
É necessária uma assinatura da JoVE para visualizar este conteúdo. Faça login ou comece sua avaliação gratuita.
Method Article
Respirometria de alta resolução para avaliar bioenergésicos em células e tecidos usando respirômetros baseados em câmara e placas
Neste Artigo
Resumo
Avaliar a fosforilação oxidativa usando respirômetros de alta resolução tornou-se parte integrante da análise funcional das mitocôndrias e do metabolismo de energia celular. Aqui, apresentamos protocolos para a análise do metabolismo de energia celular usando respirômetros de alta resolução baseados em câmara e microplatos e discutimos os principais benefícios de cada dispositivo.
Resumo
A respirometria de alta resolução (HRR) permite monitorar a fosforilação oxidativa em tempo real para análise de estados de energia celular individuais e avaliação de complexos respiratórios usando protocolos diversificados de titulação inibidora de substrato-uncoupler (SUIT). Aqui, demonstra-se o uso de dois dispositivos de respirometria de alta resolução, e uma coleção básica de protocolos aplicáveis para a análise de células cultivadas, fibras musculares esqueléticas e cardíacas, e tecidos moles como o cérebro e o fígado são apresentados. Protocolos para células e tecidos cultivados são fornecidos para um respirômetro baseado em câmara e células cultivadas para um respirômetro à base de microplato, ambos abrangendo protocolos de respiração padrão. Para fins comparativos, as células HEK293 projetadas pelo CRISPR deficientes na tradução mitocondrial que causam deficiência múltipla do sistema respiratório são usadas com ambos os dispositivos para demonstrar defeitos celulares na respiração. Ambos os respirômetros permitem uma medição abrangente da respiração celular com seus respectivos méritos técnicos e adequação dependente da questão da pesquisa e do modelo em estudo.
Introdução
Mitocôndrias cumprem a oferta-chave de energia e são uma organela compartimentalizada contribuindo para processos bioenergetics celulares essenciais e metabólicos, como anabolismo de nucleotídeos, lipídios e aminoácidos, biogênese do aglomerado de enxofre de ferro e estão implicados em sinalização como morte celular controlada 1,2,3 . A bioenergeticia mitocondrial através da fosforilação oxidativa contribui para quase todos os processos celulares dentro da célula e, consequentemente, disfunções mitocondriais de origem primária ou secundária estão associadas a um amplo espectro de condições da doença 4,5. A disfunção mitocondrial envolve não apenas alterações na estrutura ou densidade mitocondrial, mas também na qualidade e regulação do sistema respiratório6. Este elemento qualitativo abrange controle de substrato, características de acoplamento, modificações pós-translacionais, dinâmica cristae e supercomplexos respiratórios 7,8. Portanto, a análise precisa da bioenergetics mitocondrial para abordagens experimentais e diagnósticas para avaliar o metabolismo energético da célula é importante na saúde e na doença.
Fosforilação oxidativa mitocondrial (OXPHOS) é uma sequência de reações dentro do sistema respiratório ou sistema de transferência de elétrons (ETS) para a geração de energia celular através de triptosfato de adenosina (ATP)9. O passo multienzimático para aproveitar a energia do fluxo de elétrons através dos complexos I e II ao complexo IV gera um gradiente eletroquímico de prótons através da membrana mitocondrial interna, posteriormente utilizado para fosforilação de difosfato de adenosina (ADP) para ATP via complexo V (F1FO ATP synthase) (Figura 1A).
Primeiro, os portadores de dois elétrons são gerados durante o ciclo tricarboxílico (TCA), glicolise e oxidação piruvato: dinucleotídeo de adenina de nicotinamida (NADH) e dinucleotídeo de adenina dihidroflavina (FADH2). O NADH é oxidado no complexo I (NADH desidrogenase), durante o qual dois elétrons são transferidos para a coenzima Q (quinona é reduzida a quinol), enquanto prótons são bombeados para o espaço intermembrano (IMS). Em segundo lugar, o complexo II (Succinato desidrogenase) oxida FADH2 e alimenta os elétrons para coenzima Q sem bombear prótons. Em terceiro lugar, no complexo III (Cytochrome c oxidoreductase), elétrons da coenzima Q são transferidos para citocromo c enquanto prótons são bombeados para o IMS. Em quarto lugar, o citocromo c transfere os elétrons para o complexo IV (Cytochrome c oxidase), o complexo final para bombear prótons, e onde o oxigênio funciona como um aceitador de elétrons para assimilar prótons, formando água. É esse oxigênio que as mitocôndrias consomem que pode ser medido por um oxígrafo. Finalmente, os prótons gerados a partir do complexo I, complexo III e complexo IV são usados para rodar o complexo V, gerando ASSIM ATP9.
É importante ressaltar que a transferência de elétrons ocorre não apenas de forma linear, de outra forma denotada como a cadeia de transporte de elétrons. Em vez disso, os elétrons podem ser transferidos para a piscina coenzima Q através de múltiplas vias respiratórias e facilitar o fluxo de elétrons convergentes. Os substratos e succinatos do NADH, por exemplo, podem entrar por meio do complexo I e do complexo II, respectivamente. Elétrons da oxidação de ácidos graxos podem ser doados através do complexo de flavoproteína de transferência de elétrons. De fato, uma análise abrangente do OXPHOS requer uma abordagem holística com substratos de combustível apropriados (Figura 1A).

Figura 1: Fosforilação oxidativa mitocondrial e protocolos específicos de substrato e inibidor. (A) Mitoconddrion e esquema do sistema de transferência de elétrons (CI-CIV) e mitocondrial F1F0 ATP synthase (CV). Todas as estruturas são do PDB. As figuras retratam apenas substratos e inibidores descritos neste estudo). (B) Traços de amostra de fluxo de oxigênio em células HEK293 intactas usando protocolo padrão em um dispositivo mHRR. (C) Traços amostrais de fluxo de oxigênio em células HEK293 intactas usando protocolo padrão em um dispositivo cHRR. (D) Traços amostrais de fluxo de oxigênio em fibroblastos humanos permeabilizados de um doador saudável com o respectivo protocolo SUIT. Abreviaturas: 1 = Respiração rotineira de células intactas; 2 = Estado 2; 3 = Estado 3(I); 4 = Estado 3(I) com cítara; 5 = Estado 3 (I+II); 6 = Vazamento(OM); 7 = Capacidade de ETS; 8 = S(ROT); 9 = ROX; 10 = TMPD; 11 = Az. ROT = Rotenone, AM = Antimicina, ATP = Adenosina triphosfato, Az = Azide, OM = Oligomicina, FCCP = Cianeto carbonilizado p-trifluoro-metoxifenil-hidratante; Asc = Ascorbate, TMPD = N,N,N′,N'-tetramethyl-p-fenilelenediamina, Succ = Succinato, M = Malato, P = Piruado, ADP = Difosfato de Adenosina, NAD = Nicotinamida adenina dinucleotídeo, IMS = Espaço Intermembrano, FAD = Flavin adenina dinucleotídeo. Clique aqui para ver uma versão maior desta figura.
{kind=link}
A análise da capacidade mitocondrial oxphos usando HRR tornou-se um método bioquímico instrumental de valor diagnóstico não apenas para defeitos mitocondriais primários10,11, mas estendendo-se a todos os outros domínios da biologia, como câncer e envelhecimento12. O HRR permite a determinação da respiração celular pela análise da capacidade mitocondrial de OXPHOS, que reflete diretamente deficiência do complexo respiratório mitocondrial individual ou combinado, e indiretamente está associada à disfunção celular e ao metabolismo energético alterado9. Metodologicamente, as medidas respiratórias são realizadas utilizando células, tecidos ou mitocôndrias isoladas 11,13,14, com material congelado apenas parcialmente adequado15,16. O tecido congelado é mostrado ter um ETS intacto com estabilidade de supercomplex mantido15. Assim, ao contrário dos intermediários tradicionais do TCA, os respectivos substratos são diretamente alimentados no ETS. No entanto, o acoplamento entre a síntese de ETS e ATP é perdido à medida que a integridade da membrana é comprometida por danos congelantes (formação de cristais de gelo).
Experimentos de respiração normalmente ocorrem a uma temperatura fisiológica de 37 °C para endotermias em células ou tecidos não permeabilizados ou permeabilizados. Enquanto o primeiro considera o contexto metabólico citosomático, este último fornece a contribuição energética dos complexos oxphos individuais e do ATPase através da adição de substratos específicos (e inibidores). A sequência e variação de substratos e inibidores levaram ao desenvolvimento de uma matriz diversificada de protocolos SUIT17 e ensaios18 para abordar várias questões científicas da função OXPHOS (revisadas abaixo de12). O protocolo básico de respiração celular avalia quatro estados diferentes: i) respiração de rotina - a respiração em uma respectiva mídia respiratória sem qualquer adição de substratos ou inibidores que consumam, mas substratos endógenos. Este estado pode revelar defeitos de respiração geralmente ou induzidos por secundários causados, por exemplo, por perfis metabólitos alterados. Em seguida, a adição do oligomicina inibidora de ATPase revela a permeabilidade da membrana mitocondrial interna aos prótons, definida como ii) respiração de vazamento. A titulação subsequente de um prótonophore como o desacontedor cianeto de cyanide carbonyl p-trifluoro-metoxiphenyl-hydrazone (FCCP) permite determinar o estado em que a capacidade de ETS é máxima em um modo de circuito de próton aberto transmembrano, definido como iii) respiração nãocopada. É importante ressaltar que um estado desacoplado também pode ocorrer por intervenções experimentais através de danos mecânicos excessivos às membranas mitocondriais. Por outro lado, o estado não acoplado refere-se ao desacoplamento respiratório por um mecanismo intrínseco que é fisiologicamente controlado. Finalmente, a inibição completa do ETS por adição do complexo inibidor III antimicina e do inibidor complexo I rotenona determina o consumo residual de oxigênio (ROX) de processos não mitocondriais de consumo de oxigênio (Figura 1A-C).
A bioenergetics mitocondrial consiste em cinco estados de respiração distintos19,20. A respiração do estado 1 é sem substratos adicionais ou ADP, exceto pelo que está disponível de forma endógena. Após a adição de ADP, mas ainda assim, sem substratos, a respiração estadual 2 é alcançada. Quando substratos são adicionados, permitindo transferência de elétrons e síntese ATP, a respiração do estado 3 é atingida. Neste estado, a capacidade oxphos pode ser definida em concentrações saturadas de ADP, fosfato inorgânico, oxigênio, nadh- e substratos ligados ao succinato. A respiração do estado 4 ou a respiração DE VAZAMENTO podem ser definidas como um estado sem ADP ou sintetizadores ATP quimicamente inibidos enquanto têm substratos suficientes. Por fim, quando todo o oxigênio é esgotado (anoxico) em um ambiente de câmara fechada, o estado 5 respiração é observado.
Existem vários métodos para avaliar os estados de energia celular14 com dois dispositivos dominando a avaliação atual em tempo real do OXPHOS através da análise do consumo de oxigênio, medido como função da diminuição do oxigênio ao longo do tempo em um sistema de câmara fechada com diferentes aplicabilidade dependentes do modelo experimental e da questão da pesquisa: o respirômetro oroboros de alta resolução e o analisador de fluxo extracelular Seahorse XF. Ambos os dispositivos registram as taxas de consumo de oxigênio como uma diminuição nos picomoles (pmol) de oxigênio (O2) por segundo como um valor absoluto dentro da câmara ou poço de microplato. O consumo específico de oxigênio por massa é obtido pela normalização do respectivo consumo de oxigênio em uma receita tampão específica por número de células (milhões), peso tecidual (mg) ou quantidade de proteína.
O O2k (Instrumentos Oroboros) é um sistema fechado de duas câmaras equipado com um sensor de oxigênio polarográfico (abreviado como respirômetro de alta resolução baseado em câmara: cHRR). Cada câmara experimental contém 2 mL de líquido que é mantido homogêneo por agitadores magnéticos. O sensor de oxigênio polarográfico utiliza uma abordagem amperométrica para medir o oxigênio: contém um cátodo de ouro, um ânodo de cloreto de prata/prata, e entre uma solução KCI criando uma célula eletroquímica sobre a qual uma tensão (0,8 V) é aplicada. O oxigênio do ensaio difere através de uma membrana fluorina de etileno fluorado de 25 μm (O 2-permeável) e sofre redução no cátodo, produzindo peróxido de hidrogênio. No ânodo, a prata é oxidada por peróxido de hidrogênio, gerando uma corrente elétrica. Esta corrente elétrica (ampere) está linearmente relacionada com a pressão parcial de oxigênio. A pressão parcial do oxigênio e o fator de solubilidade de oxigênio do meio de ensaio são usados para calcular a concentração de oxigênio. Uma vez que a pressão parcial do oxigênio depende da temperatura experimental e as medidas polarográficas são sensíveis à temperatura, as flutuações na temperatura precisam de regulação precisa (±0,002 °C) por um bloco de aquecimento Peltier. A temperatura pode ser controlada dentro de uma faixa de 4 °C e 47 °C.
O analisador de fluxo extracelular Seahorse XF (Agilent) é um sistema baseado em placas com formato de microplaca de 24 ou 96 poços no qual três eletrodos de fluorescência medem o consumo de oxigênio ao longo do tempo em cada poço (abreviado como respirômetro de alta resolução baseado em microplaca: mHRR). Um máximo de quatro portas no cartucho de ensaio estão disponíveis para injeção automatizada durante o ensaio. Um ensaio contém vários ciclos, cada um com três fases: 1) mistura, 2) espera e 3) medição. Durante a fase de medição, as sondas de sensores são reduzidas na microplacão criando uma câmara temporariamente fechada contendo volume de 7-10 μL para medir a luz emitida. Esta luz é emitida por fluoroforos embutidos em polímero na ponta das sondas do sensor, que sentem O2 com base na saciamento da fosforescência. A intensidade do sinal de fluorescência é proporcional a O2 e influenciada pela temperatura do sensor e do meio de ensaio. Portanto, a estimativa precisa de oxigênio requer uma abordagem relativa com um poço de fundo sem qualquer amostra. A restauração da concentração de oxigênio ocorre durante a fase de mistura quando o sensor se move para cima e para baixo para misturar o volume acima da câmara temporária. Cada ciclo calcula uma taxa de consumo de oxigênio. A temperatura pode ser controlada dentro de uma faixa de 16 °C e 42 °C.
O HRR é o padrão-ouro para avaliar bioenergetics celulares em doenças primárias e mitocôndrias associadas e metabolismo celular geral. Neste estudo, protocolos básicos para HRR são fornecidos para avaliar a função OXPHOS em células e tecidos.

Figura 2: Fluxo de trabalho para preparações celulares e tecidos para rCR, e preparação celular para respirometria mHRR. (A) Esboço dos protocolos fornecidos. (B) Células mamíferas (passo 1.2): pelota HEK293 igual a 3 x 106 células (painel esquerdo). Tecido não fibroso (passo 1.3): Preparação de cerebelo murino em 2 mL Teflon potter (painel médio). Permeabilização do músculo esquelético induzida por sapona (etapa 1.4) painel direito) para respirometria de HRR. (C) Layout de semeamento de microplaca padrão (etapa 2.4) e verificação de confluência para a análise de células eucarióticas (HEK293) para respirometria mHRR. (D, E) Esquema de carregamento da porta de injeção para respirometria mHRR (etapa 2.4). Clique aqui para ver uma versão maior desta figura.
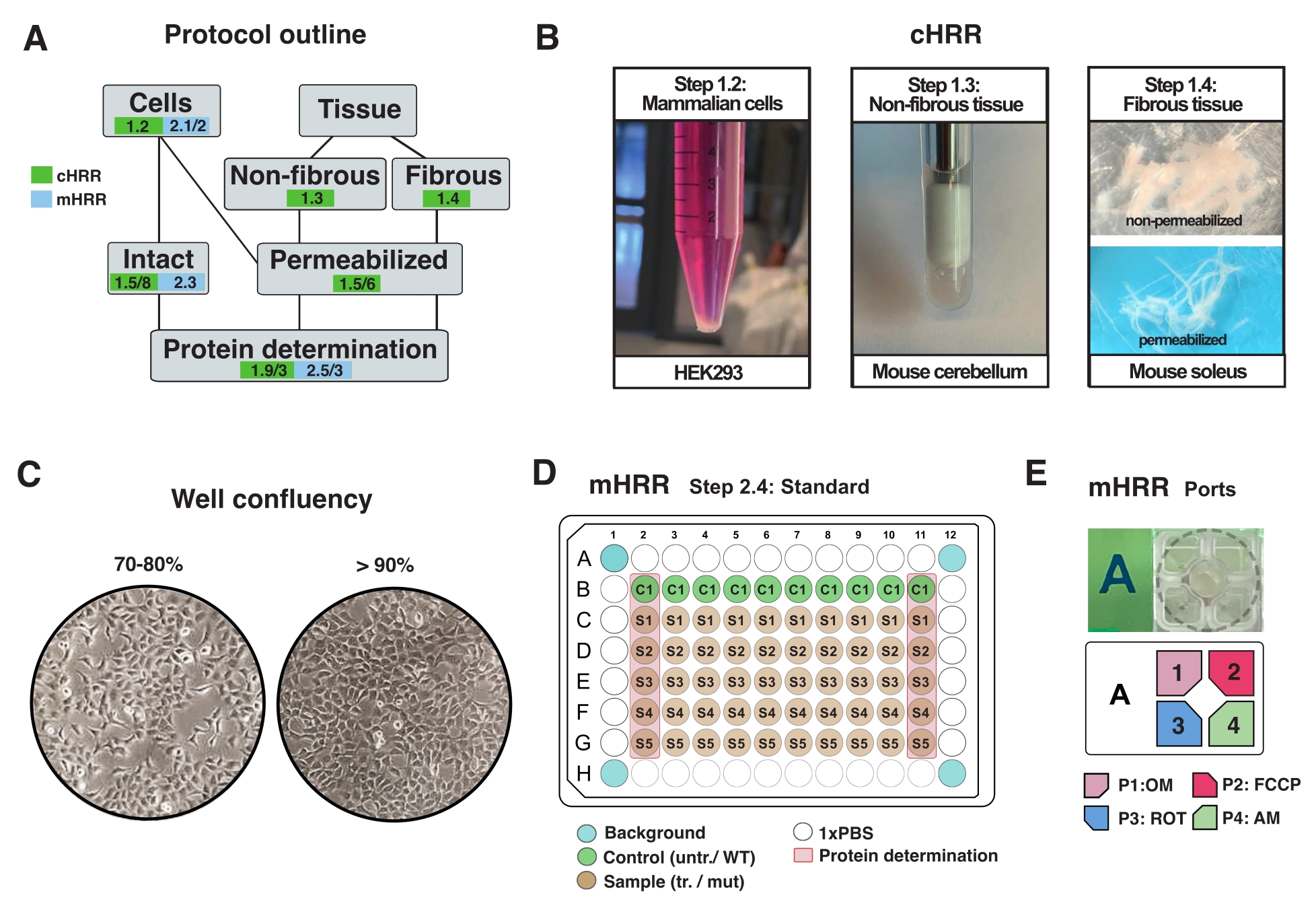{kind=link}
Protocolo
Toda a experimentação animal é realizada de acordo com o National Animal Experiment Review Board e a Agência Regional De Administração Estadual para o Sul da Finlândia. Foram utilizados neste estudo camundongos C57BL/6JOlaHsd masculinos (4-6 meses de idade). O consentimento para o uso de linhas celulares humanas foi obtido do comitê de ética institucional da Universidade de Helsinque.
1. Respirometria de alta resolução: respirômetro baseado em câmara (cHRR)
NOTA: Os experimentos nesta seção do protocolo foram realizados utilizando-se o Oroboros O2k-Core: Oxygraph-2k (Tabela de Materiais)
- Calibração dos sensores de oxigênio
- Respirômetros pré-executados a 37 °C em 2,1 mL de meio de respiração mitocondrial (MiR05, Tabela 1, fator de solubilidade: 0,92) para >45 min e realizar calibração de oxigênio conforme descrito21. Proceda se a variação da linha de base estiver dentro ± 4 pmol/s.
NOTA: Grandes flutuações no sinal de fundo podem indicar a manutenção necessária da membrana do sensor ou traços de inibidores remanescentes na câmara de experimentações anteriores. Recomenda-se uma correção instrumental de fluxo de oxigênio de fundo antes de um lote de experimentos25. - Regisso valores de calibração de oxigênio para monitorar o desempenho da membrana do sensor ao longo do tempo.
NOTA: Isso revela a função do sensor, a estabilidade do sinal ao ruído e quando a manutenção da membrana do sensor é necessária. Dependente da pressão ambiente, entre 180-200 μmol de oxigênio é solubilizado em MiR05. - Remova todo o líquido da câmara antes da adição de qualquer amostra no meio de respiração.
NOTA: Avalie o volume de câmaras de respiração para ser exatamente 2 mL regularmente.
- Respirômetros pré-executados a 37 °C em 2,1 mL de meio de respiração mitocondrial (MiR05, Tabela 1, fator de solubilidade: 0,92) para >45 min e realizar calibração de oxigênio conforme descrito21. Proceda se a variação da linha de base estiver dentro ± 4 pmol/s.
- Preparação de células para respirometria de alta resolução
- Cultura HEK293 células em pratos de 10 cmde 2 diâmetros no meio de Águia Modificada (DMEM) de Dulbecco com alta glicose suplementada com 10% de soro bovino fetal inativado por calor (FBS), GlutaMax, Aminoácidos não essenciais, e Na-Pyrunato22 e uridine23 para suportar o metabolismo com defeito oxphos em uma incubadora a 37 °C a 5% de CO2.
NOTA: Qualquer tipo de célula eucariótica pode ser cultivada. Para a maioria dos tipos de células, a cultura de um prato de 10 cm2 leva a células suficientes (geralmente >3 x 106 células). Verifique rotineiramente a infecção por mycoplasma para evitar efeitos no metabolismo celular e respiração. - Crescer células sem exceder 90% de confluência (Figura 2C).
NOTA: Células com confluência >90% podem apresentar efeitos inibitórios dependentes do crescimento na respiração (se não sincronizadas ou pós-mitóticas). - Lave as células com 1x PBS, desprende com 1 mL de trippsina quente de 0,25%, desative a trippsina adicionando DMEM quente (5 mL/10 cm2 ) e conte as células com hemocitômetro.
- Centrifugar suavemente a solução celular igualando 2,5 x 106 células a 300 x g por 5 min, remover o supernasce completamente e resuspend em 2,5 mL de MiR05 quente (1 x 106 células/mL) (Figura 2A).
- Para células de suspensão, conte e remova a solução igual a 2,5 x 106 células, pelota e continue como mencionado na etapa 1.2.4.
- Execute o protocolo SUIT para otimização de permeabilização (etapa 1.6), célula ou tecido permeabilizizado (passo 1.5) ou células intactas (etapa 1.7)
NOTA: Para resultados consistentes, recomenda-se manter a concentração celular constante (por exemplo, 1 x 106 células/mL). Embora a respiração seja independente da densidade celular no respirômetro24, substratos e inibidores estão em concentração comparável ao longo dos experimentos se os números de células forem mantidos constantes.
- Cultura HEK293 células em pratos de 10 cmde 2 diâmetros no meio de Águia Modificada (DMEM) de Dulbecco com alta glicose suplementada com 10% de soro bovino fetal inativado por calor (FBS), GlutaMax, Aminoácidos não essenciais, e Na-Pyrunato22 e uridine23 para suportar o metabolismo com defeito oxphos em uma incubadora a 37 °C a 5% de CO2.
- Preparação de tecido não fibroso (por exemplo, cérebro, fígado) para respirometria de alta resolução
- Extite um pedaço homogêneo de tecido, 30-40 mg de peso, ou use todo o órgão (cerebelo do rato neste caso).
NOTA: Se o tecido não for usado imediatamente, mantenha em 2 mL de MiR05 gelado permitindo a preservação por até 2 h para a maioria dos tecidos. Os tempos individuais de armazenamento de tecidos precisam ser avaliados em séries temporidas. - Borrite o tecido seco com um papel filtro Whatman (cuidado: a matéria de tecido mole tende a grudar).
- Coloque a peça de tecido de 30-40 mg em um politetrafluoroetileno de 2 mL resfriado ol politetrafluoroetileno homogeneizador.
- Adicione uma quantidade apropriada de MiR05 para obter 20 mg/mL para manter a relação tecido-tampão. Mantenha a quantidade total > 1,5 mL e <2 mL para evitar fluidos insuficientes ou excessivos para permeabilização mecânica adequada.
- Insira o pilão, lise o tecido lentamente, retraindo o pilão cuidadosamente, evitando a geração de um vácuo causando danos excessivos no tecido.
- Realize 7 traçados no total (1x definidos como um curso para cima e para baixo) até lise (aparente como um líquido turvo sem detritos maiores) (Figura 2B).
NOTA: O número de derrames para lise apropriada precisa ser testado para cada tecido, avaliando a integridade da membrana mitocondrial externa através da resposta citocromática C (etapa 1.5.11). Tecido conjuntivo de difícil de lise ou partes de vasos podem permanecer. - Decante o tecido líscido em um tubo de centrífuga de 15 mL.
- Lave o interior do oleiro com uma quantidade igual de MiR05 usado na etapa de lise (por exemplo, 1,5 mL) e adicione ao tubo de 15 mL agora contendo 3-4 mL de MiR05 a 10 mg/mL de tecido lysate.
- Adicione 2 mL de MiR05 simples por câmara para aquecer a 37 °C.
- Gire o tubo para distribuição igual antes de pipetar 500 μL (igualando 5 mg) de cada lise por câmara lentamente para minimizar o estresse do frio a 37 °C.
- Aguarde >3 minutos para que o conteúdo da câmara aqueça até 37 °C antes de fechar a câmara. Remova o excesso de fluido em cima da rolha (quantidade por câmara após o fechamento: 4 mgs).
- Execute o protocolo SUIT para permeabilized padrão (etapa 1.5).
- Extite um pedaço homogêneo de tecido, 30-40 mg de peso, ou use todo o órgão (cerebelo do rato neste caso).
- Preparação de tecido fibroso (músculo esquelético, músculo cardíaco) para respirometria de alta resolução
- Extrair o tecido duro, remover o tecido conjuntivo e a gordura dos músculos usando fórceps afiados em 2 mL de BIOPS gelado (Tabela 2) sob um microscópio de dissecção.
- Separe os feixes de fibra (~4 mg) ao longo do eixo longitudinal com fórceps afiados. Provoque as fibras o suficiente para obter uma estrutura em forma de malha (Figura 2B).
NOTA: A separação e permeabilização adequada da fibra mecânica é indicada pela perda da mioglobina do pigmento vermelho e pelo aumento da translucência. - Lave e permeabilize o feixe de fibras em saponina (50 μg/mL em BIOPS, preparado fresco) por 20 min a 4 °C (as fibras se tornam translúcidas, indicando permeabilização completa, Figura 2B).
- Lave as fibras duas vezes em MiR05 por 5 min por lavagem a 4 °C.
- Estaque seque com papel filtro e pese antes de adicionar à câmara cheia de 2,1 mL MiR05.
- Introduza rolhas sem fechar totalmente, em seguida, oxigenar as câmaras com 2 mL de O2 puro usando uma seringa de 20 mL e fechar as câmaras torcendo as rolhas em um movimento rotativo. Mantenha a concentração de O2 entre 300-500 μM durante o experimento para evitar a limitação da difusão de oxigênio.
- Protocolo para avaliação da respiração de rotina em células ou tecidos
- Adicione a amostra à câmara conforme mencionado nas etapas 1.5.2-1.5.3.
- Adicione 2,3 mL de suspensão celular MiR05 quente (entrada padrão: 1 x 106 células/mL como na etapa 1.2 ou 2 mg de tecido/mL como na etapa 1.3)
- Músculo esquelético e cardíaco (passo 1.4): Adicione ~4 mg de fibras de saponina permeabilized a 2,3 mL pré-equipados de MiR05 quente considerando as etapas 1.4.4-1.4.6
- Executar câmaras a 37 °C e uma velocidade de agitação de 700 rpm. Aguarde >3 minutos para permitir que a mídia desgase e feche as câmaras torcendo a rolha em um movimento rotativo. A estabilização do bloco peltier indica atingir a temperatura definida.
- (OPCIONAL) Altere a velocidade do agitador para 300 rpm para permitir que as bolhas restantes escapem através do capilar da rolha.
- Aspire qualquer excesso de líquido em cima da rolha. Aguarde 10 minutos até que um sinal de fluxo de oxigênio estável seja alcançado com qualquer tipo de amostra para registrar a respiração de rotina/estado 1, Figura 1B).
- Para medições de respiração em células permeabilizadas e tecidos, continue com a etapa 1.6. Para células intactas com passo 1.8.
- Protocolo para análise de OXPHOS em células ou tecidos permeabilizados
- Use amostras de tecido lísticula (permeabilized) ou células permeabilizes adicionando 1 μL de digitonina (8,1 mM de digitalonina em sulfoxida de dimetil (DMSO)) para uma concentração final de 5 μg/mL às células permeabilize. O fluxo cairá e deve estabilizar em >5 min.
ATENÇÃO: A digitonina é agudamente tóxica para o trato respiratório, em contato com a pele ou quando engolida.
NOTA: A injeção de todos os produtos químicos é realizada com seringas de vidro de precisão. Use seringas apenas para produtos químicos indicados para evitar contaminação cruzada e lavar completamente na água e etoh após o uso. Seringas bloqueadas podem exigir ultrassônicas em ddH2O quente ou um fio de limpeza para desalojar quaisquer tamancos químicos. Retraia sempre um excedente da respectiva solução de estoque na seringa para evitar a introdução de ar nas câmaras. Inspecione o interior das câmaras para a introdução de ar após cada injeção. Registo cada passo até o fluxo planalto. - Adicione em rápida sucessão: 5 μL de 0,4 M de malato (M) para uma concentração final de 1 mM, 5 μL de piruvato de 2,0 M (P; preparado recentemente), para uma concentração final de 5 mM, 4 μL de 2,5 M de glutamato (G) para uma concentração final de 5 mM.
- Após o fluxo anterior nivelado, adicione 5 μL (10 μL para tecido muscular) de 0,5 M difosfato de adenosina (ADP, alíquotas armazenadas a -80 °C) para uma concentração final de 1,25 mM.
NOTA: Tecidos como músculo podem precisar de uma concentração diferente para alcançar a saturação. - Adicione 5 μL de 4 mM citocromo C (ctil) para uma concentração final de 10 μM.
NOTA: Opcional para as células avaliarem a qualidade da permeabilização. - Adicione 16 μL de 1,25 M de succinato (S) para uma concentração final de 10 mM. (OPCIONAL) Adicione 3 μL de 0,5 M ADP para uma concentração final de 2 mM para controlar a saturação da concentração de ADP.
- Para células e tecido não fibroso, adicione 2 μL de oligomicina de 1 mg/mL (OM) para uma concentração final de 1 μg/mL.
ATENÇÃO: Todos os inibidores de ETS utilizados são altamente tóxicos.
NOTA: A oligomicina pode exigir titulação para uma concentração ideal, pois pode reprimir a capacidade de ETS e é omitida para tecido muscular. Reoxigenato aqui quando o tecido muscular é avaliado e se O2 está abaixo de 300 μM. - Titrate FCCP de um estoque de 2 mM, adicione 0,6 μL com etapas subsequentes de 0,2 μL até que nenhum aumento na respiração e respiração seja maximamente desacoplado (teórico: não acoplado).
- Adicione 1 μL de 1 mM rotenona (ROT) para uma concentração final de 0,5 μM. Adicione 2 μL de 1 mg/mL de antimicina (AM) para uma concentração final de 1 μg/mL.
- Reoxigenar as câmaras para alcançar um nível de oxigênio semelhante (~150 μM) em todas as câmaras, levantando lentamente o êmbolo em movimento de torção.
- Adicione 5 μL de 0,8 M ascorbate para uma concentração final de 2 mM imediatamente seguido por 5 μL de 0,2 M N,N,N',N',N'-tetramethyl-p-fenilediamina (TMPD) para uma concentração final de 0,5 mM para avaliar a atividade iv complexa (opcional).
- Adicione 5 μL de 4 M de azida para uma concentração final de 10 mM imediatamente quando o pico O2 fluxo for atingido com TMPD. Continue a corrida de >5 min para avaliar a auto-oxidação do TMPD para o cálculo complexo do nível base IV.
- Reconta as células para confirmar a contagem de células pré-corrida e continuar com a etapa 1.9.
NOTA: A digitonina-permeabilização (apenas para células) precisa ser titulada em experimentos experimentais para atingir o fluxo máximo e não afetar a integridade da membrana mitocondrial (ver passo 1.7). Amostras permeabilizadas (especialmente tecido muscular) com aumento de >10% na taxa de respiração após a adição de citocromo c devem ser excluídas de análises posteriores devido a danos na membrana mitocondrial externa. Espera-se um pequeno mergulho no fluxo após a adição de produtos químicos dissolvidos por EtOH.
- Use amostras de tecido lísticula (permeabilized) ou células permeabilizes adicionando 1 μL de digitonina (8,1 mM de digitalonina em sulfoxida de dimetil (DMSO)) para uma concentração final de 5 μg/mL às células permeabilize. O fluxo cairá e deve estabilizar em >5 min.
- Protocolo para determinar condições ideais de permeabilização para células
- Adicione células conforme descrito nas etapas 1.2 e 1.5.2.
- Pegue 10 μL de 10 mg/mL de digitonina e adicione 10 μL de DMSO para diluir para 5 mg/mL.
- Adicione 1 μL de rotenona (1 mM de estoque). Adicione 10 μL de succinato (2 mM de estoque) e 5 μL de ADP (estoque de 0,5 M).
- Titulação 1 μL de digitonina (2,5 mg por passo) repetidamente até que a respiração não aumente ainda mais e seja máxima.
NOTA: A diminuição da respiração indica uma concentração excessiva de digitonina.
- Protocolo para análise de OXPHOS em células intactas
- Após a respiração de rotina (etapa 1.6.1-1.6.6), adicione 2 μL de 0,01 mM de oligomicina para uma concentração final de 10 nM.
- Titrate FCCP a partir de 2 mM de estoque, adicione 0,6 μL com etapas subsequentes de 0,2 μL até que nenhum aumento adicional na respiração e respiração seja maximamente desacoplado (teórico: não acoplado)
- Adicione 1 μL de rotenona de 1 mM para uma concentração final de 0,5 μM. Adicione 2 μL de 1 mg/mL de antimicina para uma concentração final de 1 μg/mL.
- Reoxigenar a câmara ao mesmo nível de oxigênio (~150 μM) levantando lentamente o êmbolo em movimento de torção.
- Adicione 5 μL de 0,8 M de ascorbate para concentração final de 2 mM. Adicione imediatamente 5 μL de 0,2 M TMPD para uma concentração final de 0,5 mM para avaliar a atividade iv complexa.
NOTA: Prepare um lote fresco antes que qualquer conjunto maior de experimentos, pois o TMPD seja propenso à auto-oxidação. A atividade pode diminuir ao longo do tempo quando armazenada a -20 °C. - Adicione 5 μL de 4 M de azida para uma concentração final de 10 mM imediatamente quando o pico O2 fluxo for atingido com TMPD. Continue a execução por >5 min para avaliar a auto-oxidação do TMPD para o cálculo complexo do nível base IV.
- Reconte células para confirmar a contagem de células pré-corrida e continuar com a etapa 1.9.
- Coleta de amostras pós-escorrida
- Colete exatamente 1 mL de suspensão MiR05 de cada câmara (com agitadores ligados) em um tubo de 1,5 mL tubo de 1,5 mL.
- Centrifugar a 1000 x g para células permeabilizadas ou a 20.000 x g para lise tecidual. Remova o sobrenante e congele a pelota a -80 °C para processamento adicional (seção 3).
- Análise dos protocolos DA SUIT
- Analise o fluxo de oxigênio (pmol/s, normalizado para entrada) em cada platô após a adição de um substrato ou inibidor (Figura 1C e Figura 3A). Exporte os valores para uma planilha.
- Subtrair o valor residual de consumo de oxigênio (ROX, Figura 1C e Figura 3C) de todos os valores de cada execução experimental. Subtrair a respiração residual de azide do TMPD para obter respiração IV complexa.
- Plote os valores absolutos normalizados para entrada celular (Figura 3A, B) ou de tecido (Figura 5A,B). Calcule as razões de controle de fluxo (etapa 1,11) ou normalize-as para entrada de proteína (Figura 3C).
- Cálculo da razão de controle de fluxo
- Adquira um índice de função respiratória e controle de acoplamento utilizando razões de controle de fluxo (FCR)9,26.
NOTA: Isso permite avaliar a qualidade mitocondrial intrínseca, independente da quantidade mitocondrial. Além disso, as razões de controle de fluxo (FCR) são comparáveis dentro das mesmas linhas celulares que permitem o controle de qualidade do reagente (as respectivas FCRs são obtidas através dos valores de referência numerados indicados nas Figuras 1B-D e Figura 3C). - Calcule a razão de controle respiratório para o acoplamento de OXPHOS ao LEAK usando a Equação 1.
Equação 1: FCRADP = 5/6 = Estado 3 / Estado 4 - Calcule o FCR para avaliar a respiração dependente de NADH usando a Equação 2
Equação 2: Estado fcr3 (I) = 3/5 = Estado 3 (I) / Estado 3 (I+II) - Calcule o FCR para avaliar a respiração dependente de succinato usando a Equação 3.
Equação 3: Estado FCR3 (II) = 8/7 = Spodridão /capacidade de ETS - Calcule o FCR para avaliar acoplado a desacoplamento usando a Equação 4.
Equação 4: FCRacoplado/desacoplado = 5/7 = Estado 3 (I+II) /Capacidade de ETS - Para testar a integridade da membrana externa mitocondrial, use a Equação 5.
Equação 5: % dano de membrana externa mitocondrial = 3/4 = Estado 3 (I) / Estado 3 (I) com cyt c
- Adquira um índice de função respiratória e controle de acoplamento utilizando razões de controle de fluxo (FCR)9,26.
2. Respirometria de alta resolução: Respirômetro à base de microplaco (mHRR)
NOTA: Os experimentos nesta seção do protocolo foram realizados utilizando-se o Analisador de Fluxo Extracelular Seahorse XFe96 (Tabela de Materiais)
- Cultura celular
- Cultura qualquer tipo de célula. Os adeptos (por exemplo, colágeno, laminina) podem ser usados para facilitar o apego celular. Aqui, as células HEK293 são cultivadas como antes (passo 1.3).
- Um dia antes do experimento, desprende as células e transfira-as para uma microplaca designada mHRR 96-well para obter confluência ideal no dia do experimento (80%-100%) (Figura 2C).
NOTA: Para o mHRR, as densidades de células microplacas são críticas. As propriedades de crescimento individual das linhas de células ou tratamentos que afetam o crescimento precisam ser contabilizadas para garantir confluência comparável no dia do experimento.
- Preparação de células para respirometria de alta resolução
- Colher e resuspensar as células suficientemente antes da semeadura
NOTA: Recomenda-se às células de sementes a partir da mesma diluição para réplicas. - Semeadas as células de acordo com as taxas de crescimento de linhas celulares individuais ou propriedades de crescimento em tratamento.
NOTA: Otimize em uma microplacão padrão de 96 poços e extrapolule a densidade celular para uma microplacão específica de ensaio de 96 poços. Nesta configuração, as células 7 x 104 HEK293 WT foram semeadas por poço de um poço de 96 poços. As primeira e últimas colunas da placa de 96 poços são utilizadas para determinação proteica (Figura 2C). Os quatro poços de canto não devem conter células e são usados para correção experimental de fundo. Idealmente, poços próximos às bordas estão vazios para minimizar o efeito de borda (por exemplo, as células apresentam taxas de crescimento alteradas causadas por efeitos de temperatura) (Figura 2C, D).
- Colher e resuspensar as células suficientemente antes da semeadura
- Preparação de placas de sensores, carregamento de inibidores
- No dia do ensaio, suplementar 38,8 mL de médio com 0,4 mL de glicose de 1 M, 0,4 mL de glutamina de 200 mM e 0,4 mL de 100 mM Na-Pyruato.
NOTA: a respiração mHRR requer um meio de DMEM não tamponado especializado em pH 7.4. Em geral, 40 mL devem ser suficientes para um experimento com uma microplacão de 96 poços. - Aqueça o ensaio de respiração médio a 37 °C e troque o meio de cultura celular pelo meio de ensaio respiratório lavando duas vezes com 80 μL por poço.
- Coloque a placa com as células em uma incubadora de 37 °C sem CO2 por 60 minutos antes do ensaio.
NOTA: Esta etapa é essencial para desgasar a placa, pois o CO2 pode afetar os resultados respiratórios, e o soro no meio pode produzir bolhas durante o ensaio. - Alíquotas inibidoras pré-aquecedoras para OM, FCCP, ROT e AM a 37 °C e tire a placa do sensor da incubadora.
- Diluir OM, FCCP, ROT e AM em 3 mL de médio ensaio para uma concentração final de poço de 1,5 μM, 1,125 μM e 1 μM, respectivamente. Encha em portas separadas conforme indicado na Figura 2E.
NOTA: Recomenda-se uma pipeta multicanal para encher o cartucho do sensor. Uma vez que o ar pressurizado é usado para injetar compostos, todas as portas devem ser preenchidas com uma quantidade igual de volume líquido sempre que uma porta é preenchida com um composto. ROT e AM podem ser combinados em uma porta. Os inibidores podem ser dissolvidos em EtOH ou DMSO. - Inspecione as portas de injeção e verifique um volume de carregamento uniforme para cada porta.
NOTA: Todas as portas contêm um orifício na parte inferior para injeção. Deve-se tomar cuidado ao mover a placa do sensor. Bolhas de ar podem ser removidas usando uma agulha.
- No dia do ensaio, suplementar 38,8 mL de médio com 0,4 mL de glicose de 1 M, 0,4 mL de glutamina de 200 mM e 0,4 mL de 100 mM Na-Pyruato.
- Protocolo para avaliação de oxigênio em células intactas
- Na véspera do ensaio, realize as etapas 2.4.2-2.4.7.
- Alíquota de 20 mL da solução calibrante em um tubo cônico de 50 mL.
- Abra o Kit de Ensaio de Fluxo Extracelular e remova o conteúdo.
- Coloque o cartucho do sensor invertido ao lado da placa de utilidade. Pipeta 200 μL de solução calibrante em cada poço da placa de utilidade.
- Coloque o cartucho do sensor na placa do utilitário prestando atenção que todos os sensores estão submersos.
- Coloque a placa em uma incubadora de 37 °C sem CO2 durante a noite ou mínima de 12h. Verifique se a umidade dentro da incubadora é suficiente para evitar a evaporação do calibrador.
- Ligue o sistema e o computador baseados em microplacos para estar pronto para usar no dia seguinte (a máquina requer um mínimo de 3h para equilibrar até 37 °C antes de realizar um ensaio).
NOTA: Para estabilidade do sinal, aumente os pontos de medição para 6 em vez de 3 ciclos de medição por estado respiratório. Cada ciclo consiste em 3 min de mistura e 3 min de medição. - No dia do ensaio XF, realize as etapas 2.4.9-2.4.20.
- Verifique a confluência da placa de cultura celular, a morfologia das células, e que os poços de fundo estão vazios.
- Lave as células com o meio de respiração preparado como mencionado nas etapas 2.4.11-2.4.12.
- Remova todos, exceto 20 μL do meio de cultura de cada poço. Remova 55 μL se o meio de cultura tiver 80 μL devido à evaporação durante a noite (aproximadamente 5 μL).
- Lave as células duas vezes com 90 μL de meio de ensaio. Por fim, adicione 100 μL de meio de ensaio. O volume final deve ser de 120 μL.
NOTA: Recomenda-se uma pipeta multicanal para esta etapa para garantir que o mesmo procedimento de lavagem tenha sido aplicado a cada condição experimental (depende da configuração da placa). Ao aspirar, incline a placa para um ângulo de 45° e coloque as pontas da pipeta no canto dos poços para aspiração e injeção de líquidos. É imperativo tomar cuidado durante a lavagem, pois certas células podem facilmente se desprender do fundo da placa de cultura celular. - Coloque a placa em uma incubadora de 37 °C sem CO2 por 60 minutos antes do ensaio.
- Recupere a placa do cartucho do sensor hidratado da incubadora sem CO2.
- Descarte a solução calibrante antiga e substitua-a por uma solução calibrante fresca, pré-armada a 37 °C.
- Preparar inibidores e médio de ensaio (3 mL por inibidor para um total de 12 mL de meio de ensaio) e usar um reservatório de pipeta para o inibidor de carga em portas.
- Abra o software e execute um modelo pré-projetado ou novo. Encha o mapa da placa, ajuste as titulações e ciclos de medição e, em seguida, pressione Comece a iniciar a calibração dos sensores ópticos.
- Remova a tampa do cartucho carregado e coloque-a no slot que desliza automaticamente para fora da máquina, verificando se as marcas no canto inferior direito da placa se alinham com o triângulo no canto inferior direito da ranhura.
- Clique em Continuar a realizar a calibração automática, com duração aproximada de 20 minutos.
- Após a calibração, remova a placa de utilidade contendo o calibrante.
- Retire a tampa da microplacão contendo as células e coloque a placa no slot quando solicitada pela máquina. Clique em Continuar para iniciar a execução.
- Coleta de amostras pós-escorrida
- Retire a placa da máquina, remova cuidadosamente a mídia restante do ensaio sem perturbar as células e congele toda a placa a -80 °C para processamento adicional (seção 3).
3. Determinação da proteína utilizando o ensaio de ácido bicinchonínico (ensaio BCA)
- Prepare a albumina de soro bovino diluído (BSA) em tampão utilizado para extração de proteínas e compatível com BCA: 2 mg/mL, 1,5 mg/mL, 1 mg/mL, 0,5 mg/mL, 0,25 mg/mL e 0 mg/mL para curva padrão em duplicatas.
- Extrair proteínas reutilizando em um tampão de lise apropriado (por exemplo, RIPA) com 20 μL por poço para mHRR ou 100 μL por pelota contida dentro de um tubo de 1,5 mL para cHRR.
- Incubar a placa mHRR ou tubo de 1,5 mL contendo lises proteicas por 30 minutos no gelo.
- Centrifugar o tubo de 1,5 mL contendo o liseféico a 4 °C a 20.000 x g por 20 min e transfira o supernante resultante para um novo tubo limpo de 1,5 mL.
- Use 10 μL por amostra em duplicatas e padrões em uma placa de microtiter. Adicione 200 μL de reagente de trabalho BCA e incubar por >15 min.
- Leia a placa em um espectrofotômetro padrão em um comprimento de onda de 562 nm e calcule as concentrações proteicas usando uma curva padrão BSA.
- Normalizar os resultados da respiração para a concentração de proteínas.
NOTA: A normalização da quantidade proteica permite corroborar densidades de semeadura celular ou entrada de peso molhado. As proteínas extraídas são adequadas para a imunoblotação subsequente contra subunidades do ETS, por exemplo, mas não representam totalmente a amostra nativa (por exemplo, perda de locais de fosforilação).
Resultados
Aqui, fornecemos protocolos para determinar os bioenergetics mitocondriais em células eucarióticas, tecido não fibroso (por exemplo, cerebelo) e tecido fibroso (por exemplo, músculo esquelético). Para células eucarióticas, hek293 com nocaute projetado por CRISPR de duas proteínas diferentes associadas à tradução mitocondrial resultando em múltiplas (CRISPRKO1) e deficiência oxphos grave/completa (CRISPRKO2) foram medidas com cHRR (Figura 3A-C
Discussão
Tradicionalmente, bioenergetics mitocondriais tem sido estudado com eletrodos de oxigênio do tipo Clark. A falta de resolução e throughput, no entanto, justificava o avanço tecnológico. Até o momento, o O2k (conhecido como cHRR) e o Seahorse XF96 Flux Analyzer (conhecido como mHRR) foram amplamente adotados no campo dos bioenergésicos celulares. Aqui, apresentamos uma coleção compreensível de protocolos para análise do metabolismo de energia celular através da avaliação da respiração mitocondrial usando c...
Divulgações
Nenhum conflito de interesses para revelar.
Agradecimentos
Este trabalho foi apoiado por financiamento da Academia da Finlândia (C.B.J), da Fundação Magnus Ehrnroot (C.B.J), e de uma bolsa de doutorado da Integrated Life Sciences Graduate School (R.A.).
Materiais
| Name | Company | Catalog Number | Comments |
| 2 mL Potter-Elvehjem Glass/PTFE Tissue Grinder/Homogenizer | Omni International | 07-358029 | |
| 95% O2, 5% CO2 medical gas mixture | Potter for tissue grinding | ||
| ADP | Sigma | A 4386 | |
| Antimycin A | Sigma | A 8674 | Chemical |
| Ascorbate | Merck | PHR1279-1G | Chemical, dissolve in ethanol |
| BSA (fatty accid free) | Sigma | A 6003 | Chemical |
| CaCO3 | Sigma | C 4830 | Chemical |
| Cytochrome c | Sigma | C 7752 | Chemical |
| Digitonin | Sigma | D 5628 | Chemical |
| Dithiothreitol | Sigma | D 0632 | Chemical, dissolve in DMSO |
| D-Sucrose | Roth | 4621.1 | Chemical |
| Dulbecco’s modified Eagle’s medium (High glucose) | Fisher Scientific | 41965-039 | Chemical |
| Dulbecco’s modified Eagle’s medium (No Glucose) | Fisher Scientific | A14430-01 | |
| EGTA | Sigma | E 4378 | |
| Etomoxir | Sigma | E1905 | Chemical |
| Falcon 15 ml Conical Centrifuge Tubes | Fisher Scientific | AM12500 | Chemical |
| Falcon 50 ml Conical Centrifuge Tubes | Fisher Scientific | AM12501 | |
| FCCP | Sigma | C 2920 | |
| Glucose | Sigma | G7021 | Chemical, dissolve in ethanol |
| Glutamate | Sigma | G 1626 | Chemical |
| GlutaMax (100x) (200 nM L-alanyl-L-glutamine dipeptide) | Fisher Scientific | 35050061 | Chemical |
| HEK293 cells | ATTC | CRL-1573 | |
| Hemocytometer | Fisher Scientific | 0267151B | Instrument for cell counting |
| Hepes | Sigma | H 7523 | Chemical |
| Imidazole | Fluka | 56750 | Chemical |
| KCl | Merck | 1.04936 | Chemical |
| L-carnitine | Sigma | C0283 | Chemical |
| Malate | Sigma | M 1000 | Chemical |
| MES hydrate | Sigma | M8250 | Chemical |
| MgCl2 | Sigma | M 9272 | Chemical |
| Na2ATP | Sigma | A 2383 | Chemical |
| Na2Phosphocreatine | Sigma | P 7936 | Chemical |
| Na-pyruvate (100 mM) (100x) | Fisher Scientific | 11360070 | |
| NEAA (Non-essential amino acids) 100x | Fisher Scientific | 11140035 | |
| Normal FBS (10x) | Fisher Scientific | 10500064 | |
| O2k-Core: Oxygraph-2k | Oroboros Instruments | 10000-02 | High-resolution respirometry instrument |
| O2k-Titration Set | Oroboros Instruments | 20820-03 | Hamilton syringes for chemical injections |
| Oligomycin | Sigma | O 4876 | Chemical, dissolve in ethanol |
| Palmitoylcarnitine | Sigma | P 4509 | Chemical |
| Penicillin-Streptomycin | Fisher Scientific | 15140122 | |
| Pierce BCA Protein Assay Kit | Fisher Scientific | 23227 | |
| Pyruvate | Sigma | P 2256 | Chemical |
| RIPA-Buffer | Fisher Scientific | 89900 | Chemical |
| Rotenone | Sigma | R 8875 | Chemical, dissolve in ethanol |
| Saponin | Sigma | S7900 | Chemical |
Seahorse XF DMEM assay medium pack, pH 7.4 | Agilent, Santa Clara, CA | 103680-100 | |
| Seahorse XFe96 Extracellular Flux Analyzer | Agilent, Santa Clara, CA | High-throughput respirometry instrument | |
| Seahorse XFe96 FluxPak | Agilent, Santa Clara, CA | Includes assay plates, cartridges, loading guides for transferring compounds to the assay cartridge, and calibrant solution. | |
| Small scissors | Fisher Scientific | 08-951-20 | |
| Sodium azide | Sigma | S2002 | Chemical |
| Succinate | Sigma | S 2378 | Chemical |
| Taurine | Sigma | T 8691 | Chemical |
| TMPD | Sigma | T 3134 | Chemical |
| Trypan Blue solution | Merck | 72-57-1 | Chemical |
| Trypsin 0.25% EDTA | Fisher Scientific | 25200056 | |
| Two thin-edged forceps | Fisher Scientific | 12-000-122 | |
| Uridine stock (500x) | Sigma | U3750 | Chemical |
Referências
- McBride, H. M., Neuspiel, M., Wasiak, S. Mitochondria: More than just a powerhouse. Current Biology. 16 (14), 551-560 (2006).
- Mehta, M. M., Weinberg, S. E., Chandel, N. S. Mitochondrial control of immunity. Beyond ATP. Nature Reviews Immunology. 17 (10), 608-620 (2017).
- Spinelli, J. B., Haigis, M. C. The multifaceted contributions of mitochondria to cellular metabolism. Nature Cell Biology. 20 (7), 745-754 (2018).
- Gnaiger, E. Capacity of oxidative phosphorylation in human skeletal muscle. New perspectives of mitochondrial physiology. International Journal of Biochemistry and Cell Biology. 41 (10), 1837-1845 (2009).
- Gorman, G. S., et al. Mitochondrial diseases. Nature Reviews Disease Primers. 2, 1-23 (2016).
- Boushel, R., Gnaiger, E., Schjerling, P., Skovbro, M., Kraunsøe, R., Dela, F. Patients with type 2 diabetes have normal mitochondrial function in skeletal muscle. Diabetologia. 50 (4), 790-796 (2007).
- Cogliati, S., et al. Mitochondrial cristae shape determines respiratory chain supercomplexes assembly and respiratory efficiency. Cell. 155 (1), 160-171 (2013).
- Kühlbrandt, W. Structure and function of mitochondrial membrane protein complexes. BMC Biology. 13, 1-11 (2015).
- Gnaiger, E. Mitochondrial pathways and Respiratory control. An introduction to OXPHOS analysis. Bioenergetics communications. 5th ed. , (2020).
- Jackson, C. B., et al. Mutations in SDHD lead to autosomal recessive encephalomyopathy and isolated mitochondrial complex II deficiency. Journal of Medical Genetics. 51 (3), 170-175 (2014).
- Pesta, D., Gnaiger, E. High-resolution respirometry: OXPHOS protocols for human cells and permeabilized fibers from small biopsies of human muscle. Methods in Molecular Biology. 810, 25-58 (2012).
- Horan, M. P., Pichaud, N., Ballard, J. W. O. Review: Quantifying mitochondrial dysfunction in complex diseases of aging. Journals of Gerontology - Series A Biological Sciences and Medical Sciences. 67 (10), 1022-1035 (2012).
- Doerrier, C., Garcia-Souza, L. F., Krumschnabel, G., Wohlfarter, Y., Mészáros, A. T., Gnaiger, E. High-resolution fluorespirometry and oxphos protocols for human cells, permeabilized fibers from small biopsies of muscle, and isolated mitochondria. Methods in Molecular Biology. 1782, 31-70 (2018).
- Zhang, J., et al. Measuring energy metabolism in cultured cells, including human pluripotent stem cells and differentiated cells. Nature Protocols. 7 (6), 1068-1085 (2012).
- García-Roche, M., Casal, A., Carriquiry, M., Radi, R., Quijano, C., Cassina, A. Respiratory analysis of coupled mitochondria in cryopreserved liver biopsies. Redox Biology. 17, 207-212 (2018).
- Acin-Perez, R., et al. A novel approach to measure mitochondrial respiration in frozen biological samples. The EMBO Journal. 39 (13), 1-18 (2020).
- Cell metabolism assay kits. Seahorse assay kits and media Available from: https://www.agilent.com/en/product/cell-analysis/real-time-cell-metabolic-analysis/xf-assay-lits-reagents-cell-assay-media (2021)
- Chance, B., Williams, G. R. A method for the localization of sites for oxidative phosphorylation. Nature. 176 (4475), 250-254 (1955).
- Gnaiger, E., et al. Mitochondrial respiratory states and rates. MitoFit Preprint Arch. , (2019).
- Gnaiger, E. O2k-procedures: SOP O2k quality control 1: Polarographic oxygen sensors and accuracy of calibration Section Page. Oroboros. 03 (18), 1-21 (2020).
- Robinson, B. H., Petrova-Benedict, R., Buncic, J. R., Wallace, D. C. Nonviability of cells with oxidative defects in galactose medium: A screening test for affected patient fibroblasts. Biochemical Medicine and Metabolic Biology. 48 (2), 122-126 (1992).
- King, M. P., Attardi, G. Human cells lacking mtDNA: Repopulation with exogenous mitochondria by complementation. Science. 246 (4929), 500-503 (1989).
- Makrecka-Kuka, M., Krumschnabel, G., Gnaiger, E. High-resolution respirometry for simultaneous measurement of oxygen and hydrogen peroxide fluxes in permeabilized cells, tissue homogenate and isolated mitochondria. Biomolecules. 5 (3), 1319-1338 (2015).
- Fasching, M., Gnaiger, E. O2k quality control 2: Instrumental oxygen background correction and accuracy of oxygen flux. Mitochondrial Physiology Network. 14 (06), 1-14 (2016).
- Gnaiger, E., Lassnig, B., Kuznetsov, A., Rieger, G., Margreiter, R. Excess capacity of cytochrome c oxidase. Journal of Experimental Biology. 1139, 1129-1139 (1998).
- Gnaiger, E., et al. Mitochondria in the Cold. Life in the Cold. , 431-442 (2000).
- Fontana-Ayoub, M., Fasching, E., Gnaiger, Selected media and chemicals for respirometry with mitochondrial preparations. Mitochondrial Physiology Network. 02 (17), 1-9 (2014).
- Gerencser, A. A., et al. Quantitative microplate-based respirometry with correction for oxygen diffusion. Analytical Chemistry. 81 (16), 6868-6878 (2009).
- Krumschnabel, G., Eigentler, A., Fasching, M., Gnaiger, E. Use of safranin for the assessment of mitochondrial membrane potential by high-resolution respirometry and fluorometry. Methods in Enzymology. 542, 163-181 (2014).
- Nászai, A., Terhes, E., Kaszaki, J., Boros, M., Juhász, L. Ca(2+)N it be measured? Detection of extramitochondrial calcium movement with high-resolution fluorespirometry. Scientific Reports. 9 (1), 1-13 (2019).
- Pajak, B., et al. 2-Deoxy-d-Glucose and its analogs: From diagnostic to therapeutic agents. International Journal of Molecular Sciences. 21 (1), 234 (2019).
- Mercier-Letondal, P., Marton, C., Godet, Y., Galaine, J. Validation of a method evaluating T cell metabolic potential in compliance with ICH Q2 (R1). Journal of Translational Medicine. 19 (1), 1-15 (2021).
- Sauerbeck, A., et al. Analysis of regional brain mitochondrial bioenergetics and susceptibility to mitochondrial inhibition utilizing a microplate based system. Journal of Neuroscience Methods. 198 (1), 36-43 (2011).
- Jackman, M. R., Willis, W. T. Characteristics of mitochondria isolated from type I and type IIb skeletal muscle. American Journal of Physiology - Cell Physiology. 270 (2), 673-678 (1996).
- Ponsot, E., et al. Mitochondrial tissue specificity of substrates utilization in rat cardiac and skeletal muscles. Journal of Cellular Physiology. 203 (3), 479-486 (2005).
- Schönfeld, P., Reiser, G. Why does brain metabolism not favor burning of fatty acids to provide energy-Reflections on disadvantages of the use of free fatty acids as fuel for brain. Journal of Cerebral Blood Flow and Metabolism. 33 (10), 1493-1499 (2013).
- Calderon-Dominguez, M., Mir, J. F., Fucho, R., Weber, M., Serra, D., Herrero, L. Fatty acid metabolism and the basis of brown adipose tissue function. Adipocyte. 5 (2), 98-118 (2016).
- Divakaruni, A. S., Rogers, G. W., Murphy, A. N. Measuring mitochondrial function in permeabilized cells using the seahorse XF analyzer or a clark-type oxygen electrode. Current Protocols in Toxicology. 2014, 1-16 (2014).
- Iuso, A., Repp, B., Biagosch, C., Terrile, C., Prokisch, H. Assessing mitochondrial bioenergetics in isolated mitochondria from various mouse tissues using Seahorse XF96 analyzer. Methods in Molecular Biology. 1567, 217-230 (2017).
- Rogers, G. W., et al. High throughput microplate respiratory measurements using minimal quantities of isolated mitochondria. PLoS ONE. 6 (7), 21746 (2011).
- Jordá, A., Zaragozá, R., Portolés, M., Báguena-Cervellera, R., Renau-Piqueras, J. Long-term high-protein diet induces biochemical and ultrastructural changes in rat liver mitochondria. Archives of Biochemistry and Biophysics. 265 (2), 241-248 (1988).
- Jackson, C. B., Gallati, S., Schaller, A. QPCR-based mitochondrial DNA quantification: Influence of template DNA fragmentation on accuracy. Biochemical and Biophysical Research Communications. 423 (3), 441-447 (2012).
- Hirsch, H. M. Tissue autoxidation inhibitors: II. The presence of inhibitor in intact cells; Assay of liver and hepatoma effect on radio-oxidations. Cancer Research. 16 (11), 1076-1082 (1956).
- Picard, M., et al. Mitochondrial structure and function are disrupted by standard Isolation methods. PLoS ONE. 6 (3), 18317 (2011).
- Tanumihardja, E., Slaats, R. H., Van Der Meer, A. D., Passier, R., Olthuis, W., Van Den Berg, A. Measuring both pH and O2 with a single On-Chip sensor in cultures of human pluripotent stem cell-derived cardiomyocytes to track induced changes in cellular metabolism. ACS Sensors. 6 (1), 267-274 (2021).
- Harms, F., Stolker, R. J., Mik, E. Cutaneous respirometry as novel technique to monitor mitochondrial function: A feasibility study in healthy volunteers. PLoS ONE. 11 (7), 159544 (2016).
- Levitsky, Y., et al. Micro-respirometry of whole cells and isolated mitochondria. RSC Advances. 9 (57), 33257-33267 (2019).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados