease Primers. 2, 1-23 (2016).
- Boushel, R., Gnaiger, E., Schjerling, P., Skovbro, M., Kraunsøe, R., Dela, F. Patients with type 2 diabetes have normal mitochondrial function in skeletal muscle. Diabetologia. 50 (4), 790-796 (2007).
- Cogliati, S., et al. Mitochondrial cristae shape determines respiratory chain supercomplexes assembly and respiratory efficiency. Cell. 155 (1), 160-171 (2013).
- Kühlbrandt, W. Structure and function of mitochondrial membrane protein complexes. BMC Biology. 13, 1-11 (2015).
- Gnaiger, E. Mitochondrial pathways and Respiratory control. An introduction to OXPHOS analysis. Bioenergetics communications. 5th ed. , (2020).
- Jackson, C. B., et al. Mutations in SDHD lead to autosomal recessive encephalomyopathy and isolated mitochondrial complex II deficiency. Journal of Medical Genetics. 51 (3), 170-175 (2014).
- Pesta, D., Gnaiger, E. High-resolution respirometry: OXPHOS protocols for human cells and permeabilized fibers from small biopsies of human muscle. Methods in Molecular Biology. 810, 25-58 (2012).
- Horan, M. P., Pichaud, N., Ballard, J. W. O. Review: Quantifying mitochondrial dysfunction in complex diseases of aging. Journals of Gerontology - Series A Biological Sciences and Medical Sciences. 67 (10), 1022-1035 (2012).
- Doerrier, C., Garcia-Souza, L. F., Krumschnabel, G., Wohlfarter, Y., Mészáros, A. T., Gnaiger, E. High-resolution fluorespirometry and oxphos protocols for human cells, permeabilized fibers from small biopsies of muscle, and isolated mitochondria. Methods in Molecular Biology. 1782, 31-70 (2018).
- Zhang, J., et al. Measuring energy metabolism in cultured cells, including human pluripotent stem cells and differentiated cells. Nature Protocols. 7 (6), 1068-1085 (2012).
- García-Roche, M., Casal, A., Carriquiry, M., Radi, R., Quijano, C., Cassina, A. Respiratory analysis of coupled mitochondria in cryopreserved liver biopsies. Redox Biology. 17, 207-212 (2018).
- Acin-Perez, R., et al. A novel approach to measure mitochondrial respiration in frozen biological samples. The EMBO Journal. 39 (13), 1-18 (2020).
- Cell metabolism assay kits. Seahorse assay kits and media Available from: https://www.agilent.com/en/product/cell-analysis/real-time-cell-metabolic-analysis/xf-assay-lits-reagents-cell-assay-media (2021)
- Chance, B., Williams, G. R. A method for the localization of sites for oxidative phosphorylation. Nature. 176 (4475), 250-254 (1955).
- Gnaiger, E., et al. Mitochondrial respiratory states and rates. MitoFit Preprint Arch. , (2019).
- Gnaiger, E. O2k-procedures: SOP O2k quality control 1: Polarographic oxygen sensors and accuracy of calibration Section Page. Oroboros. 03 (18), 1-21 (2020).
- Robinson, B. H., Petrova-Benedict, R., Buncic, J. R., Wallace, D. C. Nonviability of cells with oxidative defects in galactose medium: A screening test for affected patient fibroblasts. Biochemical Medicine and Metabolic Biology. 48 (2), 122-126 (1992).
- King, M. P., Attardi, G. Human cells lacking mtDNA: Repopulation with exogenous mitochondria by complementation. Science. 246 (4929), 500-503 (1989).
- Makrecka-Kuka, M., Krumschnabel, G., Gnaiger, E. High-resolution respirometry for simultaneous measurement of oxygen and hydrogen peroxide fluxes in permeabilized cells, tissue homogenate and isolated mitochondria. Biomolecules. 5 (3), 1319-1338 (2015).
- Fasching, M., Gnaiger, E. O2k quality control 2: Instrumental oxygen background correction and accuracy of oxygen flux. Mitochondrial Physiology Network. 14 (06), 1-14 (2016).
- Gnaiger, E., Lassnig, B., Kuznetsov, A., Rieger, G., Margreiter, R. Excess capacity of cytochrome c oxidase. Journal of Experimental Biology. 1139, 1129-1139 (1998).
- Gnaiger, E., et al. Mitochondria in the Cold. Life in the Cold. , 431-442 (2000).
- Fontana-Ayoub, M., Fasching, E., Gnaiger, Selected media and chemicals for respirometry with mitochondrial preparations. Mitochondrial Physiology Network. 02 (17), 1-9 (2014).
- Gerencser, A. A., et al. Quantitative microplate-based respirometry with correction for oxygen diffusion. Analytical Chemistry. 81 (16), 6868-6878 (2009).
- Krumschnabel, G., Eigentler, A., Fasching, M., Gnaiger, E. Use of safranin for the assessment of mitochondrial membrane potential by high-resolution respirometry and fluorometry. Methods in Enzymology. 542, 163-181 (2014).
- Nászai, A., Terhes, E., Kaszaki, J., Boros, M., Juhász, L. Ca(2+)N it be measured? Detection of extramitochondrial calcium movement with high-resolution fluorespirometry. Scientific Reports. 9 (1), 1-13 (2019).
- Pajak, B., et al. 2-Deoxy-d-Glucose and its analogs: From diagnostic to therapeutic agents. International Journal of Molecular Sciences. 21 (1), 234 (2019).
- Mercier-Letondal, P., Marton, C., Godet, Y., Galaine, J. Validation of a method evaluating T cell metabolic potential in compliance with ICH Q2 (R1). Journal of Translational Medicine. 19 (1), 1-15 (2021).
- Sauerbeck, A., et al. Analysis of regional brain mitochondrial bioenergetics and susceptibility to mitochondrial inhibition utilizing a microplate based system. Journal of Neuroscience Methods. 198 (1), 36-43 (2011).
- Jackman, M. R., Willis, W. T. Characteristics of mitochondria isolated from type I and type IIb skeletal muscle. American Journal of Physiology - Cell Physiology. 270 (2), 673-678 (1996).
- Ponsot, E., et al. Mitochondrial tissue specificity of substrates utilization in rat cardiac and skeletal muscles. Journal of Cellular Physiology. 203 (3), 479-486 (2005).
- Schönfeld, P., Reiser, G. Why does brain metabolism not favor burning of fatty acids to provide energy-Reflections on disadvantages of the use of free fatty acids as fuel for brain. Journal of Cerebral Blood Flow and Metabolism. 33 (10), 1493-1499 (2013).
- Calderon-Dominguez, M., Mir, J. F., Fucho, R., Weber, M., Serra, D., Herrero, L. Fatty acid metabolism and the basis of brown adipose tissue function. Adipocyte. 5 (2), 98-118 (2016).
- Divakaruni, A. S., Rogers, G. W., Murphy, A. N. Measuring mitochondrial function in permeabilized cells using the seahorse XF analyzer or a clark-type oxygen electrode. Current Protocols in Toxicology. 2014, 1-16 (2014).
- Iuso, A., Repp, B., Biagosch, C., Terrile, C., Prokisch, H. Assessing mitochondrial bioenergetics in isolated mitochondria from various mouse tissues using Seahorse XF96 analyzer. Methods in Molecular Biology. 1567, 217-230 (2017).
- Rogers, G. W., et al. High throughput microplate respiratory measurements using minimal quantities of isolated mitochondria. PLoS ONE. 6 (7), 21746 (2011).
- Jordá, A., Zaragozá, R., Portolés, M., Báguena-Cervellera, R., Renau-Piqueras, J. Long-term high-protein diet induces biochemical and ultrastructural changes in rat liver mitochondria. Archives of Biochemistry and Biophysics. 265 (2), 241-248 (1988).
- Jackson, C. B., Gallati, S., Schaller, A. QPCR-based mitochondrial DNA quantification: Influence of template DNA fragmentation on accuracy. Biochemical and Biophysical Research Communications. 423 (3), 441-447 (2012).
- Hirsch, H. M. Tissue autoxidation inhibitors: II. The presence of inhibitor in intact cells; Assay of liver and hepatoma effect on radio-oxidations. Cancer Research. 16 (11), 1076-1082 (1956).
- Picard, M., et al. Mitochondrial structure and function are disrupted by standard Isolation methods. PLoS ONE. 6 (3), 18317 (2011).
- Tanumihardja, E., Slaats, R. H., Van Der Meer, A. D., Passier, R., Olthuis, W., Van Den Berg, A. Measuring both pH and O2 with a single On-Chip sensor in cultures of human pluripotent stem cell-derived cardiomyocytes to track induced changes in cellular metabolism. ACS Sensors. 6 (1), 267-274 (2021).
- Harms, F., Stolker, R. J., Mik, E. Cutaneous respirometry as novel technique to monitor mitochondrial function: A feasibility study in healthy volunteers. PLoS ONE. 11 (7), 159544 (2016).
- Levitsky, Y., et al. Micro-respirometry of whole cells and isolated mitochondria. RSC Advances. 9 (57), 33257-33267 (2019).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati