JoVE 비디오를 활용하시려면 도서관을 통한 기관 구독이 필요합니다. 전체 비디오를 보시려면 로그인하거나 무료 트라이얼을 시작하세요.
Method Article
챔버 및 플레이트 기반 호흡계를 사용하여 세포 및 조직의 생물 에너지를 평가하기위한 고해상도 호흡 측정법
요약
고분해능 호흡계를 사용하여 산화 인산화를 평가하는 것은 미토콘드리아와 세포 에너지 대사의 기능 분석의 필수적인 부분이되었습니다. 여기에서는 챔버 및 마이크로 플레이트 기반 고해상도 호흡계를 사용하여 세포 에너지 대사를 분석하기위한 프로토콜을 제시하고 각 장치의 주요 이점에 대해 논의합니다.
초록
고분해능 호흡측정법(HRR)을 통해 개별 세포 에너지 상태 분석 및 다양한 기질-언커플러-억제제 적정(SUIT) 프로토콜을 사용하여 호흡기 복합체를 평가하기 위해 산화적 인산화를 실시간으로 모니터링할 수 있습니다. 여기에서는 두 개의 고해상도 호흡 측정 장치의 용도가 입증되며, 배양 된 세포, 골격 및 심장 근육 섬유, 뇌 및 간과 같은 연조직의 분석에 적용 할 수있는 프로토콜의 기본 모음이 제시됩니다. 배양된 세포 및 조직을 위한 프로토콜은 표준 호흡 프로토콜을 포괄하는 마이크로플레이트 기반 호흡계를 위한 챔버 기반 호흡계 및 배양된 세포를 위해 제공된다. 비교 목적을 위해, 다수의 호흡기 시스템 결핍을 야기하는 미토콘드리아 번역이 결핍된 CRISPR 조작된 HEK293 세포는 호흡에서 세포 결함을 입증하기 위해 두 장치와 함께 사용된다. 두 호흡계는 연구중인 연구 질문 및 모델에 따라 각각의 기술적 장점과 적합성으로 세포 호흡을 포괄적으로 측정 할 수 있습니다.
서문
미토콘드리아는 에너지의 주요 공급을 충족하고 뉴클레오티드, 지질 및 아미노산의 동화 작용, 철 - 황 클러스터 생물 발생과 같은 필수 세포 생물 에너지 및 대사 과정에 기여하는 구획화 된 소기관이며 통제 된 세포 사멸과 같은 신호 전달에 연루되어 있습니다 1,2,3 . 산화적 인산화를 통한 미토콘드리아 생물에너지학은 세포 내의 거의 모든 세포 과정에 기여하며, 결과적으로, 일차 또는 이차 기원의 미토콘드리아 기능장애는 광범위한 질병 상태 4,5와 연관된다. 미토콘드리아 기능 장애는 구조 또는 미토콘드리아 밀도의 변화뿐만 아니라 호흡기의 질과 조절을 포함합니다6. 이 정성적 요소는 기질 제어, 결합 특성, 번역 후 변형, 크리스타 역학 및 호흡 초복합체 7,8을 포함합니다. 따라서, 세포의 에너지 대사를 평가하기 위한 실험 및 진단 접근법을 위한 미토콘드리아 생물에너지학의 정확한 분석은 건강 및 질병에서 중요하다.
미토콘드리아 산화 인산화 (OXPHOS)는 아데노신 삼인산염 (ATP)9을 통한 세포 에너지의 생성을위한 호흡계 또는 전자 전달 시스템 (ETS) 내의 일련의 반응입니다. 복합체 I 및 II를 통한 전자 흐름으로부터 콤플렉스 IV로의 에너지를 이용하는 다중 효소적 단계는 내부 미토콘드리아 막을 가로질러 전기화학적 양성자 구배를 생성하고, 이어서 복합 V(F1FO ATP 합성효소)를 통해 ATP로의 아데노신 디포스페이트(ADP)의 인산화에 이용된다(도 1A).
먼저, 2개의 전자 담체는 트리카르복실 사이클(TCA), 당분해 및 피루베이트 산화 동안 생성된다: 니코틴아미드 아데닌 디뉴클레오타이드(NADH) 및 디히드로플라빈 아데닌 디뉴클레오타이드(FADH2). NADH는 콤플렉스 I (NADH 탈수소 효소)에서 산화되며, 그 동안 두 개의 전자가 코엔자임 Q로 전달되고 (퀴논은 퀴놀로 환원됨), 양성자는 막간 공간 (IMS)으로 펌핑됩니다. 둘째, 착물 II (숙시네이트 탈수소효소)는 FADH2를 산화시키고 양성자를 펌핑하지 않고 전자를 코엔자임 Q에 공급한다. 셋째, 콤플렉스 III (시토크롬 c 산화환원효소)에서, 코엔자임 Q로부터의 전자는 시토크롬 c로 전달되고, 양성자는 IMS로 펌핑된다. 넷째, 시토크롬 c는 전자를 양성자를 펌핑하는 최종 복합체인 콤플렉스 IV(Cytochrome c oxidase)로 전달하고, 여기서 산소는 양성자를 동화시키는 전자 수용체로서 기능하여 궁극적으로 물을 형성한다. 미토콘드리아가 소비하는 것은 이 산소이며, 이는 옥시그래프로 측정될 수 있다. 마지막으로, 콤플렉스 I, 콤플렉스 III, 및 콤플렉스 IV로부터 생성된 양성자는 콤플렉스 V를 회전시키는데 사용되어, ATP9를 생성한다.
중요하게도, 전자 전달은 선형 방식으로 발생할뿐만 아니라, 그렇지 않으면 전자 수송 사슬로 표시된다. 대신, 전자는 다수의 호흡 경로를 통해 코엔자임 Q 풀로 전달될 수 있고 수렴 전자 흐름을 촉진할 수 있다. NADH-기질 및 숙시네이트는 예를 들어, 각각 콤플렉스 I 및 콤플렉스 II를 통해 들어갈 수 있다. 지방산 산화로부터의 전자는 전자 전달 플라보단백질 복합체를 통해 공여될 수 있다. 실제로 OXPHOS를 포괄적으로 분석하려면 적절한 연료 기판을 사용한 전체적인 접근 방식이 필요합니다(그림 1A).

도 1: 미토콘드리아 산화 인산화 및 특정 기질 및 저해제 프로토콜. (A) 미토콘드리온 및 전자 전달 시스템 (CI-CIV) 및 미토콘드리아F1F0ATP 합성 효소 (CV)의 계획. 모든 구조물은 PDB로부터이다. 도면은 본 연구에 기재된 기질 및 억제제만을 묘사한다). (b) mHRR 장치에서 표준 프로토콜을 사용하는 무손상 HEK293 세포에서 산소 플럭스의 샘플 트레이스. (c) cHRR 장치에서 표준 프로토콜을 사용하는 무손상 HEK293 세포에서 산소 플럭스의 샘플 트레이스. (d) 각각의 SUIT 프로토콜을 갖는 건강한 공여자로부터의 투과화된 인간 섬유아세포에서의 산소 플럭스의 샘플 트레이스. 약어: 1 = 손상되지 않은 세포의 일상적인 호흡; 2 = 상태 2; 3 = 상태 3(I); 4 = cytC가 있는 상태 3(I); 5 = 상태 3 (I+II); 6 = 누출(OM); 7 = ETS 용량; 8 = S(ROT); 9 = ROX; 10 = TMPD; 11 = Az. ROT = 로테논, AM = 안티마이신, ATP = 아데노신 트리포스페이트, Az = 아지드, OM = 올리고마이신, FCCP = 카르보닐 시아나이드 파라트리플루오로메톡시페닐히드라존; Asc = 아스코르베이트, TMPD = N,N,N',N'-테트라메틸-p-페닐렌디아민, Succ = 숙시네이트, M = 말레이트, P = 피루베이트, ADP = 아데노신 디포스페이트, NAD = 니코틴아미드 아데닌 디뉴클레오티드, IMS = 막간 공간, FAD = 플라빈 아데닌 디뉴클레오티드. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
HRR을 이용한 미토콘드리아 OXPHOS 용량 분석은 1차 미토콘드리아 결함10,11뿐만 아니라 암 및 노화 12와 같은 생물학의 다른 모든 영역으로 확장되는 진단 가치의 도구 생화학 적 방법이되었습니다. HRR은 개별 또는 복합 미토콘드리아 호흡 복합체 결핍을 직접 반영하고 간접적으로 세포 기능 장애 및 변경된 에너지 대사와 관련이있는 미토콘드리아 OXPHOS 용량의 분석에 의해 세포 호흡의 결정을 허용합니다9. 방법론적으로, 호흡 측정은 세포, 조직 또는 단리된 미토콘드리아(11,13,14)를 사용하여 수행되며, 동결된 물질은 부분적으로만 적합한 물질(15,16)을 사용한다. 동결 조직은 초복합체 안정성을 유지하는 무손상 ETS를 갖는 것으로 나타났다(15). 따라서, 전통적인 TCA 중간체와는 달리, 각각의 기질은 ETS로 직접 공급된다. 그러나, ETS와 ATP 합성 사이의 결합은 동결 손상(얼음 결정 형성)을 통해 막 완전성이 손상됨에 따라 손실된다.
호흡 실험은 일반적으로 비투과화 또는 투과화된 세포 또는 조직에서 흡열에 대해 37°C의 생리학적 온도에서 일어난다. 전자는 세포질 대사 맥락을 고려하는 반면, 후자는 특정 기질 (및 억제제)의 첨가를 통해 개별 OXPHOS 복합체 및 ATPase의 에너지 적 기여를 제공한다. 기질 및 억제제의 서열 및 변이는 OXPHOS 기능의 다양한 과학적 질문을 다루기 위해 SUIT 프로토콜17 및 분석 18의 다양한 어레이의 개발로 이어졌다 (12에서 검토). 세포 호흡의 기본 프로토콜은 네 가지 다른 상태를 평가합니다 : i) 일상적인 호흡 - 기질이나 억제제를 첨가하지 않고 각각의 호흡 매체에서 호흡하지만 내인성 기질을 소비합니다. 이러한 상태는 예를 들어, 변경된 대사산물 프로파일에 의해 야기된 일반적인 OXPHOS 또는 이차-유도된 호흡 결함을 나타낼 수 있다. 다음으로, ATPase 억제제인 올리고마이신의 첨가는 ii) 누출 호흡으로 정의되는 양성자에 대한 미토콘드리아 내부 막의 투과성을 드러낸다. 언커플러 카르보닐 시아나이드 p-트리플루오로메톡시페닐히드라존(FCCP)과 같은 프로토노포어의 후속 적정은 iii) 결합되지 않은 호흡으로 정의되는 개방 막횡단 양성자 회로 모드에서 ETS 용량이 최대인 상태를 결정할 수 있게 한다. 중요하게도, 결합되지 않은 상태는 또한 미토콘드리아 막에 대한 과도한 기계적 손상을 통한 실험적 개입에 의해 발생할 수 있다. 반대로, 비결합 상태는 생리학적으로 조절되는 내재적 메카니즘에 의해 호흡 언커플링을 지칭한다. 마지막으로, 콤플렉스 III 억제제인 안티마이신 및 콤플렉스 I 억제제 로테논을 첨가함으로써 ETS의 완전한 억제는 비미토콘드리아 산소 소모 공정으로부터의 잔류 산소 소비량(ROX)을 결정한다(도 1A-C).
미토콘드리아 생물에너지학은 다섯 개의 뚜렷한 호흡 상태(19,20)로 구성된다. 상태 1 호흡은 내인성으로 이용가능한 것을 제외하고는 어떠한 추가적인 기질 또는 ADP도 없다. ADP를 첨가한 후에도, 여전히 기질이 없으나, 상태 2 호흡이 달성된다. 기판이 추가되어 전자 전달 및 ATP 합성을 허용하면 상태 3 호흡에 도달합니다. 이 상태에서, OXPHOS 용량은 ADP, 무기 포스페이트, 산소, NADH- 및 숙시네이트-연결된 기질의 포화 농도에서 정의될 수 있다. 상태 4 호흡 또는 LEAK 호흡은 충분한 기질을 가지면서 ADP 또는 화학적으로 억제된 ATP 합성효소가 없는 상태로 정의될 수 있다. 마지막으로, 닫힌 챔버 설정에서 모든 산소가 고갈(anoxic)될 때, 상태 5 호흡이 관찰된다.
실험 모델 및 연구 질문에 따라 상이한 적용성을 갖는 폐쇄 챔버 시스템에서 시간에 따른 산소 감소의 함수로서 측정되는 산소 소비의 분석을 통해 OXPHOS의 현재 실시간 평가를 지배하는 두 개의 장치를 사용하여 세포 에너지 상태(14 )를 평가하기 위한 몇 가지 방법이 존재한다: Oroboros 2k 고분해능 호흡계와 Seahorse XF 세포외 플럭스 분석기. 두 장치 모두 챔버 또는 마이크로플레이트 웰 내의 절대값으로서 초당 산소(O2)의 피코몰(pmol)의 감소로서 산소 소모율을 기록한다. 질량당 특정 산소 소비량은 세포 수(백만), 조직 중량(mg), 또는 단백질 양당 특정 완충 레시피에서 각각의 산소 소비를 정상화함으로써 얻어진다.
O2k(Oroboros Instruments)는 편광 산소 센서(챔버 기반 고해상도 호흡계: cHRR)가 장착된 폐쇄형 2챔버 시스템입니다. 각 실험 챔버는 자기 교반기에 의해 균질하게 유지되는 2 mL의 액체를 보유한다. 편광 산소 센서는 산소를 측정하기 위해 양면계 접근 방식을 사용합니다 : 금 음극, 은/염화은 양극, KCI 용액 사이에 전압(0.8V)이 가해지는 전기화학 전지를 생성합니다. 분석 매질로부터의 산소는 25 μm 플루오르화 에틸렌 프로필렌 막 (O2-투과성)을 통해 확산되고 캐소드에서 환원을 겪고, 과산화수소를 생성한다. 양극에서 은은 과산화수소에 의해 산화되어 전류를 생성합니다. 이러한 전류(암페어)는 분위 산소 압력과 선형적으로 관련된다. 산소의 분압 및 분석 매질의 산소 용해도 인자는 산소 농도를 계산하는데 사용된다. 산소 분압은 실험 온도에 의존하고 편광 측정은 온도에 민감하기 때문에 온도 변동은 펠티에 가열 블록에 의한 정밀한(±0.002°C) 조절이 필요합니다. 온도는 4°C 및 47°C의 범위 내에서 제어될 수 있다.
Seahorse XF 세포외 플럭스 분석기(애질런트)는 24웰 또는 96웰 마이크로플레이트 포맷의 플레이트 기반 시스템으로, 세 개의 형광 전극이 각 웰에서 시간에 따른 산소 소비량을 측정합니다(마이크로플레이트 기반 고분해능 호흡계: mHRR). 분석 카트리지 내의 최대 네 개의 포트는 분석 동안 자동 주입을 위해 이용가능하다. 분석에는 여러 주기가 포함되며, 각 주기에는 1) 혼합, 2) 대기 및 3) 측정의 세 단계가 있습니다. 측정 단계 동안, 센서 프로브는 마이크로플레이트 내로 하강되어 방출된 광을 측정하기 위해 7-10 μL 부피를 포함하는 일시적으로 닫힌 챔버를 생성한다. 이 광은 센서 프로브의 팁에 폴리머 포매된 형광단에 의해 방출되며, 이는 인광 담금질에 기초하여O2 를 감지한다. 형광 신호의 강도는O2 에 비례하고 센서 및 분석 매질의 온도에 의해 영향을 받는다. 따라서, 정확한 산소 추정은 어떠한 샘플도 없이 배경 우물을 갖는 상대적인 접근법을 필요로 한다. 산소 농도 복원은 혼합 단계에서 센서가 임시 챔버 위의 부피를 혼합하기 위해 위아래로 움직일 때 발생합니다. 각 사이클은 하나의 산소 소모율을 계산합니다. 온도는 16°C 및 42°C의 범위 내에서 제어될 수 있다.
HRR은 원발성 및 미토콘드리아 관련 질환 및 일반적인 세포 대사에서 세포 생물 에너지를 평가하는 황금 표준입니다. 이 연구에서는 세포 및 조직에서 OXPHOS 기능을 평가하기 위해 HRR에 대한 기본 프로토콜이 제공됩니다.

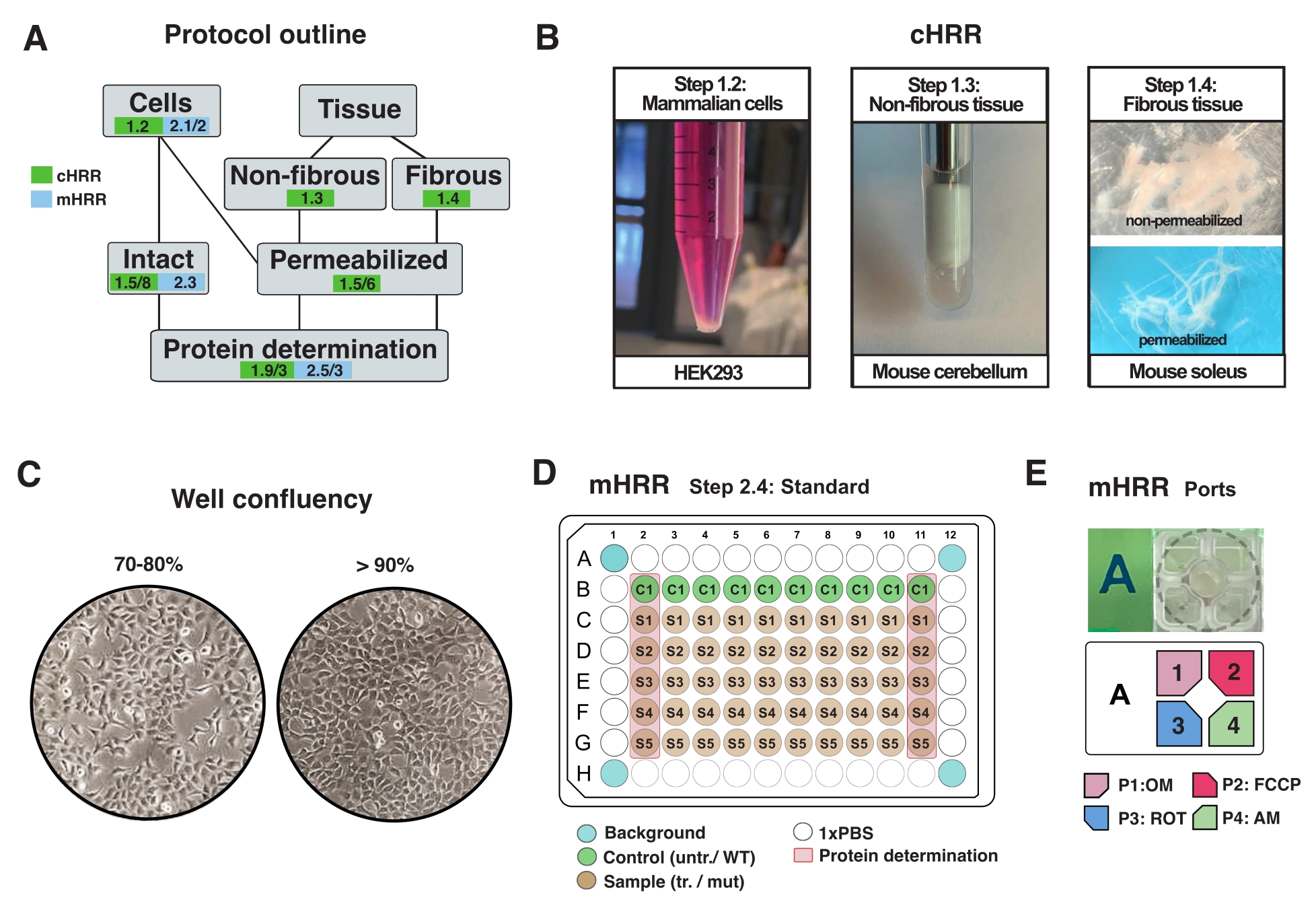
그림 2: cHRR을 위한 세포 및 조직 제제를 위한 워크플로우 및 mHRR 호흡측정을 위한 세포 준비 . (A) 제공된 프로토콜의 개요. (b) 포유동물 세포 (단계 1.2): 3 x 106 세포와 동등한 HEK293 펠릿 (왼쪽 패널). 비섬유성 조직 (단계 1.3): 2 mL 테프론 포터 (중간 패널) 중의 뮤린 소뇌 용해물의 제조. 사포닌-유도된 골격근 투과 (단계 1.4) cHRR 호흡측정을 위한 우측 패널. (c) mHRR 호흡측정법을 위한 표준 마이크로플레이트 시딩 레이아웃(단계 2.4) 및 진핵 세포(HEK293)의 분석을 위한 컨플루언시 확인. (D, E) mHRR 호흡 측정을 위한 주입 포트 로딩 방식(단계 2.4). 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
프로토콜
모든 동물 실험은 핀란드 남부의 국가 동물 실험 검토위원회 및 지역 국가 행정 기관에 따라 수행됩니다. 수컷 C57BL/6JOlaHsd 마우스(4-6개월령)를 본 연구에 사용하였다. 인간 세포주의 사용에 대한 동의는 헬싱키 대학의 제도적 윤리위원회로부터 입수하였다.
1. 고해상도 호흡 측정법 : 챔버 기반 호흡계 (cHRR)
참고: 프로토콜의 이 섹션에 있는 실험은 Oroboros O2k-Core: Oxygraph-2k(Table of Materials)를 사용하여 수행되었습니다.
- 산소 센서의 교정
- 2.1 mL의 미토콘드리아 호흡 배지 (MiR05, 표 1, 용해도 계수: 0.92)에서 37°C에서 >45분 동안 호흡계를 사전 실행하고21에 기재된 바와 같이 산소 교정을 수행한다. 기준선 변동이 ± 4pmol/s 이내인 경우 진행하십시오.
참고: 배경 신호의 큰 변동은 센서 멤브레인의 유지 보수가 필요하거나 이전 실험에서 챔버에 남아있는 억제제의 흔적을 나타낼 수 있습니다. 도구 배경 산소 플럭스 보정은 실험25의 배치 이전에 권장된다. - 산소 보정 값을 기록하여 시간 경과에 따른 센서 멤브레인 성능을 모니터링합니다.
참고: 이는 센서 기능, 신호 대 잡음 안정성 및 센서 멤브레인 유지 보수가 필요한 경우를 나타냅니다. 주위 압력에 따라 180-200 μmol의 산소가 MiR05에서 가용화됩니다. - 호흡 매체에 샘플을 첨가하기 전에 챔버 내의 모든 액체를 제거하십시오.
참고: 호흡실의 부피를 정확히 2mL로 정기적으로 평가하십시오.
- 2.1 mL의 미토콘드리아 호흡 배지 (MiR05, 표 1, 용해도 계수: 0.92)에서 37°C에서 >45분 동안 호흡계를 사전 실행하고21에 기재된 바와 같이 산소 교정을 수행한다. 기준선 변동이 ± 4pmol/s 이내인 경우 진행하십시오.
- 고해상도 호흡 측정을 위한 세포의 제조
- HEK293 세포를 10% 열 불활성화된 태아 소 혈청(FBS), 글루타맥스, 비필수 아미노산, Na-피루베이트22 및 우리딘23으로 보충된 고포도당을 갖는 둘베코 변형 이글 배지(DMEM)에서 직경 10cm 2의 접시에 배양하여 5%CO2의 37°C에서 OXPHOS 결함 대사를 지원한다.
참고: 모든 유형의 진핵 세포를 배양할 수 있습니다. 대부분의 세포 유형에 대해, 10cm2 디쉬를 배양하면 충분한 세포 (보통 >3 x 106 세포)가 생깁니다. 세포 신진 대사와 호흡에 영향을 미치지 않도록 마이코 플라스마 감염을 정기적으로 확인하십시오. - 90% 컨플루언시를 초과하지 않고 세포를 성장시킨다(그림 2C).
참고: >90% 플루언시를 가진 세포는 호흡에 대한 성장 의존적 억제 효과를 나타낼 수 있습니다(동기화되지 않은 경우 또는 유사분열 후). - 세포를 1x PBS로 세척하고, 따뜻한 0.25% 트립신 1 mL로 분리하고, 따뜻한 DMEM (5 mL/10cm2 플레이트)을 첨가하여 트립신을 비활성화하고, 혈구측정기로 세포를 계수한다.
- 300 x g에서 2.5 x 10 6 세포와 동일한 세포 용액을 5분 동안 부드럽게 원심분리하고, 상층액을 완전히 제거한 다음, 2.5 mL의 따뜻한 MiR05(1 x 106 cells/mL)에 재현탁시킨다(그림 2A).
- 현탁액 세포의 경우, 2.5 x 106 세포와 동등한 용액을 카운트 및 제거하고, 펠릿을 제거하고 단계 1.2.4에서 언급된 바와 같이 계속한다.
- 투과화 최적화를 위한 SUIT 프로토콜 실행(단계 1.6), 투과화된 세포 또는 조직(단계 1.5), 또는 무손상 세포(단계 1.7)
참고: 일관된 결과를 얻으려면 세포 농도를 일정하게 유지하는 것이 좋습니다(예: 1 x 106 cells/mL). 호흡은 호흡계(24)에서의 세포 밀도와 무관하지만, 세포 수가 일정하게 유지된다면 기질 및 억제제는 실험 전반에 걸쳐 유사한 농도에 있다.
- HEK293 세포를 10% 열 불활성화된 태아 소 혈청(FBS), 글루타맥스, 비필수 아미노산, Na-피루베이트22 및 우리딘23으로 보충된 고포도당을 갖는 둘베코 변형 이글 배지(DMEM)에서 직경 10cm 2의 접시에 배양하여 5%CO2의 37°C에서 OXPHOS 결함 대사를 지원한다.
- 고분해능 호흡 측정을 위한 비섬유성 조직(예: 뇌, 간)의 제조
- 중량 30-40mg의 균질 한 조직을 소비하거나 전체 장기 (이 경우 마우스 소뇌)를 사용하십시오.
참고: 조직을 즉시 사용하지 않는 경우, 얼음처럼 차가운 MiR05 2mL에 보관하여 대부분의 조직에 대해 최대 2시간 동안 보존할 수 있습니다. 개별 조직 저장 시간은 시계열로 평가되어야 합니다. - Whatman 여과지로 건조한 조직을 블롯하십시오 (조심하십시오 : 연조직 물질이 달라 붙는 경향이 있음).
- 30-40 mg 조직 조각을 빙냉된 2 mL 폴리테트라플루오로포터 엘베젬 호모게나이저에 넣는다.
- 적절한 양의 MiR05를 첨가하여 조직 대 완충액 비율을 유지하기 위해 20 mg / mL를 얻으십시오. 적절한 기계적 투과를 위해 불충분하거나 과도한 유체를 피하기 위해 총량을 >1.5 mL 및 <2 mL로 유지하십시오.
- 유모차를 삽입하고 과도한 조직 손상을 일으키는 진공의 생성을 피하면서 조심스럽게 유모차를 수축시켜 조직을 천천히 용해시킵니다.
- 용해될 때까지(큰 파편이 없는 탁한 액체로 나타남) 총 7번의 스트로크(1x 상하 스트로크로 정의됨)를 수행합니다(그림 2B).
참고: 적절한 용해를 위한 뇌졸중의 수는 시토크롬 C 반응을 통해 외부 미토콘드리아 막 완전성을 평가함으로써 각 조직에 대해 시험될 필요가 있다(단계 1.5.11). 분해하기 어려운 결합 조직 또는 용기 부분이 남아있을 수 있습니다. - 용해된 조직을 15 mL 원심분리 튜브에 데칸트한다.
- 용해 단계에서 사용되는 동량의 MiR05 (예를 들어, 1.5 mL)로 포터의 내부를 세척하고 10 mg / mL 조직 용해물에서 MiR05 3-4 mL를 함유하는 15 mL 튜브에 첨가하십시오.
- 챔버 당 2mL의 일반 MiR05를 첨가하여 37°C로 가온한다.
- 챔버 당 500 μL (5 mg과 같음)의 각 용해물을 피펫팅하기 전에 동일한 분포를 위해 튜브를 천천히 소용돌이 치며 37 °C까지 추위로부터의 응력을 최소화하십시오.
- 챔버를 닫기 전에 챔버 내용물이 37°C로 가온될 때까지 >3분 동안 기다리십시오. 마개 위에 과도한 유체를 제거하십시오 (닫은 후 챔버 당 양 : 4mg).
- 표준 투과에 대해 SUIT 프로토콜을 실행합니다(1.5단계).
- 중량 30-40mg의 균질 한 조직을 소비하거나 전체 장기 (이 경우 마우스 소뇌)를 사용하십시오.
- 고해상도 호흡측정을 위한 섬유조직(골격근, 심장근육)의 제조
- 해부 현미경으로 얼음처럼 차가운 BIOPS 2 mL의 날카로운 포셉을 사용하여 경질 조직을 추출하고, 결합 조직 및 지방을 근육으로부터 제거하였다(표 2).
- 날카로운 포셉으로 세로 축을 따라 섬유 다발 (~ 4mg)을 분리하십시오. 메쉬형 구조를 얻기 위해 섬유를 충분히 떼어냅니다(그림 2B).
참고 : 적절한 기계적 섬유 분리 및 투과는 적색 안료 미오글로빈의 손실과 반투명도 증가로 나타납니다. - 섬유 다발을 4°C에서 20분 동안 사포닌(BIOPS에서 50μg/mL, 신선하게 제조됨)으로 세척하고 투과시킨다(섬유는 반투명이 되어 완전한 투과를 나타냄, 도 2B).
- 섬유를 4°C에서 세척당 5분 동안 MiR05로 두 번 세척한다.
- 여과지로 블롯 건조시키고 2.1 mL MiR05로 채워진 챔버에 첨가하기 전에 무게를 재십시오.
- 완전히 닫지 않고 마개를 도입 한 다음 20 mL 주사기를 사용하여 2 mL의 순수한O2 로 챔버에 산소를 공급하고 마개를 회전 운동으로 비틀어 챔버를 닫습니다. 산소 확산 제한을 피하기 위해 실험 동안O2 농도를 300-500 μM 사이로 유지하십시오.
- 세포 또는 조직의 일상적인 호흡을 평가하기위한 프로토콜
- 단계 1.5.2-1.5.3에서 언급한 바와 같이 샘플을 챔버에 첨가한다.
- 따뜻한 MiR05 세포 현탁액 2.3mL를 첨가하십시오 (표준 입력: 단계 1.2에서와 같이 1 x 106 세포/mL 또는 단계 1.3에서와 같이 조직/mL의 2 mg)
- 골격근 및 심장 근육 (단계 1.4) : 1.4.4-1.4.6 단계를 고려하여 따뜻한 MiR05 2.3 mL에 사포닌 투과 섬유를 ~ 4mg 첨가하십시오.
- 챔버를 37°C에서 실행하고 700 rpm의 교반 속도로 실행한다. 매체가 탈기되도록 >3 분 동안 기다렸다가 회전하는 동작으로 스토퍼를 비틀어 챔버를 닫으십시오. 펠티에 블록 안정화는 설정 온도에 도달했음을 나타낸다.
- (선택 사항) 교반기 속도를 300 rpm으로 변경하여 나머지 기포가 마개의 모세관을 통해 빠져나갈 수 있도록 하십시오.
- 마개 위에 과도한 액체를 흡인하십시오. 모든 샘플 유형에서 안정적인 산소 플럭스 신호가 달성되어 루틴/상태 1 호흡을 기록할 때까지 10분 동안 기다립니다( 그림 1B).
- 투과된 세포 및 조직에서의 호흡 측정을 위해, 단계 1.6으로 계속하십시오. 단계 1.8을 갖는 온전한 세포의 경우.
- 투과된 세포 또는 조직에서의 OXPHOS 분석을 위한 프로토콜
- 용해된(permeabilized) 조직 샘플을 사용하거나 세포를 투과시키기 위해 5 μg/mL의 최종 농도에 대해 1 μL의 디지토닌 (디메틸설폭사이드(DMSO)의 8.1 mM 디지토닌 스톡)을 첨가하여 세포를 투과시킨다. 플럭스는 떨어지고 >5 분에 안정화되어야합니다.
주의: 디지토닌은 호흡기에 심각한 독성을 갖거나, 피부에 닿거나, 삼킬 때 독성이 있습니다.
참고 : 모든 화학 물질의 주입은 정밀 유리 주사기로 수행됩니다. 교차 오염을 피하기 위해 표시된 화학 물질에만 주사기를 사용하고 사용 후 물과 EtOH로 철저히 씻으십시오. 막힌 주사기는 따뜻한ddH2O또는 화학 막힘을 제거하기 위해 세척 와이어에서 초음파가 필요할 수 있습니다. 챔버에 공기가 유입되지 않도록 항상 각 원액의 잉여를 주사기로 철회하십시오. 챔버 내부를 검사하여 각 주사 후 공기가 유입되는지 검사하십시오. 플럭스가 고원화될 때까지 각 단계를 기록하십시오. - 5 μL의 최종 농도에 대해 0.4 M 말레이트(M)를 1 mM, 5 μL의 2.0 M 피루베이트(P; 갓 제조), 최종 농도 5 mM, 4 μL의 2.5 M 글루타메이트(G)를 최종 농도 5 mM에 대해 첨가한다.
- 이전의 플럭스를 고원화한 후, 1.25 mM의 최종 농도를 위해 0.5 M 아데노신 디포스페이트 (ADP, -80°C에서 저장된 분취량)의 5 μL (근육 조직에 대해 10 μL)를 첨가한다.
참고: 근육과 같은 조직은 포화에 도달하기 위해 다른 농도가 필요할 수 있습니다. - 5 μL의 4 mM 시토크롬 C (cytC)를 10 μM의 최종 농도에 첨가한다.
참고: 세포가 투과의 품질을 평가하는 것은 선택 사항입니다. - 10 mM의 최종 농도에 대해 1.25 M 숙시네이트(S)의 16 μL를 첨가한다. (선택 사항) ADP 농도의 포화를 조절하기 위해 2 mM의 최종 농도에 대해 3 μL의 0.5 M ADP를 첨가한다.
- 세포 및 비섬유성 조직의 경우, 1 mg/mL 올리고마이신(OM) 2 μL를 첨가하여 최종 농도 1 μg/mL를 섭취하십시오.
주의: 사용된 모든 ETS 억제제는 독성이 강합니다.
참고 : Oligomycin은 ETS 용량을 억제 할 수 있고 근육 조직에 대해 생략 될 수 있으므로 최적의 농도를 위해 적정이 필요할 수 있습니다. 근육 조직이 분석될 때 여기에서 재산소화되고O2 가 300 μM 미만이면 여기에서 재산소화된다. - FCCP를 2 mM 스톡으로부터 적정하고, 호흡 및 호흡의 증가가 최대로 결합되지 않을 때까지 후속 0.2 μL 단계와 함께 0.6 μL를 첨가한다(이론적: 비결합).
- 최종 농도 0.5 μM에 대해 1 mM 로테논(ROT) 1 μL를 첨가하십시오. 최종 농도 1 μg/mL에 대해 1 mg/mL 안티마이신(AM) 스톡 2 μL를 첨가하십시오.
- 챔버를 재산소화하여 플런저를 비틀림 동작으로 천천히 들어 올려 모든 챔버에서 유사한 산소 수준(~150 μM)을 달성한다.
- 5 μL의 0.8 M 아스코르베이트를 2 mM의 최종 농도에 대해 즉시 첨가한 다음, 0.2 mM의 0.2 M N,N,N',N'-테트라메틸-p-페닐렌디아민 (TMPD)을 5 μL로 첨가하여 복합체 IV 활성을 평가하였다 (선택적).
- TMPD로 피크O2 플럭스에 도달할 때 즉시 10 mM의 최종 농도에 대해 5 μL의 4 M 아지드를 첨가한다. 복합 IV 염기 수준 계산을 위해 TMPD의 자동 산화 검정을 하기 위해 >5분 동안 실행을 계속한다.
- 셀 카운트를 다시 계산하여 셀 카운트 사전 실행을 확인하고 1.9단계를 계속하십시오.
참고: 디지토닌-투과화(세포에만 해당)는 최대 플럭스에 도달하고 미토콘드리아 막 완전성에 영향을 미치지 않도록 시험 실험에서 적정해야 합니다(단계 1.7 참조). 시토크롬 c의 첨가 후 호흡률의 >10% 증가로 투과화된 샘플(특히 근육 조직)은 외부 미토콘드리아 막 손상으로 인한 추가 분석으로부터 배제되어야 한다. EtOH 용해 화학 물질의 첨가 후 플럭스에서 단시간 딥이 예상된다.
- 용해된(permeabilized) 조직 샘플을 사용하거나 세포를 투과시키기 위해 5 μg/mL의 최종 농도에 대해 1 μL의 디지토닌 (디메틸설폭사이드(DMSO)의 8.1 mM 디지토닌 스톡)을 첨가하여 세포를 투과시킨다. 플럭스는 떨어지고 >5 분에 안정화되어야합니다.
- 세포에 대한 최적의 투과 조건을 결정하는 프로토콜
- 단계 1.2 및 1.5.2에 설명된 대로 셀을 추가합니다.
- 10mg/mL 디지토닌 스톡 10μL를 취하여 DMSO 10μL를 첨가하여 5mg/mL로 희석합니다.
- 1 μL의 로테논 (1 mM 스톡)을 첨가한다. 10 μL의 숙시네이트 (2 mM 스톡) 및 5 μL의 ADP (0.5 M 스톡)를 첨가한다.
- 호흡이 더 이상 증가하지 않고 최대가 될 때까지 1 μL의 디지토닌 (단계 당 2.5 mg)을 반복적으로 적정하십시오.
참고: 호흡의 감소는 디지토닌의 과도한 농도를 나타냅니다.
- 무손상 세포에서 OXPHOS 분석을 위한 프로토콜
- 일상적인 호흡 후 (단계 1.6.1-1.6.6), 10 nM의 최종 농도에 대해 0.01 mM 올리고마이신 2 μL를 첨가한다.
- 2 mM 스톡으로부터 FCCP를 적정하고, 호흡 및 호흡의 추가 증가가 최대로 결합되지 않을 때까지 후속 0.2 μL 단계와 함께 0.6 μL를 첨가한다 (이론적: 비결합)
- 최종 농도 0.5 μM에 대해 1 mM 로테논 1 μL를 첨가하십시오. 최종 농도 1 μg/mL에 대해 1 mg/mL 안티마이신 스톡 2 μL를 첨가하십시오.
- 꼬이는 동작에서 플런저를 천천히 들어 올려 챔버를 동일한 산소 수준(~150 μM)으로 재산소화한다.
- 2 mM의 최종 농도를 위해 0.8 M 아스코르베이트 5 μL를 첨가한다. 복합 IV 활성을 평가하기 위해 0.5 mM의 최종 농도에 대해 5 μL의 0.2 M TMPD를 즉시 첨가한다.
참고 : TMPD가 자동 산화되기 쉽기 때문에 더 큰 실험 세트 전에 새로운 배치를 준비하십시오. 활동은 -20°C에서 저장될 때 시간이 지남에 따라 감소할 수 있다. - TMPD로 피크O2 플럭스에 도달할 때 즉시 10 mM의 최종 농도에 대해 5 μL의 4 M 아지드를 첨가한다. 복합 IV 염기 수준 계산을 위해 TMPD의 자동 산화 검정을 위해 >5분 동안 계속 실행한다.
- 셀 카운트를 재계산하여 셀 카운트가 사전 실행되었는지 확인하고 1.9단계를 계속합니다.
- 실행 후 샘플 수집
- 각 챔버(교반기가 켜져 있음)에서 MiR05 현탁액 정확히 1mL를 1.5mL 튜브 1.5mL 튜브에 수집합니다.
- 투과화된 세포의 경우 1000 x g에서 또는 조직 용해물의 경우 20,000 x g에서 원심분리한다. 상청액을 제거하고 추가 처리를 위해 펠렛을 -80°C에서 동결시킨다(섹션 3).
- SUIT 프로토콜 분석
- 기질 또는 억제제를 첨가한 후 각 고원에서 산소 플럭스(pmol/s, 입력으로 정규화됨)를 분석합니다(그림 1C 및 그림 3A). 값을 스프레드시트로 내보냅니다.
- 각 실험 실행의 모든 값에서 잔류 산소 소비량(ROX, 그림 1C 및 그림 3C) 값을 뺍니다. TMPD에서 아지드 잔류 호흡을 빼서 복잡한 IV 호흡을 얻습니다.
- 세포(그림 3A, B) 또는 조직 입력(그림 5A, B)에 대해 정규화된 절대값을 플로팅합니다. 플럭스 조절 비율을 계산하거나(단계 1.11) 단백질 입력으로 정규화합니다(그림 3C).
- 플럭스 제어비 계산
- 플럭스 제어 비율 (FCR) 9,26을 사용하여 호흡 기능 및 커플링 제어 지수를 획득하십시오.
참고: 이를 통해 미토콘드리아 양과 무관하게 내재적 미토콘드리아 품질을 평가할 수 있습니다. 또한, 플럭스 제어 비율 (FCR)은 시약 품질 관리를 허용하는 동일한 세포주 내에서 필적한다 (각각의 FCR은 도 1B-D 및 도 3C에 표시된 번호가 매겨진 기준 값을 통해 얻어진다). - 수학식 1을 사용하여 OXPHOS와 LEAK의 결합에 대한 호흡 조절 비율을 계산한다.
수학식 1: FCRADP = 5/6 = 상태 3 / 상태 4 - 방정식 2를 사용하여 NADH 의존성 호흡을 평가하기 위해 FCR을 계산하십시오.
수학식 2: FCR 상태 3(I) = 3/5 = 상태 3(I) /상태 3( I+II) - FCR을 계산하여 수학식 3을 사용하여 숙시네이트 의존성 호흡을 평가한다.
수학식 3: FCR상태 3(II) = 8/7 = S로트 /ETS용량 - FCR을 계산하여 수학식 4를 사용하여 결합되지 않은 결합에 대한 결합을 평가한다.
수학식 4: FCR 결합/비결합 = 5/7 = 상태 3(I+II)/ETS용량 - 미토콘드리아 외막 무결성을 테스트하려면 방정식 5를 사용하십시오.
수학식 5: % 미토콘드리아 외막 손상 = 3/4 = 상태 3 ( I) / 상태 3 (I)와 cyt c
- 플럭스 제어 비율 (FCR) 9,26을 사용하여 호흡 기능 및 커플링 제어 지수를 획득하십시오.
2. 고해상도 호흡 측정: 마이크로플레이트 기반 호흡계(mHRR)
참고 : 프로토콜의이 섹션의 실험은 Seahorse XFe96 세포외 플럭스 분석기 (표 자료)를 사용하여 수행되었습니다.
- 세포 배양
- 모든 종류의 세포를 배양하십시오. 부착제 (예를 들어, 콜라겐, 라미닌)는 세포 부착을 촉진하기 위해 사용될 수 있다. 여기서, HEK293 세포는 이전과 같이 배양된다(단계 1.3).
- 실험 전날, 세포를 분리하고 지정된 mHRR 96-웰 마이크로플레이트로 옮겨 실험 당일에 이상적인 컨플루언시를 얻었다(80%-100%)(도 2C).
참고: mHRR의 경우 마이크로플레이트 셀 밀도가 중요합니다. 성장에 영향을 미치는 세포주 또는 치료법의 개별적인 성장 특성은 실험 당일에 필적할만한 합류성을 보장하기 위해 설명될 필요가 있다.
- 고해상도 호흡 측정을 위한 세포의 제조
- 시딩 전에 세포를 충분히 수확하고 재현탁하십시오.
참고: 반복실험에 대해 동일한 희석액으로부터 세포를 시드하는 것이 좋습니다. - 처리중인 개별 세포주의 성장 속도 또는 성장 특성에 따라 세포를 시드한다.
참고: 표준 96웰 마이크로플레이트에서 최적화하고 세포 밀도를 96웰 분석 특이적 마이크로플레이트로 추정하십시오. 이 셋업에서, 7 x 104 HEK293 WT 세포를 96-웰의 웰당 시딩하였다. 96-웰 플레이트의 첫 번째 및 마지막 컬럼은 단백질 결정에 사용된다(도 2C). 네 모서리 웰에는 세포가 없어야하며 실험 배경 보정에 사용됩니다. 이상적으로는, 에지에 가까운 웰은 에지 효과를 최소화하기 위해 비어 있다(예를 들어, 세포는 온도 효과에 의해 야기된 변화된 성장 속도를 나타냄)(도 2C, D).
- 시딩 전에 세포를 충분히 수확하고 재현탁하십시오.
- 센서 플레이트의 제조, 억제제의 로딩
- 분석 당일, 배지 38.8 mL를 0.4 mL의 1 M 글루코스, 0.4 mL의 200 mM 글루타민 및 0.4 mL의 100 mM Na-Pyruvate로 보충한다.
참고: mHRR 호흡은 pH 7.4에서 전문화된 비완충된 DMEM 배지를 필요로 한다. 일반적으로 40mL는 하나의 96웰 마이크로플레이트를 사용한 한 번의 실험에 충분해야 합니다. - 호흡 분석 배지를 37°C로 가온하고, 웰 당 80 μL로 두 번 세척하여 세포 배양 배지를 호흡 분석 배지로 교환하였다.
- 세포를 검정 전에 60분 동안CO2 가 없는 37°C 인큐베이터에서 플레이트를 설정한다.
참고: 이 단계는CO2 가 호흡 결과에 영향을 미칠 수 있고 배지 내의 혈청이 분석 중에 기포를 생성할 수 있기 때문에 플레이트를 탈기시키는 데 필수적입니다. - OM, FCCP, ROT 및 AM에 대한 억제제 분취량을 37°C로 예열하고 센서 플레이트를 인큐베이터 밖으로 꺼낸다.
- OM, FCCP, ROT, 및 AM을 각각 1.5 μM, 1.125 μM, 및 1 μM의 최종 웰 농도로 3 mL의 분석 배지에 희석한다. 그림 2E에 표시된 대로 별도의 포트에 채우십시오.
주: 센서 카트리지를 채우려면 다중 채널 파이펫을 사용하는 것이 좋습니다. 가압 공기는 화합물을 주입하는 데 사용되기 때문에 포트가 화합물로 채워질 때마다 모든 포트는 동일한 양의 액체 부피로 채워져야합니다. ROT와 AM은 하나의 포트에서 결합할 수 있습니다. 억제제는 EtOH 또는 DMSO에 용해될 수 있다. - 주입 포트를 검사하고 각 포트에 대해 균일한 로딩 볼륨을 확인합니다.
참고: 모든 포트에는 하단에 주입용 구멍이 있습니다. 센서 플레이트를 이동할 때는주의해야합니다. 기포는 바늘을 사용하여 제거 할 수 있습니다.
- 분석 당일, 배지 38.8 mL를 0.4 mL의 1 M 글루코스, 0.4 mL의 200 mM 글루타민 및 0.4 mL의 100 mM Na-Pyruvate로 보충한다.
- 온전한 세포에서 산소 평가를 위한 프로토콜
- 분석 전날, 단계 2.4.2-2.4.7을 수행한다.
- 교정액 용액 20 mL를 분취량 50 mL 코니컬 튜브에 넣었다.
- 세포외 플럭스 분석 키트를 열고 내용물을 제거합니다.
- 센서 카트리지를 유틸리티 플레이트 옆에 반전 배치합니다. 200 μL의 교정 용액을 유틸리티 플레이트의 각 웰에 피펫한다.
- 센서 카트리지를 유틸리티 플레이트에 부착하여 모든 센서가 잠겼는지 확인합니다.
- 플레이트를 CO2가 없는 37°C 인큐베이터에 하룻밤 동안 또는 최소12 시간 동안 설정한다. 인큐베이터 내부의 습도가 교정제의 증발을 방지하기에 충분한지 확인하십시오.
- 마이크로플레이트 기반 시스템 및 컴퓨터를 켜서 다음날 사용할 수 있도록 준비한다(기계는 분석을 수행하기 전에 37°C로 평형화하기 위해 최소 3 h를 필요로 한다).
참고: 신호 안정성을 위해 호흡 상태당 3회의 측정 주기 대신 측정 지점을 6개로 늘립니다. 각 사이클은 3 분의 혼합과 3 분의 측정으로 구성됩니다. - XF 분석의 날에 단계 2.4.9-2.4.20을 수행한다.
- 세포 배양 플레이트의 합류성, 세포의 형태 및 배경 웰이 비어 있는지 확인하십시오.
- 단계 2.4.11-2.4.12에 언급된 바와 같이 제조된 호흡 배지로 세포를 세척한다.
- 20 μL를 제외한 모든 배지를 각 웰로부터 제거하였다. 배양 배지가 80 μL인 경우 55 μL를 밤새 증발시키기 때문에 제거하였다(대략 5 μL).
- 90 μL의 분석 배지로 세포를 두 번 세척한다. 마지막으로, 100 μL의 분석 배지를 첨가한다. 최종 부피는 120 μL이어야 한다.
참고: 이 단계에서는 각 실험 조건에 동일한 세척 절차가 적용되었는지 확인하기 위해 다중 채널 피펫을 사용하는 것이 좋습니다(플레이트 설정에 따라 다름). 흡인 할 때 플레이트를 45 ° 각도로 기울이고 피펫 팁을 웰 모서리에 배치하여 액체를 흡인 및 주입하십시오. 특정 세포가 세포 배양 플레이트의 바닥에서 쉽게 분리 될 수 있으므로 세척 중에주의해야합니다. - 플레이트를 검정 전에 60분 동안CO2 가 없는 37°C 인큐베이터에 설정한다.
- 수화된 센서 카트리지 플레이트를CO2-비함유 인큐베이터로부터 회수한다.
- 오래된 교정 용액을 버리고 37 °C로 예열된 신선한 교정 용액으로 교체하십시오.
- 억제제 및 분석 배지 (총 12 mL의 분석 배지에 대해 억제제 당 3 mL)를 준비하고 억제제 로딩을 위해 피펫 저장소를 사용하여 포트에 로딩하십시오.
- 소프트웨어를 열고 미리 디자인된 템플릿 또는 새 템플릿을 실행합니다. 플레이트 맵을 채우고 적정 및 측정 주기를 조정한 다음 시작 을 눌러 광 센서의 교정을 시작합니다.
- 로드된 카트리지에서 뚜껑을 분리하고 기계에서 자동으로 미끄러지는 슬롯에 놓고, 플레이트의 오른쪽 아래 모서리에 있는 표시가 슬롯의 오른쪽 아래 모서리에 있는 삼각형과 정렬되는지 확인합니다.
- 계속을 클릭하여 약 20분 동안 지속되는 자동 교정을 수행합니다.
- 교정 후 교정제가 포함된 유틸리티 플레이트를 제거합니다.
- 세포가 들어있는 마이크로플레이트에서 뚜껑을 분리하고 기계에 의해 메시지가 표시되면 플레이트를 슬롯에 놓습니다. 계속 을 클릭하여 실행을 시작하십시오.
- 실행 후 샘플 수집
- 플레이트를 기계 밖으로 꺼내고, 세포를 교란시키지 않고 남아있는 분석 배지를 조심스럽게 제거하고, 추가 처리를 위해 -80°C에서 전체 플레이트를 동결시킨다(섹션 3).
3. 비신코닌산 분석법(BCA assay)을 이용한 단백질 정량
- 단백질 추출에 사용되고 BCA와 호환되는 완충액에 희석된 소 혈청 알부민(BSA)을 준비하십시오: 2 mg/mL, 1.5 mg/mL, 1 mg/mL, 0.5 mg/mL, 0.25 mg/mL 및 0 mg/mL를 중복하여 표준 곡선을 준비하십시오.
- 추출 단백질은 cHRR용 1.5 mL 튜브 내에 포함된 mHRR 또는 펠릿 당 100 μL에 대해 웰당 20 μL의 적절한 용해 완충액 (예를 들어, RIPA)에 재현탁시킨다.
- 단백질 용해물을 함유하는 mHRR 플레이트 또는 1.5 mL 튜브를 얼음 상에서 30분 동안 인큐베이션한다.
- 단백질 용해물을 함유하는 1.5 mL 튜브를 20분 동안 20,000 x g 에서 4°C에서 20분 동안 원심분리하고, 생성된 상청액을 새로운 깨끗한 1.5 mL 튜브로 옮긴다.
- 샘플당 10 μL를 마이크로타이터 플레이트에서 중복 및 표준으로 사용하십시오. 200 μL의 BCA 작동 시약을 첨가하고, >15분 동안 인큐베이션한다.
- 플레이트를 파장 562 nm의 표준 분광 광도계에서 판독하고 BSA 표준 곡선을 사용하여 단백질 농도를 계산하십시오.
- 호흡 결과를 단백질 농도로 정상화하십시오.
참고: 단백질 양에 대한 정상화는 세포 시딩 밀도 또는 습윤 중량 입력을 확증할 수 있습니다. 추출된 단백질은 예를 들어 ETS의 서브유닛에 대한 후속 면역블롯팅에 적합하지만 천연 샘플을 완전히 나타내지는 않는다(예를 들어, 인산화 부위의 손실).
결과
여기서, 우리는 진핵 세포, 비섬유성 조직(예를 들어, 소뇌), 및 섬유성 조직(예를 들어, 골격근)에서 미토콘드리아 생물에너지를 결정하기 위한 프로토콜을 제공한다. 진핵 세포의 경우, 미토콘드리아 번역과 관련된 두 개의 상이한 단백질의 CRISPR 조작된 녹아웃을 갖는 HEK293을 다중 (CRISPR KO1) 및 심각한/완전한 OXPHOS 결핍 (CRISPRKO2)으로 cHRR (도 3A-C) 또는 mHRR (
토론
전통적으로 미토콘드리아 생물에너지학은 클락형 산소전극으로 연구되어 왔다. 그러나 해상도와 처리량의 부족은 기술 발전을 보장합니다. 현재까지 O2k (cHRR이라고 함) 및 Seahorse XF96 Flux Analyzer (mHRR이라고 함)는 세포 생물 에너지 분야에서 널리 채택되었습니다. 여기에서는 cHRR 또는 mHRR을 사용하여 미토콘드리아 호흡 평가를 통해 세포 에너지 대사를 분석하기위한 이해할 수있는 프로토콜 모음을...
공개
공개 할 이해 상충이 없습니다.
감사의 말
이 연구는 핀란드 아카데미 (C.B.J), Magnus Ehrnroot Foundation (C.B.J) 및 통합 생명 과학 대학원 (R.A.)의 박사 학위 펠로우십의 기금으로 지원되었습니다.
자료
| Name | Company | Catalog Number | Comments |
| 2 mL Potter-Elvehjem Glass/PTFE Tissue Grinder/Homogenizer | Omni International | 07-358029 | |
| 95% O2, 5% CO2 medical gas mixture | Potter for tissue grinding | ||
| ADP | Sigma | A 4386 | |
| Antimycin A | Sigma | A 8674 | Chemical |
| Ascorbate | Merck | PHR1279-1G | Chemical, dissolve in ethanol |
| BSA (fatty accid free) | Sigma | A 6003 | Chemical |
| CaCO3 | Sigma | C 4830 | Chemical |
| Cytochrome c | Sigma | C 7752 | Chemical |
| Digitonin | Sigma | D 5628 | Chemical |
| Dithiothreitol | Sigma | D 0632 | Chemical, dissolve in DMSO |
| D-Sucrose | Roth | 4621.1 | Chemical |
| Dulbecco’s modified Eagle’s medium (High glucose) | Fisher Scientific | 41965-039 | Chemical |
| Dulbecco’s modified Eagle’s medium (No Glucose) | Fisher Scientific | A14430-01 | |
| EGTA | Sigma | E 4378 | |
| Etomoxir | Sigma | E1905 | Chemical |
| Falcon 15 ml Conical Centrifuge Tubes | Fisher Scientific | AM12500 | Chemical |
| Falcon 50 ml Conical Centrifuge Tubes | Fisher Scientific | AM12501 | |
| FCCP | Sigma | C 2920 | |
| Glucose | Sigma | G7021 | Chemical, dissolve in ethanol |
| Glutamate | Sigma | G 1626 | Chemical |
| GlutaMax (100x) (200 nM L-alanyl-L-glutamine dipeptide) | Fisher Scientific | 35050061 | Chemical |
| HEK293 cells | ATTC | CRL-1573 | |
| Hemocytometer | Fisher Scientific | 0267151B | Instrument for cell counting |
| Hepes | Sigma | H 7523 | Chemical |
| Imidazole | Fluka | 56750 | Chemical |
| KCl | Merck | 1.04936 | Chemical |
| L-carnitine | Sigma | C0283 | Chemical |
| Malate | Sigma | M 1000 | Chemical |
| MES hydrate | Sigma | M8250 | Chemical |
| MgCl2 | Sigma | M 9272 | Chemical |
| Na2ATP | Sigma | A 2383 | Chemical |
| Na2Phosphocreatine | Sigma | P 7936 | Chemical |
| Na-pyruvate (100 mM) (100x) | Fisher Scientific | 11360070 | |
| NEAA (Non-essential amino acids) 100x | Fisher Scientific | 11140035 | |
| Normal FBS (10x) | Fisher Scientific | 10500064 | |
| O2k-Core: Oxygraph-2k | Oroboros Instruments | 10000-02 | High-resolution respirometry instrument |
| O2k-Titration Set | Oroboros Instruments | 20820-03 | Hamilton syringes for chemical injections |
| Oligomycin | Sigma | O 4876 | Chemical, dissolve in ethanol |
| Palmitoylcarnitine | Sigma | P 4509 | Chemical |
| Penicillin-Streptomycin | Fisher Scientific | 15140122 | |
| Pierce BCA Protein Assay Kit | Fisher Scientific | 23227 | |
| Pyruvate | Sigma | P 2256 | Chemical |
| RIPA-Buffer | Fisher Scientific | 89900 | Chemical |
| Rotenone | Sigma | R 8875 | Chemical, dissolve in ethanol |
| Saponin | Sigma | S7900 | Chemical |
Seahorse XF DMEM assay medium pack, pH 7.4 | Agilent, Santa Clara, CA | 103680-100 | |
| Seahorse XFe96 Extracellular Flux Analyzer | Agilent, Santa Clara, CA | High-throughput respirometry instrument | |
| Seahorse XFe96 FluxPak | Agilent, Santa Clara, CA | Includes assay plates, cartridges, loading guides for transferring compounds to the assay cartridge, and calibrant solution. | |
| Small scissors | Fisher Scientific | 08-951-20 | |
| Sodium azide | Sigma | S2002 | Chemical |
| Succinate | Sigma | S 2378 | Chemical |
| Taurine | Sigma | T 8691 | Chemical |
| TMPD | Sigma | T 3134 | Chemical |
| Trypan Blue solution | Merck | 72-57-1 | Chemical |
| Trypsin 0.25% EDTA | Fisher Scientific | 25200056 | |
| Two thin-edged forceps | Fisher Scientific | 12-000-122 | |
| Uridine stock (500x) | Sigma | U3750 | Chemical |
참고문헌
- McBride, H. M., Neuspiel, M., Wasiak, S. Mitochondria: More than just a powerhouse. Current Biology. 16 (14), 551-560 (2006).
- Mehta, M. M., Weinberg, S. E., Chandel, N. S. Mitochondrial control of immunity. Beyond ATP. Nature Reviews Immunology. 17 (10), 608-620 (2017).
- Spinelli, J. B., Haigis, M. C. The multifaceted contributions of mitochondria to cellular metabolism. Nature Cell Biology. 20 (7), 745-754 (2018).
- Gnaiger, E. Capacity of oxidative phosphorylation in human skeletal muscle. New perspectives of mitochondrial physiology. International Journal of Biochemistry and Cell Biology. 41 (10), 1837-1845 (2009).
- Gorman, G. S., et al. Mitochondrial diseases. Nature Reviews Disease Primers. 2, 1-23 (2016).
- Boushel, R., Gnaiger, E., Schjerling, P., Skovbro, M., Kraunsøe, R., Dela, F. Patients with type 2 diabetes have normal mitochondrial function in skeletal muscle. Diabetologia. 50 (4), 790-796 (2007).
- Cogliati, S., et al. Mitochondrial cristae shape determines respiratory chain supercomplexes assembly and respiratory efficiency. Cell. 155 (1), 160-171 (2013).
- Kühlbrandt, W. Structure and function of mitochondrial membrane protein complexes. BMC Biology. 13, 1-11 (2015).
- Gnaiger, E. Mitochondrial pathways and Respiratory control. An introduction to OXPHOS analysis. Bioenergetics communications. 5th ed. , (2020).
- Jackson, C. B., et al. Mutations in SDHD lead to autosomal recessive encephalomyopathy and isolated mitochondrial complex II deficiency. Journal of Medical Genetics. 51 (3), 170-175 (2014).
- Pesta, D., Gnaiger, E. High-resolution respirometry: OXPHOS protocols for human cells and permeabilized fibers from small biopsies of human muscle. Methods in Molecular Biology. 810, 25-58 (2012).
- Horan, M. P., Pichaud, N., Ballard, J. W. O. Review: Quantifying mitochondrial dysfunction in complex diseases of aging. Journals of Gerontology - Series A Biological Sciences and Medical Sciences. 67 (10), 1022-1035 (2012).
- Doerrier, C., Garcia-Souza, L. F., Krumschnabel, G., Wohlfarter, Y., Mészáros, A. T., Gnaiger, E. High-resolution fluorespirometry and oxphos protocols for human cells, permeabilized fibers from small biopsies of muscle, and isolated mitochondria. Methods in Molecular Biology. 1782, 31-70 (2018).
- Zhang, J., et al. Measuring energy metabolism in cultured cells, including human pluripotent stem cells and differentiated cells. Nature Protocols. 7 (6), 1068-1085 (2012).
- García-Roche, M., Casal, A., Carriquiry, M., Radi, R., Quijano, C., Cassina, A. Respiratory analysis of coupled mitochondria in cryopreserved liver biopsies. Redox Biology. 17, 207-212 (2018).
- Acin-Perez, R., et al. A novel approach to measure mitochondrial respiration in frozen biological samples. The EMBO Journal. 39 (13), 1-18 (2020).
- Cell metabolism assay kits. Seahorse assay kits and media Available from: https://www.agilent.com/en/product/cell-analysis/real-time-cell-metabolic-analysis/xf-assay-lits-reagents-cell-assay-media (2021)
- Chance, B., Williams, G. R. A method for the localization of sites for oxidative phosphorylation. Nature. 176 (4475), 250-254 (1955).
- Gnaiger, E., et al. Mitochondrial respiratory states and rates. MitoFit Preprint Arch. , (2019).
- Gnaiger, E. O2k-procedures: SOP O2k quality control 1: Polarographic oxygen sensors and accuracy of calibration Section Page. Oroboros. 03 (18), 1-21 (2020).
- Robinson, B. H., Petrova-Benedict, R., Buncic, J. R., Wallace, D. C. Nonviability of cells with oxidative defects in galactose medium: A screening test for affected patient fibroblasts. Biochemical Medicine and Metabolic Biology. 48 (2), 122-126 (1992).
- King, M. P., Attardi, G. Human cells lacking mtDNA: Repopulation with exogenous mitochondria by complementation. Science. 246 (4929), 500-503 (1989).
- Makrecka-Kuka, M., Krumschnabel, G., Gnaiger, E. High-resolution respirometry for simultaneous measurement of oxygen and hydrogen peroxide fluxes in permeabilized cells, tissue homogenate and isolated mitochondria. Biomolecules. 5 (3), 1319-1338 (2015).
- Fasching, M., Gnaiger, E. O2k quality control 2: Instrumental oxygen background correction and accuracy of oxygen flux. Mitochondrial Physiology Network. 14 (06), 1-14 (2016).
- Gnaiger, E., Lassnig, B., Kuznetsov, A., Rieger, G., Margreiter, R. Excess capacity of cytochrome c oxidase. Journal of Experimental Biology. 1139, 1129-1139 (1998).
- Gnaiger, E., et al. Mitochondria in the Cold. Life in the Cold. , 431-442 (2000).
- Fontana-Ayoub, M., Fasching, E., Gnaiger, Selected media and chemicals for respirometry with mitochondrial preparations. Mitochondrial Physiology Network. 02 (17), 1-9 (2014).
- Gerencser, A. A., et al. Quantitative microplate-based respirometry with correction for oxygen diffusion. Analytical Chemistry. 81 (16), 6868-6878 (2009).
- Krumschnabel, G., Eigentler, A., Fasching, M., Gnaiger, E. Use of safranin for the assessment of mitochondrial membrane potential by high-resolution respirometry and fluorometry. Methods in Enzymology. 542, 163-181 (2014).
- Nászai, A., Terhes, E., Kaszaki, J., Boros, M., Juhász, L. Ca(2+)N it be measured? Detection of extramitochondrial calcium movement with high-resolution fluorespirometry. Scientific Reports. 9 (1), 1-13 (2019).
- Pajak, B., et al. 2-Deoxy-d-Glucose and its analogs: From diagnostic to therapeutic agents. International Journal of Molecular Sciences. 21 (1), 234 (2019).
- Mercier-Letondal, P., Marton, C., Godet, Y., Galaine, J. Validation of a method evaluating T cell metabolic potential in compliance with ICH Q2 (R1). Journal of Translational Medicine. 19 (1), 1-15 (2021).
- Sauerbeck, A., et al. Analysis of regional brain mitochondrial bioenergetics and susceptibility to mitochondrial inhibition utilizing a microplate based system. Journal of Neuroscience Methods. 198 (1), 36-43 (2011).
- Jackman, M. R., Willis, W. T. Characteristics of mitochondria isolated from type I and type IIb skeletal muscle. American Journal of Physiology - Cell Physiology. 270 (2), 673-678 (1996).
- Ponsot, E., et al. Mitochondrial tissue specificity of substrates utilization in rat cardiac and skeletal muscles. Journal of Cellular Physiology. 203 (3), 479-486 (2005).
- Schönfeld, P., Reiser, G. Why does brain metabolism not favor burning of fatty acids to provide energy-Reflections on disadvantages of the use of free fatty acids as fuel for brain. Journal of Cerebral Blood Flow and Metabolism. 33 (10), 1493-1499 (2013).
- Calderon-Dominguez, M., Mir, J. F., Fucho, R., Weber, M., Serra, D., Herrero, L. Fatty acid metabolism and the basis of brown adipose tissue function. Adipocyte. 5 (2), 98-118 (2016).
- Divakaruni, A. S., Rogers, G. W., Murphy, A. N. Measuring mitochondrial function in permeabilized cells using the seahorse XF analyzer or a clark-type oxygen electrode. Current Protocols in Toxicology. 2014, 1-16 (2014).
- Iuso, A., Repp, B., Biagosch, C., Terrile, C., Prokisch, H. Assessing mitochondrial bioenergetics in isolated mitochondria from various mouse tissues using Seahorse XF96 analyzer. Methods in Molecular Biology. 1567, 217-230 (2017).
- Rogers, G. W., et al. High throughput microplate respiratory measurements using minimal quantities of isolated mitochondria. PLoS ONE. 6 (7), 21746 (2011).
- Jordá, A., Zaragozá, R., Portolés, M., Báguena-Cervellera, R., Renau-Piqueras, J. Long-term high-protein diet induces biochemical and ultrastructural changes in rat liver mitochondria. Archives of Biochemistry and Biophysics. 265 (2), 241-248 (1988).
- Jackson, C. B., Gallati, S., Schaller, A. QPCR-based mitochondrial DNA quantification: Influence of template DNA fragmentation on accuracy. Biochemical and Biophysical Research Communications. 423 (3), 441-447 (2012).
- Hirsch, H. M. Tissue autoxidation inhibitors: II. The presence of inhibitor in intact cells; Assay of liver and hepatoma effect on radio-oxidations. Cancer Research. 16 (11), 1076-1082 (1956).
- Picard, M., et al. Mitochondrial structure and function are disrupted by standard Isolation methods. PLoS ONE. 6 (3), 18317 (2011).
- Tanumihardja, E., Slaats, R. H., Van Der Meer, A. D., Passier, R., Olthuis, W., Van Den Berg, A. Measuring both pH and O2 with a single On-Chip sensor in cultures of human pluripotent stem cell-derived cardiomyocytes to track induced changes in cellular metabolism. ACS Sensors. 6 (1), 267-274 (2021).
- Harms, F., Stolker, R. J., Mik, E. Cutaneous respirometry as novel technique to monitor mitochondrial function: A feasibility study in healthy volunteers. PLoS ONE. 11 (7), 159544 (2016).
- Levitsky, Y., et al. Micro-respirometry of whole cells and isolated mitochondria. RSC Advances. 9 (57), 33257-33267 (2019).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유